Cryptic Species Discrimination in Western Pine Beetle, Dendroctonus brevicomis LeConte (Curculionidae: Scolytinae), Based on Morphological Characters and Geometric Morphometrics

 ,
,
Abstract

{kind=link}
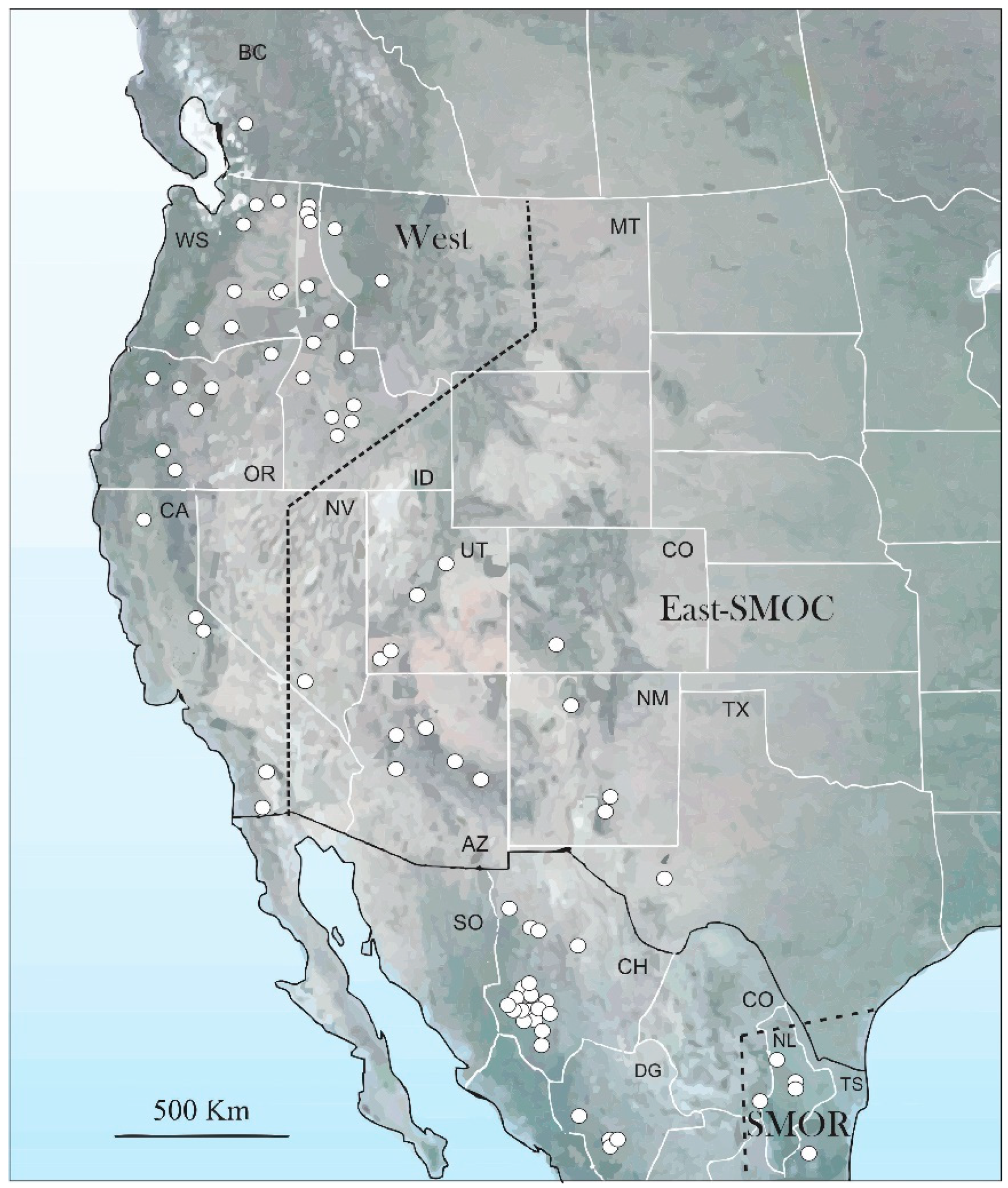{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
1. Introduction
2. Materials and Methods
2.1. Samples
2.2. Characters
2.3. Morphometrics Analysis
2.4. Geometric Morphometrics
3. Results
3.1. Multivariate Analysis of Non-Geometric Morphological Data
3.2. Relative Taxonomic Weight of Individual Characters
3.3. Geometric Morphometrics
4. Discussion
4.1. Geometric Morphometrics
4.2. Taxonomic Considerations
4.3. Redescriptions
4.3.1. Dendroctonus brevicomis LeConte 1876
4.3.2. Description of Female
4.3.3. Description of Male.
4.3.4. Dendroctonus Barberi Hopkins 1909
4.3.5. Description of the Female
4.3.6. Description of the Male
5. Final Considerations
6. Conclusions
Supplementary Materials
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
References
- DeMars, C.J.; Roettgering, B.H. Western pine beetle. In Forest Service and Disease Leaflet, I; USDA Forest Service: Washington, DC, USA, 1982; p. 8. [Google Scholar]
- Valerio-Mendoza, O.; Armendáriz-Toledano, F.; Cuellar-Rodríguez, G.; Negrón, J.F.; Zúñiga, G. The current status of the distribution range of the western pine beetle, Dendroctonus brevicomis (Curculionidae: Scolytinae) in northern Mexico. J. Insect Sci. 2017, 17, 1–7. [Google Scholar] [CrossRef] [PubMed][Green Version]
- Miller, J.C.; Keen, F.P. Biology and Control of the Western pine beetle: A Summary of the First Fifty Years of Research; Misc. Publication 800; United States Department of Agriculture: Washington, DC, USA, 1960; p. 381.
- Six, D.L.; Bracewell, R.R. Dendroctonus. In Bark Beetles: Biology and Ecology of Native and Invasive Species; Vega, F.E., Hofstetter, R.W., Eds.; Academic Press: Cambridge, MA, USA; Elsevier: London, UK, 2015; p. 350. [Google Scholar]
- Salinas-Moreno, Y.; Mendoza, M.G.; Barrios, M.A.; Cisneros, R.; Macías-Sámano, J.; Zúñiga, G. Aerography of the genus Dendroctonus (Coleoptera: Curculionidae: Scolytinae) in Mexico. J. Biogeogr. 2004, 31, 1163–1177. [Google Scholar] [CrossRef]
- Lanier, G.N.; Hendrichs, J.P.; Flores, J.E. Biosystematics of the Dendroctonus frontalis (Coleoptera: Scolytidae) Complex. Ann. Entomol. Soc. Am. 1988, 81, 403–418. [Google Scholar] [CrossRef]
- Armendáriz-Toledano, F.; Niño-Domínguez, A.; Sullivan, B.T.; Kirkendall, L.R.; Zúñiga, G. A new species of bark beetle, Dendroctonus mesoamericanus sp. nov. (Curculionidae: Scolytinae) in southern Mexico and Central America. Ann. Entomol. Soc. Am. 2015, 108, 403–414. [Google Scholar] [CrossRef]
- Zúñiga, G.; Cisneros, R.; Salinas-Moreno, Y. Coexistencia de Dendroctonus frontalis Zimmerman y D. mexicanus Hopkins (Coleoptera: Scolytidae) sobre un mismo hospedero. Acta Zool. Mex. (Nueva Serie) 1995, 64, 59–62. [Google Scholar] [CrossRef]
- Zúñiga, G.; Mendoza-Correa, G.; Cisneros, R.; Salinas-Moreno, Y. Zonas de sobreposición geográfica de las especies mexicanas de Dendroctonus Erichson (Coleoptera: Scolytidae) y sus implicaciones ecológico-evolutivas. Acta Zool. Mex. (Nueva Serie) 1999, 77, 1–22. [Google Scholar] [CrossRef]
- Moser, J.C.; Fitzgibbon, B.A.; Klepzig, K.D. The Mexican pine beetle, Dendroctonus mexicanus: First record in the United States and co-occurrence with the southern pine beetle Dendroctonus frontalis (Coleoptera: Scolytidae or Curculionidae: Scolytinae). Entomol. News 2005, 116, 235–243. [Google Scholar]
- LeConte, J.L. The Rhynchophora of America north of Mexico. Proc. Am. Philos. Soc. 1876, 15, 1–452. [Google Scholar]
- Hopkins, A.D. Contributions toward a Monograph of the Scolytid Beetles. I. The Genus Dendroctonus; United States Department of Agriculture, Series No. 17, Part 1; Forgotten Books: London, UK, 1909; pp. 1–164. [Google Scholar]
- Swaine, J.M. Canadian bark beetles, Part II. A Preliminary classification with an account of the habits and means of controls. Can. Dep. Agric. 1918, 14, 250. [Google Scholar]
- Wood, S.L. A revision of the bark beetles genus Dendroctonus Erichson (Coleoptera: Scolytidae). Great Basin Nat. 1963, 23, 1–116. [Google Scholar]
- Wood, S.L. The bark and ambrosia beetles of North and Central America (Coleoptera: Scolytidae). A taxonomic monograph. Great Basin Nat. 1982, 6, 1–1359. [Google Scholar]
- Kelley, S.; Mitton, J.B.; Paine, T.D. Strong differentiation in mitochondrial DNA of Dendroctous brevicomis (Coleoptera: Scolytidae) on different subspecies of Ponderosa pine. Ann. Entomol. Soc. Am. 1999, 92, 193–197. [Google Scholar] [CrossRef]
- Pureswaran, D.S.; Hofstetter, R.W.; Sullivan, B.T.; Grady, A.M.; Brownie, C. Western pine beetle populations in Arizona and California differ in the composition of their aggregation pheromones. J. Chem. Ecol. 2016, 42, 404–413. [Google Scholar] [CrossRef] [PubMed]
- Bracewell, R.R.; Vanderpool, D.; Good, J.M.; Six, D.L. Cascading speciation among mutualists and antagonists in a tree–beetle–fungi interaction. Proc. R. Soc. B. 2018, 285, 20180694. [Google Scholar] [CrossRef]
- Sullivan, B.T. (USDA Forest Service: Pineville, LA, USA). Personal Communication, 2019.
- Armendáriz-Toledano, F.; Zúñiga, G. Illustrated key to species of genus Dendroctonus (Coleoptera: Curculionidae) occurring in Mexico and Central America. J. Insect Sci. 2017, 34, 1–15. [Google Scholar] [CrossRef]
- Lyon, R.L. A useful secondary sex character in Dendroctonus bark beetles. Can. Entomol. 1958, 90, 582–584. [Google Scholar] [CrossRef]
- Wood, S.L. New synonymy and records of American bark beetles (Coleoptera: Scolytidae). Great Basin Nat. 1974, 34, 277–290. [Google Scholar]
- López, M.F.; Armendáriz-Toledano, F.; Macías-Sámano, J.E.; Shibayama-Salas, M.; Zúñiga, G. Comparative Study of the antennae of Dendroctonus rhizophagus and Dendroctonus valens (Curculionidae: Scolytinae): Sensilla types, distribution and club shape. Ann. Entomol. Soc. Am. 2014, 107, 1130–1143. [Google Scholar] [CrossRef]
- Gower, J.C. A general coefficient of similarity and some of its properties. Biometrics 1971, 27, 857–874. [Google Scholar] [CrossRef]
- Zar, J.H. Bioestatistical Analysis; Prentice Hall: Upper Saddle River, NJ, USA, 2010; p. 944. [Google Scholar]
- Bookstein, F.L. Morphometrics Tools for Landmark Data: Geometry and Biology; Cambridge University Press: Cambridge, UK, 1997; p. 512. [Google Scholar]
- Zelditch, L.M.; Swiderski, D.L.; Sheets, H.D.; Fink, W.L. Geometric Morphometrics for Biologist: A Primer; Maple Vail: New York, NY, USA, 2004; p. 416. [Google Scholar]
- Sheets, H.D. IMP–Integrated Morphometrics Package; Department of Physics, Canisius College: Buffalo, NY, USA, 2003; Available online: www.filogenetica.org/cursos/Morfometria/IMP_installers/index.php (accessed on 9 October 2019).
- Rohlf, F.J. TpsDig. Version 1.40. Ecology and Evolution; State University of New York: Suny at Stony Brook, NY, USA, 2004; Available online: https://life.bio.sunysb.edu/morph/ (accessed on 10 January 2018).
- Hammer, O.; Harper, D.; Ryan, P. Paleontological statistics software package for education and data analysis. Paleontol. Electrón. 2001, 4, 1–9. [Google Scholar]
- Lanier, G.N.; Wood, D.L. Controlled mating, karyology, morphology, and sex-ratio in the Dendroctonus ponderosa complex. Ann. Entomol. Soc. Am. 1968, 61, 517–526. [Google Scholar] [CrossRef]
- Pajares, J.A.; Lanier, G.N. Biosystematics of the turpentine beetles Dendroctonus terebrans and D. valens (Coleoptera: Scolytidae). Ann. Entomol. Soc. Am. 1990, 83, 171–188. [Google Scholar] [CrossRef]
- García-Román, J.; Armendáriz-Toledano, F.; Valerio-Mendoza, O.; Zúñiga, G. An assessment of old and new characters using traditional and geometric morphometrics for the identification of Dendroctonus approximatus and D. parallelocollis (Curculionidae: Scolytinae). J. Insect Sci. 2019, 19, 1–13. [Google Scholar] [CrossRef] [PubMed]
- Armendáriz-Toledano, F.; Niño-Domínguez, A.; Sullivan, B.T.; Macías-Sámano, J.; Victor, J.; Clarke, S.; Zúñiga, G. Two species within Dendroctonus frontalis (Coleoptera: Curculionidae): Evidence from morphological, karyological, molecular, and crossing studies. Ann. Entomol. Soc. Am. 2014, 107, 11–27. [Google Scholar] [CrossRef]
- López, M.F.; Armendáriz-Toledano, F.; Albores-Medina, A.; Zúñiga, G. Morphology of antennae of Dendroctonus vitei (Coleoptera: Curculionidae: Scolytinae), with special reference to sensilla clustered into pit craters. Can. Entomol. 2018, 150, 471–480. [Google Scholar] [CrossRef]
- Vité, J.P.; Islas, S.F.; Renwich, J.A.A.; Hughes, P.R.; Kliefoth, R.A. Biochemical and biological variation of southern pine beetle population in North and Central America. J. Appl. Entomol. 1974, 75, 422–435. [Google Scholar] [CrossRef]
- Vité, J.P.; Hughes, R.F.; Renwich, J.A.A. Pine beetles of the genus Dendroctonus: Pest population in Central America. FAO Plant Prot. Bull. 1975, 6, 178–184. [Google Scholar]
- Ríos-Reyes, A.V.; Valdez-Carrasco, J.; Equihua-Martínez, A.; Moya-Raygoza, G. Identification of Dendroctonus frontalis (Zimmermann) and D. mexicanus (Hopkins) (Coleoptera: Curculionidae: Scolytinae) through structures of the female genitalia. Coleopt Bull. 2008, 62, 99–103. [Google Scholar] [CrossRef]
- Armendáriz-Toledano, F.; García-Román, J.; López, M.F.; Sullivan, B.T.; Zúñiga, G. New characters and redescription of Dendroctonus vitei (Coleoptera: Curculionidae: Scolytinae). Can. Entomol. 2017, 149, 413–433. [Google Scholar] [CrossRef]
- Furniss, M.M. A new subspecies of Dendroctonus (Coleoptera: Scolytidae) from Mexico. Ann. Entomol. Soc. Am. 2001, 94, 21–25. [Google Scholar] [CrossRef]
- Sullivan, B.T.; Niño-Domínguez, A.; Moreno, B.; Brownie, C.; Macías-Sámano, J.; Clarke, S.R.; Kirkendall, L.R.; Zúñiga, G. Biochemical evidence that Dendroctonus frontalis consists of two sibling species in Belize and Chiapas, Mexico. Ann. Entomol. Soc. Am. 2012, 105, 817–831. [Google Scholar] [CrossRef]
- Bright, D.E. A Taxonomic Monograph of the Bark and Ambrosia Beetles of the West Indies (Coleoptera: Curculionoidea: Scolytidae). Studies on West Indian Scolytidae (Coleoptera) 7. Occas. Papers Fla. State Collection Arthropods 2019, 12, 1–491. [Google Scholar]
- Cognato, A.I.; Gillette, N.E.; Campos-Bolaños, R.; Sperling, F.A.H. Mitochondrial phylogeny of pine cone beetles (Scolytinae, Conophthorus) and their affiliation with geographic area and host. Mol. Phylogenet. Evol. 2005, 36, 494–508. [Google Scholar] [CrossRef] [PubMed]
- Pernek, M.; Navtzis, D.; Hrašovec, B.; Diminic, D.; Wegensteiner, R.; Stauffer, C.; Cognato, A.I. Novel morphological and genetic markers for the discrimination of three European Pityokteines (Coleoptera: Curculionidae: Scolytinae) species. Periodicum Biol. 2008, 110, 329–334. [Google Scholar]
- Ruíz, E.A.; Rinehart, J.E.; Hayes, J.L.; Zúñiga, G. Effect of geographic isolation on /genetic differentiation in the Douglas-fir beetle, Dendroctonus pseudotsugae Hopkins (Coleoptera: Curculionidae: Scolytinae). Hereditas 2009, 146, 79–92. [Google Scholar] [CrossRef]
- Ruíz, E.A.; Victor, J.; Hayes, J.L.; Zúñiga, G. Molecular and morphological analysis of Dendroctonus pseudotsugae Hopkins (Coleoptera: Curculionidae: Scolytinae): An assessment of the taxonomic status of subspecies. Ann. Entomol. Soc. Am. 2009, 102, 982–997. [Google Scholar] [CrossRef]
- Cerezke, H.F. The morphology and function of the reproductive systems of Dendroctonus monticolae Hopk. (Coleoptera: Scolytidae). Can. Entomol. 1964, 96, 477–500. [Google Scholar] [CrossRef]
- Willyard, A.; Gernandt, D.S.; Potter, K.; Hipkins, V.; Marquardt, P.; Mahalovich, M.F.; Langer, S.K.; Telewski, F.W.; Cooper, B.; Douglas, C.; et al. Pinus ponderosa: A checkered past obscured four species. Am. J. Bot. 2017, 104, 161–181. [Google Scholar] [CrossRef]










© 2019 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Valerio-Mendoza, O.; García-Román, J.; Becerril, M.; Armendáriz-Toledano, F.; Cuéllar-Rodríguez, G.; Negrón, J.F.; Sullivan, B.T.; Zúñiga, G. Cryptic Species Discrimination in Western Pine Beetle, Dendroctonus brevicomis LeConte (Curculionidae: Scolytinae), Based on Morphological Characters and Geometric Morphometrics. Insects 2019, 10, 377. https://doi.org/10.3390/insects10110377
Valerio-Mendoza O, García-Román J, Becerril M, Armendáriz-Toledano F, Cuéllar-Rodríguez G, Negrón JF, Sullivan BT, Zúñiga G. Cryptic Species Discrimination in Western Pine Beetle, Dendroctonus brevicomis LeConte (Curculionidae: Scolytinae), Based on Morphological Characters and Geometric Morphometrics. Insects. 2019; 10(11):377. https://doi.org/10.3390/insects10110377
Chicago/Turabian StyleValerio-Mendoza, Osiris, Jazmín García-Román, Moises Becerril, Francisco Armendáriz-Toledano, Gerardo Cuéllar-Rodríguez, José F. Negrón, Brian T. Sullivan, and Gerardo Zúñiga. 2019. "Cryptic Species Discrimination in Western Pine Beetle, Dendroctonus brevicomis LeConte (Curculionidae: Scolytinae), Based on Morphological Characters and Geometric Morphometrics" Insects 10, no. 11: 377. https://doi.org/10.3390/insects10110377
APA StyleValerio-Mendoza, O., García-Román, J., Becerril, M., Armendáriz-Toledano, F., Cuéllar-Rodríguez, G., Negrón, J. F., Sullivan, B. T., & Zúñiga, G. (2019). Cryptic Species Discrimination in Western Pine Beetle, Dendroctonus brevicomis LeConte (Curculionidae: Scolytinae), Based on Morphological Characters and Geometric Morphometrics. Insects, 10(11), 377. https://doi.org/10.3390/insects10110377