Insecticidal Activity of Compounds of Plant Origin on Mahanarva spectabilis (Hemiptera: Cercopidae)

 ,
, {kind=link}
{kind=link}
{kind=link}
Abstract
:1. Introduction
2. Materials and Methods
2.1. Acquisition and Maintenance of Insects and Plants
2.2. Reagents
2.3. Evaluation of Insecticidal Activity
2.4. Statistical Analysis
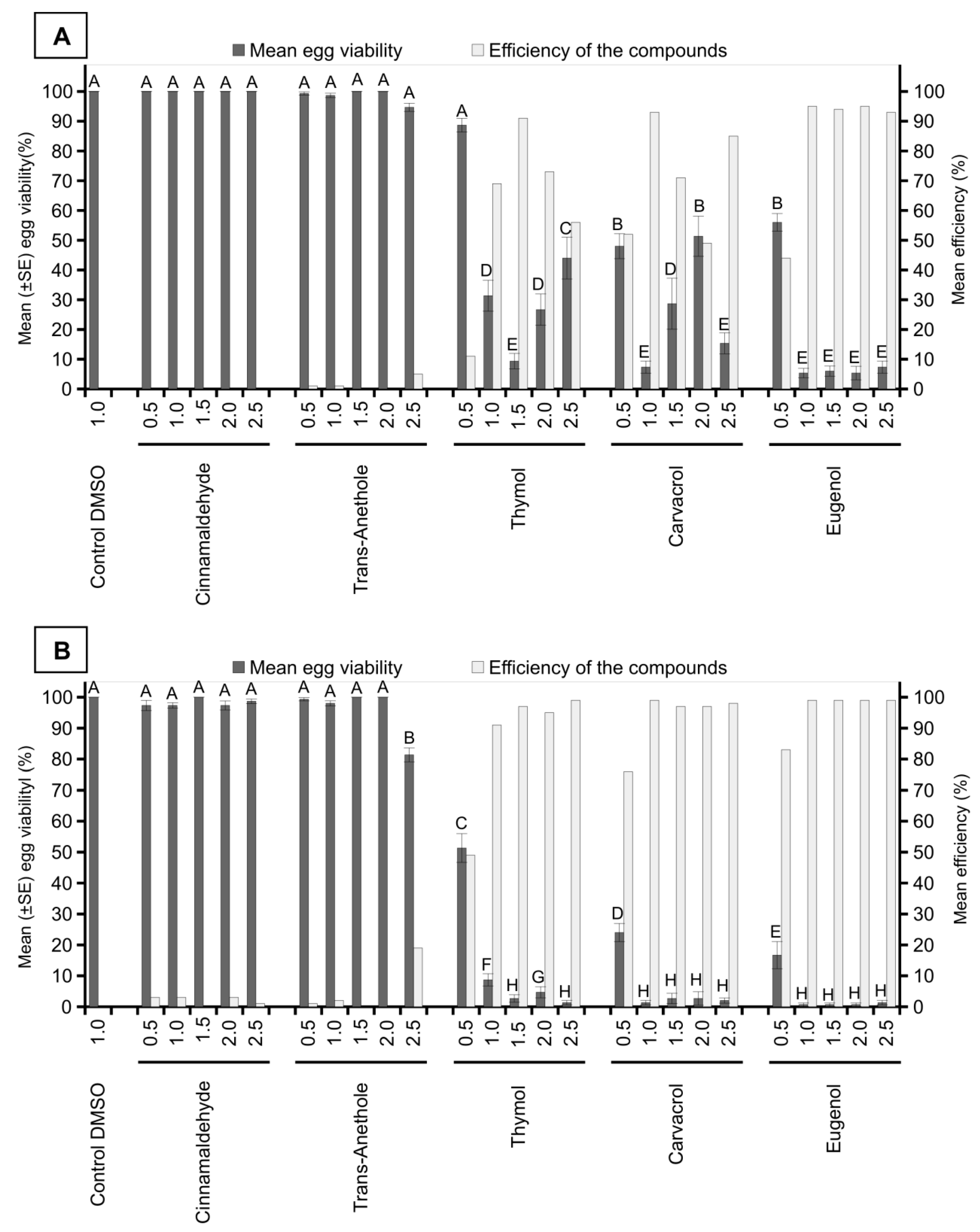
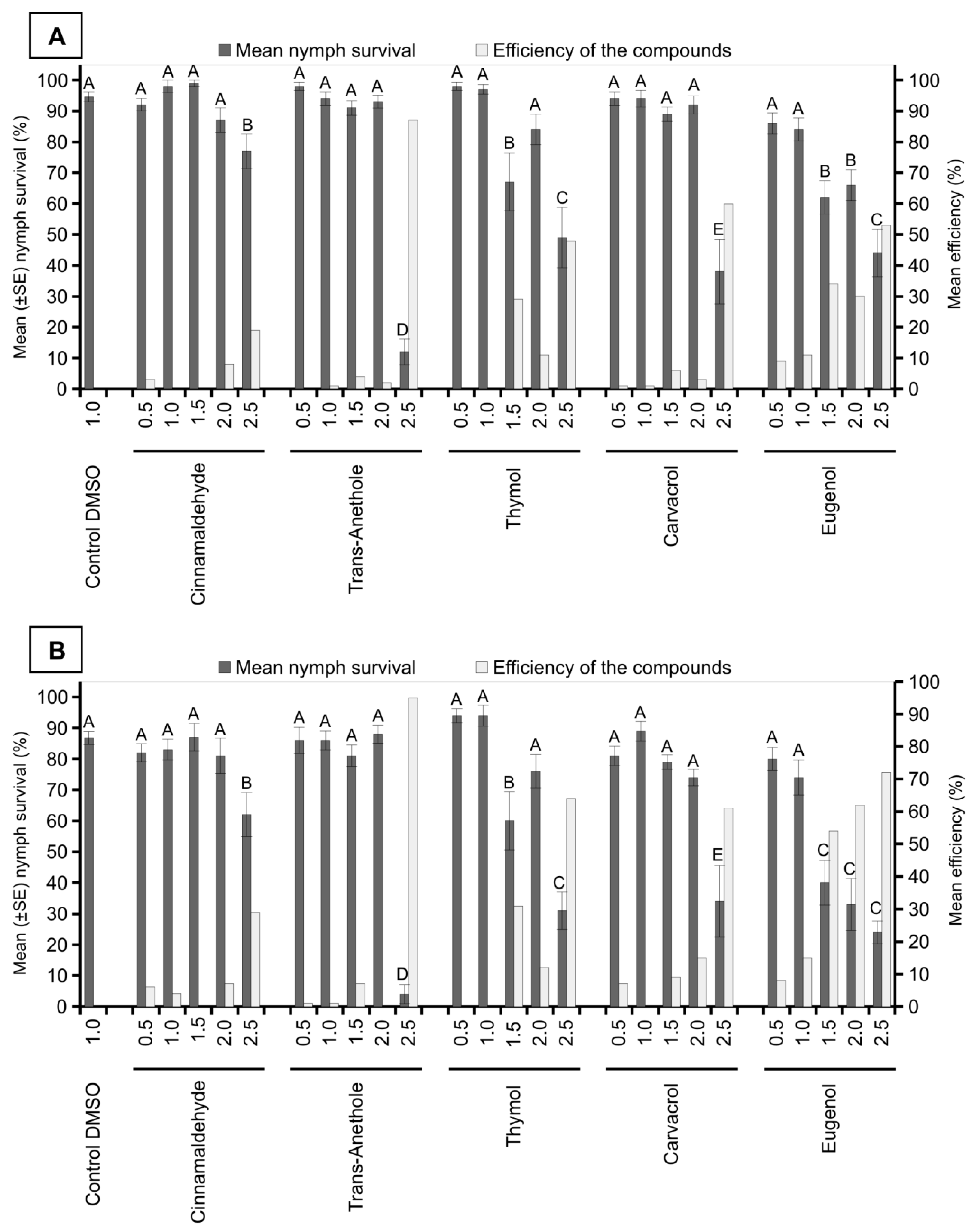
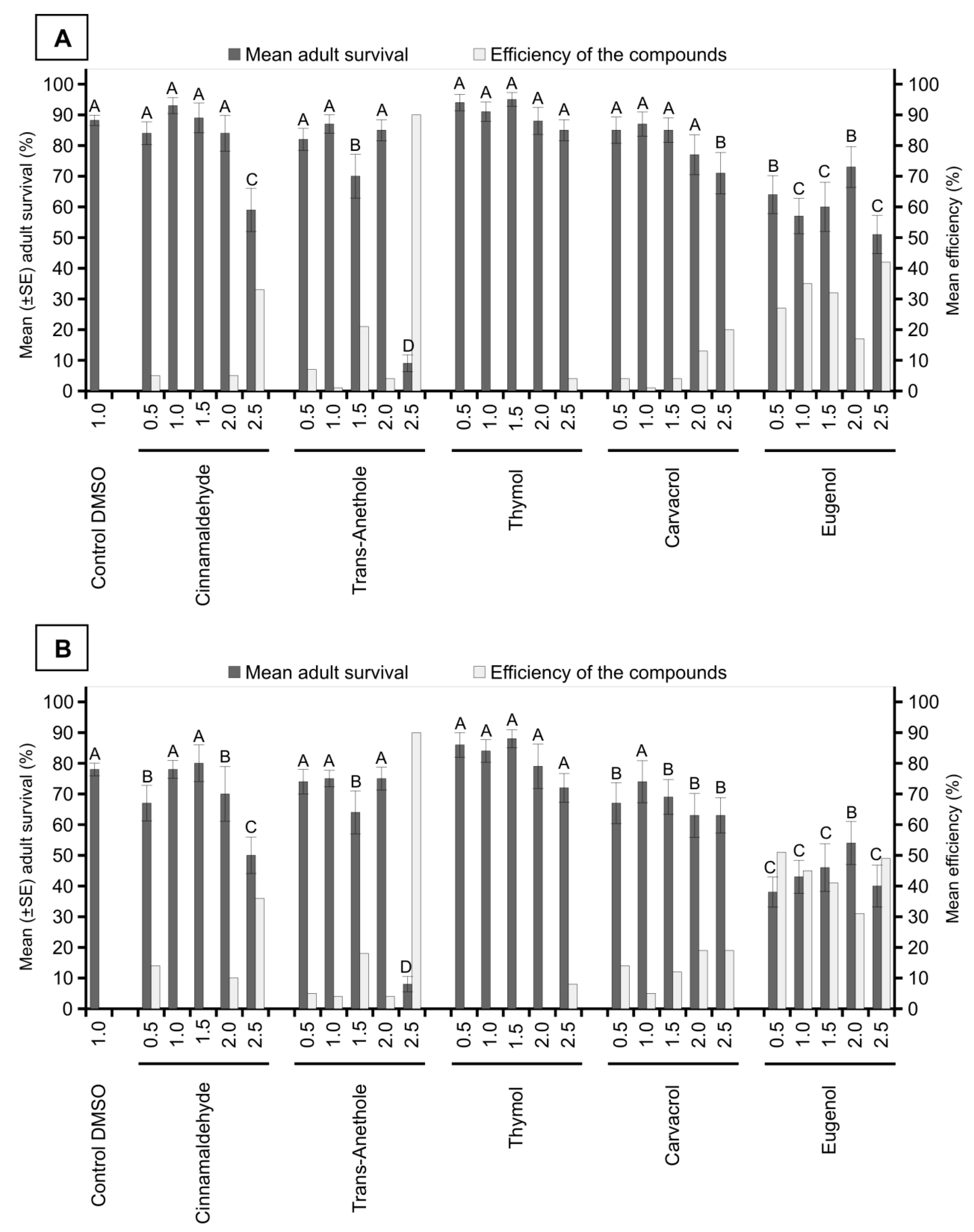
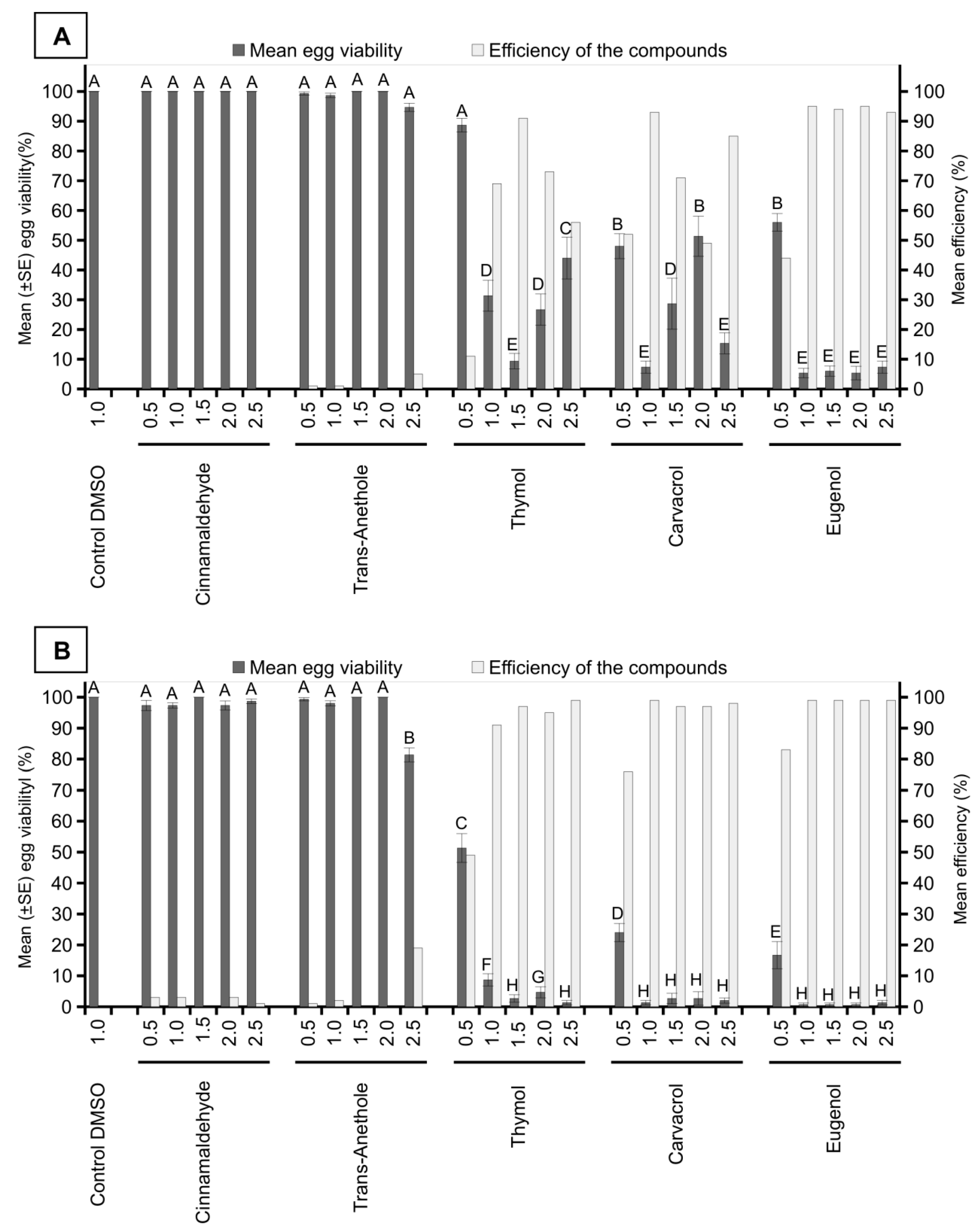
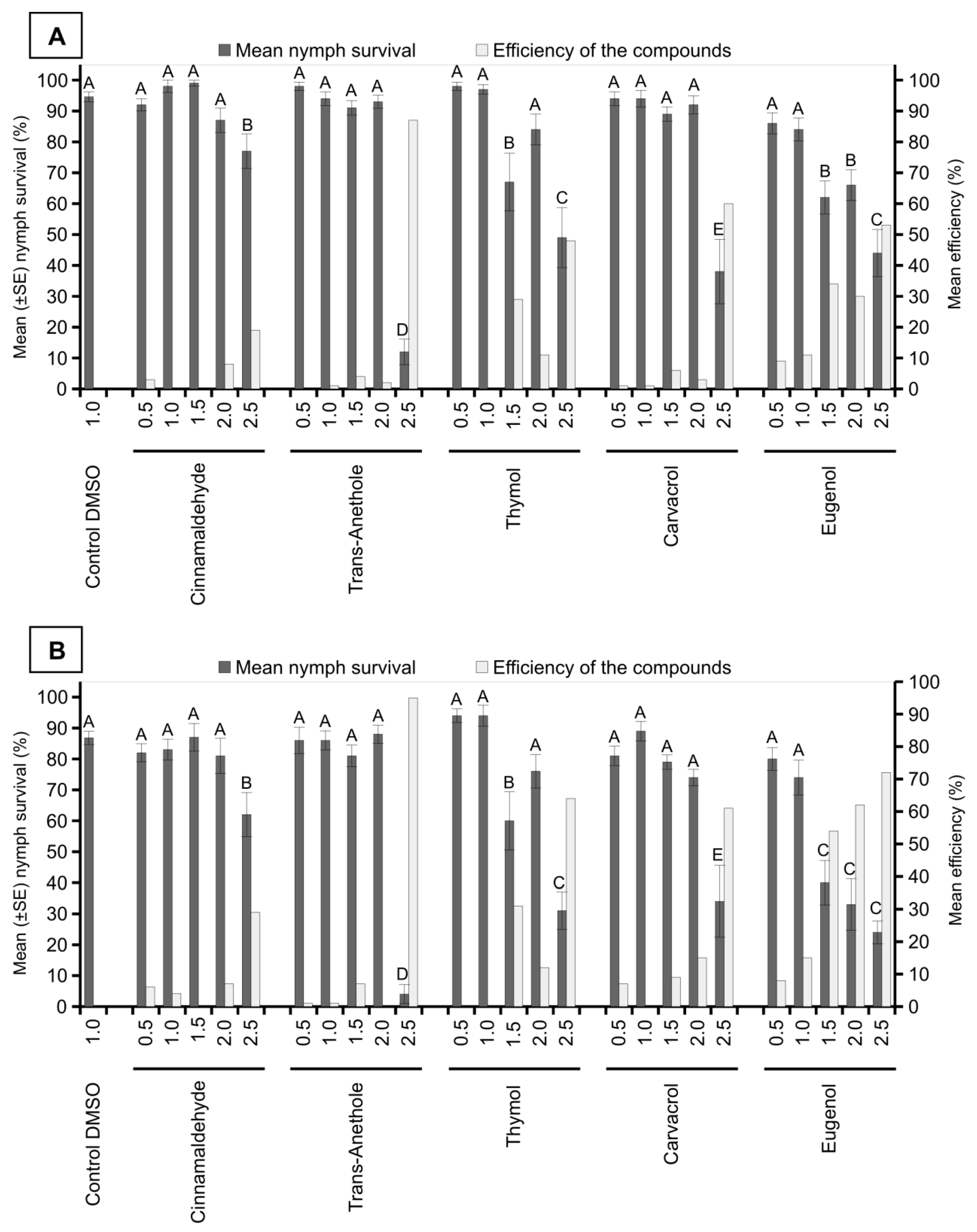
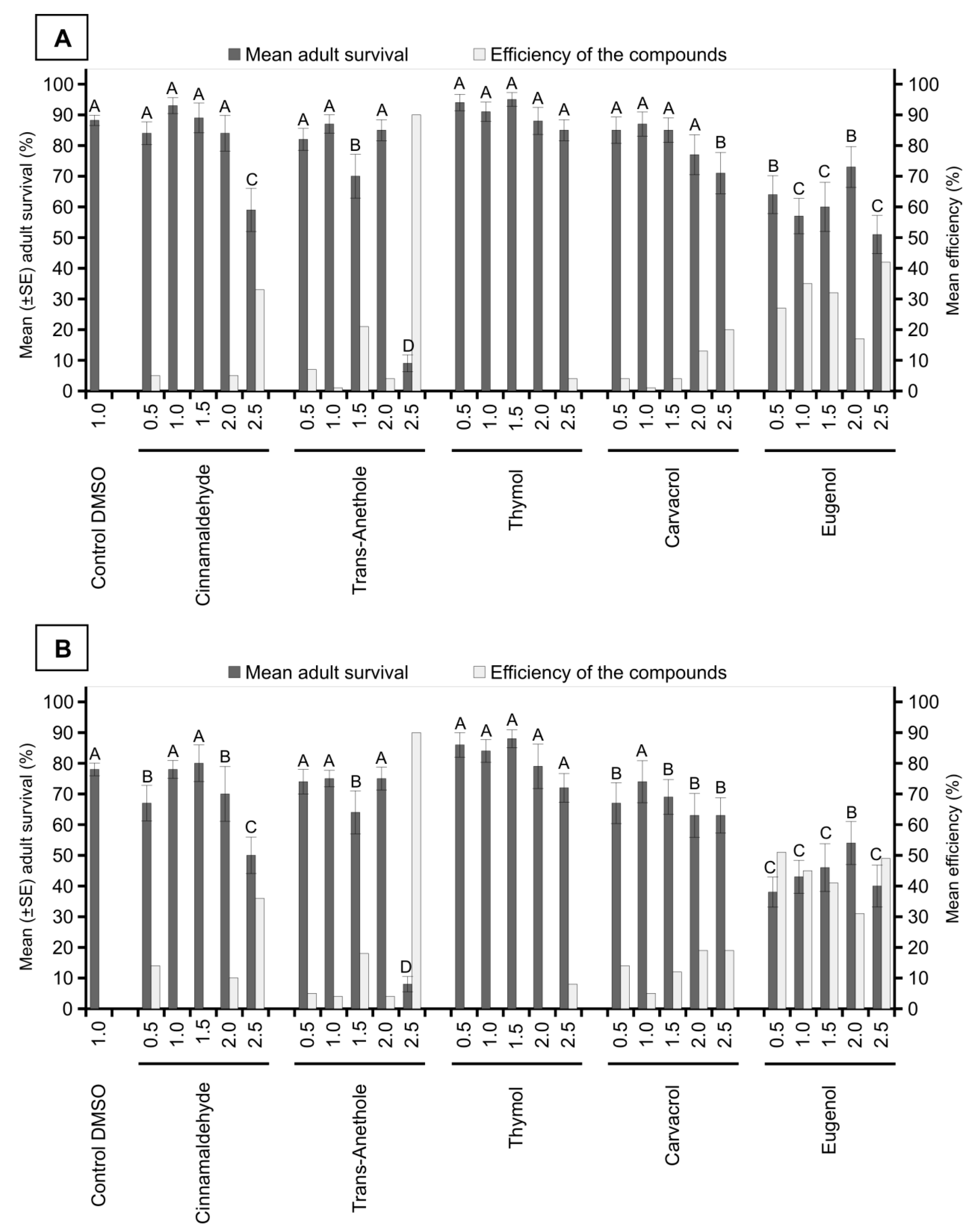
3. Results
4. Discussion
5. Conclusions
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
References
- Valério, J.R.; Nakano, O. Danos causados pelo adulto da cigarrinha Zulia entreriana na produção e qualidade de Brachiaria decumbens. Pesqui. Agropecu. Bras. 1988, 23, 447–453. [Google Scholar]
- Bernardo, E.R.D.A.; Rocha, V.D.F.; Puga, O.; Silva, R.A.D. Espécies de cigarrinhas-das-pastagens (Hemiptera: Cercopidae) no meio-norte do Mato Grosso. Cienc Rural 2003, 33, 369–371. [Google Scholar] [CrossRef]
- Thompson, V. Associative nitrogen fixation, C 4 photosynthesis, and the evolution of spittlebugs (Hemiptera: Cercopidae) as major pests of neotropical sugarcane and forage grasses. Bull. Entomol. Res. 2004, 94, 189–200. [Google Scholar] [CrossRef] [PubMed]
- Auad, A.M.; Simões, A.D.; Pereira, A.V.; Braga, A.L.F.; Sobrinho, F.S.; Lêdo, F.J.S.; Paula-Moraes, S.V.; Oliveira, S.A.; Ferreira, R.B. Seleção de genótipos de capim-elefante quanto à resistência à cigarrinha-das-pastagens. Pesqui. Agropecu. Bras. 2007, 42, 1077–1081. [Google Scholar] [CrossRef]
- Hewitt, G.B. Grazing management as a means of regulating spittlebug (Homoptera, Cercopidae) numbers in central Brazil. Pesqui. Agropecu. Bras. 1988, 23, 697–707. [Google Scholar]
- Oliveira, M.A.S.; Alves, P.M.P. Novas opções de gramíneas no controle da cigarrinha-das-pastagens em Rondônia; Boletim de pesquisa EMBRAPA-UEPAE: Porto Velho, Brazil, 1988; Volume 9, pp. 1–18. [Google Scholar]
- Souza, J.C.; Silva, R.A.; Reis, P.R.; Queiroz, D.S.; Silva, D.B. Cigarrinhas-das-Pastagens: Histórico, Bioecologia, Prejuízos, Monitoramento e Medidas de Controle; Circular técnica EPAMIG: Belo Horizonte, Brazil, 2008; Volume 42, pp. 1–8. [Google Scholar]
- Valério, J.R. Cigarrinha das Pastagens; Circular Técnica, Embrapa Gado de Corte: Campo Grande, Brazil, 2009; Volume 179, pp. 1–50. [Google Scholar]
- Auad, A.M.; Resende, T.T. Use of chemical inducers as a resistance trigger in Brachiaria grasses and sugarcane. Fla Entomol. 2018, 101, 119–124. [Google Scholar] [CrossRef]
- Ferrufino, A.; Lapointe, S.L. Host plant resistance in Brachiaria grasses to the spittlebug Zulia colombiana. Entomol. Exp. Appl. 1989, 51, 155–162. [Google Scholar] [CrossRef]
- Valério, J.R. Insetos-praga em pastagens tropicais. Inf. Agropecu. 2005, 26, 98–110. [Google Scholar]
- Sarwar, M. The killer chemicals as controller of agriculture insect pests: The conventional insecticides. Int. J. Chem. Biomol. Sci. 2015, 1, 141–147. [Google Scholar]
- Isman, M.B. Botanical insecticides, deterrents, and repellents in modern agriculture and an increasingly regulated world. Annu. Rev. Entomol. 2006, 51, 45–66. [Google Scholar] [CrossRef]
- Pavela, R. Insecticidal properties of Pimpinella anisum essential oils against the Culex quinquefasciatus and the non-target organism Daphnia magna. J. Asia-Pac. Entomol. 2014, 17, 287–293. [Google Scholar] [CrossRef]
- Isman, M.B. A renaissance for botanical insecticides? Pest Manag. Sci. 2015, 71, 1587–1590. [Google Scholar] [CrossRef] [PubMed]
- Pavela, R.; Benelli, G. Essential oils as ecofriendly biopesticides? Challenges and constraints. Trends Plant Sci. 2016, 21, 1000–1007. [Google Scholar] [CrossRef] [PubMed]
- Govindarajan, M.; Mathivanan, T.; Elumalai, K.; Krishnappa, K.; Anandan, A. Ovicidal and repellent activities of botanical extracts against Culex quinquefasciatus, Aedes aegypti and Anopheles stephensi (Diptera: Culicidae). Asian Pac. J. Trop. Biomed. 2011, 1, 43–48. [Google Scholar] [CrossRef]
- Benelli, G.; Maggi, F.; Pavela, R.; Murugan, K.; Govindarajan, M.; Vaseeharan, B.; Petrelli, R.; Cappellacci, L.; Kumar, S.; Hofer, A.; et al. Mosquito control with green nanopesticides: Towards the one health approach? A review of non-target effects. Environ. Sci. Pollut. Res. 2017, 25, 10184–10206. [Google Scholar] [CrossRef] [PubMed]
- Webster, A.E.; Manning, P.; Sproule, J.M.; Faraone, N.; Cutler, G.C. Insecticidal and synergistic activity of two monoterpenes against diamondback moth (Lepidoptera: Plutellidae). Can. Entomol. 2018, 150, 258–264. [Google Scholar] [CrossRef]
- Bakkali, F.; Averbeck, S.; Averbeck, D.; Idaomar, M. Biological effects of essential oils—A review. Food Chem. Toxicol. 2008, 46, 446–475. [Google Scholar] [CrossRef]
- Tripathi, A.K.; Upadhyay, S.; Bhuiyan, M.; Bhattacharya, P.R. A review on prospects of essential oils as biopesticide in insect-pest management. J. Pharmacogn. Phytother. 2009, 1, 52–63. [Google Scholar]
- Gonzalez-Coloma, A.; Reina, M.; Diaz, C.E.; Fraga, B.M. Comprehensive Natural Products Chemistry; Mander, L., Liu, H., Eds.; Elsevier: Amsterdam, The Netherlands, 2009; Volume 3, pp. 237–268. [Google Scholar]
- Almeida, R.N.; Agra, M.D.F.; Souto Maior, F.N.; Sousa, D.P. Essential oils and their constituents: Anticonvulsant activity. Molecules 2011, 16, 2726–2742. [Google Scholar] [CrossRef]
- Tak, J.H.; Jovel, E.; Isman, M.B. Contact, fumigant, and cytotoxic activities of thyme and lemongrass essential oils against larvae and an ovarian cell line of the cabbage looper, Trichoplusia ni. J. Pest Sci. 2016, 89, 183–193. [Google Scholar] [CrossRef]
- Park, J.H.; Jeon, Y.J.; Lee, C.H.; Chung, N.; Lee, H.S. Insecticidal toxicities of carvacrol and thymol derived from Thymus vulgaris Lin. against Pochazia shantungensis Chou & Lu., newly recorded pest. Sci. Rep. 2017, 7, 40902. [Google Scholar] [PubMed]
- Mètomè, G.; Adjou, E.S.; Gbaguidi, B.A.; Dahouenon-Ahoussi, E.; Sohounhloue, D. Insecticidal and repellent effects of essential oils of Cynnamomum zeylanicum and Syzygium aromaticum from Benin against the weevil of Pigeon pea (Cajanus cajan L. Millspaugh). J. Pharmacogn. Phytochem. 2010, 6, 2074–2077. [Google Scholar]
- Li, S.G.; Zhou, B.G.; Li, M.Y.; Liu, S.; Hua, R.M.; Lin, H.F. Chemical composition of Illicium verum fruit extract and its bioactivity against the peach–potato aphid, Myzus persicae (Sulzer). Arthropod-Plant Interact. 2017, 11, 203–212. [Google Scholar] [CrossRef]
- Chansang, A.; Champakaew, D.; Junkum, A.; Amornlerdpison, D.; Chaithong, U.; Jitpakdi, A.; Riyong, D.; Wannasan, A.; Intirach, J.; Muangmoon, R.; et al. Potential of natural essential oils and Cinnamaldehyde as insecticides against the dengue vector Aedes aegypti (Diptera: Culicidae). Southeast Asian J. Trop. Med. Public Health 2018, 49, 6–22. [Google Scholar]
- Garcia, J.F.; Grisoto, E.; Vendramim, J.D.; Botelho, P.S.M. Bioactivity of neem, Azadirachta indica, against spittlebug Mahanarva fimbriolata (Hemiptera: Cercopidae) on sugarcane. J. Econ. Entomol. 2006, 99, 2010–2014. [Google Scholar] [CrossRef] [PubMed]
- Jankowska, M.; Rogalska, J.; Wyszkowska, J.; Stankiewicz, M. Molecular targets for components of essential oils in the insect nervous system—A review. Molecules 2017, 23, 34. [Google Scholar] [CrossRef] [PubMed]
- Peck, D.C. Distribucion y reconocimiento del salivazo de los pastos (Homoptera: Cercopidae) en la Costa Caribe de Colômbia. Pasturas Tropicale 2002, 24, 4–15. [Google Scholar]
- Brasil. Manual de Testes de Eficácia em Produtos Desinfestantes; Anvisa, Ed.; Agência Nacional de Vigilância Sanitária: Brasília, Brazil, 2009; p. 50.
- Abbott, W.S. A method of computing the effectiveness of an insecticide. J. Econ. Entomol. 1925, 18, 265–267. [Google Scholar] [CrossRef]
- RStudio. RStudio: Integrated Development Environment for R; R Studio, Inc.: Boston, MA, USA, 2018. [Google Scholar]
- R Core Team 3.5.1. R: A Language and Environment for Statistical Computing; R Foundation for Statistical Computing: Vienna, Austria, 2018. [Google Scholar]
- Moreira, M.D.; Picanço, M.C.; Barbosa, L.C.A.; Guedes, R.N.C.; Barros, E.C.; Campos, M.R. Compounds from Ageratum conyzoides: Isolation, structural elucidation and insecticidal activity. Pest Manag. Sci. 2007, 63, 615–621. [Google Scholar] [CrossRef]
- Caperucci, D.; Camargo-Mathias, M.I. Ultrastructural study of the ovary of the sugarcane spittlebug Mahanarva fimbriolata (Hemiptera). Micron 2006, 37, 633–639. [Google Scholar] [CrossRef]
- Beament, J.W.L. The role of cuticle and egg-shell membranes in the penetration of insecticides. Ann. Appl. Biol. 1952, 39, 142–143. [Google Scholar] [CrossRef]
- Figueiredo, M.B.; Gomes, G.A.; Santangelo, J.M.; Pontes, E.G.; Azambuja, P.; Garcia, E.S.; Carvalho, M.G. Lethal and sublethal effects of essential oil of Lippia sidoides (Verbenaceae) and monoterpenes on Chagas’ disease vector Rhodnius prolixus. Mem. Inst. Oswaldo Cruz 2017, 112, 63–69. [Google Scholar] [CrossRef] [PubMed]
- Hong, T.K.; Perumalsamy, H.; Jang, K.H.; Na, E.S.; Ahn, Y.J. Ovicidal and larvicidal activity and possible mode of action of phenylpropanoids and ketone identified in Syzygium aromaticum bud against Bradysia procera. Pestic. Biochem. Physiol. 2018, 145, 29–38. [Google Scholar] [CrossRef] [PubMed]
- Priestley, C.M.; Williamson, E.M.; Wafford, K.A.; Sattelle, D.B. Thymol, a constituent of thyme essential oil, is a positive allosteric modulator of human GABAA receptors and a homo-oligomeric GABA receptor from Drosophila melanogaster. Br. J. Pharmacol. 2003, 140, 1363–1372. [Google Scholar] [CrossRef] [PubMed]
- Enan, E.E. Molecular and pharmacological analysis of an octopamine receptor from American cockroach and fruit fly in response to plant essential oils. Arch. Insect Biochem. Physiol. 2005, 59, 161–171. [Google Scholar] [CrossRef] [PubMed]
- Enan, E.E. Molecular response of Drosophila melanogaster tyramine receptor cascade to plant essential oils. Insect Biochem. Mol. Biol. 2005, 35, 309–321. [Google Scholar] [CrossRef] [PubMed]
- Anderson, J.A.; Coats, J.R. Acetylcholinesterase inhibition by nootkatone and carvacrol in arthropods. Pestic. Biochem. Physiol. 2012, 102, 124–128. [Google Scholar] [CrossRef] [Green Version]
- Obeng-Ofori, D.; Reichmuth, C.H. Bioactivity of eugenol, a major component of essential oil of Ocimum suave (Wild.) against four species of stored-product Coleoptera. Int. J. Pest Manag. 1997, 43, 89–94. [Google Scholar] [CrossRef]
- Enan, E. Insecticidal activity of essential oils: Octopaminergic sites of action. Comp. Biochem. Physiol. Part C Toxicol. Pharmacol. 2001, 130, 325–337. [Google Scholar] [CrossRef]
- Kovaříková, K.; Holý, K.; Skuhrovec, J.; Saska, P. The efficacy of insecticides against eggs and nymphs of Aleyrodes proletella (Hemiptera: Aleyrodidae) under laboratory conditions. Crop Prot. 2017, 98, 40–45. [Google Scholar] [CrossRef]
- Whittaker, J.B. Cercopid spittle as a microhabitat. Oikos 1970, 21, 59–64. [Google Scholar] [CrossRef]
- Henderson, G.; Hoffman, G.D.; Jeanne, R.L. Predation on cercopids and material use of the spittle in aphid-tent construction by prairie ants. Psyche 1990, 97, 43–53. [Google Scholar] [CrossRef]
- Tonelli, M.; Gomes, G.; Silva, W.D.; Magri, N.T.; Vieira, D.M.; Aguiar, C.L.; Bento, J.M.S. Spittlebugs produce foam as a thermoregulatory adaptation. Sci. Rep. 2018, 8, 4729. [Google Scholar] [CrossRef] [PubMed]
- Chen, X.; Meyer-Rochow, V.B.; Fereres, A.; Morente, M.; Liang, A.P. The role of biofoam in shielding spittlebug nymphs (Insecta, Hemiptera, Cercopidae) against bright light. Ecol. Entomol. 2018, 43, 273–281. [Google Scholar] [CrossRef]
- Moretti, A.N.; Zerba, E.N.; Alzogaray, R.A. Behavioral and toxicological responses of Rhodnius prolixus and Triatoma infestans (Hemiptera: Reduviidae) to 10 monoterpene alcohols. J. Med. Entomol. 2013, 50, 1046–1054. [Google Scholar] [CrossRef] [PubMed]
- Camarillo, G.R.; Ortega, A.L.D.; Serrato, C.M.A.; Rodriguez, H.C. Biological activity of Tagetes filifolia (Asteraceae) on Trialeurodes vaporariorum (Hemiptera: Aleyrodidae). Rev. Colomb. Entomol. 2009, 35, 177–184. [Google Scholar]
- Cruz, G.S.; Wanderley-Teixeira, V.; Oliveira, J.V.; D’assunção, C.G.; Cunha, F.M.; Teixeira, Á.A.; Guedes, C.A.; Dutra, K.A.; Barbosa, D.R.S.; Breda, M.O. Effect of trans-anethole, limonene and your combination in nutritional components and their reflection on reproductive parameters and testicular apoptosis in Spodoptera frugiperda (Lepidoptera: Noctuidae). Chem.-Biol. Interact. 2017, 263, 74–80. [Google Scholar] [CrossRef] [PubMed]
- Hashem, A.S.; Awadalla, S.S.; Zayed, G.M.; Maggi, F.; Benelli, G. Pimpinella anisum essential oil nanoemulsions against Tribolium castaneum—Insecticidal activity and mode of action. Environ. Sci. Pollut. Res. 2018, 25, 18802–18812. [Google Scholar] [CrossRef]
- López, M.D.; Pascual-Villalobos, M.J. Mode of inhibition of acetylcholinesterase by monoterpenoids and implications for pest control. Ind. Crops Prod. 2010, 31, 284–288. [Google Scholar] [CrossRef]
- Kim, J.R.; Jeong, I.H.; Lee, Y.S.; Lee, S.G. Insecticidal activity of cinnamon essential oils, constituents, and (E) -cinnamaldehyde analogues against Metcalfa pruinosa Say (Hemiptera: Flatidae) nymphs and adults. Korean J. Appl. Entomol. 2015, 54, 375–382. [Google Scholar] [CrossRef]
- Batista, E.; Auad, A.M.; Resende, T.T.; Frias, M.P.; Monteiro, C.M.O. Nematoides entomopatogênicos infectam ovos e adultos de cigarrinha-das-pastagens. Arq. Inst. Biol. 2011, 78, 475–478. [Google Scholar]
- Park, C.G.; Shin, E.; Kim, J. Insecticidal activities of essential oils, Gaultheria fragrantissima and Illicium verum, their components and analogs against Callosobruchus chinensis adults. J. Asia-Pac. Entomol. 2016, 19, 269–273. [Google Scholar] [CrossRef]
- Andrade-Ochoa, S.; Sánchez-Aldana, D.; Chacón-Vargas, K.F.; Rivera-Chavira, B.E.; Sánchez-Torres, L.E.; Camacho, A.D.; Nogueda-Torres, B.; Nevárez-Moorillón, G.V. Oviposition deterrent and larvicidal and pupaecidal activity of seven essential oils and their major components against Culex quinquefasciatus Say (Diptera: Culicidae): Synergism—Antagonism effects. Insects 2018, 9, 25. [Google Scholar] [CrossRef] [PubMed]
- Pandey, S.K.; Upadhyay, S.; Tripathi, A.K. Insecticidal and repellent activities of thymol from the essential oil of Trachyspermum ammi (Linn) Sprague seeds against Anopheles stephensi. Parasitol. Res. 2009, 105, 507–512. [Google Scholar] [CrossRef] [PubMed]
- Oliveira, A.P.; Santos, A.A.; Santana, A.S.; Lima, A.P.S.; Melo, C.R.; Santana, E.D.; Sampaio, T.S.; Blank, A.F.; Araújo, A.P.A.; Cristaldo, P.F.; et al. Essential oil of Lippia sidoides and its major compound thymol: Toxicity and walking response of populations of Sitophilus zeamais (Coleoptera: Curculionidae). Crop Prot. 2018, 112, 33–38. [Google Scholar] [CrossRef]
© 2019 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Dias, M.L.; Auad, A.M.; Magno, M.C.; Resende, T.T.; Fonseca, M.G.; Silva, S.E.B. Insecticidal Activity of Compounds of Plant Origin on Mahanarva spectabilis (Hemiptera: Cercopidae). Insects 2019, 10, 360. https://doi.org/10.3390/insects10100360
Dias ML, Auad AM, Magno MC, Resende TT, Fonseca MG, Silva SEB. Insecticidal Activity of Compounds of Plant Origin on Mahanarva spectabilis (Hemiptera: Cercopidae). Insects. 2019; 10(10):360. https://doi.org/10.3390/insects10100360
Chicago/Turabian StyleDias, Marcelle L., Alexander M. Auad, Milena C. Magno, Tiago T. Resende, Marcy G. Fonseca, and Sandra E. B. Silva. 2019. "Insecticidal Activity of Compounds of Plant Origin on Mahanarva spectabilis (Hemiptera: Cercopidae)" Insects 10, no. 10: 360. https://doi.org/10.3390/insects10100360