Cleonis pigra (Scopoli, 1763) (Coleoptera: Curculionidae: Lixinae): Morphological Re-Description of the Immature Stages, Keys, Tribal Comparisons and Biology


Abstract
:1. Introduction
2. Materials and Methods
2.1. Insect Collection and Laboratory Breeding
- (1)
- Sheep pasture on the alluvial floodplain, near the Kamyana Mohyla Reserve (46°57′01″ N, 35°28′12″ E). Altitude: up to 11 m.a.s.l. Bedrock: sandy chernozem. Dominant plant species: Cirsium ukranicum Besser ex DC., and Carduus hamulosus Ehrh. (syn. C. pseudocollinus (Schmalh.) Klokov.), with occasional trees of Elaeagnus commutata Bernh. ex Rydb. growing among the grass (i.e., Echinops ritro subsp. ruthenicus L. (M. Bieb.) Nyman (syn. E. ruthenicus), and Centaurea adpressa Ledeb. ex Steud., Melilotus albus Medik.) in the clearings.
- (2)
- Pishchanska Balka near Melitopol (46°49′50″ N, 35°20′18″ E). Altitude: up to 22 m.a.s.l. Bedrock: sandy chernozem with herbaceous covering. Dominant plant species: Calamagrostis epigeios (L.) Roth, Linaria genistifolia (L.) Mill., Artemisia absinthium L., Echium vulgare L., Sisymbrium loeselii L., Achillea millefolium L., Hieracium umbellatum L., Melilotus albus Medik., Centaurea diffusa Lam., Cirsium arvense (L.) Scop. (syn. C. setosum (Willd.) Besser), and Chenopodium urbicum L.
2.2. Morphological Description
3. Results and Discussion
3.1. The Morphology of Immature Stages of Cleonis pigra
3.1.1. Materials Examined
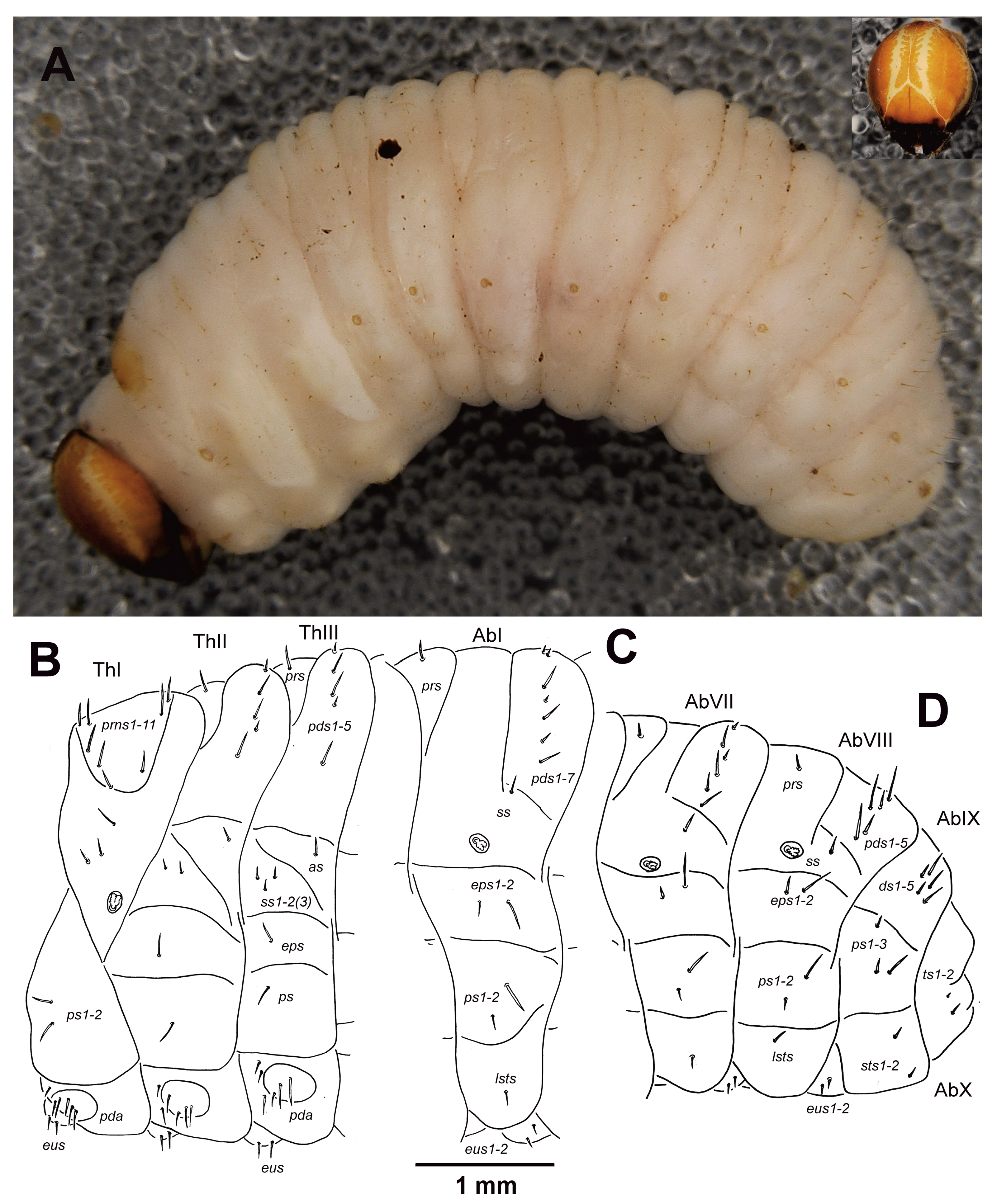
3.1.2. Description of Mature Larvae
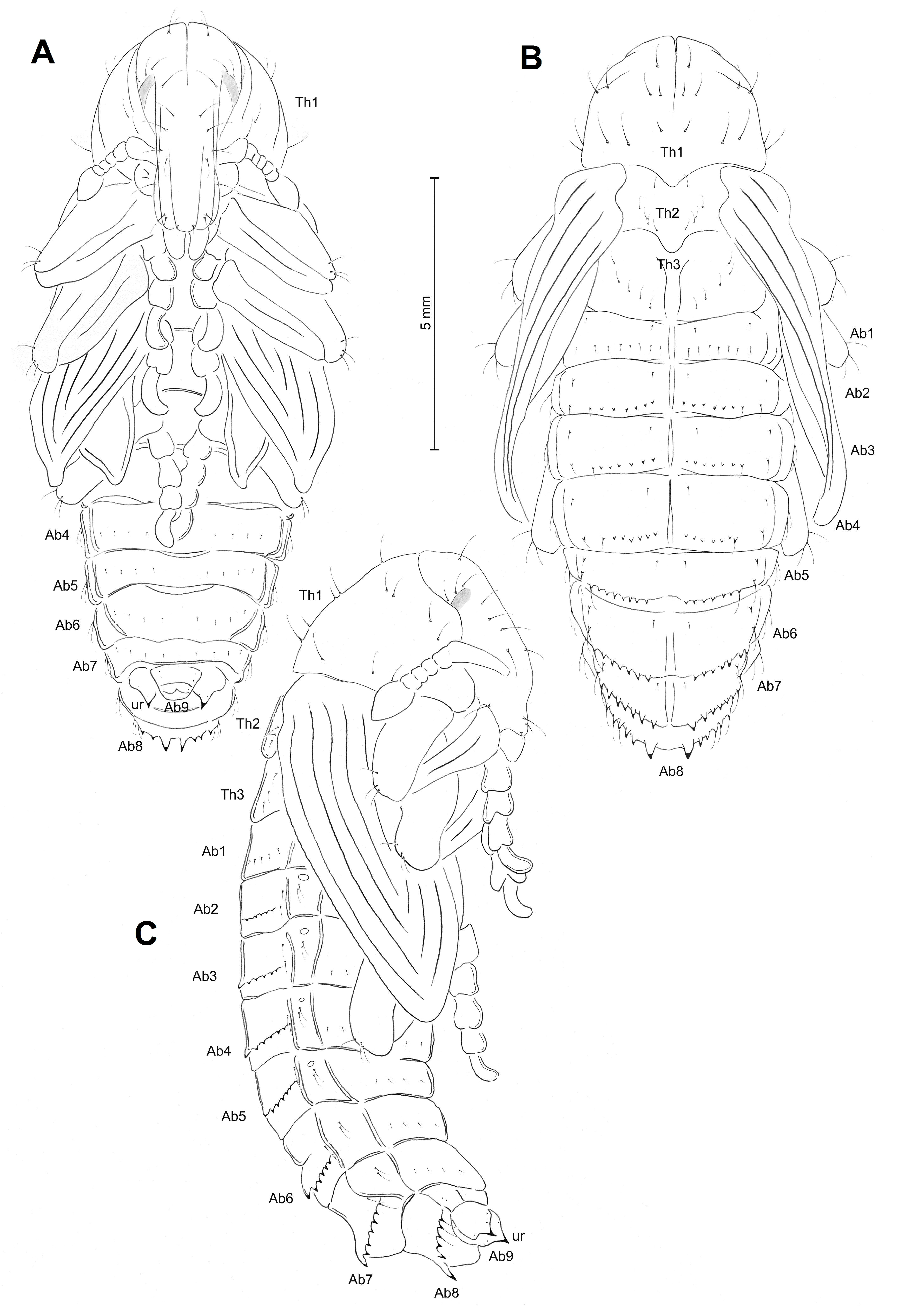
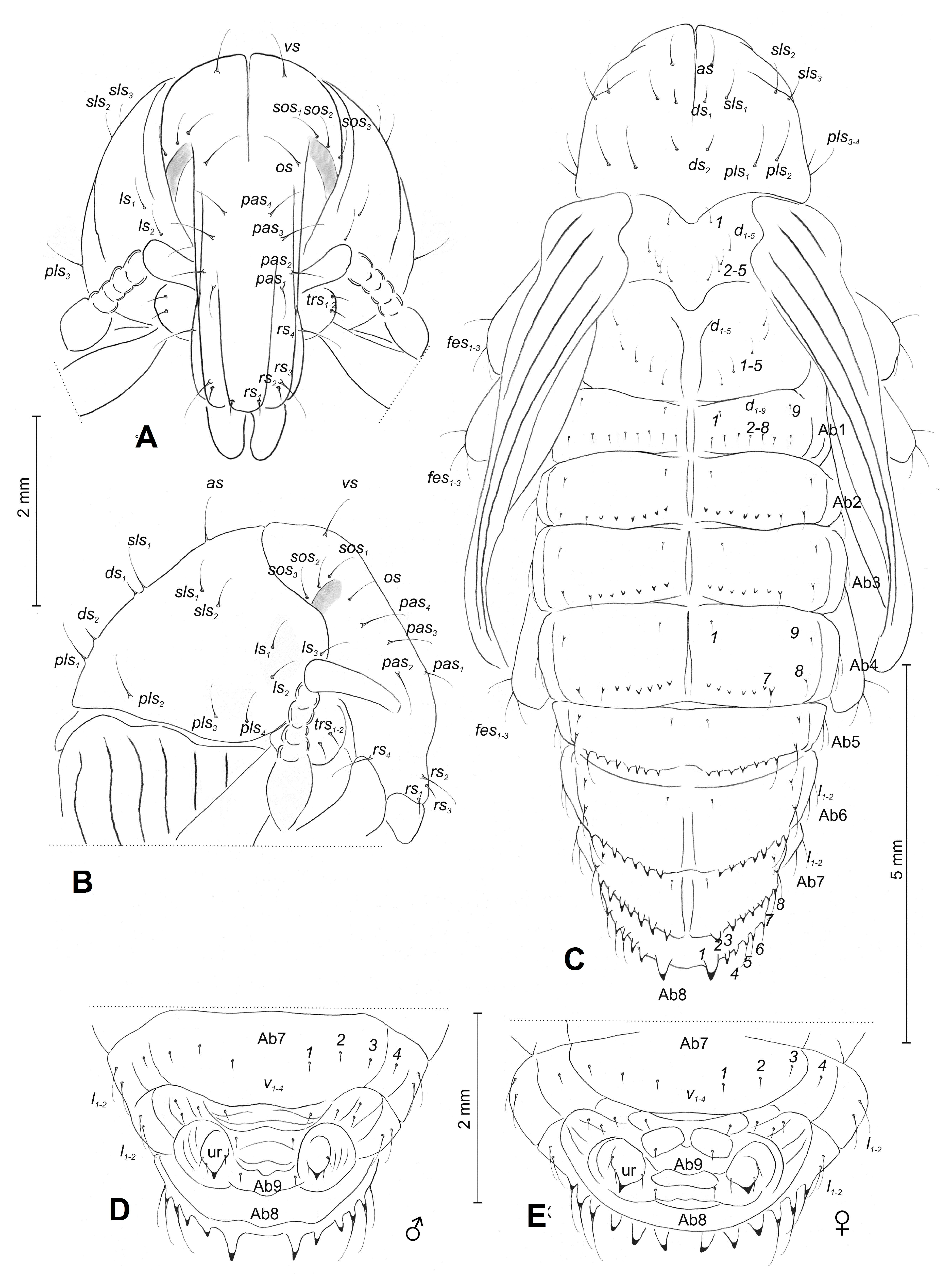
3.1.3. Description of Pupae
3.1.4. Remarks on the Previous Descriptions of Immature Stages of Cleonis pigra
3.1.5. A Comparison with Larvae of Other Cleonini
3.1.6. Key to the Known Immature Stages of Cleonini
Larvae (Last Instar)
Pupae
3.1.7. Measurements of Larval Instars
3.2. Biology and Ecology of Cleonis pigra
3.2.1. Habitats
3.2.2. Adult Behavior
3.2.3. Host Plant
3.2.4. Life Cycle
3.2.5. Biotic Interactions
3.2.6. What Are the Other Effects of the Weevil?
4. Conclusions
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
References
- Meregalli, M. 3.7.6. Lixinae Schoenherr, 1823. In Handbook of Zoology; Arthropoda: Insecta. Coleoptera, Beetles: Morphology and Systematics; Kristensen, N.P., Beutel, R.G., Eds.; De Gruyter: Göttingen, Germany, 2014; Volume 3, pp. 523–529. [Google Scholar]
- Arzanov, Y.G.; Grebennikov, V.V. Cleonini (Coleoptera: Curculionidae: Lixinae) are monophyletic and flightless: tribe overview, rampant adult homoplasy and illustrated global diversity. Zootaxa 2017, 4329, 1–63. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Gültekin, L.; Fremuth, J. Tribe Lixini. In Catalogue of Palaearctic Coleoptera, Volume 8, Curculionoidea II; Löbl, I., Smetana, A., Eds.; Brill: Leiden, The Netherlands, 2013; pp. 456–472. [Google Scholar]
- Anderson, R.S. Purealus beckelorum, a new genus and species of cleonine weevil from western Texas and eastern New Mexico (Coleoptera, Curculionidae, Lixinae, Cleonini). ZooKeys 2018, 785, 1–10. [Google Scholar] [CrossRef] [PubMed]
- Alonso-Zarazaga, M.A.; Lyal, C.H.C. A World Catalogue of Families and Genera of Curculionoidea (Insecta Coleoptera) (excepting Scolytidae and Platypodidae); Entomopraxis: Barcelona, Spain, 1999; p. 315. [Google Scholar]
- Meregalli, M.; Fremuth, J. Tribe Cleonini. In Catalogue of Palaearctic Coleoptera, Volume 8, Curculionoidea II; Löbl, I., Smetana, A., Eds.; Brill: Leiden, The Netherlands, 2013; pp. 437–456. [Google Scholar]
- Meregalli, M. World catalogue of the Curculionidae: Lixinae: Cleonini 2017. Available online: http://weevil.info/content/world-catalogue-curculionidae-lixinae-cleonini# (accessed on 1 February 2019).
- Volovnik, S.V. Weevils Lixinae (Coleoptera, Curculionidae) as Gall Formers. Entomol. Rev. 2010, 90, 585–590, [Original Russian text published in Zoologicheskiy Zhurnal, 2010, 89, 828–833]. [Google Scholar] [CrossRef]
- Trnka, F.; Stejskal, R.; Skuhrovec, J. Biology and morphology of immature stages of Adosomus roridus (Coleoptera: Curculionidae: Lixinae). Zootaxa 2015, 4021, 433–446. [Google Scholar] [CrossRef]
- Zotov, A.A. Morphology of the preimaginal stages of weevils of the tribe Cleonini sensu lato (Coleoptera: Curculionidae). Cauc. Entomol. Bull. 2011, 7, 153–162. [Google Scholar] [CrossRef]
- Stejskal, R.; Trnka, F.; Skuhrovec, J. Biology and morphology of immature stages of Coniocleonus nigrosuturatus (Coleoptera: Curculionidae: Lixinae). Acta Entomol. Musei Nat. Pragae 2014, 54, 337–354. [Google Scholar]
- Stinson, C.S.A.; Schroeder, D.; Marquardt, K. Investigations on Cyphocleonus achates (Fahr.) (Col., Curculionidae), a potential biological control agent of spotted knapweed (Centaurea maculosa Lam.) and diffuse knapweed (C. diffusa Lam.) Compositae in North America. J. Appl. Entomol. 1994, 117, 35–50. [Google Scholar] [CrossRef]
- Story, J.M.; Callan, N.W.; Corn, J.G.; White, L.J. Decline of spotted knapweed density at two sites in western Montana with large populations of the introduced root weevil, Cyphocleonus achates (Fahraeus). Biol. Control 2006, 38, 227–232. [Google Scholar] [CrossRef]
- Huber, J.T.; Vayssieres, J.F. Life cycle and host specificity of the Heliotrope weevil, Pachycerus cordiger (=madius auct.) [Col.: Curculionidae]. Entomophaga 1990, 35, 475–484. [Google Scholar] [CrossRef]
- Louda, S.M.; Pemberton, R.W.; Johnson, M.T.; Follett, P.A. Nontarget Efects—The Achilles’ Heel of Biological Control? Retrospective Analyses to Reduce Risk Associated with Biocontrol Introductions. Ann. Rev. Entomol. 2003, 48, 365–396. [Google Scholar] [CrossRef]
- Alonso-Zarazaga, M.A.; Barrios, H.; Borovec, R.; Bouchard, P.; Caldara, R.; Colonnelli, E.; Gultekin, L.; Hlavač, P.; Korotyaev, B.; Lyal, C.H.C.; et al. Cooperative Catalogue of Palaerctic Coleoptera Curculionoidea. Monogr. Electron. SEA 2017, 8, 729. [Google Scholar]
- Sanwal, N.; Kumar, T.; Kumar, A.; Sharma, V. Diversity, distribution and host plants of Cleonini (Curculionidae: Lixinae) in India. In National Symposium on Entomology as a Science and IPM as a Technology—The Way Forward; Ramamurthy, V.V., Subramanin, S., Eds.; Pasighat, Arunanchal Pradesh, Entomological Society of India: New Delhi, India, 2014; pp. 17–18. [Google Scholar]
- Siitonen, J.; Silfverberg, H. Kovakuoriaisten maakuntaluettelo 2015: Provincial List of Finnish Coleoptera 2015. Sahlbergia 2015, 21 (Suppl. 1), 164. [Google Scholar]
- Averenskiy, A.I. The Catalogue of Beetles of Yakutia; Yakutsk Scientific Center SD RAS Publ.: Yakutsk, Russia, 1999; p. 96. [Google Scholar]
- Yunakov, N.; Nazarenko, V.; Filimonov, R.; Volovnik, S. A survey of the weevils of Ukraine (Coleoptera: Curculionoidea) (excluding Platypodinae and Scolytinae). Zootaxa 2018, 4404, 494. [Google Scholar] [CrossRef] [PubMed]
- Ødegaard, J.A.; Oddvar, H.; Torstein, K.; Stefan, O. Coleoptera. In The 2010 Norwegian Red List for Species; Kålås, J.A., Viken, Å., Henriksen, S., Skjelseth, S., Eds.; Norwegian Biodiversity Information Centre: Trondheim, Norway, 2010; pp. 257–290. [Google Scholar]
- Hyvärinen, E.; Mannerkoski, I.; Clayhills, T.; Helve, E.; Karjalainen, S.; Laurinharju, E.; Martikainen, P.; Mattila, J.; Muona, J.; Pentinsaari, M.; et al. Beetles Coleoptera. In The 2010 Red List of Finnish Species; Rassi, P., Hyvarinen, E., Juslen, A., Mannerkoski, I., Eds.; Ymparistoministerio & Suomen Ymparistokeskus: Helsinki, Finland, 2010; pp. 545–582. [Google Scholar]
- Anderson, R.S. Systematics, phylogeny and biogeography of New World weevils traditionally of the tribe Cleonini (Coleoptera: Curculionidae; Cleoninae). Quaest. Entomol. 1987, 23, 431–709. [Google Scholar]
- Lindroth, C.H. The Faunal Connections between Europe and North America; Wiley & Sons Inc., Almquist & Wicksell: Stockholm, Sweden, 1957; p. 334. [Google Scholar]
- Cawthra, E.M. The Occurrence of Cleonus piger Scop. (Col., Curculionidae) at Aberlady Bay, East Lothian, with Some Notes upon Its Larval Instars. Entomol. Mon. Mag. 1958, 94, 204–206. [Google Scholar]
- O’Brien, C.W.; Wibmer, G.J. Annotated checklist of the weevils (Curculionidae sensu lato) of North America, Central America, and the West Indies (Coleoptera: Curculionoidea). Mem. Am. Ent. Inst. 1982, 34, 188. [Google Scholar]
- Scherf, H. Die Entwicklungsstadien der mitteleuropäischen Curculioniden (Morphologie, Bionomie, Ökologie). Abh. Senckenberg. Naturf. Ges. 1964, 506, 1–335. [Google Scholar]
- Peschken, D.P. Cirsium arvense (L.) Scop., Canada thistle (Compositae). In Programmes against Insects and Weeds in Canada 1969–1980; Kelleher, J.S., Hulme, M.A., Eds.; Commonwealth Agricultural Bureaux: London, UK, 1984; pp. 139–146. [Google Scholar]
- La Ferla, A. Contributo alla conoscenza dei cleono del carciofo (Cleonus piger Scop.). Boll. Lab. Entomol. Agrar. Portici 1939, 3, 25–33. [Google Scholar]
- Ortu, S.; Lentini, A.; Foxi, C.; Murgia, G. I principali parassiti animali del carciofo e relative metodi di lotta. In Filiera Agro Alimentare del Medio Campi Dano. 1. Il Carciofo: Storia, Tecnica Colturale, Politica Comunitaria, Ricerca, Trasformazione; ERSAT, Servizio Territoriale del Medio Campidano Sanluri, Centro Zonale di Serramanna: Sardinia, Italy, 2003; pp. 136–173. [Google Scholar]
- Cripps, M.G.; Gassmann, A.; Fowler, S.V.; Bourdot, G.W.; McClay, A.S.; Edwards, G.R. Classical biological control of Cirsium arvense: Lessons from the past. Biol. Control 2011, 57, 165–174. [Google Scholar] [CrossRef]
- Andrzejewska, J.; Lamparski, R.; Skinder, Z. Yield and quality of raw material of milk thistle (Silybum marianum [L.] Gaertn.) grown in monoculture and in crop rotation. Herba Pol. 2006, 52, 11–17. [Google Scholar]
- Luchnik, V. Vrednye nasekomye Stavropolskogo okruga v 1928 g. [Pests of Stavropol District in 1928]. Raboty Stavropolskoj Stantsii Zashchity Rastenij 1929, 2, 1–120. [Google Scholar]
- Tamarina, N.A. K biologii nasekomykh, vrediashchikh v usloviyakh stepnogo lesorazvedeniya (To biology of insects that damage to man-made forests in the steppe]. Zool. Zhurnal 1957, 36, 1187–1198. (In Russian) [Google Scholar]
- Tielecke, H. Ein Beitrag zur Biologie des Distelrulers (Cleonus piger Scop.). Nachrbl. Deut. Pflanzenschutzd. 1951, 5, 31–35. [Google Scholar]
- Skuhrovec, J.; Bogusch, P. The morphology of the immature stages of Metadonus vuillefroyanus (Capiomont, 1868) (Coleoptera, Curculionidae, Hyperini) and notes on its biology. Zookeys 2016, 589, 123–142. [Google Scholar] [CrossRef]
- May, B.M. An introduction to the immature stages of Australian Curculionoidea, subfamily Bagoinae. In Australian Weevils (Coleoptera: Curculionidae); Zimmerman, E.C., Ed.; CSIRO: Melbourne, Australia, 1994; Volume 2, pp. 533–535. [Google Scholar]
- May, B.M. Immature stages of Curculionidae: larvae of soil dwelling weevils of New Zealand. J. R. Soc. N. Z. 1977, 72, 189–228. [Google Scholar] [CrossRef]
- Marvaldi, A.E. Larvae of South American Rhytirrhininae (Coleoptera: Curculionidae). Coleopts. Bull. 1998, 52, 71–89. [Google Scholar]
- Marvaldi, A.E. Morfología larval en Curculionidae (Insecta: Coleoptera). Acta Zool. Lilloana 1999, 45, 7–24. [Google Scholar]
- Caldara, R.; Skuhrovec, J.; Gosik, R.; Marvaldi, A.E. On the affinities and systematic position of Pachytychius Jekel, a genus currently incertae sedis in Coleoptera, Curculionidae: evidence from immature stages and the COI gene support its placement in Smicronychini. Zool. Anz. 2018, 277, 218–230. [Google Scholar] [CrossRef]
- Hoffmann, A. Fauna de France 52 Coléoptères curculionides (Première partie); Lechevalier: Paris, France, 1950; p. 486. [Google Scholar]
- Anderson, R.S. Larva and pupa of Cleonidius erysimi (Fall) with a discussion of the phylogenetic position of Lixini (sensu Kuschel) (Coleoptera: Curculionidae). Proc. Entomol. Soc. Wash. 1991, 93, 288–297. [Google Scholar]
- May, B.M. Fauna of New Zealand, Larvae of Curculionoidea (Insecta: Coleoptera): A Systematic Overview; Manaaki Whenua Press: Lincoln, New Zealand, 1993; Volume 28, p. 226. [Google Scholar]
- Zotov, A.A. Morphology of the preimaginal stages of three species of weevil of the Lixini (Coleoptera: Curculionidae). Cauc. Entomol. Bull. 2009, 5, 81–90, [Original text published in Russian in Kavkazskiy Entomologicheskiy Byulleten, 5, 81–90]. [Google Scholar] [CrossRef]
- Zotov, A.A. Morphology of preimaginal stages of the genus Larinus Dejean, 1821 (Coleoptera: Curculionidae). Part I. Cauc. Entomol. Bull. 2010, 6, 171–178. [Google Scholar] [CrossRef]
- Gosik, R.; Skuhrovec, J. Descriptions of mature larvae and pupae of the genus Larinus (Coleoptera: Curculionidae, Lixinae). Zootaxa 2011, 3019, 1–25. [Google Scholar] [CrossRef]
- Skuhrovec, J.; Volovnik, S.; Gosik, R. Description of the immature stages of Larinus vulpes and notes on its biology (Coleoptera: Curculionidae: Lixinae). ZooKeys 2017, 679, 107–137. [Google Scholar] [CrossRef]
- Lee, C.Y.; Morimoto, K. Larvae of the family curculionidae of Japan. Part 2. Hyperinae to Cioninae (Insecta: Coleoptera). J. Fac. Agric. Kyushu Univ. 1988, 33, 131–152. [Google Scholar]
- Nikulina, O.N. Larval morphology of the weevil genus Lixus (Coleoptera, Curculionidae) from Middle Asia. Entomol. Rev. 2001, 81, 809–823, [Original text published in Russian in Zool. Zhurnal, 80, 183–195]. [Google Scholar]
- Nikulina, O.N. New data on larvae of weevils of the genus Lixus (Coleoptera, Curculionidae) from Central Asia. Entomol. Rev. 2007, 87, 750–756, [Original text published in Russian in Zool. Zhurnal, 86, 1086–1092]. [Google Scholar] [CrossRef]
- Zotov, A.A. Morphology of the preimaginal stages of weevil Lixus iridis Olivier, 1807 (Coleoptera: Curculionidae). Cauc. Entomol. Bull. 2009, 5, 249–252, [Original text published in Russian in Kavkazskiy Entomologicheskiy Byulleten, 5, 249–252]. [Google Scholar] [CrossRef]
- Nikulina, O.N.; Gültekin, L. Larval morphology of Lixus cardui Olivier and Lixus filiformis (Fabricius) (Coleoptera: Curculionidae): biological control agents for Scotch and musk thistles. Aust. J. Entomol. 2011, 50, 253–257. [Google Scholar] [CrossRef]
- Gosik, R.; Wanat, M. Descriptions of immature stages of the weevil Lixus punctiventris Boheman, 1835 (Coleoptera, Curculionidae, Lixini). Zootaxa 2014, 3754, 159–172. [Google Scholar] [CrossRef]
- Skuhrovec, J.; Volovnik, S. Biology and morphology of immature stages of Lixus canescens (Coleoptera: Curculionidae: Lixinae). Zootaxa 2015, 4033, 350–362. [Google Scholar] [CrossRef]
- Gosik, R.; Sprick, P.; Skuhrovec, J.; Derus, M.; Hommes, M. Morphology and identification of the mature larvae of several species of the genus Otiorhynchus (Coleoptera, Curculionidae, Entiminae) from Central Europe with an update of life history traits. Zootaxa 2016, 4108, 1–67. [Google Scholar] [CrossRef] [PubMed]
- Skuhrovec, J.; Gosik, R.; Caldara, R. Immatures of Palaearctic species of the weevil genus Tychius (Coleoptera, Curculionidae): New descriptions and new bionomic data with an evaluation of their value in a phylogenetic reconstruction of the genus. Zootaxa 2014, 3839, 1–84. [Google Scholar] [CrossRef] [PubMed]
- Skuhrovec, J.; Gosik, R.; Caldara, R.; Košťál, M. Immatures of Palaearctic species of the weevil genus Sibinia (Coleoptera, Curculionidae): New descriptions and new bionomic data with suggestions on their potential value in a phylogenetic reconstruction of the genus. Zootaxa 2015, 3955, 151–187. [Google Scholar] [CrossRef] [PubMed]
- Gosik, R.; Skuhrovec, J.; Toševski, I.; Caldara, R. Morphological evidence from immature stages further suggests Lignyodina being close to Tychiina (Coleoptera, Curculionidae, Curculioninae, Tychiini). Zootaxa 2017, 4320, 426–446. [Google Scholar] [CrossRef]
- Skuhrovec, J.; Gosik, R.; Caldara, R.; Toševski, I.; Łętowski, J.; Szwaj, E. Importance of morphological characters of immatures for a better distinction of Palaearctic species of Cleopomiarus and Miarus from each other and other Mecinini (Coleoptera, Curculionidae, Curculioninae). ZooKeys 2018, 808, 23–92. [Google Scholar] [CrossRef] [PubMed]
- Rowe, D.J.; Kok, L.T. Determination of larval instars, and comparison of field and artifical diet-reared larval stages of Rhinocyllus conicus (Col: Curculionidae). Va. J. Sci. 1985, 36, 277–280. [Google Scholar]
- Torres, B.C.P.; Skuhrovec, J.; Marín-Cevada, V.; Elizade-González, M.P. Conotrachelusdimidiatus Champion, 1904 (Coleoptera: Curculionidae: Molytinae): Morphological re-description of the immature stages, keys, tribal comparisons and biology. Zootaxa 2018, 4433, 127–140. [Google Scholar] [CrossRef]
- Von Hormuzaki, C. Beiträge zur Käferfauna der Bukovina und Nordrumäliens. Entomol. Nachr. 1888, 14, 148–156. [Google Scholar]
- Sar, J.; Dudas, G. Bogarkozossegek (Coleoptera) vizsgalata pionir es rekultivalt elohelyen. Nat. Somogy. 2002, 3, 35–43. [Google Scholar]
- Stejskal, R.; Trnka, F. Nosatci tribu Cleonini a rodu Lixus v České republice (Coleoptera: Curculionidae: Lixinae) (Weevils of the tribe Cleonini and the genus Lixus in the Czech Republic (Coleoptera: Curculionidae: Lixinae)). Klapalekiana 2013, 49, 111–184, (In Czech, English summary). [Google Scholar]
- Fremuth, J. Cleoninae aus Türkei und den angrenzenden Gebieten (Coleoptera, Curculionidae). Fragm. Entomol. 1982, 16, 239–258. [Google Scholar]
- Ghahari, H.; Arzanov, Y.G.; Legalov, A.A.; Ostovan, H. Weevils (Coleoptera: Curculionidae) from Iranian rice fields and surrounding grasslands. Munis Entomol. Zool. 2010, 5, 163–169. [Google Scholar]
- Bouchard, P.; Lesage, L.; Goulet, H.; Bostanian, N.J.; Vincent, C.H.; Zmudzinska, A.; Lasnier, J. Weevil (Coleoptera: Curculionoidea) Diversity and Abundance in Two Quebec Vineyards. Ann. Entomol. Soc. Am. 2005, 98, 565–574. [Google Scholar] [CrossRef] [Green Version]
- Lindenberg, V.O. Do biologii lychynok dvokyljovogo dovgonosyka (Cleonus piger Scop.) [On the larval biology of the sluggish weevil (Cleonus piger Scop.)]. Naukovi Pratsi Instytutu Entomologii ta Fitopatologii 1950, 1, 96–100. [Google Scholar]
- MacNay, C.G. Highlights of the occurrence of insects and other arthropods in Canada, 1964. Coop. Econ. Insect Rep. 1965, 15, 21. [Google Scholar]
- Ioannisiani, T.G. Zhuki-dolgonosiki (Coleoptera, Curculionidae) Belorussii [Weevils of Belorussia (Coleoptera, Curculionidae)]; Nauka i Tekhnika Publ.: Minsk, Belarus, 1972; p. 352. [Google Scholar]
- Crosby, C.R.; Blauvelt, W.E. A European Beetle found in New York (Coleop.: Curculionidae). Entomol. News 1930, 41, 164. [Google Scholar]
- Aiping, L.; Linbo, X.; Hui, W. Biology and host range tests of Cleonus piger, a natural enemy of Cirsium arvense. Plant Prot. (Beijing) 2007, 33, 62–64, (In Chinese Cited by English Abstract). [Google Scholar]
- Isaev, A.Y. Ecologo-Faunistichesjkij Obzor Zhukov-Dolgonosikov [Coleoptera: Apionidae, Rhynchophoridae, Curculionidae] Ulyanovskoj Oblasti [An Ecologo-Faunistic Survey of Weevils [Coleoptera: Apionidae, Rhynchophoridae, Curculionidae] of Ulyanovsk Oblast; Branch of MSU: Ulyanosk, Russia, 1994; p. 104. [Google Scholar]
- Batra, S.W.T.; Coulson, J.R.; Dunn, P.H.; Boldt, P.E. Insects and fungi associated with Carduus thistles (Compositae). Technol. Bull. U.S. Dep. Agric. 1981, 1616, 100. [Google Scholar]
- Jolivet, P. Interrelationship between Insects and Plants; CRC Press: Boca Raton, FA, USA; London, UK; New York, NY, USA; Washington, DC, USA, 1998; p. 336. [Google Scholar]
- Forsyth, S.F.; Watson, A.K. Stress Inflicted by Organisms of Canada Thistle. In Proceedings of the 6th International Symposium Biological Control of Weeds, Vancouver, BC, Canada, 19–25 August 1984; Deflosse, E.S., Ed.; Agriculture Canada: Vancouver, BC, Canada, 1985; pp. 425–431. [Google Scholar]
- Sheppard, A.W.; Aeschlimann, J.P.; Sagliocco, J.L.; Vitou, J. Below-ground Herbivory in Carduus nutans (Asteraceae) and the Potential for Biological Control. Biocontrol. Sci. Technol. 1995, 5, 261–270. [Google Scholar] [CrossRef]
- Schädler, M.; Jung, G.; Brandl, R.; Auge, H. Secondary succession is influenced by belowground insect herbivory on a productive site. Oecologia 2004, 138, 242–252. [Google Scholar] [CrossRef]
- Watson, A.K.; Keogh, W.J. Mortality of Canada thistle due to Puccinia punctiformis. In Proceedings of the V International Symposium Biological Control of Weeds, Brisbane, Australia, 22–29 July 1980; CSIRO: Melbourne, Australia, 1980; pp. 325–332. [Google Scholar]
- Volovnik, S.V. On distribution and ecology of some species of Cleonine weevils (Coleoptera: Curculionidae) I. Tribe Cleonini. Entomol. Rev. 1989, 68, 138–144, [Original Russian text published in Entomol. Obozr., 68, 86–92]. [Google Scholar]
- Anderson, D.M. Notes on Cleonus piger (Scop.) in the United States (Coleoptera, Curculionidae). Coleopts. Bull. 1956, 1110, 81–85. [Google Scholar]
- Dieckmann, L. Beiträge zur Insektenfauna der DDR: Coleoptera—Curculionidae (Tanymecinae, Leptopiinae, Cleoninae, Tanyrhynchinae, Cossoninae, Raymondionyminae, Bagoinae, Tanysphyrinae). Beitr. Entomol. 1983, 33, 257–381. [Google Scholar]
- Müller, H.; Stinson, C.S.A.; Marquardt, K.; Schröeder, D. The entomofaunas of roots of Centaurea maculosa Lam., C. diffusa Lam., C. vallesiana Jordan in Europe. J. Appl. Entomol. 1989, 107, 83–95. [Google Scholar] [CrossRef]
- Müller, H. Structural Analysis of the Phytophagous Insect Guilds Associated with the Roots of Centaurea maculosa Lam., C. diffusa Lam., and C. vallesiana Jordan in Europe: 1. Field Observations. Oecologia 1989, 78, 41–52. [Google Scholar] [CrossRef] [PubMed]
- Nan, S.; Zhao, P.; Xin, H. Study on the occurring regular pattern and life history of the Cleonis piger. J. Heilongjiang Bayi Agric. Univ. 2000, 4. (In Chinese Cited by English Abstract). [Google Scholar]
- Pomerantsev, D.V.; Shevyrjov, I.Y. Znachenie nasekomojadnykh ptits v lesu i stepi [The importance of insectivorous birds in the forest and in the steppe]. Trudy po Lesnomu Opytnomu delu v Rossii 1910, 24, 1–99. [Google Scholar]
- Didier, R. L‘utilité de la Perdix Grise (Perdix perdix [L.]). Rev. Fr. D’ornithologie 1914, 3, 221–222. [Google Scholar]
- Antczak, M.; Konwerski, S.; Grobelny, S.; Tryjanowski, P. The Food Composition of Immature and Non-breeding White Storks in Poland. Waterbirds 2002, 25, 424–428. [Google Scholar] [CrossRef]
- Fahringer, J. Hymenopterologische Ergebnisse einer wissenschaftlichen Studienreise nach der Turkei und Kleinasien (mit Ausschluss des Amanusgebirges). Arch. Naturgesch. (A) 1922, 88, 149–222. [Google Scholar]
- Lou, S.X.; Li, Z.X. Preliminary observations on Cerceris rufipes evecta, a natural enemy of some weevils. For. Pest Dis. 1990, 2, 8, (In Chinese Cited by English Abstract). [Google Scholar]
- Volovnik, S.V. On parasites and predators of Cleoninae weevils (Col. Curculionidae) in Ukrainian steppe. Anz. Schadlingskde, Pflanzenschutz, Umweltschutz 1994, 67, 77–79. [Google Scholar] [CrossRef]
- Urban, S. Beauveria bassiana (BALS.) VUILL. (Fungi imperfecti) als Krankheitserreger bei einigen Russelkäferarten. Entomol. Nachr. 1967, 11, 93–96. [Google Scholar]
- Papp, J. A revision of the Bracon (Lucobracon) species described by Szepligeti from the western Palaearctic Region (Hymenoptera: Braconidae, Braconinae). Ann. Hist.-Nat. Musei Natl. Hung. 2005, 97, 197–224. [Google Scholar]
- Shaw, M.R.; Wahl, D.B. The biology, egg and larvae of Acaenitus dubitator (Panzer) (Hymenoptera, Ichneumonidae: Acaenitinae). Syst. Entomol. 1989, 14, 117–125. [Google Scholar] [CrossRef]
- Rotheray, G.E. Larval morphology and feeding patterns of four Cheilosia species (Diptera: Syrphidae) associated with Cirsium palustre L. Scopoli (Compositae) in Scotland. J. Nat. Hist. 1988, 22, 17–25. [Google Scholar] [CrossRef]
- Rizza, A.; Campobasso, G.; Dunn, P.H.; Stazi, M. Cheilosia corydon (Diptera: Syrphidae). A Candidate for the Biological Control of Musk Thistle in North America. Ann. Entomol. Soc. Am. 1988, 81, 225–232. [Google Scholar] [CrossRef]
- Ball, S.G.; Morris, R. Britain’s Hoverflies: An Introduction to the Hoverflies of Britain; Princeton University Press: Woodstock, UK, 2013; p. 297. [Google Scholar]
- Schröder, D. The Search for Effective Biological Control Agents in Europe. 1. Diffuse and Spotted Knapweed. In Proceedings of the VI International Symposium on Biological Control of Weeds, Vancouver, BC, Canada, 19–25 August 1984; Delfosse, E.S., Ed.; Agriculture of Canada: Ottawa, ON, Canada, 1985; pp. 103–119. [Google Scholar]
- Story, J.M.; Piper, G.L. Status of biological control efforts against spotted and diffuse knapweed. In Proceedings of the First International Knapweed Symposium of the Twenty-First Century, Coeur d’Alene, ID, USA, 15–16 March 2001; Smith, L., Ed.; pp. 11–17. [Google Scholar]
- Donald, W.W. The biology of Canada thistle (Cirsium arvense). Rev. Weed Sci. 1994, 6, 77–101. [Google Scholar]





{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| 1. | Endrocarina present. | 2 |
| - | Endrocarina absent. | 9 |
| 2. | Labrum with three lrms. | 3 |
| - | Labrum with two lrms. | Scaphomorphus erysimi |
| 3. | Setae on labrum (lrms) in a triangle position. | 4 |
| - | Setae on labrum (lrms) in a line. | 6 |
| 4. | Dorsum on abdominal segments I–VII with six or more pds. | |
| Ligula with three setae. Mandibula with two setae. | 5 | |
| - | Dorsum on abdominal segments I–VII with four pds. Ligula with two setae. Mandibula with one seta. | Asproparthenis carinicollis |
| 5. | Dorsum on abdominal segments I–VII with six pds (not in one line) and two ss (one not in line with pds). Dorsum on abdominal segment VIII with five pds and two ss. | Adosomus roridus |
| - | Dorsum on abdominal segments I–VII with seven pds and one ss (all in one line). Dorsum on abdominal segment VIII with five pds and one ss. | Cleonis pigra |
| 6. | Head with four des. | 7 |
| - | Head with five des. | 8 |
| 7. | Two des more than twice as long as the remaining two setae | Bothynoderes declivis |
| - | All four des of almost the same size | Bothynoderes affinis |
| 8. | Mandibula with one seta. Maxilla with one mbs. | Cyphocleonus achates |
| - | Mandibula with two setae. Maxilla without mbs. | Rhabdorrhynchus karelinii |
| 9. | Setae on labrum (lrms) in a triangle position. | 10 |
| - | Setae on labrum (lrms) in a line. | Pachycerus scabrosus |
| sensu Scherf (1964) | ||
| 10. | Mandibula with two setae. Epipharynx with four als. Ligula with two setae. | Cyphocleonus dealbatus |
| - | Mandibula with one seta. Epipharynx with six als. Ligula with three setae. | Coniocleonus nigrosuturatus |
| 1 | Mesonotum with five or six setae. Dorsum of abdominal segments I–VI with nine or ten setae (sometimes setae replaced by thorn-like asperities). | 2 |
| - | Mesonotum with three setae. Dorsum of abdominal segments I–VI with six or eight setae (sometimes setae replaced by thorn-like asperities). | 6 |
| 2. | Head with two or three sos. | 3 |
| - | Head with one sos. Mesonotum with five setae. Dorsum of abdominal segments I–VI with ten setae. | Rhabdorrhynchus karelinii |
| 3 | Mesonotum with six setae. | 4 |
| -. | Mesonotum with five setae. Rostrum with four rs. Dorsum of abdominal segments I–VI with nine setae. Each apex of femora with two fes. | Cleonis pigra |
| 4 | Rostrum with one rs. Dorsum of abdominal segments I–VI with nine setae. | 5 |
| - | Rostrum with two rs. Dorsum of abdominal segments I–VI with ten setae. Each apex of femora with two fes. | Adosomus roridus |
| 5. | Pronotum with ten setae. Head with four pas. Each apex of femora with three fes. | Coniocleonus nigrosuturatus |
| - | Pronotum with eleven setae. Head with five pas. | Scaphomorphuserysimi |
| 6 | Head with two sos. Metanotum with three setae. | 7 |
| -. | Head with one sos. Metanotum with four setae. | Bothynoderes affinis |
| 7. | Dorsum of abdominal segments I–VI with six setae. | Asproparthenis carinicollis |
| - | Dorsum of abdominal segments I–VI with eight setae. | Cyphocleonus achates |
© 2019 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Skuhrovec, J.; Volovnik, S.; Gosik, R.; Stejskal, R.; Trnka, F. Cleonis pigra (Scopoli, 1763) (Coleoptera: Curculionidae: Lixinae): Morphological Re-Description of the Immature Stages, Keys, Tribal Comparisons and Biology. Insects 2019, 10, 325. https://doi.org/10.3390/insects10100325
Skuhrovec J, Volovnik S, Gosik R, Stejskal R, Trnka F. Cleonis pigra (Scopoli, 1763) (Coleoptera: Curculionidae: Lixinae): Morphological Re-Description of the Immature Stages, Keys, Tribal Comparisons and Biology. Insects. 2019; 10(10):325. https://doi.org/10.3390/insects10100325
Chicago/Turabian StyleSkuhrovec, Jiří, Semyon Volovnik, Rafał Gosik, Robert Stejskal, and Filip Trnka. 2019. "Cleonis pigra (Scopoli, 1763) (Coleoptera: Curculionidae: Lixinae): Morphological Re-Description of the Immature Stages, Keys, Tribal Comparisons and Biology" Insects 10, no. 10: 325. https://doi.org/10.3390/insects10100325
APA StyleSkuhrovec, J., Volovnik, S., Gosik, R., Stejskal, R., & Trnka, F. (2019). Cleonis pigra (Scopoli, 1763) (Coleoptera: Curculionidae: Lixinae): Morphological Re-Description of the Immature Stages, Keys, Tribal Comparisons and Biology. Insects, 10(10), 325. https://doi.org/10.3390/insects10100325