Comb Irradiation Has Limited, Interactive Effects on Colony Performance or Pathogens in Bees, Varroa destructor and Wax Based on Two Honey Bee Stocks



Abstract
1. Introduction
2. Materials and Methods
2.1. Establishing Test Colonies
2.2. Measuring Colony Performance and Mite Infestation
2.3. Pathogen Load Analyses
2.4. Data Analyses
3. Results
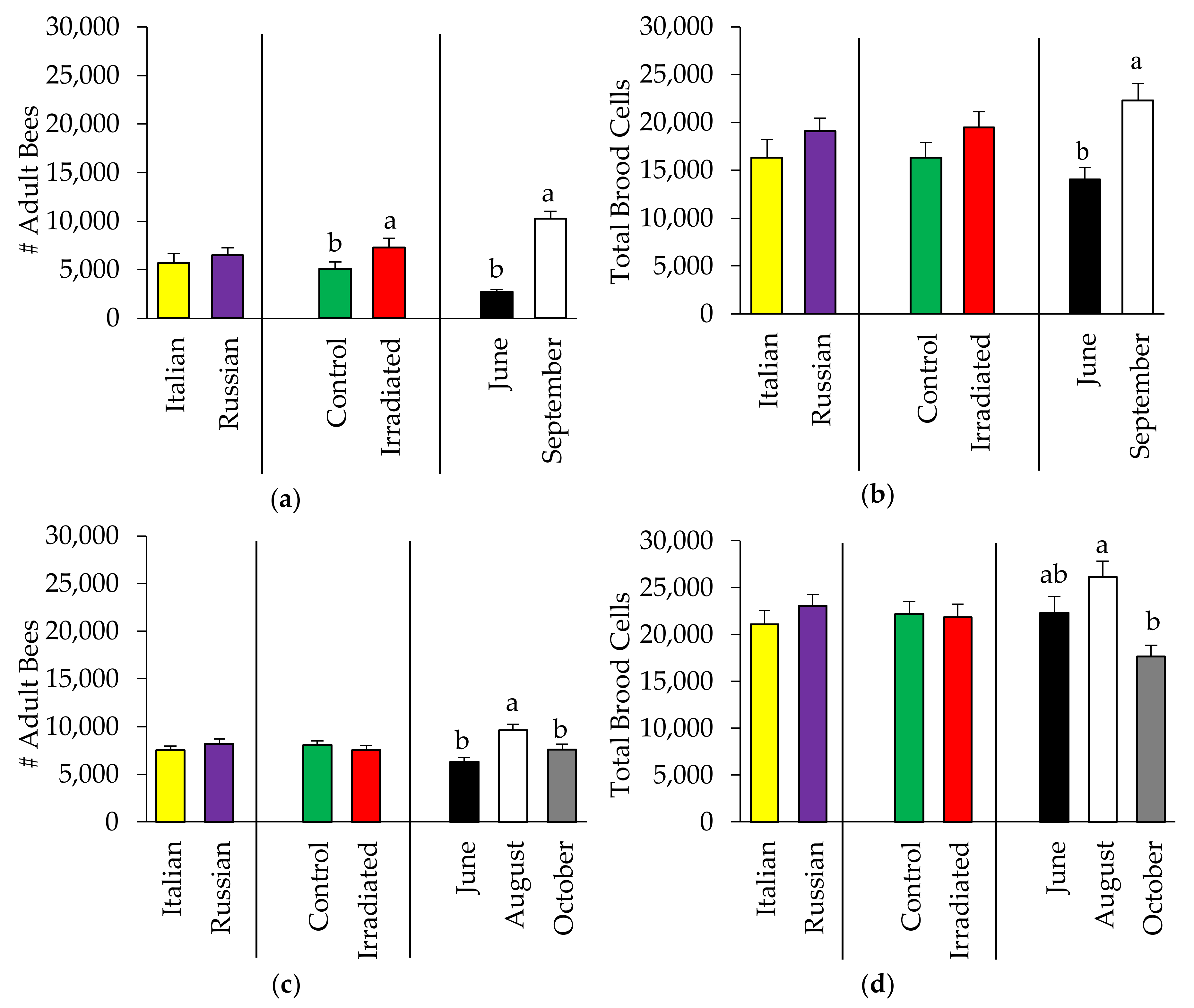
3.1. Effects of Honey Bee Stock and Comb Irradiation on Colony Performance
3.1.1. Bee Population
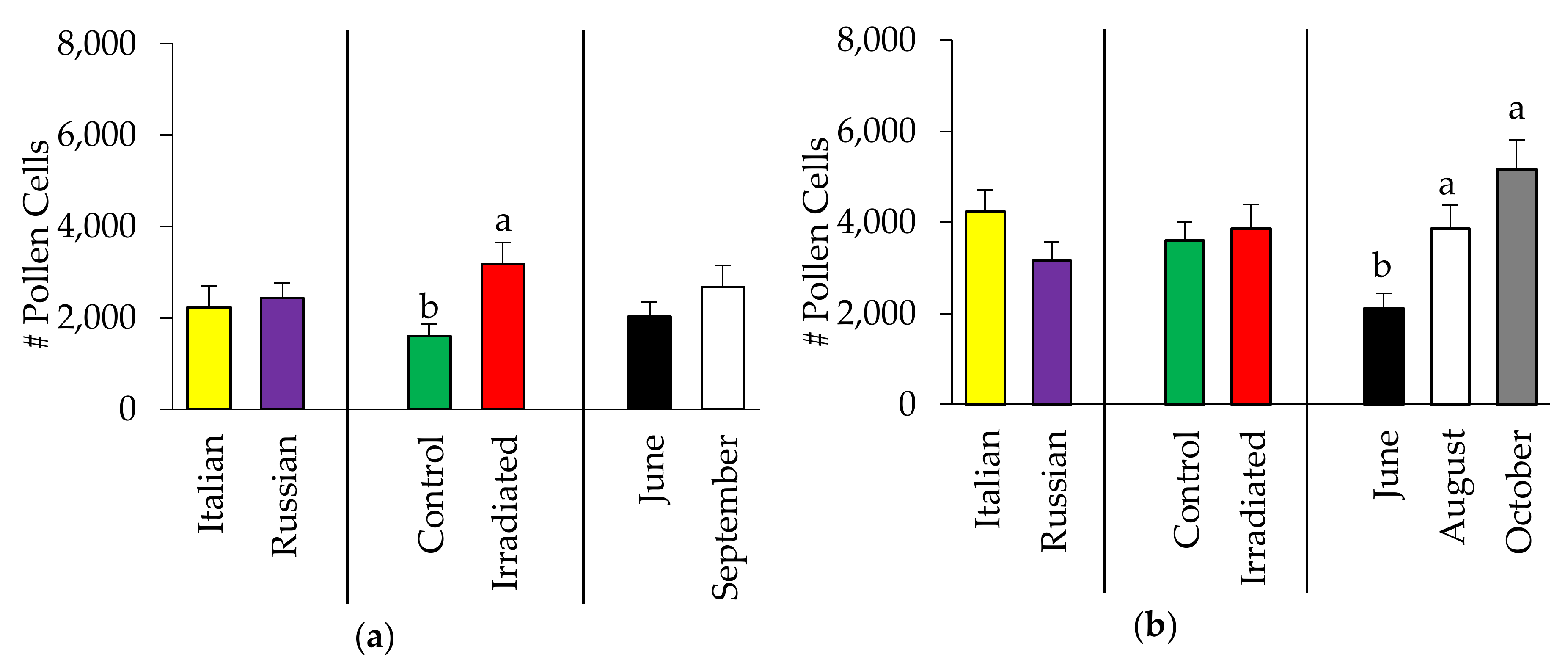
3.1.2. Pollen Availability
3.2. Effects of Honey Bee Stock and Comb Irradiation on Varroa Population
Colony Mites
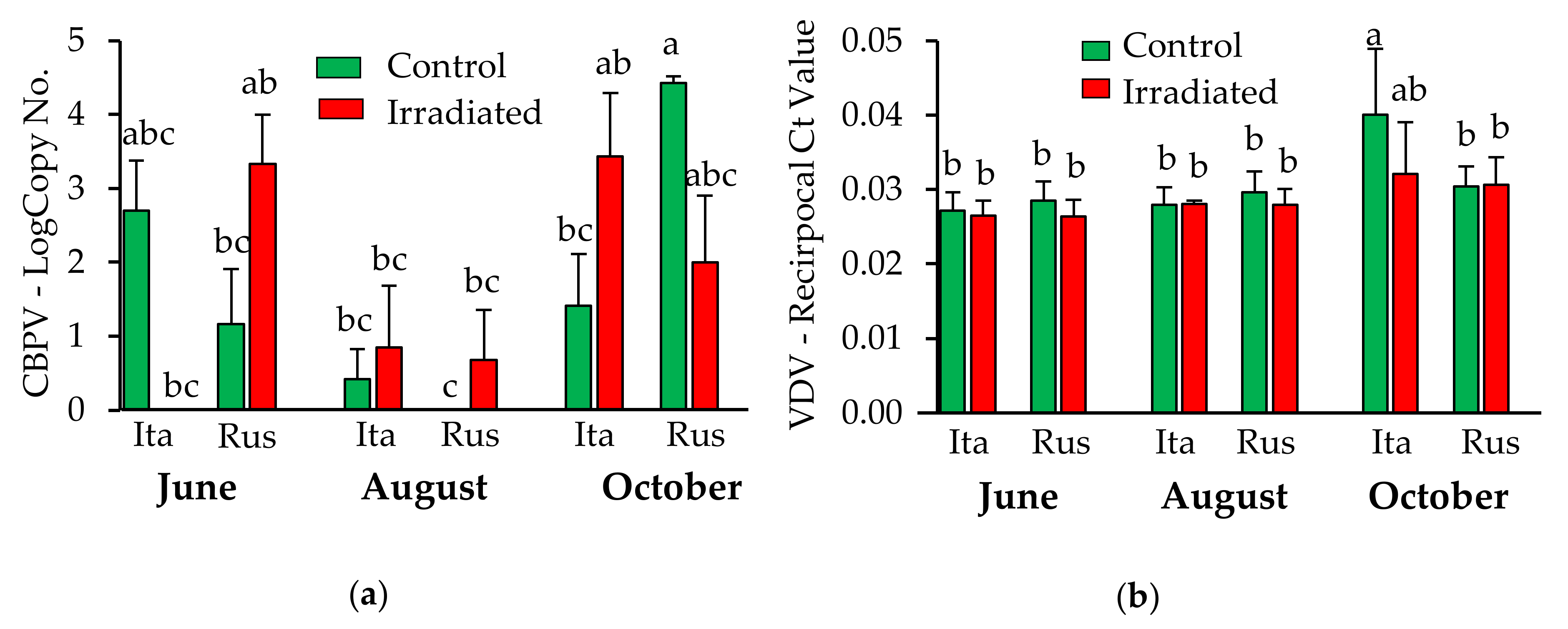
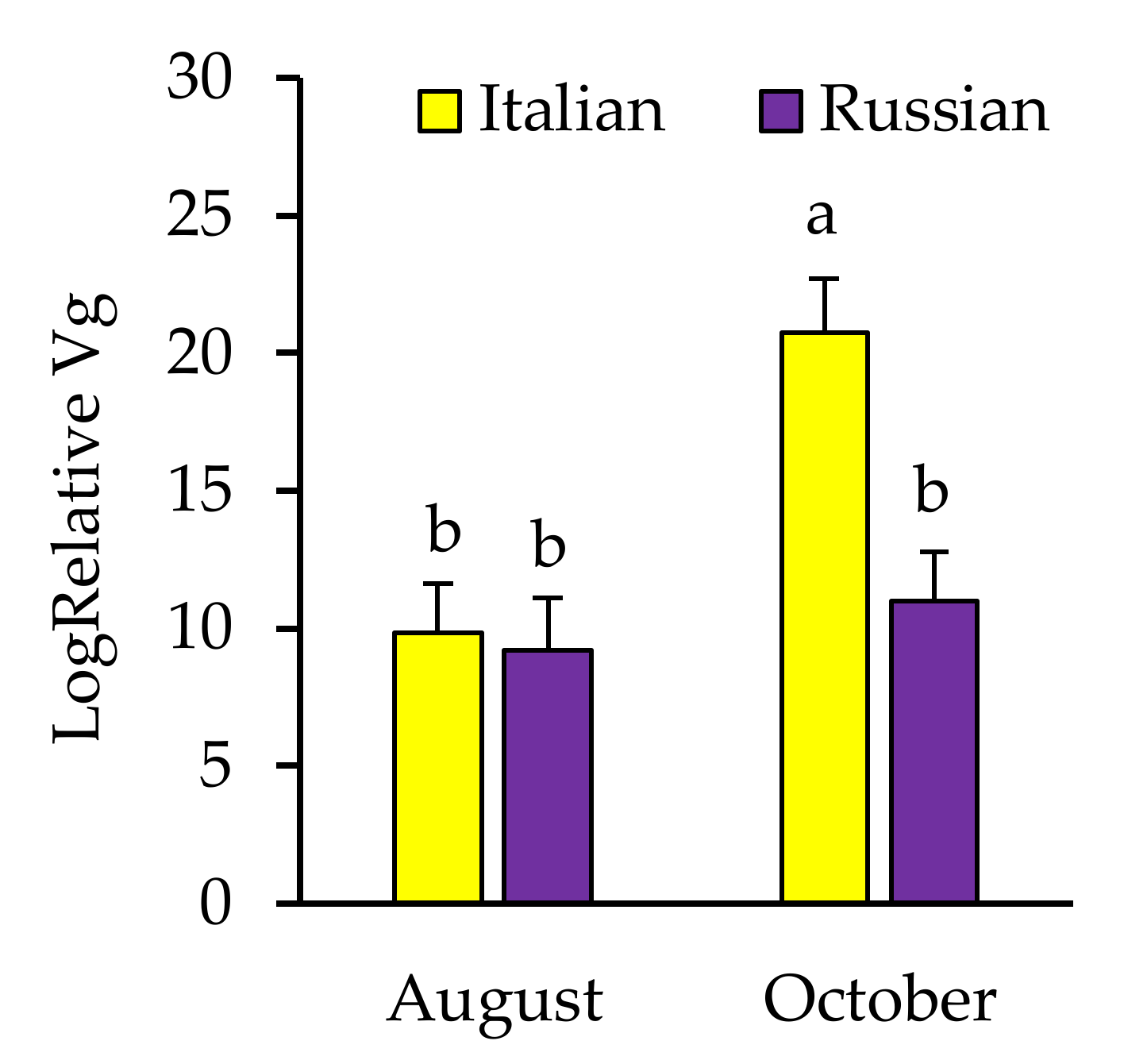
3.3. Effects of Honey Bee Stock and Comb Irradiation on Pathogen Load
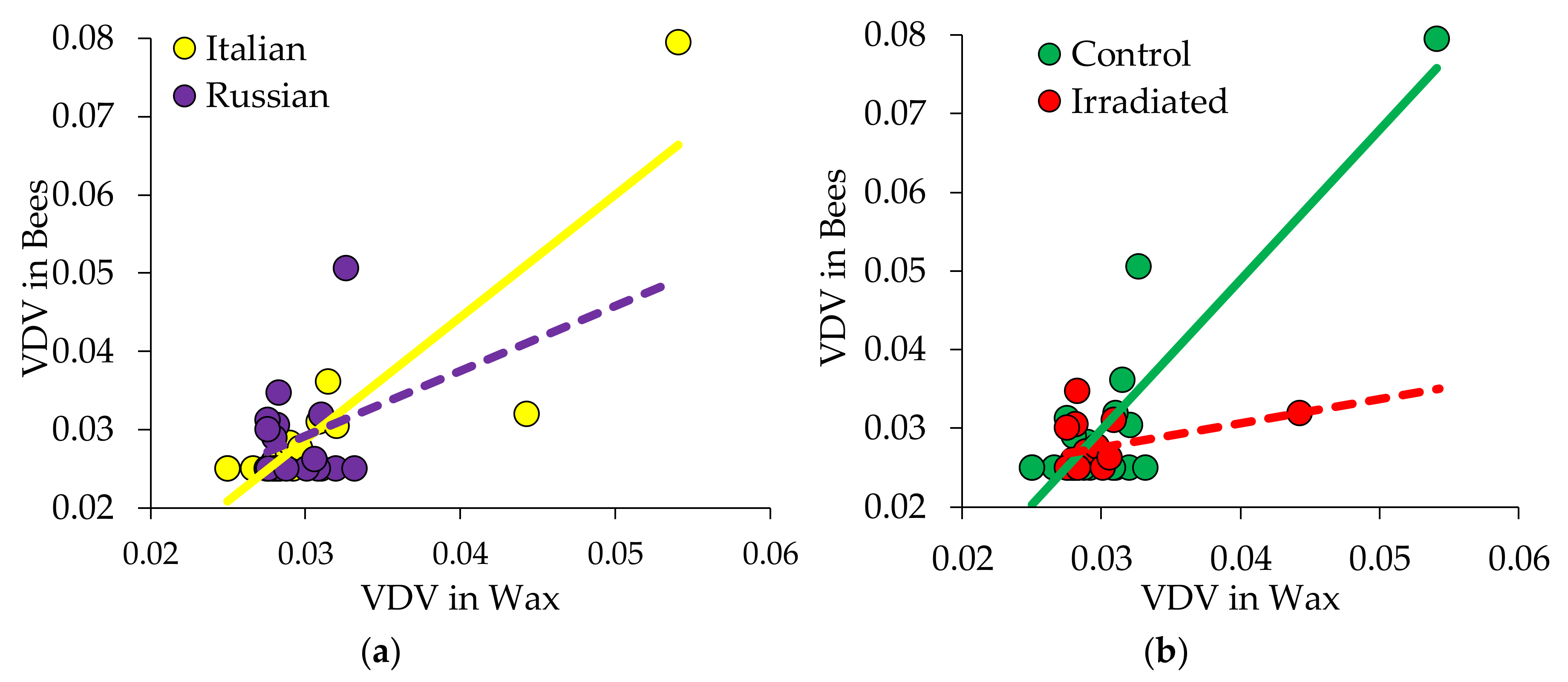
3.3.1. Pathogens Detected in Wax
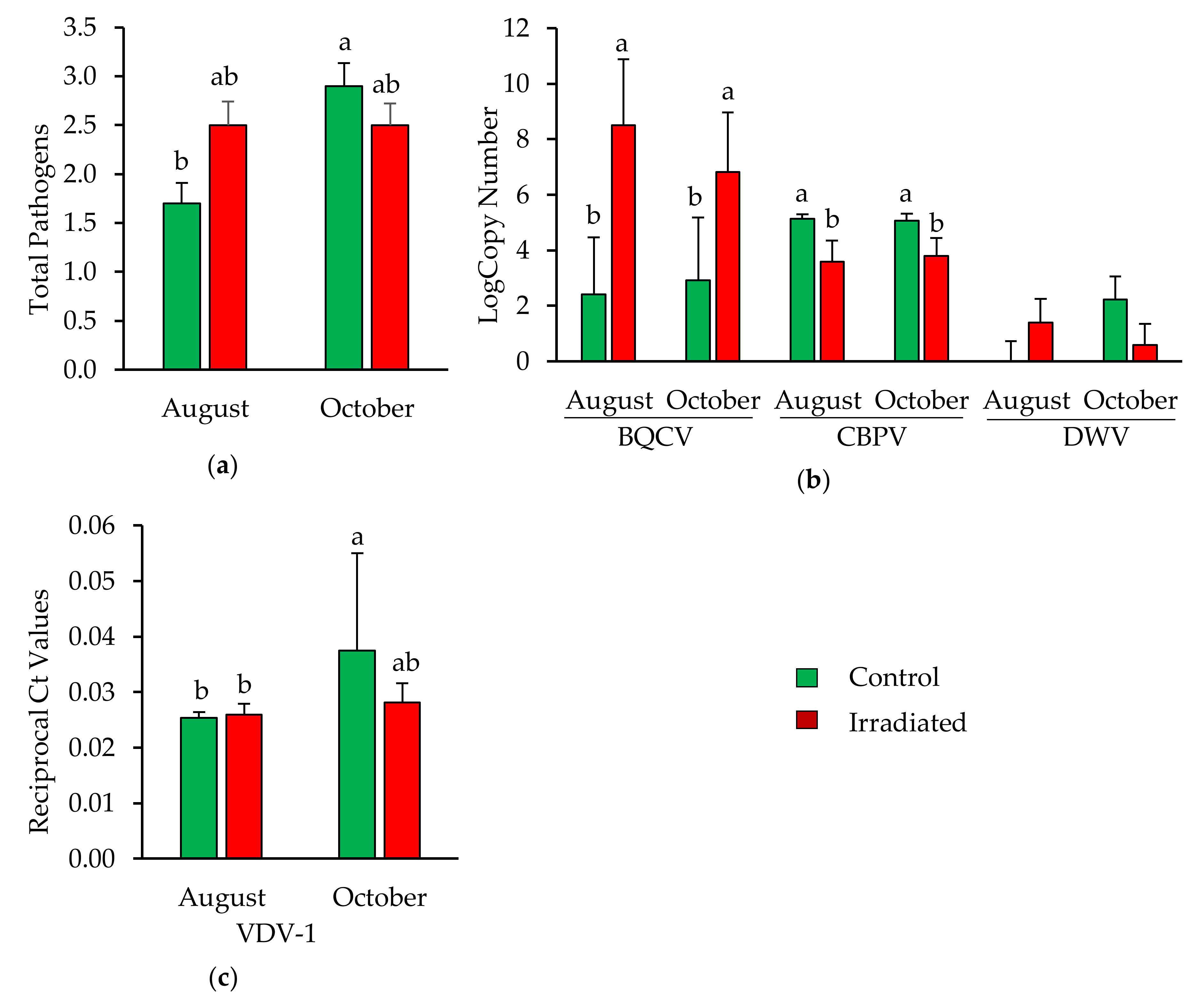
3.3.2. Pathogens Detected in Newly Emerged Bees
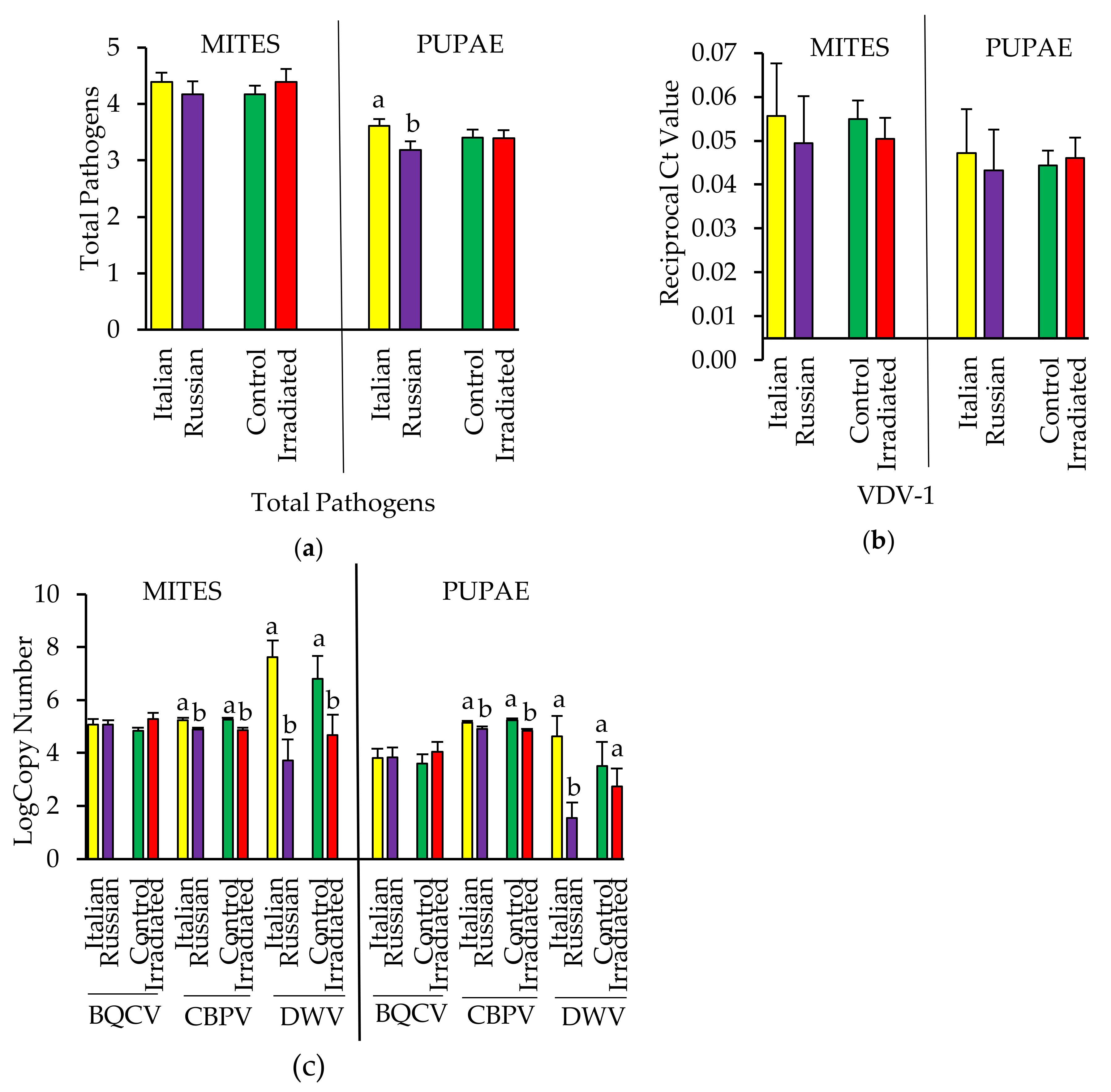
3.3.3. Pathogens Detected in Pupae and Their Infesting Varroa
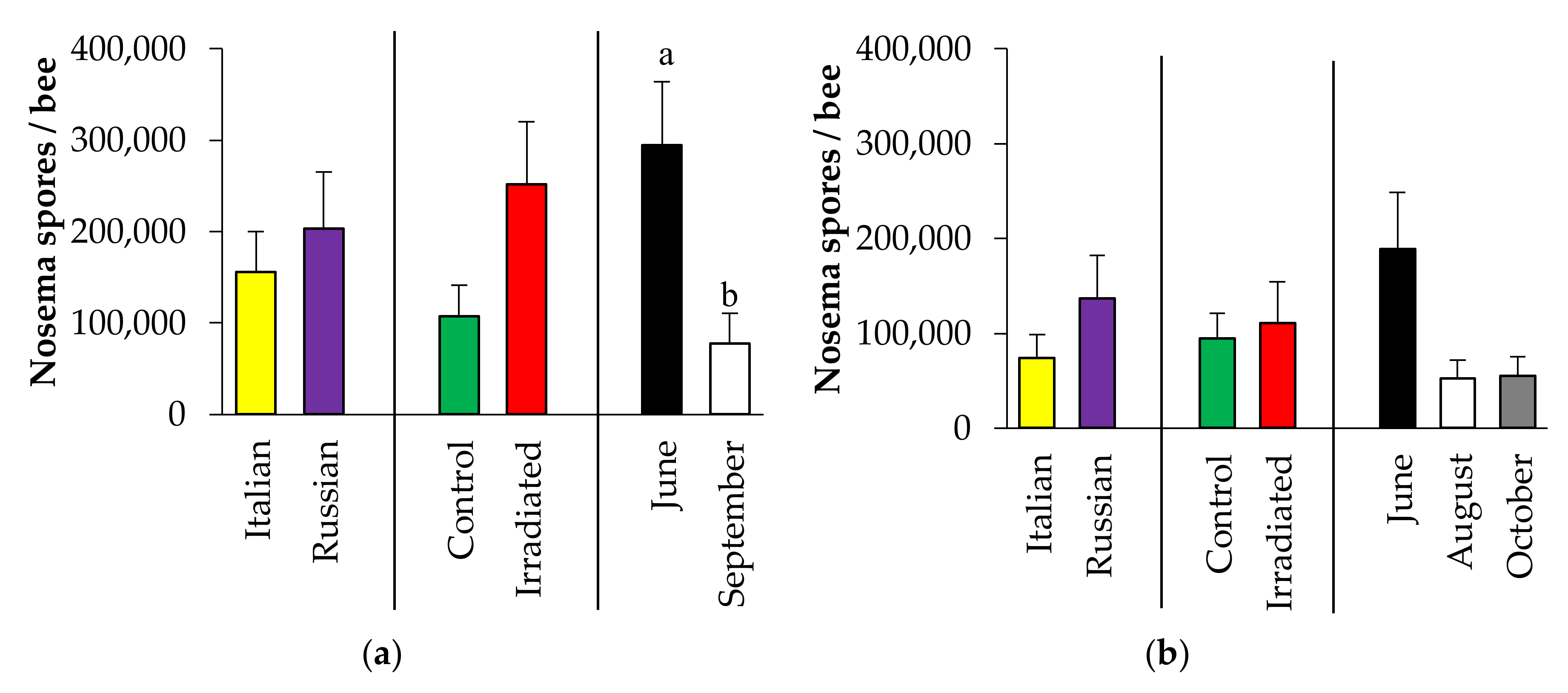
3.3.4. Nosema Spore Count
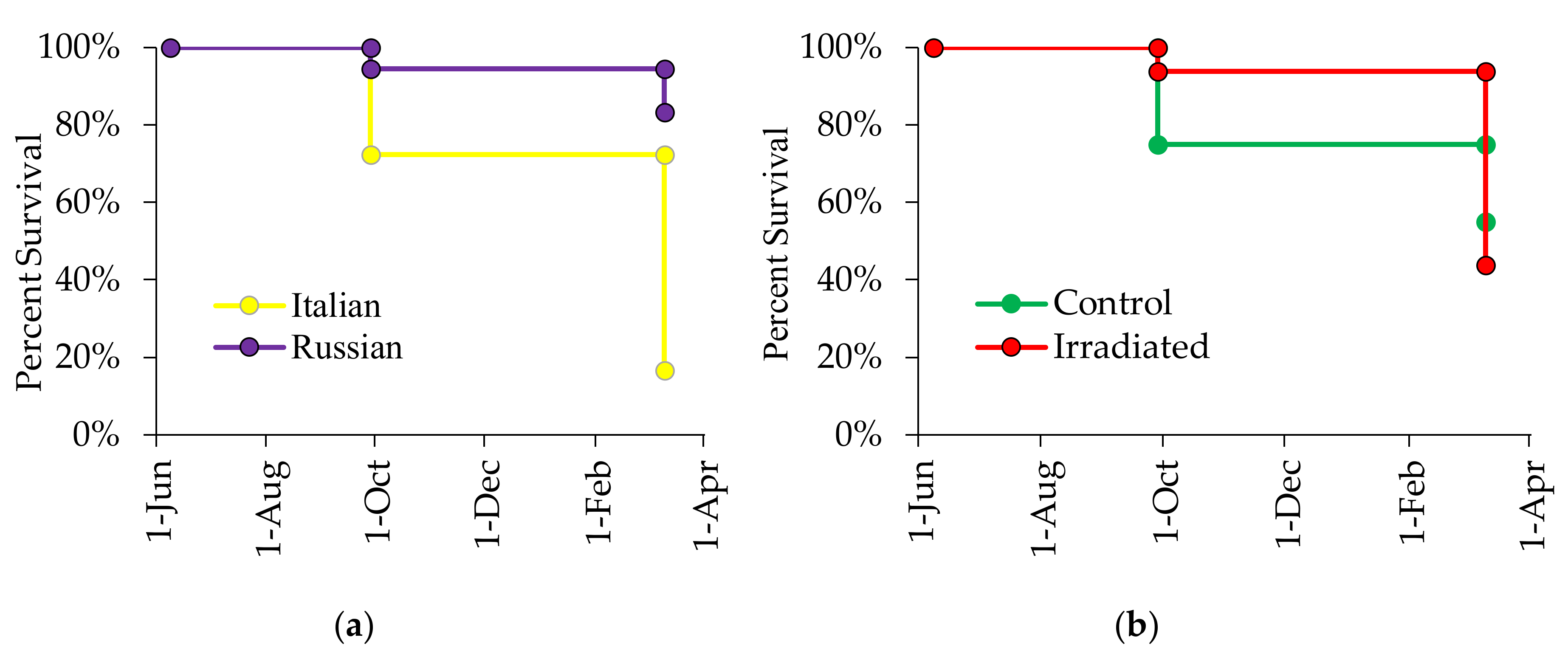
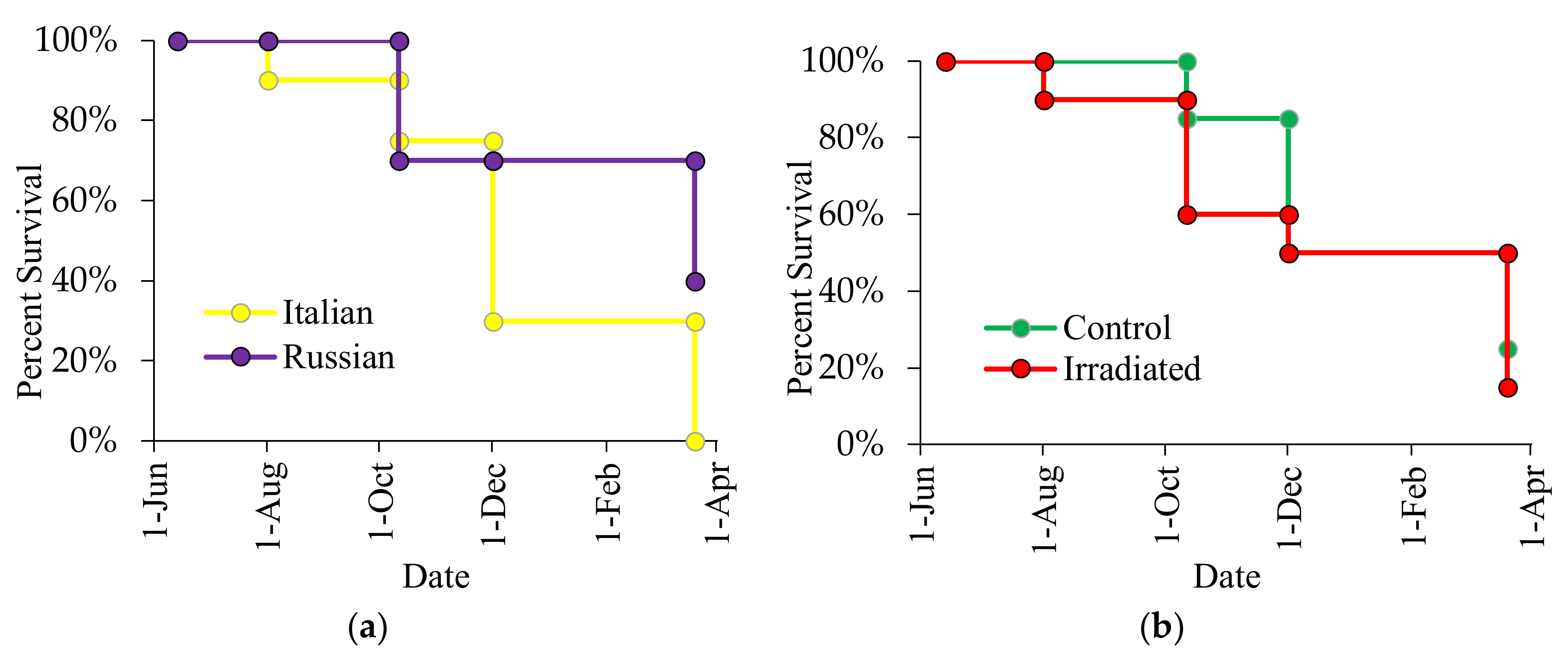
3.4. Effects of Honey Bee Stock and Comb Irradiation on Colony Survival
4. Discussion
5. Conclusions
Supplementary Materials
Author Contributions
Acknowledgments
Conflicts of Interest
References
- vanEngelsdorp, D.; Meixner, M.D. A historical review of managed honey bee populations in Europe and the United States and the factors that may affect them. J. Invertebr. Pathol. 2010, 103, S80–S95. [Google Scholar] [CrossRef] [PubMed]
- Mullin, C.A.; Frazier, M.; Frazier, J.L.; Ashcraft, S.; Simonds, R.; vanEngelsdorp, D.; Pettis, J.S. High levels of miticides and agrochemicals in North American apiaries: Implications for honey bee health. PLoS ONE 2010, 5, e9754. [Google Scholar] [CrossRef] [PubMed]
- Danka, R.G.; de Guzman, L.I.; Rinderer, T.E.; Allen Sylvester, H.; Wagener, C.M.; Bourgeois, A.L.; Harris, J.W.; Villa, J.D. Functionality of varroa-resistant honey bees (hymenoptera: Apidae) when used in migratory beekeeping for crop pollination. J. Econ. Entomol. 2012, 105, 313–321. [Google Scholar] [CrossRef] [PubMed]
- Rinderer, T.E.; Danka, R.G.; Johnson, S.; Bourgeois, A.L.; Frake, A.M.; Villa, J.D.; De Guzman, L.I.; Harris, J.W. Functionality of varroa-resistant honey bees (hymenoptera: Apidae) when used for western U.S. Honey production and almond pollination. J. Econ. Entomol. 2014, 107, 523–530. [Google Scholar] [CrossRef] [PubMed]
- Büchler, R.; Berg, S.; Le Conte, Y. Breeding for resistance to Varroa destructor in Europe. Apidologie 2010, 41, 393–408. [Google Scholar] [CrossRef]
- Rinderer, T.E.; Harris, J.W.; Hunt, G.; de Guzman, L.I. Breeding for resistance to Varroa destructor in North America. Apidologie 2010, 41, 409–424. [Google Scholar] [CrossRef]
- de Guzman, L.I.; Rinderer, T.E.; Frake, A.M. Growth of Varroa destructor (acari: Varroidae) populations in Russian honey bee (hymenoptera: Apidae) colonies. Ann. Entomol. Soc. Am. 2007, 100, 187–195. [Google Scholar]
- Delaplane, K.S.; Berry, J.A.; Skinner, J.A.; Parkman, J.P.; Hood, W.M. Integrated pest management against Varroa destructor reduces colony mite levels and delays treatment threshold. J. Apic. Res. 2005, 44, 157–162. [Google Scholar] [CrossRef]
- Wantuch, H.A.; Tarpy, D.R. Removal of drone brood from Apis mellifera (Hymenoptera: Apidae) colonies to control Varroa destructor (acari: Varroidae) and retain adult drones. J. Econ. Entomol. 2009, 102, 2033–2040. [Google Scholar] [CrossRef] [PubMed]
- Piccirillo, G.A.; De Jong, D. The influence of brood comb cell size on the reproductive behavior of the ectoparasitic mite Varroa destructor in africanized honey bee colonies. Genet. Mol. Res. GMR 2003, 2, 36–42. [Google Scholar] [PubMed]
- Message, D.; Gonçalves, L.S. Effect of the size of worker brood cells of africanized honey bees on infestation and reproduction of the ectoparasitic mite Varroa jacobsoni Oud. Apidologie 1995, 26, 381–386. [Google Scholar] [CrossRef]
- Martin, S.J.; Kryger, P. Reproduction of Varroa destructor in South African honey bees: Does cell space influence Varroa male survivorship? Apidologie 2002, 33, 51–61. [Google Scholar] [CrossRef]
- Nganso, B.T.; Fombong, A.T.; Yusuf, A.A.; Pirk, C.W.W.; Stuhl, C.; Torto, B. Low fertility, fecundity and numbers of mated female offspring explain the lower reproductive success of the parasitic mite Varroa destructor in African honeybees. Parasitology 2018, 145, 1633–1639. [Google Scholar] [CrossRef] [PubMed]
- Harbo, J.R.; Harris, J.W. Suppression of mite reproduction: A characteristic of honey bees that produces resistance to Varroa destructor. Am. Bee J. 2001, 141, 889. [Google Scholar]
- De Guzman, L.I.; Rinderer, T.E.; Frake, A.M. Comparative reproduction of Varroa destructor in different types of Russian and Italian honey bee combs. Exp. Appl. Acarol. 2008, 44, 227–238. [Google Scholar] [CrossRef] [PubMed]
- Jay, C.S. The development of honeybees in their cells. J. Agric. Res. 1963, 2, 117–134. [Google Scholar] [CrossRef]
- Hepburn, H.R.; Kurstjens, S.P. The combs of honeybees as composite materials. Apidologie 1988, 19, 25–36. [Google Scholar] [CrossRef]
- Free, J.B.; Williams, I.H. Factors determining food storage and brood rearing in honeybee (Apis mellifera L.) comb. J. Entomol. Ser. A 1974, 49, 47–63. [Google Scholar] [CrossRef]
- Traynor, K.S.; Pettis, J.S.; Tarpy, D.R.; Mullin, C.A.; Frazier, J.L.; Frazier, M.; vanEngelsdorp, D. In-hive pesticide exposome: Assessing risks to migratory honey bees from in-hive pesticide contamination in the eastern United States. Sci. Rep. 2016, 6, 33207. [Google Scholar] [CrossRef]
- Katznelson, H.; Jamieson, C.; Lawton, E.; Bellamy, W. Studies on the treatment of contaminated combs and honey with high velocity electrons. Can. J. Technol. 1952, 30, 95–103. [Google Scholar]
- James, R.R. Potential of ozone as a fumigant to control pests in honey bee (Hymenoptera: Apidae) hives. J. Econ. Entomol. 2011, 104, 353–359. [Google Scholar] [CrossRef] [PubMed]
- Colwell, M.J.; Currie, R.W.; Pernal, S.F. Viruses in unexpected places: New transmission routes of European honey bee (Apis mellifera) viruses. In Proceedings of the New Transmission Routes of European Honey Bee (Apis mellifera) Viruses, American Bee Research Conference, Galveston, TX, USA, 12–13 January 2017; Simone-Finstrom, M., Ed.; Bee World: Colorado Springs, CO, USA, 2017; pp. 106–107. [Google Scholar]
- Fries, I. Comb replacement and nosema disease (Nosema apis Z.) in honey bee colonies. Apidologie 1988, 19, 343–354. [Google Scholar] [CrossRef]
- Berry, J.A.; Delaplane, K.S. Effects of comb age on honey bee colony growth and brood survivorship. J. Apic. Res. 2001, 40, 3–8. [Google Scholar] [CrossRef]
- Seeley, T.D. Honey Bee Ecology; Princeton University Press: Princeton, NJ, USA, 1985; p. 201. [Google Scholar]
- Fries, I. The influence of comb building before and during the main honeyflow on the swarm tendency and honey yield of honey bee colonies. Am. Bee J. 1981, 121, 651–652, 655–656. [Google Scholar]
- Pettis, J.; vanEngelsdorp, D.; Cox-Foster, D. Colony collapse disorder working group pathogen sub-group progress report. Am. Bee J. 2007, 147, 595–597. [Google Scholar]
- Gochnauer, T.A.; Hamilton, H.A. Disinfection of honeybee combs by gamma irradiation I. American foul brood disease. J. Apic. Res. 1970, 9, 87–94. [Google Scholar] [CrossRef]
- Gosselin, P.; Charbonneau, R. Disinfection of the bee hive’s american foulbrood by gamma radiation from cobalt-60. Int. J. Radiat. Appl. Instrum. Part C Radiat. Phys. Chem. 1990, 35, 292–295. [Google Scholar] [CrossRef]
- The Bee Informed Partnership. The Bee Informed Partnership Management Survey Results (2012) Brood Comb Management. Available online: https://beeinformed.org/wp-content/uploads/2013/08/title-Brood-Comb-managment.pdf (accessed on 10 September 2018).
- de Guzman, L.I.; Rinderer, T.E.; Lancaster, V.A.; Delatte, G.T.; Stelzer, J.A. Varroa in the mating yard: III. The effects of formic acid gel formulation on drone production. Am. Bee J. 1999, 139, 304–307. [Google Scholar]
- Rinderer, T.E.; de Guzman, L.I.; Vancaster, V.; Delatte, G.T.; Stelzer, J.A. Varroa in the mating yard: I. The effects of Varroa jacobsoni and Apistan® on drone honey bees. Am. Bee J. 1999, 139, 134–139. [Google Scholar]
- De Guzman, Z.M.; Cervancia, C.R.; Dimasuay, K.G.B.; Tolentino, M.M.; Abrera, G.B.; Cobar, M.L.C.; Fajardo, A.C.; Sabino, N.G.; Manila-Fajardo, A.C.; Feliciano, C.P. Radiation inactivation of Paenibacillus larvae and sterilization of American foul brood (AFB) infected hives using co-60 gamma rays. Appl. Radiat. Isotopes 2011, 69, 1374–1379. [Google Scholar] [CrossRef]
- Simone-Finstrom, M.; Aronstein, K.; Goblirsch, M.; Rinkevich, F.; de Guzman, L. Gamma irradiation inactivates honey bee fungal, microsporidian, and viral pathogens and parasites. J. Invertebr. Pathol. 2018, 153, 57–64. [Google Scholar] [CrossRef] [PubMed]
- Graystock, P.; Jones, J.C.; Pamminger, T.; Parkinson, J.F.; Norman, V.; Blane, E.J.; Rothstein, L.; Wäckers, F.; Goulson, D.; Hughes, W.O.H. Hygienic food to reduce pathogen risk to bumblebees. J. Invertebr. Pathol. 2016, 136, 68–73. [Google Scholar] [CrossRef] [PubMed]
- Meeus, I.; Mosallanejad, H.; Niu, J.; de Graaf, D.C.; Wackers, F.; Smagghe, G. Gamma irradiation of pollen and eradication of Israeli acute paralysis virus. J. Invertebr. Pathol. 2014, 121, 74–77. [Google Scholar] [CrossRef]
- de Guzman, L.; Frake, A.; Simone-Finstrom, M. Comparative flight activities and pathogen load of two stocks of honey bees reared in gamma-irradiated combs. Insects 2017, 8, 127. [Google Scholar] [CrossRef] [PubMed]
- Rogers, L.E.; Gilbert, R.O.; Burgett, M. Sampling honeybee colonies for brood production: A double sampling technique. J. Apic. Res. 1983, 22, 232–241. [Google Scholar] [CrossRef]
- Burgett, M.; Burikam, I. Number of adult honey bees (Hymenoptera: Apidae) occupying a comb: A standard for estimating colony populations. J. Econ. Entomol. 1985, 78, 1154–1156. [Google Scholar] [CrossRef]
- Rinderer, T.; de Guzman, L.; Sylvester, H.A. Re-examination of the accuracy of a detergent solution for varroa mite detection. Am. Bee J. 2004, 144, 560–562. [Google Scholar]
- Dietemann, V.; Nazzi, F.; Martin, S.J.; Anderson, D.L.; Locke, B.; Delaplane, K.S.; Wauquiez, Q.; Tannahill, C.; Frey, E.; Ziegelmann, B.; et al. Standard methods for varroa research. J. Apic. Res. 2013, 52, 1–54. [Google Scholar] [CrossRef]
- Rinderer, T.E.; de Guzman, L.I.; Delatte, G.T.; Stelzer, J.A.; Williams, J.L.; Beaman, L.D.; Kuznetsov, V.; Bernard, S.J.; Bigalk, M.; Tubbs, H. Multi-state field trials of ARS Russian honey bees 1. Responses to Varroa destructor 1999, 2000. Am. Bee J. 2001, 141, 658–661. [Google Scholar]
- Ryabov, E.V.; Childers, A.K.; Chen, Y.; Madella, S.; Nessa, A.; vanEngelsdorp, D.; Evans, J.D. Recent spread of Varroa destructor virus-1, a honey bee pathogen, in the United States. Sci. Rep. 2017, 7, 17447. [Google Scholar] [CrossRef]
- Mordecai, G.J.; Wilfert, L.; Martin, S.J.; Jones, I.M.; Schroeder, D.C. Diversity in a honey bee pathogen: First report of a third master variant of the deformed wing virus quasispecies. Isme J. 2016, 10, 1264. [Google Scholar] [CrossRef] [PubMed]
- Chen, Y.P.; Nakashima, N.; Christian, P.D.; Bakonyi, T.; Bonning, B.C.; Valles, S.M.; Lightner, D.V. Family iflaviridae Virus Taxonomy: Ninth Report of the International Committee on Taxonomy of Viruses; Andrew, M.Q., King, M.J.A., Carstens, E.B., Lefkowitz, E.J., Eds.; Elsevier Academic: London, UK, 2012; pp. 846–849. [Google Scholar]
- Boncristiani, H.F.; Evans, J.D.; Chen, Y.P.; Pettis, J.; Murphy, C.; Lopez, D.L.; Simone-Finstrom, M.; Strand, M.; Tarpy, D.R.; Rueppell, O. In vitro infection of pupae with Israeli acute aralysis virus suggests disturbance of transcriptional homeostasis in honey bees (Apis mellifera). PLoS ONE 2013, 8, e73429. [Google Scholar] [CrossRef] [PubMed]
- Ryabov, E.V.; Wood, G.R.; Fannon, J.M.; Moore, J.D.; Bull, J.C.; Chandler, D.; Mead, A.; Burroughs, N.; Evans, D.J. A virulent strain of Deformed wing virus (DWV) of honeybees (apis mellifera) prevails after Varroa destructor-mediated, or in vitro, transmission. PLoS Pathog. 2014, 10, e1004230. [Google Scholar] [CrossRef] [PubMed]
- Schmittgen, T.D.; Livak, K.J. Analyzing real-time pcr data by the comparative ct method. Nat. Protoc. 2008, 3, 1101–1108. [Google Scholar] [CrossRef] [PubMed]
- Fries, I.; Chauzat, M.-P.; Chen, Y.-P.; Doublet, V.; Genersch, E.; Gisder, S.; Higes, M.; McMahon, D.P.; Martín-Hernández, R.; Natsopoulou, M.; et al. Standard methods for nosema research. J. Apic. Res. 2013, 52, 1–28. [Google Scholar] [CrossRef]
- Rinderer, T.E.; de Guzman, L.I.; Delatte, G.T.; Stelzer, J.A.; Kuznetsov, V.N.; Beaman, L.D.; Watts, R.; Harris, J. Resistance to the parasitic mite Varroa destructor in honey bees from far-eastern Russia. Apidologie 2001, 32, 381–394. [Google Scholar] [CrossRef]
- De Guzman, L.I.; Rinderer, T.E.; Bigalk, M.; Tubbs, H.; Bernard, S.J. Russian honey bee (Hymenoptera: Apidae) colonies: Acarapis woodi (acari: Tarsonemidae) infestations and overwintering survival. J. Econ. Entomol. 2005, 98, 1796–1801. [Google Scholar] [CrossRef]
- Villa, J.D.; Rinderer, T.E.; Bigalk, M. Overwintering of Russian honey bees in northeastern Iowa. Sci. Bee Cult. 2009, 1, 19–21. [Google Scholar]
- de Groot, A.P. Protein and amino acid requirements of the honey-bee (Apis mellifera L.). Physiol. Comp. Oecol. 1953, 3, 197–285. [Google Scholar]
- McLellan, A.R. Growth and decline of honeybee colonies and inter-relationships of adult bees, brood, honey and pollen. J. Appl. Ecol. 1978, 15, 155–161. [Google Scholar] [CrossRef]
- Carroll, M.J.; Brown, N.; Goodall, C.; Downs, A.M.; Sheenan, T.H.; Anderson, K.E. Honey bees preferentially consume freshly-stored pollen. PLoS ONE 2017, 12, e0175933. [Google Scholar] [CrossRef] [PubMed]
- Tentcheva, D.; Gauthier, L.; Zappulla, N.; Dainat, B.; Cousserans, F.; Colin, M.E.; Bergoin, M. Prevalence and seasonal variations of six bee viruses in Apis mellifera L. and Varroa destructor mite populations in France. Appl. Environ. Microbiol. 2004, 70, 7185–7191. [Google Scholar] [CrossRef] [PubMed]
- Martin, S.J.; Highfield, A.C.; Brettell, L.; Villalobos, E.M.; Budge, G.E.; Powell, M.; Nikaido, S.; Schroeder, D.C. Global honey bee viral landscape altered by a parasitic mite. Science 2012, 336, 1304–1306. [Google Scholar] [CrossRef] [PubMed]
- Khongphinitbunjong, K.; de Guzman, L.I.; Rinderer, T.E.; Tarver, M.R.; Frake, A.M.; Chen, Y.; Chantawannakul, P. Responses of varroa-resistant honey bees (Apis mellifera L.) to Deformed wing virus. J. Asia-Pac. Entomol. 2016, 19, 921–927. [Google Scholar] [CrossRef]
- Mondet, F.; Kim, S.H.; de Miranda, J.R.; Beslay, D.; Le Conte, Y.; Mercer, A.R. Specific cues associated with honey bee social defence against Varroa destructor infested brood. Sci. Rep. 2016, 6, 25444. [Google Scholar] [CrossRef] [PubMed]
- Schoning, C.; Gisder, S.; Geiselhardt, S.; Kretschmann, I.; Bienefeld, K.; Hilker, M.; Genersch, E. Evidence for damage-dependent hygienic behaviour towards Varroa destructor-parasitised brood in the western honey bee, Apis mellifera. J. Exp. Biol. 2012, 215, 264–271. [Google Scholar] [CrossRef] [PubMed]
- Guzman, L.I.d.; Rinderer, T.E.; Delatte, G.T.; Stelzer, J.A.; Beaman, L.D.; Harper, C. Hygienic behavior by honey bees from far-eastern Russia. Am. Bee J. 2002, 142, 58–60. [Google Scholar]
- Kirrane, M.J.; de Guzman, L.I.; Holloway, B.; Frake, A.M.; Rinderer, T.E.; Whelan, P.M. Phenotypic and genetic analyses of the Varroa sensitive hygienic trait in Russian honey bee (hymenoptera: Apidae) colonies. PLoS ONE 2015, 10, e0116672. [Google Scholar] [CrossRef]
- De Guzman, L.I.; Rinderer, T.E.; Frake, A.M.; Kirrane, M.J. Brood removal influences fall of Varroa destructor in honey bee colonies. J. Apic. Res. 2015, 54, 216–225. [Google Scholar] [CrossRef]
- Kirrane, M.J.; de Guzman, L.I.; Whelan, P.M.; Frake, A.M.; Rinderer, T.E. Evaluations of the removal of Varroa destructor in Russian honey bee colonies that display different levels of Varroa sensitive hygienic activities. J. Insect Behav. 2018, 31, 283–297. [Google Scholar] [CrossRef]
- Munch, D.; Amdam, G.V.; Wolschin, F. Ageing in a eusocial insect: Molecular and physiological characteristics of life span plasticity in the honey bee. Funct. Ecol. 2008, 22, 407–421. [Google Scholar] [CrossRef] [PubMed]
- Traver, B.E.; Williams, M.R.; Fell, R.D. Comparison of within hive sampling and seasonal activity of Nosema ceranae in honey bee colonies. J. Invertebr. Pathol. 2012, 109, 187–193. [Google Scholar] [CrossRef] [PubMed]
- Smith, M.L. The honey bee parasite Nosema ceranae: Transmissible via food exchange? PLoS ONE 2012, 7, e43319. [Google Scholar] [CrossRef] [PubMed]
- Porrini, M.P.; Sarlo, E.G.; Medici, S.K.; Garrido, P.M.; Porrini, D.P.; Damiani, N.; Eguaras, M.J. Nosema ceranae development in Apis mellifera: Influence of diet and infective inoculum. J. Apic. Res. 2011, 50, 35–41. [Google Scholar] [CrossRef]
- Rinderer, T.E.; de Guzman, L.I.; Bourgeois, L.; Frake, A.M. The effects of hive size, feeding and Nosema ceranae on the size of winter clusters of Russian honey bee colonies. Sci. Bee Cult. 2010, 2, 1–6. [Google Scholar]
- Bourgeois, A.L.; Rinderer, T.E.; Sylvester, H.A.; Holloway, B.; Oldroyd, B.P. Patterns of Apis mellifera infestation by Nosema ceranae support the parasite hypothesis for the evolution of extreme polyandry in Eusocial insects. Apidologie 2012, 43, 539–548. [Google Scholar] [CrossRef]
- Chen, Y.-W.; Chung, W.-P.; Wang, C.-H.; Solter, L.F.; Huang, W.-F. Nosema ceranae infection intensity highly correlates with temperature. J. Invertebr. Pathol. 2012, 111, 264–267. [Google Scholar] [CrossRef]
- Di Pasquale, G.; Salignon, M.; Le Conte, Y.; Belzunces, L.P.; Decourtye, A.; Kretzschmar, A.; Suchail, S.; Brunet, J.-L.; Alaux, C. Influence of pollen nutrition on honey bee health: Do pollen quality and diversity matter? PLoS ONE 2013, 8, e72016. [Google Scholar] [CrossRef]
- Van Dooremalen, C.; Stam, E.; Gerritsen, L.; Cornelissen, B.; van der Steen, J.; van Langevelde, F.; Blacquiere, T. Interactive effect of reduced pollen availability and Varroa destructor infestation limits growth and protein content of young honey bees. J. Insect Physiol. 2013, 59, 487–493. [Google Scholar] [CrossRef]
- Nazzi, F.; Le Conte, Y. Ecology of Varroa destructor, the major ectoparasite of the western honey bee, Apis mellifera. Annu. Rev. Entomol. 2016, 61, 417–432. [Google Scholar] [CrossRef]












{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Treatment Type | VDV1 | DWV | BQVC | CBPV | Total Pathogens Detected |
|---|---|---|---|---|---|
| Control | r = 0.897, p < 0.0001 | r = 0.674, p = 0.003 | r = 0.173, p = 0.508 | r = 0.312, p = 0.222 | r = 0.122, p = 0.642 |
| Irradiated | r = 0.833, p < 0.0001 | r = 0.585, p = 0.011 | r = 0.404, p = 0.096 | r = 0.285, p = 0.251 | r = 0.324, p = 0.189 |
| Italian | r = 0.922, p < 0.0001 | r = 0.687, p = 0.002 | r = 0.475, p = 0.046 | r = 0.352, p = 0.152 | r = 0.121, p = 0.631 |
| Russian | r = 0.737, p < 0.0001 | r = 0.324, p = 0.204 | r = 0.180, p = 0.489 | r = 0.534, p = 0.024 | r = 0.255, p = 0.323 |
| Italian, Control (n = 9) | r = 0.968, p < 0.0001 | r = 0.802, p = 0.009 | r = 0.413, p = 0.269 | r = −0.456, p = 0.218 | r = −0.791, p = 0.011 |
| Italian, Irradiated (n = 9) | r = 0.865, p = 0.003 | r = 0.829, p = 0.006 | r = 0.527, p = 0.145 | r = −0.034, p = 0.932 | r = 0.661, p = 0.052 |
| Russian, Control (n = 8) | r = 0.781, p = 0.022 | r = 0.574, p = 0.137 | r = −0.084, p = 0.843 | r = 0.236, p = 0.574 | r = 0.535, p = 0.172 |
| Russian, Irradiated (n = 9) | r = 0.835, p = 0.005 | r = −0.083, p = 0.831 | r = 0.245, p = 0.525 | r = 0.816, p = 0.007 | r = 0.124, p = 0.751 |
| Overall (n = 35) | r = 0.846, p < 0.0001 | r = 0.636, p < 0.0001 | r = 0.350, p = 0.039 | r = 0.492, p = 0.003 | r = 0.236, p = 0.172 |
© 2019 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
De Guzman, L.I.; Simone-Finstrom, M.; Frake, A.M.; Tokarz, P. Comb Irradiation Has Limited, Interactive Effects on Colony Performance or Pathogens in Bees, Varroa destructor and Wax Based on Two Honey Bee Stocks. Insects 2019, 10, 15. https://doi.org/10.3390/insects10010015
De Guzman LI, Simone-Finstrom M, Frake AM, Tokarz P. Comb Irradiation Has Limited, Interactive Effects on Colony Performance or Pathogens in Bees, Varroa destructor and Wax Based on Two Honey Bee Stocks. Insects. 2019; 10(1):15. https://doi.org/10.3390/insects10010015
Chicago/Turabian StyleDe Guzman, Lilia I., Michael Simone-Finstrom, Amanda M. Frake, and Philip Tokarz. 2019. "Comb Irradiation Has Limited, Interactive Effects on Colony Performance or Pathogens in Bees, Varroa destructor and Wax Based on Two Honey Bee Stocks" Insects 10, no. 1: 15. https://doi.org/10.3390/insects10010015
APA StyleDe Guzman, L. I., Simone-Finstrom, M., Frake, A. M., & Tokarz, P. (2019). Comb Irradiation Has Limited, Interactive Effects on Colony Performance or Pathogens in Bees, Varroa destructor and Wax Based on Two Honey Bee Stocks. Insects, 10(1), 15. https://doi.org/10.3390/insects10010015