Electrophysiological and Alarm Responses of Solenopsis invicta Buren (Hymenoptera: Formicidae) to 2-Ethyl-3,5-dimethylpyrazine


{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
1. Introduction
2. Materials and Methods
2.1. Ant Sources and Colony Maintenance
2.2. Chemical Synthesis
2.3. Antennal Sensitivity Recordings
2.4. Alarm Response
2.5. Enhancement in Bait Attraction
2.6. Statistical Analyses
3. Results
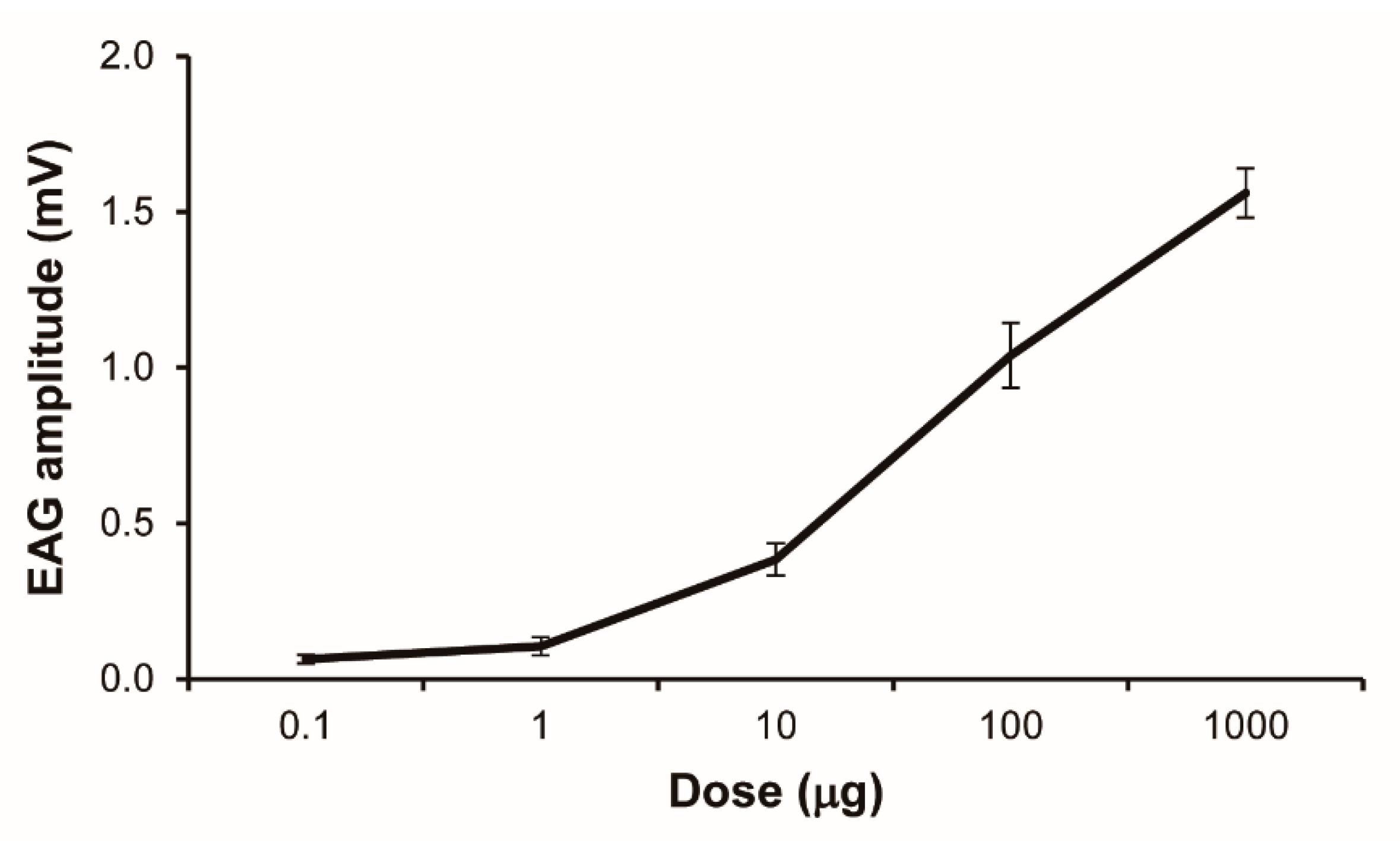
3.1. Antennal Sensitivity
3.2. Alarm Activity
3.3. Enhancement in Bait Attraction
4. Discussion
5. Conclusions
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
References
- Blum, M.S. Alarm pheromone. In Comprehensive Insect Physiology Biochemistry and Pharmacology; Kerkut, G.A., Gilbert, L.I., Eds.; Pergamon Press Inc.: New York, NY, USA, 1985; Volume 9, pp. 194–224. [Google Scholar]
- Wilson, E.O.; Regnier, F.E., Jr. The evolution of the alarm defense system in the formicine ants. Am. Nat. 1971, 105, 279–289. [Google Scholar] [CrossRef]
- Vander Meer, R.K.; Alonso, L.E. Pheromone directed behavior in ants. In Pheromone Communication in Social Insects; Vander Meer, R.K., Breed, M.D., Espelie, K.E., Winston, M.L., Eds.; Westview Press: Boulder, CO, USA, 1998; pp. 159–192. [Google Scholar]
- Verheggen, F.J.; Haubruge, E.; Mescher, M.C. Alarm pheromones—chemical signaling in response to danger. In Vitamins & Hormones; Litwack, G., Ed.; Academic Press: Cambridge, MA, USA, 2010; Volume 83, pp. 215–239. [Google Scholar]
- Hölldobler, B.; Wilson, E.O. The Ants; The Belknap Press and Harvard University Press: Cambridge, MA, USA, 1990. [Google Scholar]
- Brophy, J.J. Pyrazines obtained from insects: Their source, identification, synthesis and function. In Studies in Nature Products Chemistry vol. 5 Structure Elucidation (Part B); Atta-ur-Rahman, Ed.; Elsevier: Amsterdam, The Netherlands, 1989; Volume 5, pp. 221–273. [Google Scholar]
- Attygalle, A.B.; Morgan, E.D. Chemicals from the glands of ants. Chem. Soc. Rev. 1984, 13, 245–278. [Google Scholar] [CrossRef]
- Xu, S.; Errabeli, R.; Feener, D.H.; Noble, K.; Attygalle, A.B. Alkyl-dimethylpyrazines in mandibular gland secretions of four Odontomachus ant species (Formicidae: Ponerinae). J. Chem. Ecol. 2018, 44, 444–451. [Google Scholar] [CrossRef] [PubMed]
- Morgan, E.D.; Nascimento, R.R.d.; Keegans, S.J.; Billen, J. Comparative study of mandibular gland secretions of workers of ponerine ants. J. Chem. Ecol. 1999, 25, 1395–1409. [Google Scholar] [CrossRef]
- Torres-Contreras, H.; Olivares-Donoso, R.; Niemeyer, H.M. Solitary foraging in the ancestral South American ant, Pogonomyrmex vermiculatus. Is it due to constraints in the production or perception of trail pheromones? J. Chem. Ecol. 2007, 33, 435–440. [Google Scholar] [CrossRef]
- Hölldobler, B.; Morgan, E.D.; Oldham, N.J.; Liebig, J. Recruitment pheromone in the harvester ant genus Pogonomyrmex. J. Insect Physiol. 2001, 47, 369–374. [Google Scholar] [CrossRef]
- Keegans, S.J.; Billen, J.; Morgan, E.D.; Gökcen, O.A. Volatile glandular secretions of three species of New World army ants, Eciton burchelli, Labidus coecus, and Labidus praedator. J. Chem. Ecol. 1993, 19, 2705–2719. [Google Scholar] [CrossRef]
- Sainz-Borgo, C.; Cabrera, A.; Hernández, J.V. Nestmate recognition in the ant Odontomachus bauri (Hymenoptera: Formicidae). Sociobiology 2011, 58, 701–718. [Google Scholar]
- Cavill, G.W.K.; Robertson, P.L.; Brophy, J.J.; Duke, R.K.; McDonald, J.; Plant, W.D. Chemical ecology of the meat ant, Iridomyrmex purpureus Sens. Strict. Insect Biochem. 1984, 14, 505–513. [Google Scholar] [CrossRef]
- Vander Meer, R.K.; Preston, C.; Choi, M.-Y. Isolation of a pyrazine alarm pheromone component from the fire ant, Solenopsis invicta. J. Chem. Ecol. 2010, 36, 163–170. [Google Scholar] [CrossRef]
- Wheeler, J.W.; Blum, M.S. Alkylpyrazine alarm pheromones in ponerine ants. Science 1973, 182, 501–503. [Google Scholar] [CrossRef] [PubMed]
- Longhurst, C.; Baker, R.; Howse, P.E.; Speed, W. Alkylpyrazines in ponerine ants: Their presence in three genera, and caste specific behavioural responses to them in Odontomachus troglodytes. J. Insect Physiol. 1978, 24, 833–837. [Google Scholar] [CrossRef]
- Guan, D.; Lu, Y.-Y.; Liao, X.-L.; Wang, L.; Chen, L. Electroantennogram and behavioral responses of the imported fire ant, Solenopsis invicta Buren, to an alarm pheromone component and its analogues. J. Agric. Food Chem. 2014, 62, 11924–11932. [Google Scholar] [CrossRef] [PubMed]
- Liu, X.-F.; Chen, L.; Li, J.-K. Analysis of the alarm pheromone components of the red imported fire ant, Solenopsis invicta Buren (Hymenoptera: Formicidae). Chin. J. Biosaf. 2014, 32, 126–130. [Google Scholar]
- Sharma, K.; Vander Meer, R.K.; Fadamiro, H.Y. Phorid fly, Pseudacteon tricuspis, response to alkylpyrazine analogs of a fire ant, Solenopsis invicta, alarm pheromone. J. Insect Physiol. 2011, 57, 939–944. [Google Scholar] [CrossRef]
- Sharma, K.R.; Fadamiro, H.Y. Fire ant alarm pheromone and venom alkaloids act in concert to attract parasitic phorid flies, Pseudacteon spp. J. Insect Physiol. 2013, 59, 1119–1124. [Google Scholar] [CrossRef]
- Ngumbi, E.; Fadamiro, H. Comparative responses of four Pseudacteon phorid fly species to host fire ant alarm pheromone and analogs. Chemoecology 2015, 25, 85–92. [Google Scholar] [CrossRef]
- Du, Y.; Grodowitz, M.J.; Chen, J. Electrophysiological responses of eighteen species of insects to fire ant alarm pheromone. Insects 2019, 10, 403. [Google Scholar] [CrossRef]
- Sun, Y.; Shao, K.-M.; Lu, Y.-Y.; Shi, Q.-H.; Wang, W.-K.; Chen, L. Electrophysiological and alarm behavioral responses of Solenopsis invicta Buren (Hymenoptera: Formicidae) to alkoxypyrazines. J. Asia-Pacif. Entomol. 2017, 20, 541–546. [Google Scholar] [CrossRef]
- Fang, M.; Cadwallader, K.R. Convenient synthesis of stable deuterium-labeled alkylpyrazines for use in stable isotope dilution assays. J. Agric. Food Chem. 2013, 61, 3580–3588. [Google Scholar] [CrossRef]
- Li, Y.-Y.; Lu, Y.-Y.; Lu, M.; Wei, H.-Y.; Chen, L. HPLC separation of 2-ethyl-5(6)-methylpyrazine and its electroantennogram and alarm activities on fire ants (Solenopsis invicta Buren). Molecules 2018, 23, 1661. [Google Scholar] [CrossRef] [PubMed]
- Chen, L.; Li, Y.-Y.; Shao, K.-M. A practical technique for electrophysiologically recording from lamellated antenna of scarab beetle. J. Chem. Ecol. 2019, 45, 392–401. [Google Scholar] [CrossRef] [PubMed]
- SAS Institute. SAS User Guide; SAS Institute: Cary, NC, USA, 2004. [Google Scholar]
- Morgan, E.D.; Inwood, M.R.; Cammaerts, M.-C. The mandibular gland secretion of the ant, Myrmica scabrinodis. Physiol. Entomol. 1978, 3, 107–114. [Google Scholar] [CrossRef]
- Andryszak, N.A.; Payne, T.L.; Dickens, J.C.; Moser, J.C.; Fisher, R.W. Antennal olfactory responsiveness of the Texas leaf cutting ant (Hymenoptera: Formicidae) to trail pheromone and its two alarm substances. J. Entomol. Sci. 1990, 25, 593–600. [Google Scholar] [CrossRef]
- Fujiwara-Tsujii, N.; Yamagata, N.; Takeda, T.; Mizunami, M.; Yamaoka, R. Behavioral responses to the alarm pheromone of the ant Camponotus obscuripes (Hymenoptera: Formicidae). Zool. Sci. 2006, 23, 353–358. [Google Scholar] [CrossRef]
- Derstine, N.T.; Troyer, E.J.; Suttles, C.N.; Siderhurst, L.A.; Jang, E.B.; Siderhurst, M.S. Field trapping the little fire ant, Wasmannia auropunctata. J. Insect Sci. 2012, 12, 93. [Google Scholar] [CrossRef][Green Version]
- Welzel, K.F.; Choe, D.H. Development of a pheromone-assisted baiting technique for Argentine ants (Hymenoptera: Formicidae). J. Econ. Entomol. 2016, 109, 1303–1309. [Google Scholar] [CrossRef]
- Greenberg, L.; Klotz, J.H. Argentine ant (Hymenoptera: Formicidae) trail pheromone enhances consumption of liquid sucrose solution. J. Econ. Entomol. 2000, 93, 119–122. [Google Scholar] [CrossRef]
- Hughes, W.O.H.; Howse, P.E.; Vilela, E.F.; Knapp, J.J.; Goulson, D. Field evaluation of potential of alarm pheromone compounds to enhance baits for control of grass-cutting ants (Hymenoptera: Formicidae). J. Econ. Entomol. 2002, 95, 537–543. [Google Scholar] [CrossRef]
- Hughes, W.O.H.; Goulson, D. The use of alarm pheromones to enhance bait harvest by grass-cutting ants. Bull. Entomol. Res. 2002, 92, 213–218. [Google Scholar] [CrossRef]
- Joo, K.; Ho, C.-T. Quantitative analysis of alkylpyrazines in regular- and low-fat commercial peanut butter preparations. Biosci. Biotechnol. Biochem. 1997, 61, 171–173. [Google Scholar] [CrossRef] [PubMed]




© 2019 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Li, Y.-Y.; Liu, D.; Chen, L. Electrophysiological and Alarm Responses of Solenopsis invicta Buren (Hymenoptera: Formicidae) to 2-Ethyl-3,5-dimethylpyrazine. Insects 2019, 10, 451. https://doi.org/10.3390/insects10120451
Li Y-Y, Liu D, Chen L. Electrophysiological and Alarm Responses of Solenopsis invicta Buren (Hymenoptera: Formicidae) to 2-Ethyl-3,5-dimethylpyrazine. Insects. 2019; 10(12):451. https://doi.org/10.3390/insects10120451
Chicago/Turabian StyleLi, Ya-Ya, Deguang Liu, and Li Chen. 2019. "Electrophysiological and Alarm Responses of Solenopsis invicta Buren (Hymenoptera: Formicidae) to 2-Ethyl-3,5-dimethylpyrazine" Insects 10, no. 12: 451. https://doi.org/10.3390/insects10120451
APA StyleLi, Y.-Y., Liu, D., & Chen, L. (2019). Electrophysiological and Alarm Responses of Solenopsis invicta Buren (Hymenoptera: Formicidae) to 2-Ethyl-3,5-dimethylpyrazine. Insects, 10(12), 451. https://doi.org/10.3390/insects10120451