Influence of Distance from the Host on Parasitisation by Psyttalia concolor (Szépligeti) (Hymenoptera: Braconidae)

 , , and
, , and 
Abstract
:1. Introduction
2. Materials and Methods
2.1. Insects
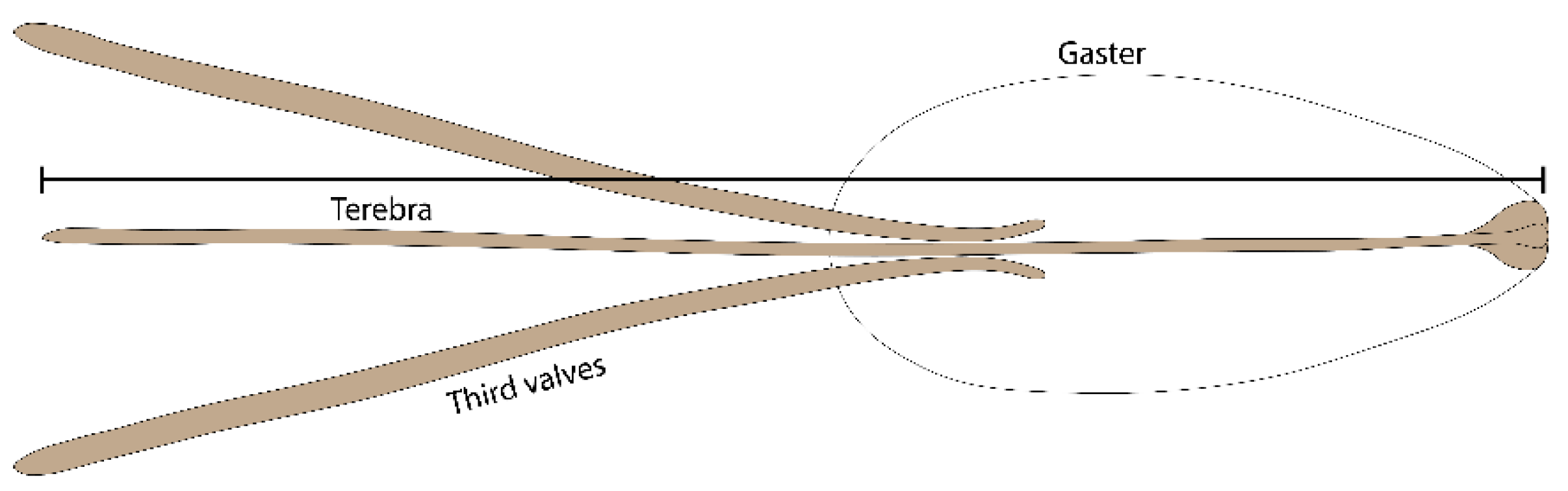
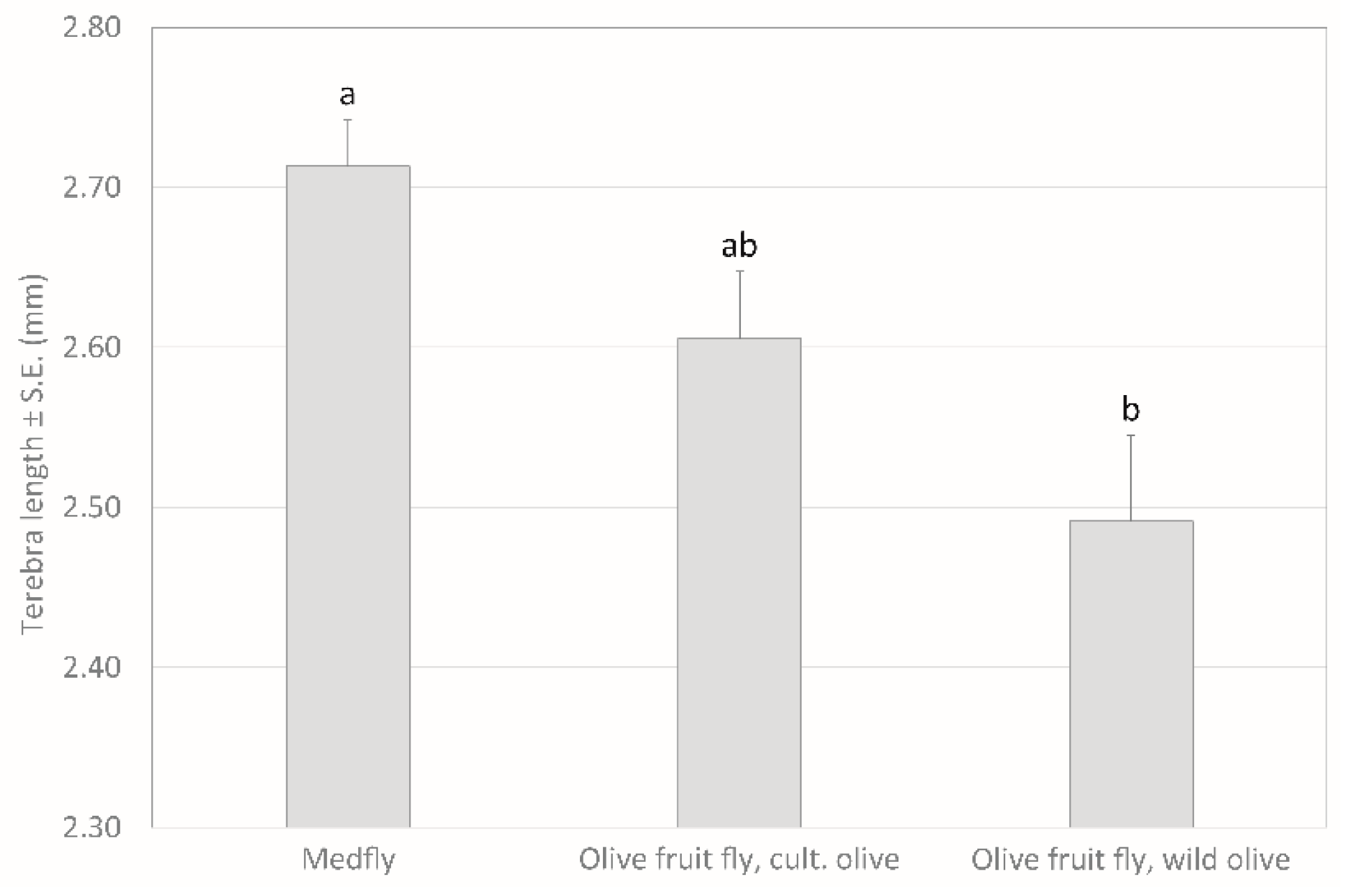
2.2. Ovipositor Length of Psyttalia concolor
2.3. Parasitisation Assays
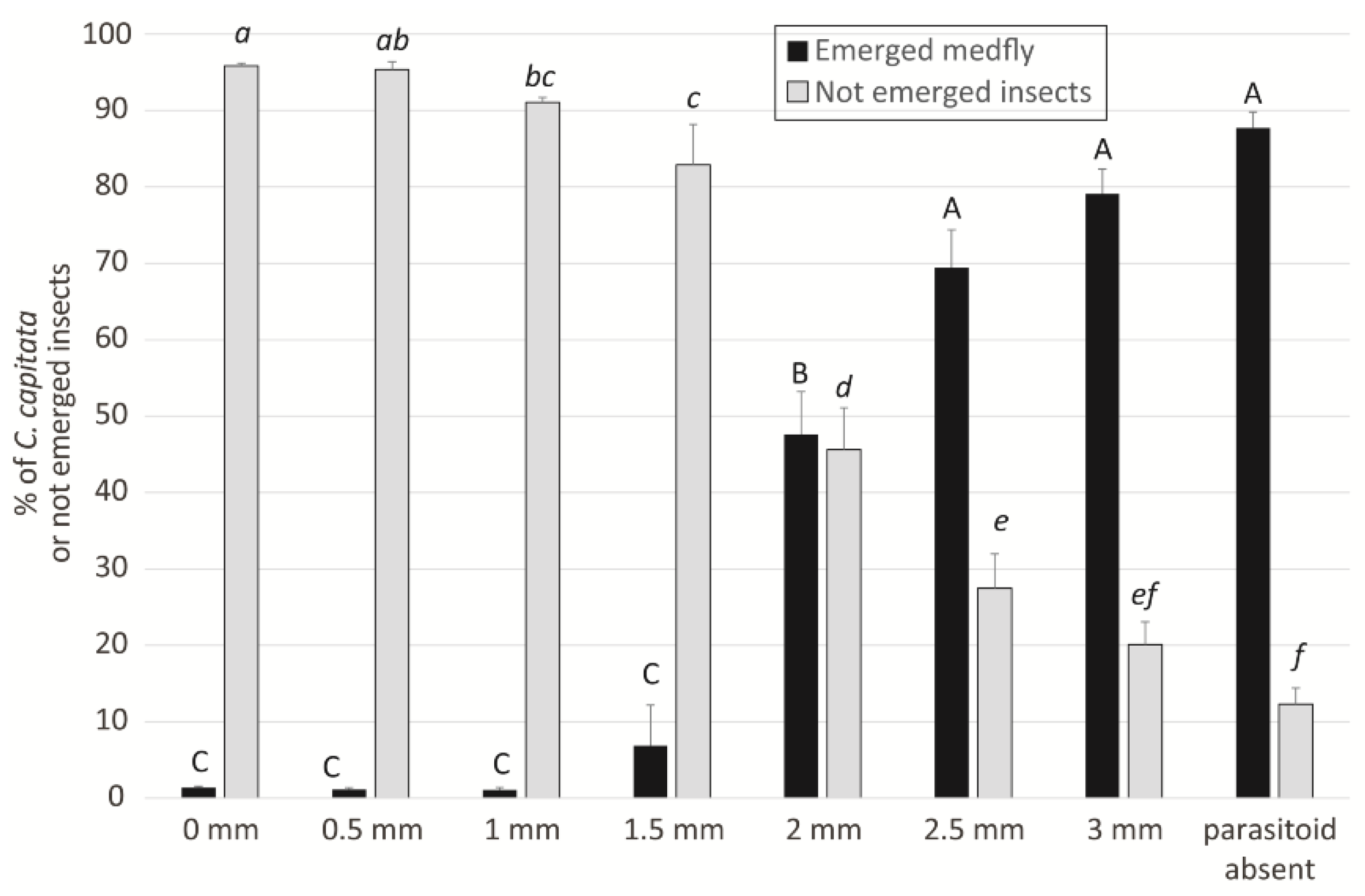
2.4. Parasitisation Tests on Ogliarola Barese Olives
2.5. Data Analysis
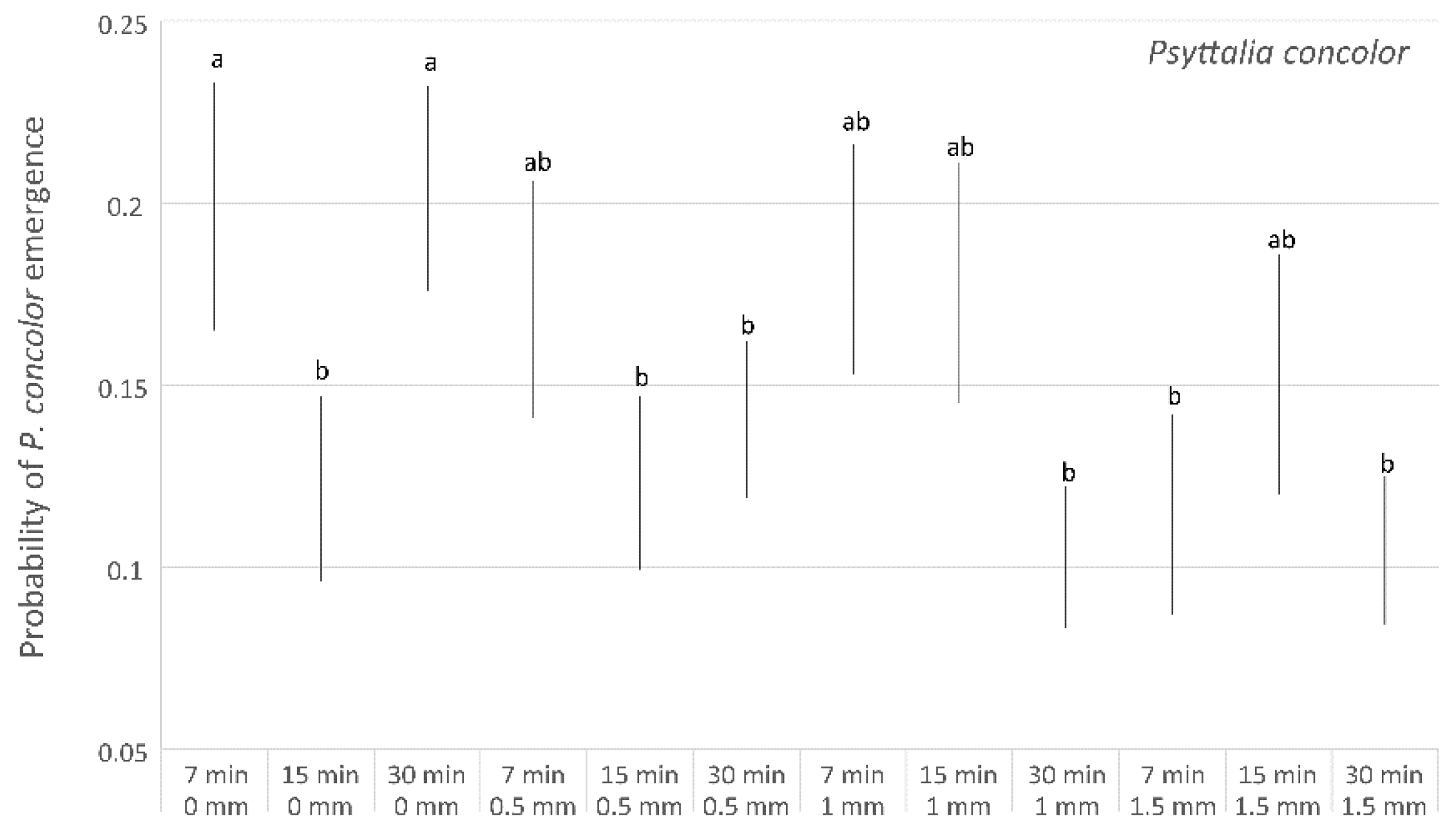
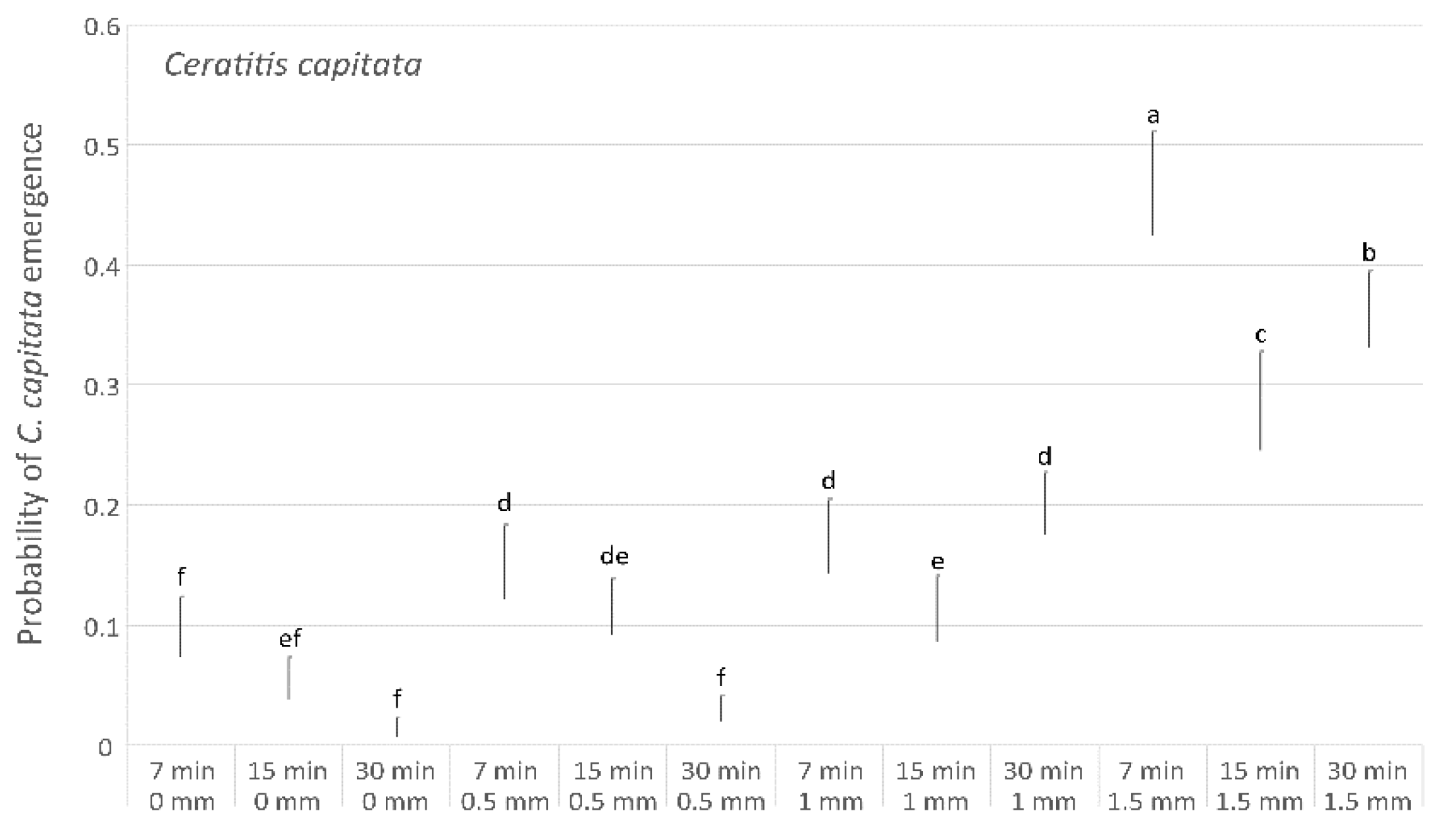
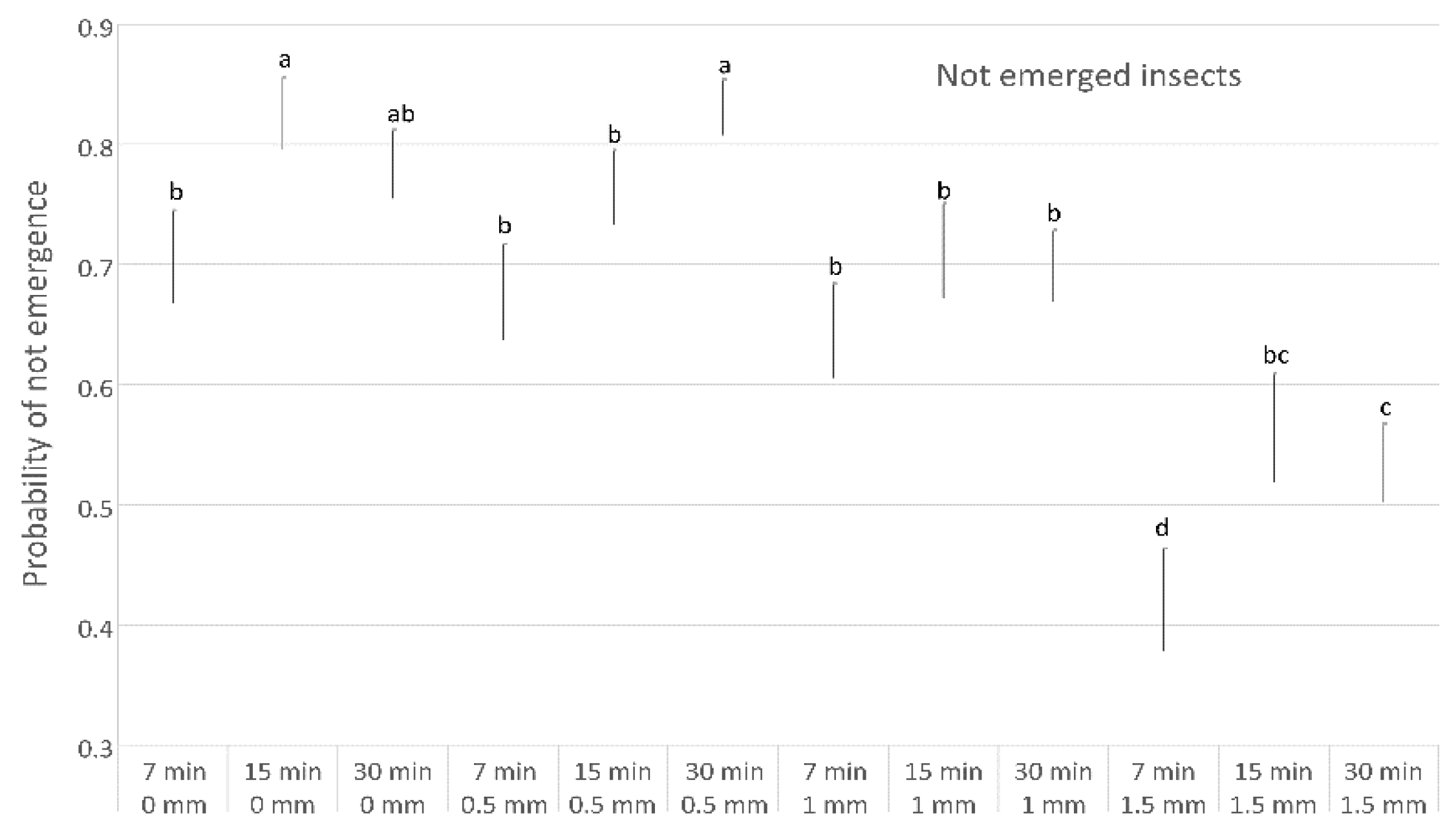
3. Results
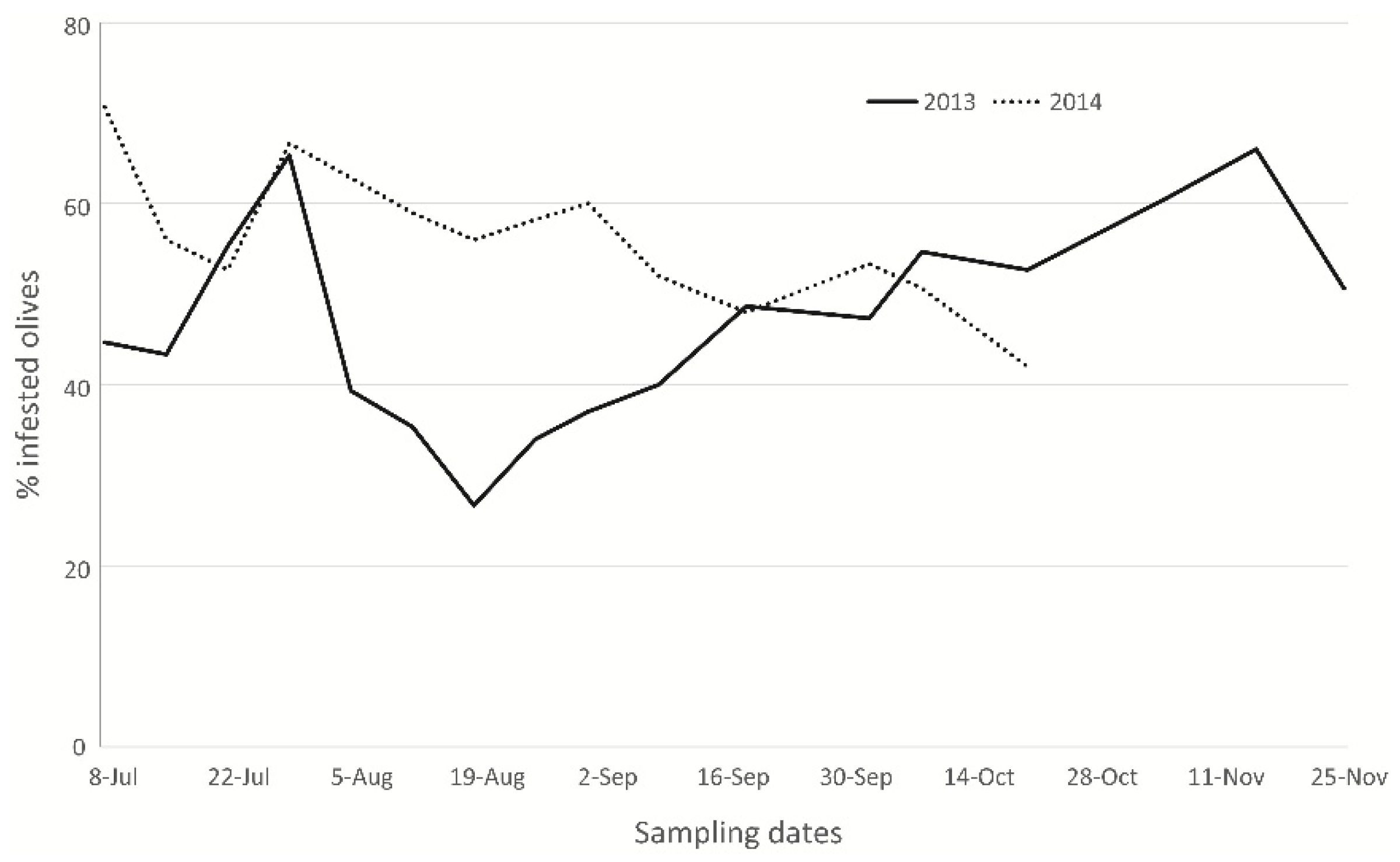
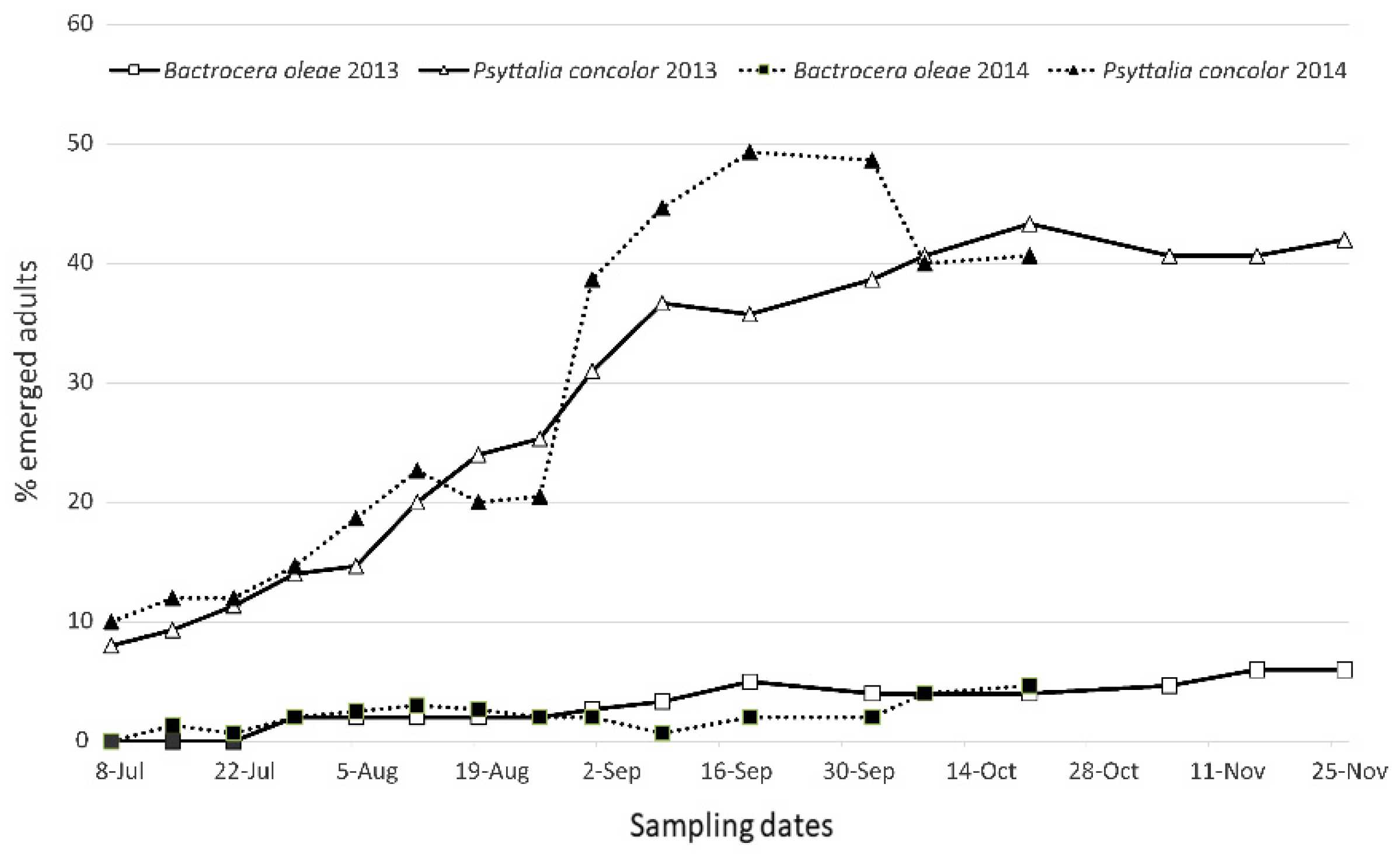
Parasitisation Tests on Ogliarola Barese Olive
4. Discussion
5. Conclusions
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
References
- Montiel Bueno, A.; Jones, O.T. Alternative methods for controlling the olive fly Bactrocera olea involving semiochemicals. IOBC WPRS Bull. 2002, 25, 147–155. [Google Scholar]
- Caleca, V.; Gıacalone, C.; Maltese, M.; Tortorıcı, F. Contenimento naturale di Bactrocera oleae (Rossi): Clima o parassitoidi? Confronto tra Western Cape (Sud Africa) e Sicilia. Atti Acc. Naz. Ital. Entomol. 2017, 64, 99–105. [Google Scholar]
- Skouras, P.J.; Margaritopoulos, J.T.; Seraphides, N.A.; Ioannides, I.M.; Kakani, E.G.; Mathiopoulos, K.D.; Atsitsipis, J. Organophosphate resistance in olive fruit fly, Bactrocera oleae, populations in Greece and Cyprus. Pest Manag. Sci. 2007, 63, 42–48. [Google Scholar] [CrossRef] [PubMed]
- Margaritopoulos, J.T.; Skavdis, G.; Kalogiannis, N.; Nikou, D.; Morou, E.; Skouras, P.J.; Tsitsipis, J.A.; Vontas, J. Efficacy of the pyrethroid alpha-cypermethrin against Bactrocera oleae populations from Greece, and improved diagnostic for an iAChE mutation. Pest Manag. Sci. 2008, 64, 900–908. [Google Scholar] [CrossRef] [PubMed]
- Kakani, E.G.; Zygouridis, N.E.; Tsoumani, K.T.; Seraphides, N.; Zalom, F.G.; Mathiopoulos, K.D. Spinosad resistance development in wild olive fruit fly Bactrocera oleae (Diptera: Tephritidae) populations in California. Pest Manag. Sci. 2010, 66, 447–453. [Google Scholar]
- Tzanakakis, M.E. Insects and Mites Feeding on Olive: Distribution, İmportance, Habits, Seasonal Development and Dormancy; Brill Academic Publisher: Leiden, The Netherlands, 2006; p. 198. [Google Scholar]
- Daane, K.M.; Johnson, M.W. Olive Fruit Fly: Managing an Ancient Pest in Modern Times. Annu. Rev. Entomol. 2010, 55, 151–169. [Google Scholar] [CrossRef] [PubMed]
- Dinis, A.M.; Pereira, J.A.; Pimenta, M.C.; Oliveira, J.; Benhadi-Marín, J.; Santos, S.A.P. Suppression of Bactrocera oleae (Diptera: Tephritidae) pupae by soil arthropods in the olive grove. J. Appl. Entomol. 2016, 140, 677–687. [Google Scholar] [CrossRef]
- Monastero, S.; Genduso, P. Esperimenti di lotta biologica artificiale contro la mosca delle olive (Dacus oleae Gmel.) eseguiti nel 1964 nell’isola di Salina (Eolie). Boll. Ist. Entomol. Agr. Oss. Fitopat. Palermo 1964, 5, 281–289. [Google Scholar]
- Monastero, S.; Delanoue, P. Lutte biologique expérimentale contre la mouche de l’olive (Dacus oleae Gmel.) au moyen d’Opius concolor Szepl. siculus Mon. dans les îles Éoliennes (Sicile) en 1965. Entomophaga 1966, 11, 411–432. [Google Scholar] [CrossRef]
- Monastero, S. La prima grande applicazione di lotta biologica artificiale contro la mosca delle olive (Dacus oleae Gmel.) (32 milioni di Opius per difendere 300 mila piante di olivo). Boll. Ist. Entomol. Agr. Oss. Fitopat. Palermo 1967, 7, 63–100. [Google Scholar]
- Liotta, G.; Mineo, G. Lotta biologica artificiale contro la mosca delle olive a mezzo dell’Opius concolor siculus in Sicilia nel 1968. Boll. Ist. Entomol. Agr. Oss. Fitopat. Palermo 1969, 7, 183–196. [Google Scholar]
- Genduso, P.; Ragusa, S. Lotta biologica artificiale contro la mosca delle olive a mezzo dell’Opius c. siculus in Puglia nel 1968. Boll. Ist. Entomol. Agr. Oss. Fitopat. Palermo 1969, 7, 197–216. [Google Scholar]
- Kapatos, E.; Fletcher, B.S.; Pappas, S.; Laudeho, Y. The release of Opius concolor and O. concolor var. siculus [Hym.: Braconidae] against the spring generation of Dacus oleae [Dipt.: Trypetidae] on Corfu. Entomophaga 1977, 22, 265–270. [Google Scholar] [CrossRef]
- Liaropoulos, C.; Louskas, C.; Canard, M.; Laudeho, Y. Release of Opius concolor (Hym.: Braconidae) on spring populations of Dacus oleae (Dipt.: Trypetidae): 1. Experiments in continental Greece. Entomophaga 1977, 22, 259–264. [Google Scholar] [CrossRef]
- Michelakis, S.E. Parasitoids for the control of Dacus oleae in Crete. EPPO Bull. 1986, 16, 389–393. [Google Scholar] [CrossRef]
- Jimenez, A. Potential value of entomophagous in the olive pests control. In Proceedings of the CEC/FAO/IOBC International Joint Meeting Integrated Pest Control in Olive-groves, Pisa, Italy, 3–6 April 1984; Cavalloro, R., Crovetti, A., Eds.; August Aimé Balkema Publisher: Rotterdam, The Netherlands, 1985; pp. 441–450. [Google Scholar]
- Alexandrakis, V.Z. Use of entomophagous insects in controlling olive pests. Bull. OEPP 1986, 16, 375–381. [Google Scholar] [CrossRef]
- Jimenez, A.; Castillo, E.; Lorite, P. Supervivencia del himenòptero bracònido Opius concolor Szep. parasito de Dacus oleae Gmelin en olivares de Jaén. Bol. San. Plagas 1990, 16, 97–103. [Google Scholar]
- Raspi, A.; Loni, A. Alcune note sull’allevamento massale di Opius concolor Szépligeti (Hym.: Braconidae) e su recenti tentative d’introduzione della specie in Toscana e Liguria. Frustula Entomol. 1994, 30, 135–145. [Google Scholar]
- Raspi, A. Lotta biologica in olivicoltura. In Proceedings of the Atti del convegno su Tecniche, norme e qualità in Olivicoltura, Potenza, Italy, 15–17 December 1993; pp. 483–495. [Google Scholar]
- Abdel-Magid, J.E.; Hammad, K.A. Seasonal abundance of Bactrocera (Dacus) oleae Gmelin and its parasitoid, Opius concolor (Hymenoptera: Braconidae) in El-Khattara district, Sharkia Governorate, Egypt. Zagazig J. Agric. Res. 2000, 26, 163–175. [Google Scholar]
- Delrio, G.; Lentini, A.; Satta, A. Biological control of olive fruit fly through inoculative releases of Opius concolor Szepl. IOBC WPRS Bull. 2005, 30, 53–58. [Google Scholar]
- Yokoyama, V.Y.; Rendón, P.A.; Wang, X.-G.; Opp, S.B.; Johnson, M.W.; Daane, K.M. Response of Psyttalia humilis (Hymenoptera: Braconidae) to Olive Fruit Fly (Diptera: Tephritidae) and Conditions in California Olive Orchards. Environ. Entomol. 2011, 40, 315–323. [Google Scholar] [CrossRef]
- Daane, K.M.; Wang, X.; Nieto, D.J.; Pickett, C.H.; Hoelmer, K.A.; Blanchet, A.; Johnson, M.W. Classic biological control of olive fruit fly in California, USA: Release and recovery of introduced parasitoids. BioControl 2015, 60, 317–330. [Google Scholar] [CrossRef]
- Rugman-Jones, P.F.; Wharton, R.; van Noort, T.; Stouthamer, R. Molecular differentiation of the Psyttalia concolor (Szépligeti) species complex (Hymenoptera: Braconidae) associated with olive fly, Bactrocera oleae (Rossi) (Diptera: Tephritidae), in Africa. Biol. Control 2009, 49, 17–26. [Google Scholar] [CrossRef]
- Delucchi, V. Les parasites de la mouche del olives. Entomophaga 1957, 2, 107–118. [Google Scholar] [CrossRef]
- Monastero, S. Un nuovo parassita endofago della mosca delle olive trovato in Altavilla Milicia (Sicilia) (Fam. Braconidae Gen. Opius). Atti R. Accade. Sci. Let. Arti Palermo 1931, 16, 1–7. [Google Scholar]
- Silvestri, F. La lotta biologica contro le mosche dei frutti della famiglia Trypetidae. In Proceedings of the VII International Congress of Entomology, Berlin, Germany, 15–20 August 1938; pp. 2396–2418. [Google Scholar]
- Gıacalone, C. Il Controllo di Bactrocera oleae (Rossi) e di Altri Carpofagi Negli Oliveti Biologici in Sicilia e Sud Africa. Ph.D. Thesis, Dipartimento DEMETRA, Università Degli Studi di Palermo, Palermo, Italy, 2011; p. 147. [Google Scholar]
- Canale, A. Effect of parasitoid/host ratio on superparasitism of Ceratitis capitata (Wiedemann) larvae (Diptera: Tephritidae) by Opius concolor Szépligeti (Hymenoptera: Braconidae). Frustula Entomol. 1998, 21, 137–148. [Google Scholar]
- Loni, A. Impact of host exposure time on mass-rearing of Psyttalia concolor (Hymenoptera Braconidae) on Ceratitis capitata (Diptera Tephritidae). Bull. Insectol. 2003, 56, 277–282. [Google Scholar]
- Canale, A. Studi Bio-Etologici su parassitoidi di Ditteri Tefritidi di İnteresse Economico: Il Superparassitismo in Opius concolor Szépligeti. (Hymenoptera Braconidae). Ph.D. Thesis, Dipartimento di Coltivazione e Difesa delle Specie legnose "G. Scaramuzzi", Universita Degli Studi di Pisa, Pisa, Italy, 1999; p. 94. [Google Scholar]
- Raspi, A.; Canale, A. Effect of superparasitism on Ceratitis capitata (Wiedemann) (Diptera Tephritidae) second instar larvae by Psyttalia concolor (Szepligeti) (Hymenoptera Braconidae). Redia 2000, 83, 123–131. [Google Scholar]
- Benelli, G.; Gennari, G.; Canale, A. Host discrimination ability in the tephritid parasitoid Psyttalia concolor (Hymenoptera: Braconidae). J. Pest Sci. 2013, 86, 245–251. [Google Scholar] [CrossRef]
- Giunti, G.; Benelli, G.; Conte, G.; Mele, M.; Caruso, G.; Gucci, R.; Flamini, G.; Canale, A. VOCs-Mediated Location of Olive Fly Larvae by the Braconid Parasitoid Psyttalia concolor: A Multivariate Comparison among VOC Bouquets from Three Olive Cultivars. BioMed Res. Int. 2016, 2016, 7827615. [Google Scholar] [CrossRef]
- Giunti, G.; Benelli, G.; Messing, R.H.; Canale, A. Early adult learning affects host preferences in the tephritid parasitoid Psyttalia concolor (Hymenoptera: Braconidae). J. Pest Sci. 2016, 89, 529–537. [Google Scholar] [CrossRef]
- Yokoyama, V.Y.; Rendón, P.A.; Sivinski, J. Psyttalia cf. concolor (Hymenoptera: Braconidae) for Biological Control of Olive Fruit Fly (Diptera: Tephritidae) in California. Environ. Entomol. 2008, 37, 764–773. [Google Scholar] [PubMed]
- Daane, K.M.; Sime, K.R.; Wang, X.G.; Nadel, H.; Johnson, M.W.; Walton, V.M.; Kirk, A.; Pickett, C.H. Psyttalia lounsburyi (Hymenoptera: Braconidae), potential biological control agent for the olive fruit fly in California. Biol. Control 2008, 44, 79–89. [Google Scholar] [CrossRef]
- Blanchet, A.; Hurtrel, B.; Roche, M.; Kirk, A.A.; Jones, W.A. A novel rearing technique for the olive fruit fly parasitoid Psyttalia lounsburyi (Hymenoptera: Braconidae) on Ceratitis capitata (Diptera: Tephritidae) in artificial diet. IOBC WPRS Bull. 2009, 59, 161. [Google Scholar]
- La-Spina, M.; Pickett, C.; Daane, K.M.; Hoelmer, K.; Blanchet, A.; Williams, L. Effect of exposure time on mass-rearing production of the olive fruit fly parasitoid, Psyttalia lounsburyi (Hymenoptera: Braconidae). J. Appl. Entomol. 2018, 142, 319–326. [Google Scholar] [CrossRef]
- Garantonakis, N.; Varikou, K.; Birouraki, A. Parasitism of Psytallia concolor (Hymenoptera: Braconidae) on Bactrocera oleae (Diptera: Tephritidae) infesting different olive varieties. Phytoparasitica 2017, 45, 461–469. [Google Scholar] [CrossRef]
- Wang, X.G.; Johnson, M.W.; Daane, K.M.; Yokoyama, V.Y. Larger olive fruit size reduces the efficiency of Psyttalia concolor, as a parasitoid of the olive fruit fly. Biol. Control 2009, 49, 45–51. [Google Scholar] [CrossRef]
- Wang, X.G.; Nadel, H.; Johnson, M.W.; Daane, K.M.; Hoelmer, K.; Walton, V.M.; Pickett, C.H.; Sime, K.R. Crop domestication relaxes both top-down and bottom-up effects on a specialist herbivore. Basic Appl. Ecol. 2009, 10, 216–227. [Google Scholar] [CrossRef]
- Loni, A. Developmental rate of Opius concolor (Hym.: Braconidae) at various constant temperatures. Entomophaga 1997, 42, 359–366. [Google Scholar] [CrossRef]
- Genduso, P. Attuale tecnica di allevamento dell’Opius concolor Szépl. siculus Mon. Boll. Ist. Ent. Agr. Fitopat. Palermo 1969, 7, 9–40. [Google Scholar]
- Oreste, M.; Baser, N.; Bubici, G.; Tarasco, E. Effect of Beauveria bassiana strains on the Ceratitis capitata—Psyttalia concolor system. Bull. Insectol. 2015, 68, 265–272. [Google Scholar]
- Cavalloro, R.; Girolami, V. Miglioramenti nell’allevamento in massa di Ceratitis capitata Wiedemann (Diptera, Trypetidae). Redia 1969, 51, 315–327. [Google Scholar]
- International Olive Council. World Catalogue of Olive Varieties; IOC: Madrid, Spain, 2000; p. 360. [Google Scholar]
- Ueno, T. Effects of host size and laboratory rearing on offspring development and sex ratio in the solitary parasitoid Agrothereutes lanceolatus (Hymenoptera: Ichneumonidae). Eur. J. Entomol. 2015, 112, 281–287. [Google Scholar] [CrossRef]
- Salvo, A.; Valladares, G. Intraspecific size variation in polyphagous parasitoids (Hym.: Parasitica), of leaf miners and its relation to host size. Entomophaga 1995, 40, 273–280. [Google Scholar] [CrossRef]
- Burrack, H.J.; Zalom, F.G. Olive Fruit Fly (Diptera: Tephritidae) Ovipositional Preference and Larval Performance in Several Commercially Important Olive Varieties in California. J. Econ. Entomol. 2008, 101, 750–758. [Google Scholar] [CrossRef]
- Canale, A. Psyttalia concolor (Hymenoptera Braconidae): Role of host movement and host substrate in ovipositor-probing behaviour. Bull. Insectol. 2003, 56, 211–213. [Google Scholar]
- Canale, A.; Loni, A. Host location and acceptance in Psyttalia concolor: Role of host instar. Bull. Insectology 2006, 59, 7–10. [Google Scholar]











{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Psyttalia concolor | DF | Adj Dev | Adj Mean | Chi-Square | p-Value |
|---|---|---|---|---|---|
| Regression | 11 | 144.65 | 13.15 | 144.65 | <0.001 |
| Distance | 3 | 28.78 | 9.593 | 28.78 | <0.001 |
| Minutes | 2 | 35.14 | 17.571 | 35.14 | <0.001 |
| Distance ∗ Minutes | 6 | 81.55 | 13.592 | 81.55 | <0.001 |
| Error | 99 | 716.56 | 7.238 | ||
| Total | 110 | 861.21 | |||
| Ceratitis capitata | |||||
| Regression | 11 | 1767.29 | 160.663 | 1767.29 | <0.001 |
| Distance | 3 | 404.78 | 134.928 | 404.78 | <0.001 |
| Minutes | 2 | 88.64 | 44.318 | 88.64 | <0.001 |
| Distance ∗ Minutes | 6 | 222.67 | 37.112 | 222.67 | <0.001 |
| Error | 99 | 864.42 | 8.732 | ||
| Total | 110 | 2631.71 | |||
| Not emerged | |||||
| Regression | 11 | 881.37 | 80.125 | 881.37 | <0.001 |
| Distance | 3 | 190.46 | 63.486 | 190.46 | <0.001 |
| Minutes | 2 | 40.8 | 20.398 | 40.8 | <0.001 |
| Distance ∗ Minutes | 6 | 41.69 | 6.948 | 41.69 | <0.001 |
| Error | 99 | 496.25 | 5.013 | ||
| Total | 110 | 1377.62 |
© 2019 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Baser, N.; Lo Verde, G.; Kaçar, G.; Lamaj, F.; Verrastro, V.; Lombardo, A.; Tortorici, F.; Caleca, V. Influence of Distance from the Host on Parasitisation by Psyttalia concolor (Szépligeti) (Hymenoptera: Braconidae). Insects 2019, 10, 316. https://doi.org/10.3390/insects10100316
Baser N, Lo Verde G, Kaçar G, Lamaj F, Verrastro V, Lombardo A, Tortorici F, Caleca V. Influence of Distance from the Host on Parasitisation by Psyttalia concolor (Szépligeti) (Hymenoptera: Braconidae). Insects. 2019; 10(10):316. https://doi.org/10.3390/insects10100316
Chicago/Turabian StyleBaser, Nuray, Gabriella Lo Verde, Gülay Kaçar, Flutura Lamaj, Vincenzo Verrastro, Alberto Lombardo, Francesco Tortorici, and Virgilio Caleca. 2019. "Influence of Distance from the Host on Parasitisation by Psyttalia concolor (Szépligeti) (Hymenoptera: Braconidae)" Insects 10, no. 10: 316. https://doi.org/10.3390/insects10100316
APA StyleBaser, N., Lo Verde, G., Kaçar, G., Lamaj, F., Verrastro, V., Lombardo, A., Tortorici, F., & Caleca, V. (2019). Influence of Distance from the Host on Parasitisation by Psyttalia concolor (Szépligeti) (Hymenoptera: Braconidae). Insects, 10(10), 316. https://doi.org/10.3390/insects10100316