
Cytotoxic Activity of Melatonin Alone and in Combination with Doxorubicin and/or Dexamethasone on Diffuse Large B-Cell Lymphoma Cells in In Vitro Conditions

 , ,
, , {kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
:1. Introduction
2. Materials and Methods
2.1. Cell Line
2.2. MLT and Drug Preparation
2.3. Treatment Protocol
2.4. Cytotoxicity Assay
2.5. Assessment of Apoptosis
2.6. Flow Cytometry Analysis
2.7. Statistical Analysis
3. Results
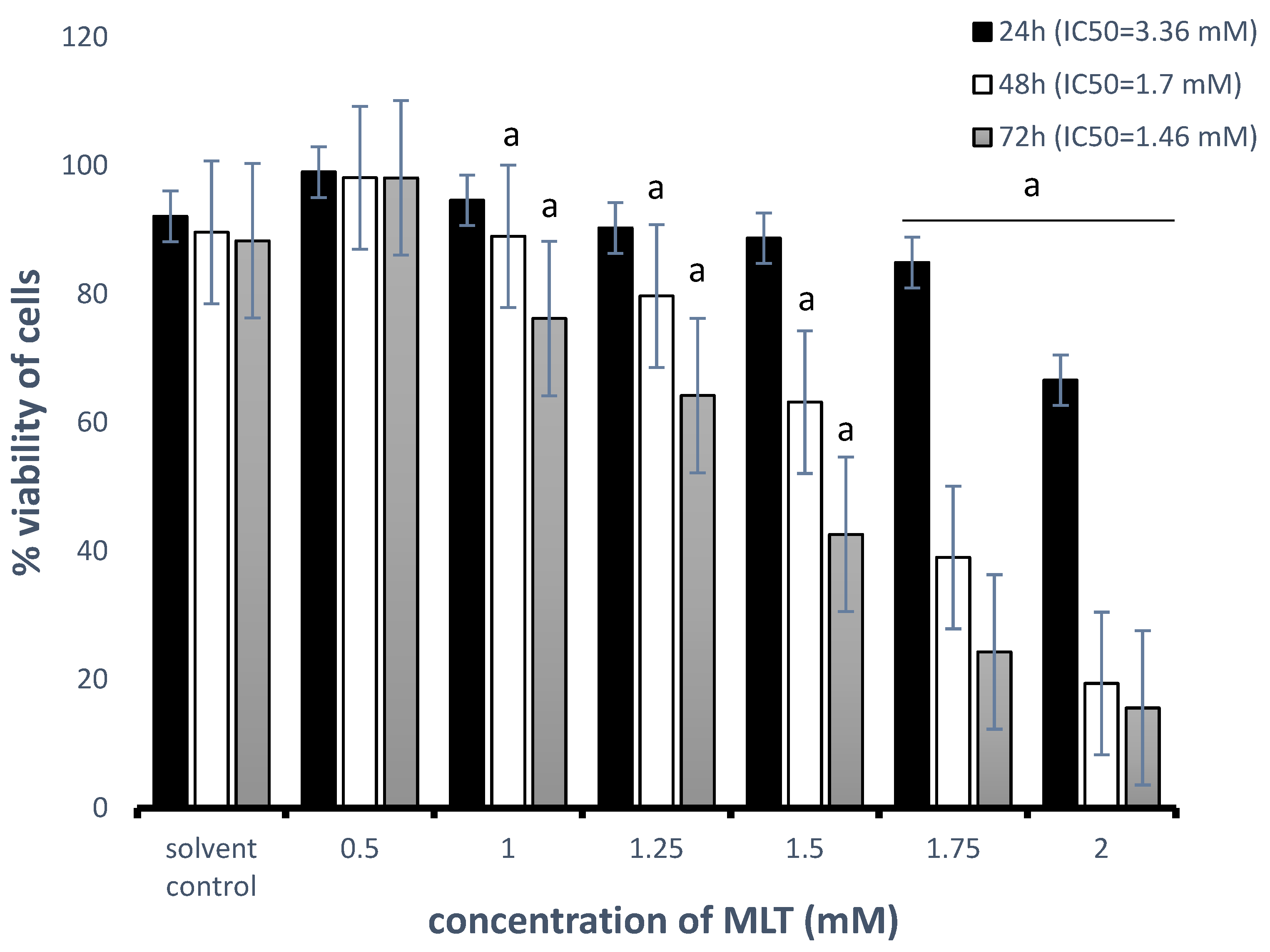
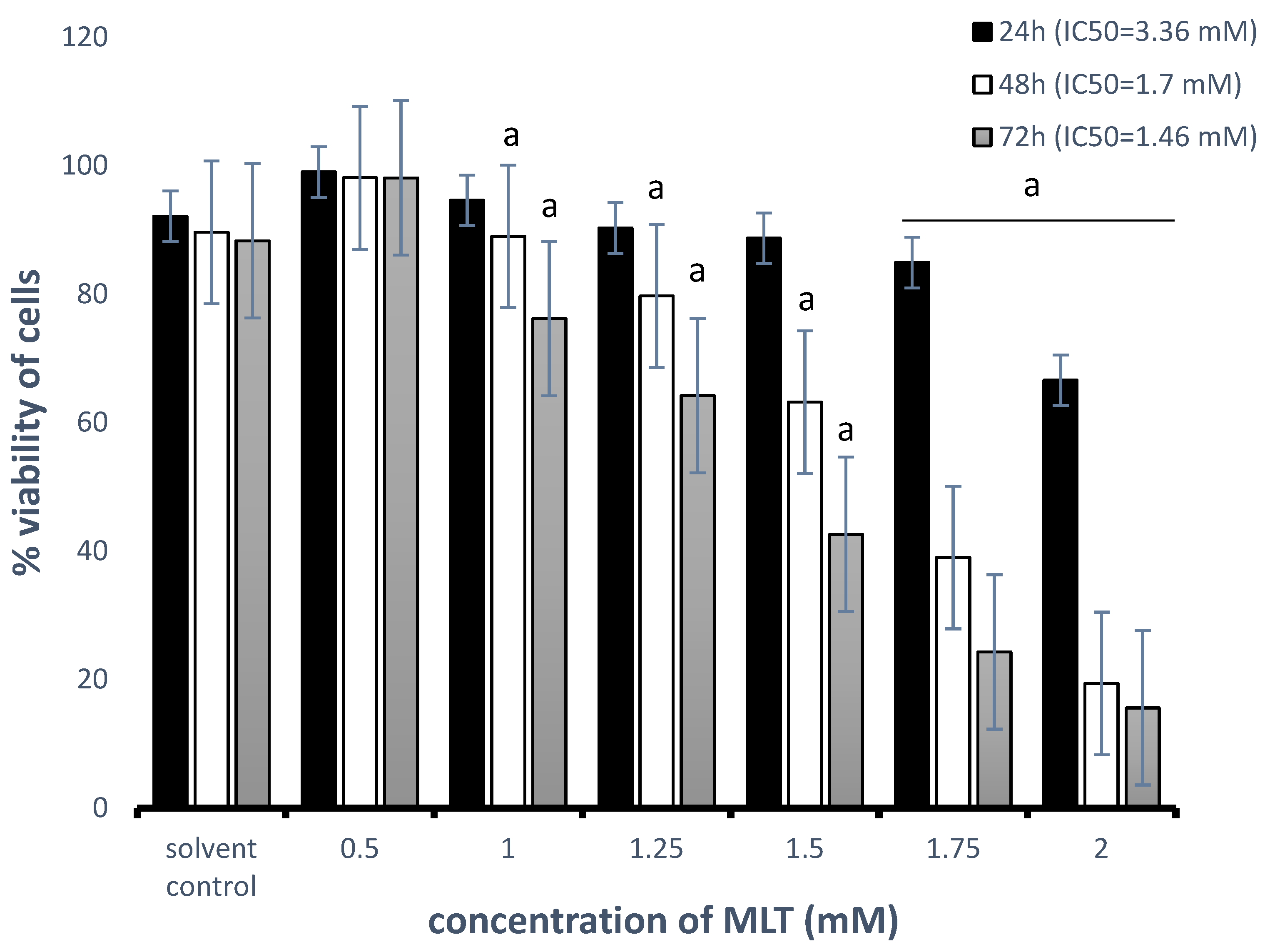
3.1. Cytotoxic Effect of MLT Alone and in Combination with Drugs on Toledo Cells
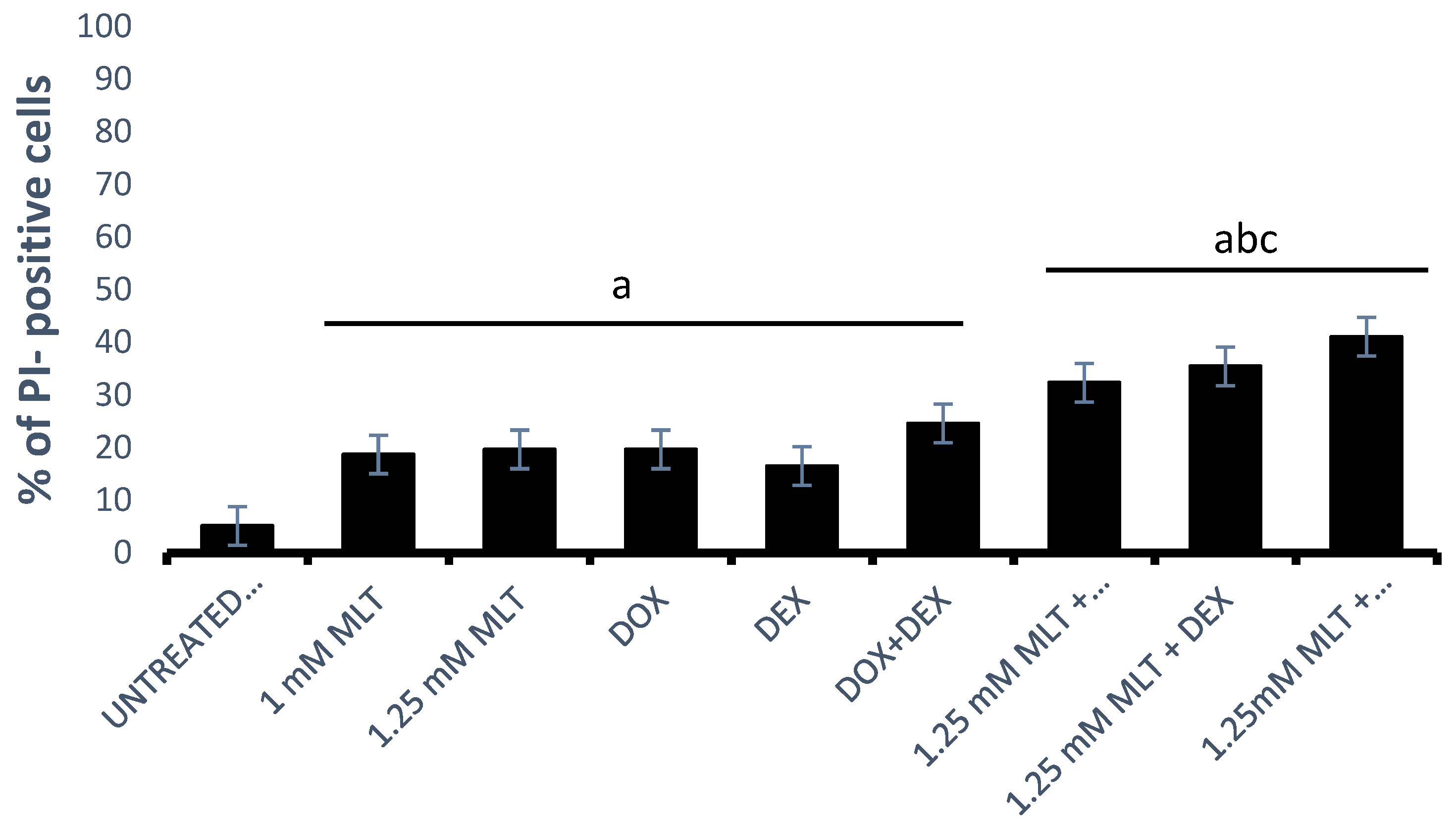
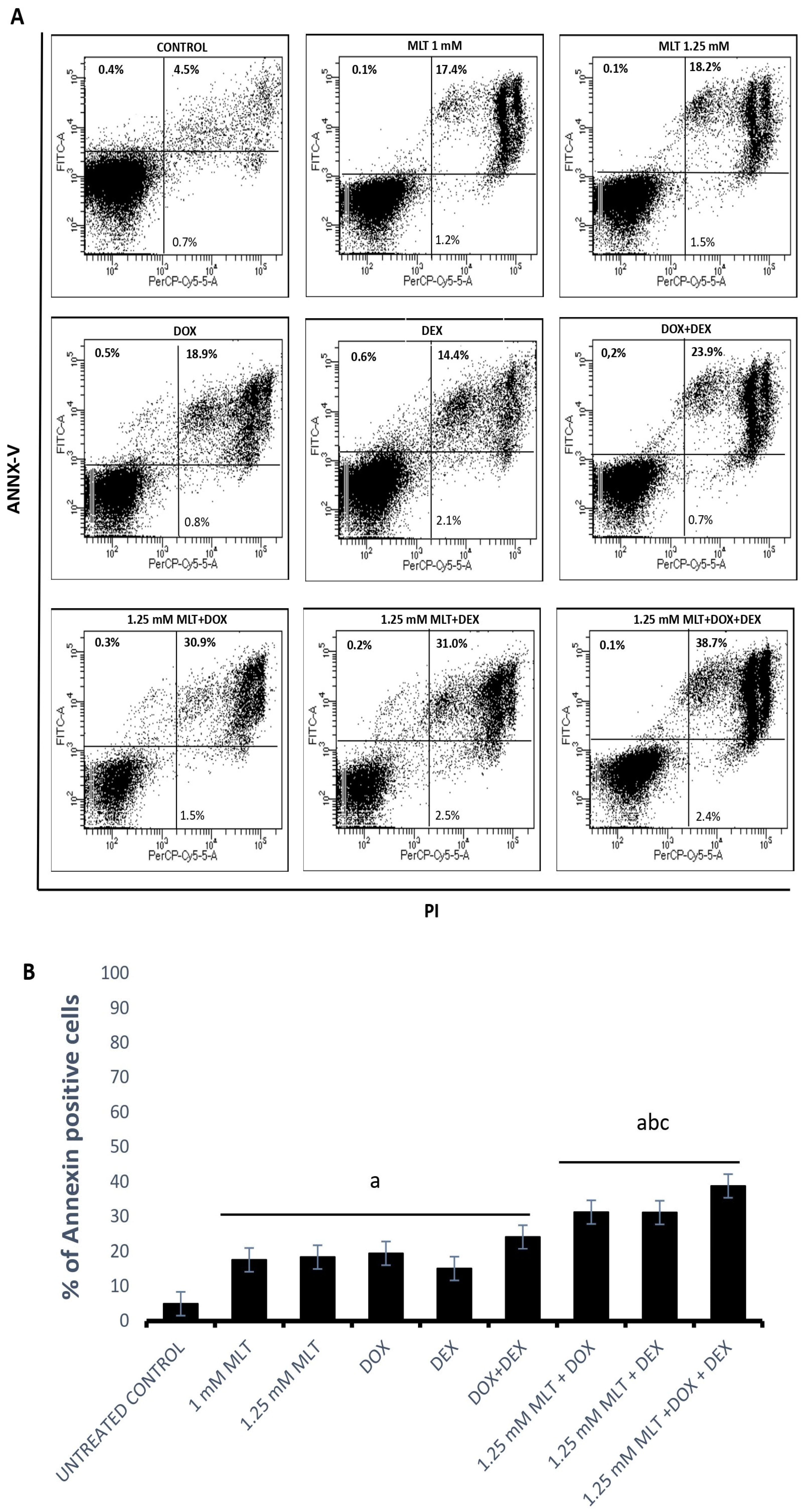
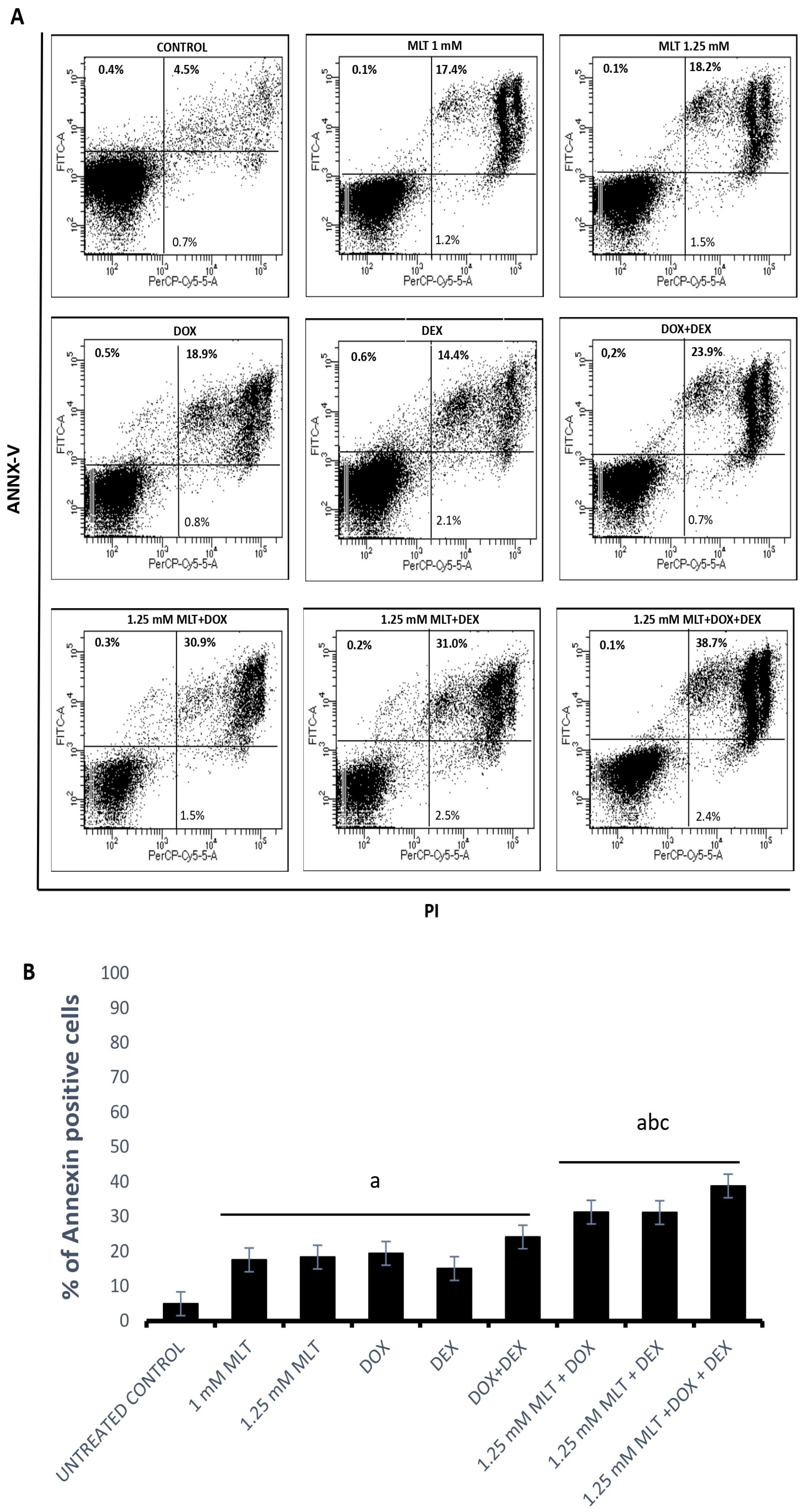
3.2. Pro-Apoptotic Effects of MLT Alone and in Combination with Other Drugs
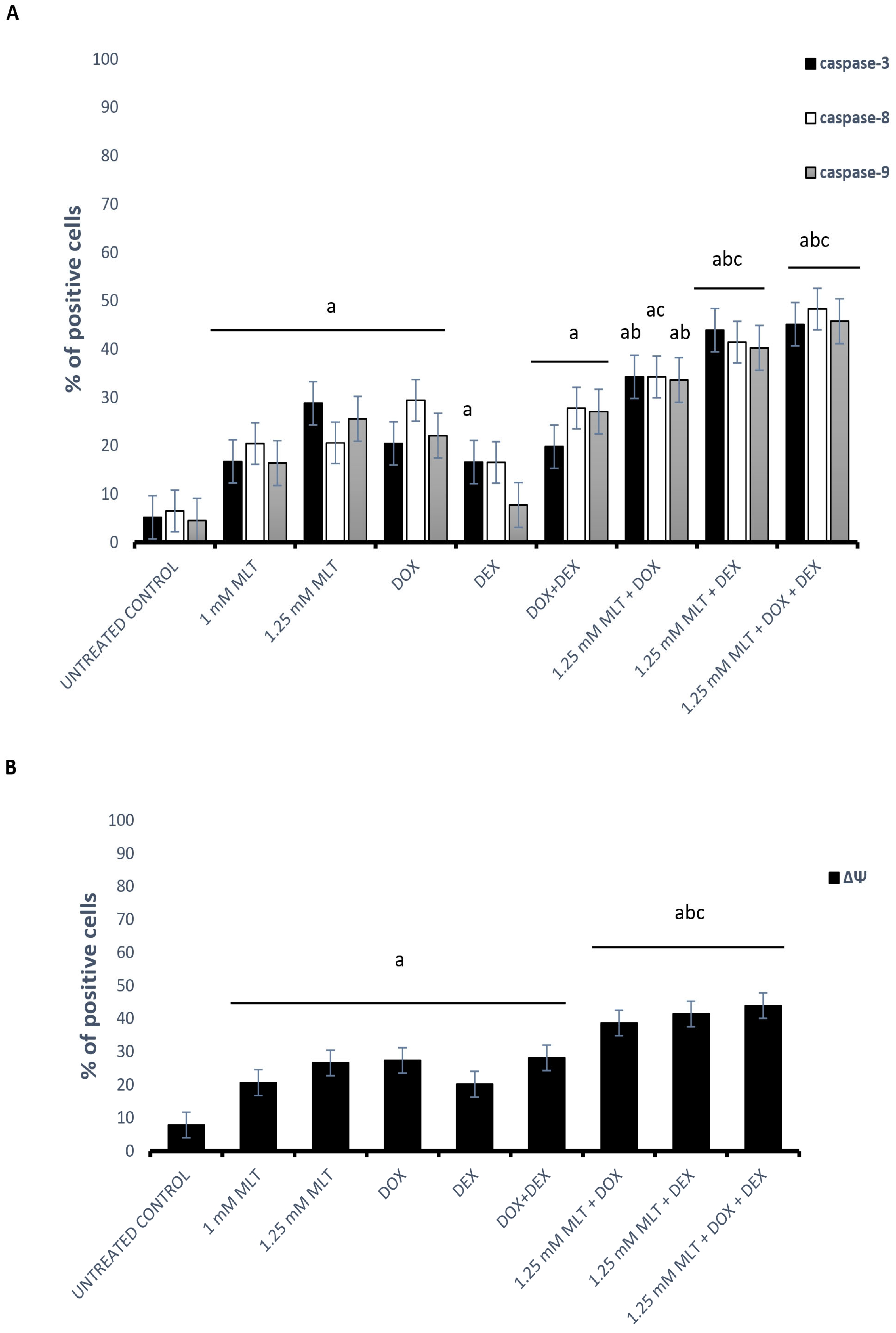
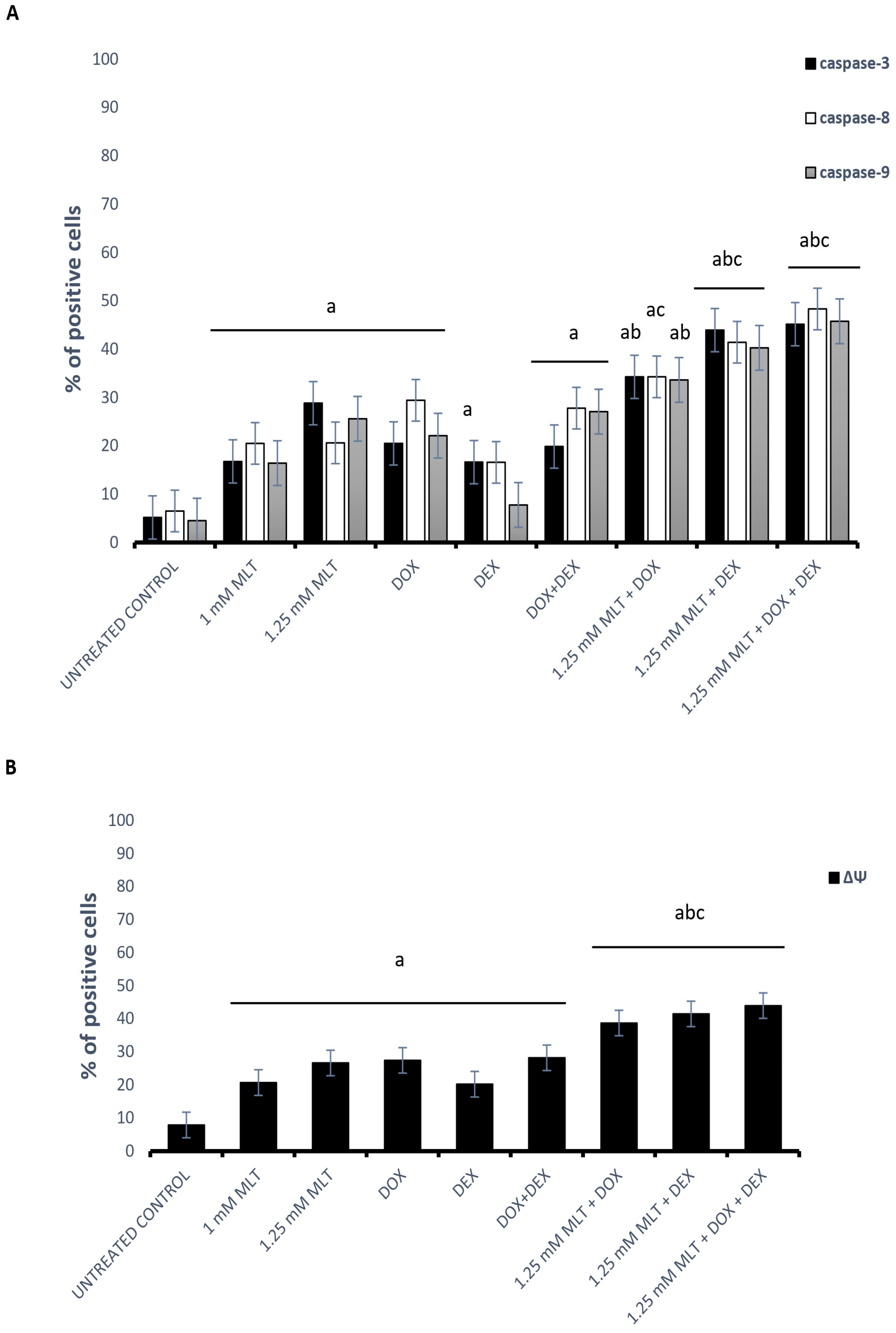
3.3. The Mechanisms of the Pro-Apoptotic Activity of MLT Alone and in Combination with Other Drugs
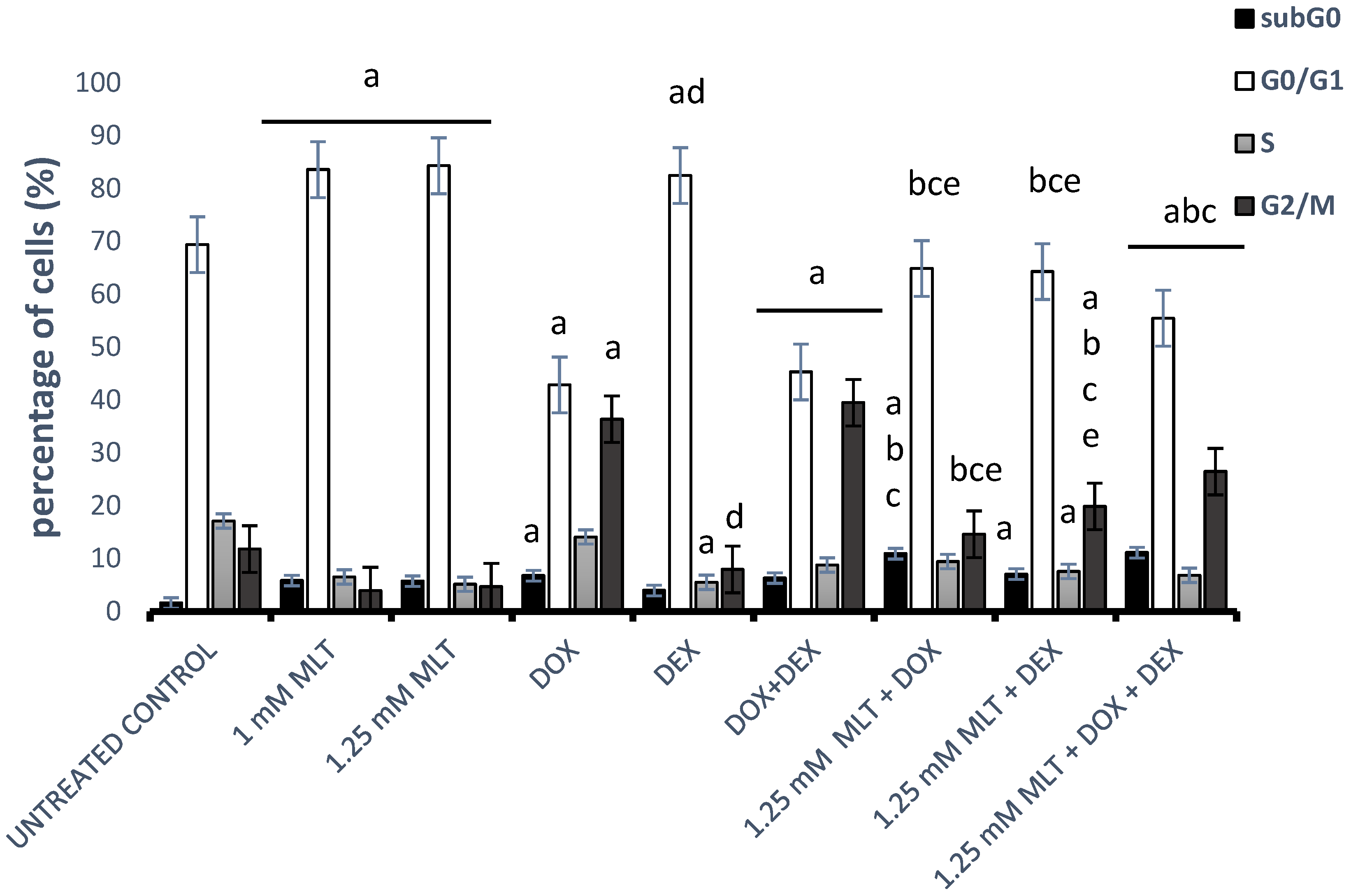
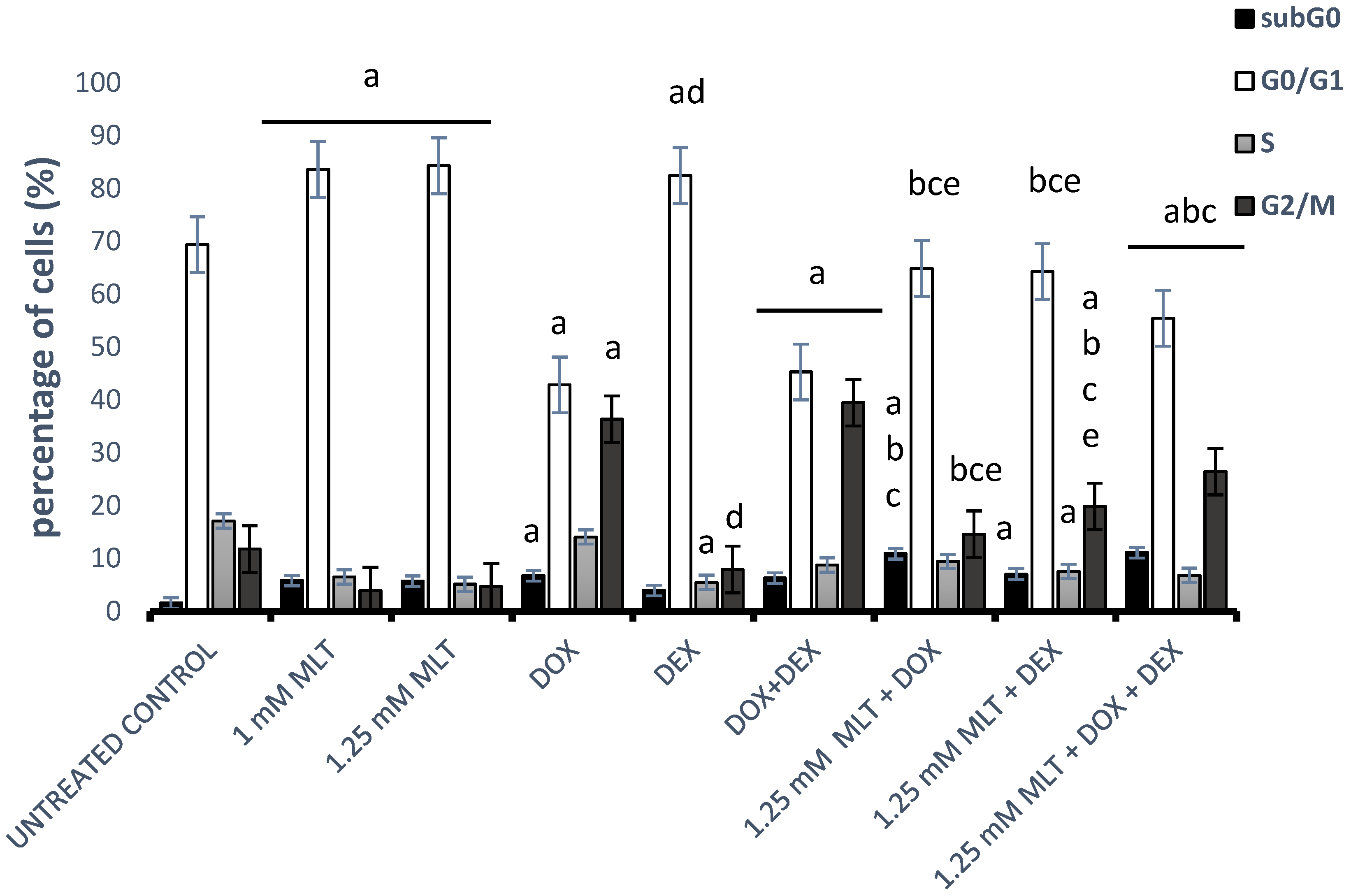
3.4. Effect of MLT Alone and in Combination with Other Drugs on Cell Cycle Distribution of DLBCL-Derived Cells
4. Discussion
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Sehn, L.H.; Gascoyne, R.D. Diffuse large B-cell lymphoma: Optimizing outcome in the context of clinical and biologic heterogeneity. Blood 2015, 125, 22–32. [Google Scholar] [CrossRef]
- Thieblemont, C.; Coiffier, B. Lymphoma in older patients. J. Clin. Oncol. 2007, 25, 1916–1923. [Google Scholar] [CrossRef] [PubMed]
- Alizadeh, A.A.; Eisen, M.B.; Davis, R.E.; Ma, C.; Lossos, I.S.; Rosenwald, A.; Boldrick, J.C.; Sabet, H.; Tran, T.; Yu, X.; et al. Distinct types of diffuse large B-cell lymphoma identified by gene expression profiling. Nature 2000, 403, 503–511. [Google Scholar] [CrossRef] [PubMed]
- Pandi-Perumal, S.R.; Srinivasan, V.; Maestroni, G.J.M.; Cardinali, D.P.; Poeggeler, B.; Hardeland, R. Melatonin: Nature’s most versatile biological signal? FEBS J. 2006, 273, 2813–2838. [Google Scholar] [CrossRef] [PubMed]
- Carrillo-Vico, A.; Calvo, J.R.; Abreu, P.; Lardone, P.J.; García-Mauriño, S.; Reiter, R.J.; Guerrero, J.M. Evidence of melatonin synthesis by human lymphocytes and its physiological significance: Possible role as intracrine, autocrine, and/or paracrine substance. FASEB J. 2004, 18, 537–539. [Google Scholar] [CrossRef]
- Boutin, J.A.; Kennaway, D.J.; Jockers, R. Melatonin: Facts, Extrapolations and Clinical Trials. Biomolecules 2023, 13, 943. [Google Scholar] [CrossRef]
- Sanchez-Barcelo, E.J.; Rueda, N.; Mediavilla, M.D.; Martinez-Cue, C.; Reiter, R.J. Clinical Uses of Melatonin in Neurological Diseases and Mental and Behavioural Disorders. Curr. Med. Chem. 2017, 24, 3851–3878. [Google Scholar] [CrossRef] [PubMed]
- Karasek, M. Melatonin, human aging, and age-related diseases. Exp. Gerontol. 2004, 39, 1723–1729. [Google Scholar] [CrossRef] [PubMed]
- Srinivasan, V.; Spence, D.W.; Trakht, I.; Pandi-Perumal, S.R.; Cardinali, D.P.; Maestroni, G.J. Immunomodulation by melatonin: Its significance for seasonally occurring diseases. Neuroimmunomodulation 2008, 15, 93–101. [Google Scholar] [CrossRef]
- Hardeland, R. Melatonin in aging and disease -multiple consequences of reduced secretion, options and limits of treatment. Aging Dis. 2012, 3, 194–225. [Google Scholar]
- Rana, S.; Munawar, M.; Shahid, A.; Malik, M.; Ullah, H.; Fatima, W.; Mohsin, S.; Mahmood, S. Deregulated expression of circadian clock and clock-controlled cell cycle genes in chronic lymphocytic leukemia. Mol. Biol. Rep. 2014, 41, 95–103. [Google Scholar] [CrossRef]
- Grubisic, M.; Haim, A.; Bhusal, P.; Dominoni, D.M.; Gabriel, K.M.; Jechow, A.; Kupprat, F.; Lerner, A.; Marchant, P.; Riley, W.; et al. Light Pollution, Circadian Photoreception, and Melatonin in Vertebrates. Sustainability 2019, 11, 6400. [Google Scholar] [CrossRef]
- Zhong, C.; Franklin, M.; Wiemels, J.; McKean-Cowdin, R.; Chung, N.T.; Benbow, J.; Wang, S.S.; Lacey, J.V., Jr.; Longcore, T. Outdoor artificial light at night and risk of non-Hodgkin lymphoma among women in the California Teachers Study cohort. Cancer Epidemiol. 2020, 69, 101811. [Google Scholar] [CrossRef]
- Cocco, P.; Piro, S.; Meloni, F.; Montagna, A.; Pani, M.; Pilia, I.; Padoan, M.; Miligi, L.; Magnani, C.; Gambelunghe, A.; et al. Night shift work and lymphoma: Results from an Italian multicentre case-control study. Occup. Environ. Med. 2022, 79, 452–459. [Google Scholar] [CrossRef] [PubMed]
- Zhang, Y.; Birmann, B.M.; Papantoniou, K.; Zhou, E.S.; Erber, A.C.; Schernhammer, E.S. Rotating Nightshift Work and Hematopoietic Cancer Risk in US Female Nurses. JNCI Cancer Spectr. 2020, 4, 106. [Google Scholar] [CrossRef] [PubMed]
- Talib, W.H.; Alsayed, A.R.; Abuawad, A.; Daoud, S.; Mahmod, A.I. Melatonin in Cancer Treatment: Current Knowledge and Future Opportunities. Molecules 2021, 26, 2506. [Google Scholar] [CrossRef] [PubMed]
- Li, T.; Yang, Z.; Jiang, S.; Di, W.; Ma, Z.; Hu, W.; Chen, F.; Reiter, R.J.; Yang, Y. Melatonin: Does it have utility in the treatment of haematological neoplasms? Br. J. Pharmacol. 2018, 175, 3251–3262. [Google Scholar] [CrossRef] [PubMed]
- Cardinali, D.P.; Escames, G.; Acuña Castroviejo, D.; Ortiz, F.; Fernández Gil, B.; Guerra Librero, A.; García López, S.; Shen, Y.; Florido, J. Melatonin-Induced Oncostasis, Mechanisms and Clinical Relevance. J. Integr. Oncol. 2016, S1, 006. [Google Scholar] [CrossRef]
- Zemła, A.; Grzegorek, I.; Dzięgiel, P.; Jabłońska, K. Melatonin Synergizes the Chemotherapeutic Effect of Cisplatin in Ovarian Cancer Cells Independently of MT1 Melatonin Receptors. In Vivo 2017, 31, 801–809. [Google Scholar] [CrossRef]
- Casado-Zapico, S.; Rodriguez-Blanco, J.; García-Santos, G.; Martín, V.; Sánchez-Sánchez, A.M.; Antolín, I.; Rodriguez, C. Synergistic antitumor effect of melatonin with several chemotherapeutic drugs on human Ewing sarcoma cancer cells: Potentiation of the extrinsic apoptotic pathway. J. Pineal Res. 2010, 48, 72–80. [Google Scholar] [CrossRef]
- Zhelev, Z.; Ivanova, D.; Bakalova, R.; Aoki, I.; Higashi, T. Synergistic Cytotoxicity of Melatonin and New-generation Anticancer Drugs Against Leukemia Lymphocytes But Not Normal Lymphocytes. Anticancer Res. 2017, 37, 149–159. [Google Scholar] [CrossRef]
- Liu, K.; Song, J.; Yan, Y.; Zou, K.; Che, Y.; Wang, B.; Li, Z.; Yu, W.; Guo, W.; Zou, L.; et al. Melatonin increases the chemosensitivity of diffuse large B-cell lymphoma cells to epirubicin by inhibiting P-glycoprotein expression via the NF-κB pathway. Transl. Oncol. 2021, 14, 100876. [Google Scholar] [CrossRef]
- Srinivasan, V.; Pandi-Perumal, S.R.; Brzezinski, A.; Bhatnagar, K.P.; Cardinali, D.P. Melatonin, immune function and cancer. Recent. Pat. Endocr. Metab. Immune Drug Discov. 2011, 5, 109–123. [Google Scholar] [CrossRef]
- Pfeffer, C.M.; Singh, A.T. Apoptosis: A Target for Anticancer Therapy. Int. J. Mol. Sci. 2018, 19, 448. [Google Scholar] [CrossRef]
- Sainz, R.M.; Mayo, J.C.; Rodriguez, C.; Tan, D.X.; Lopez-Burillo, S.; Reiter, R.J. Melatonin and cell death: Differential actions on apoptosis in normal and cancer cells. Cell. Mol. Life Sci. 2003, 60, 1407–1426. [Google Scholar] [CrossRef] [PubMed]
- Rubio, S.; Estévez, F.; Cabrera, J.; Reiter, R.J.; Loro, J.; Quintana, J. Inhibition of proliferation and induction of apoptosis by melatonin in human myeloid HL-60 cells. J. Pineal Res. 2007, 42, 131–138. [Google Scholar] [CrossRef] [PubMed]
- Paternoster, L.; Radogna, F.; Accorsi, A.; Cristina Albertini, M.; Gualandi, G.; Ghibelli, L. Melatonin as a modulator of apoptosis in B-lymphoma cells. Ann. N. Y. Acad. Sci. 2009, 1171, 345–349. [Google Scholar] [CrossRef] [PubMed]
- Trubiani, O.; Recchioni, R.; Moroni, F.; Pizzicannella, J.; Caputi, S.; Di Primio, R. Melatonin provokes cell death in human B-lymphoma cells by mitochondrial-dependent apoptotic pathway activation. J. Pineal Res. 2005, 39, 425–431. [Google Scholar] [CrossRef] [PubMed]
- Casado-Zapico, S.; Martín, V.; García-Santos, G.; Rodríguez-Blanco, J.; Sánchez-Sánchez, A.M.; Luño, E.; Suárez, C.; García-Pedrero, J.M.; Menendez, S.T.; Antolin, I.; et al. Regulation of the expression of death receptors and their ligands by melatonin in haematological cancer cell lines and in leukaemia cells from patients. J. Pineal Res. 2011, 50, 345–355. [Google Scholar] [CrossRef]
- Sánchez-Hidalgo, M.; Lee, M.; De La Lastra, C.A.; Guerrero, J.M.; Packham, G. Melatonin inhibits cell proliferation and induces caspase activation and apoptosis in human malignant lymphoid cell lines. J. Pineal Res. 2012, 53, 366–373. [Google Scholar] [CrossRef]
- Büyükavcı, M.; Özdemir, Ö.; Buck, S.; Stout, M.; Ravindranath, Y.; Savaşan, S. Melatonin cytotoxicity in human leukemia cells: Relation with its pro-oxidant effect. Fundam. Clin. Pharmacol. 2006, 20, 73–79. [Google Scholar] [CrossRef]
- Sanchez-Hidalgo, M.; Lee, M.; De La Lastra, C.A.; Guerrero, J.M.; Packham, G. Melatonin inhibits cell proliferation and induces apoptosis in human acute leukemia T and lymphoma B cells via a receptor independent mechanism. Cancer Res. 2010, 70, 3662. [Google Scholar] [CrossRef]
- Sherr, C.J. Cancer cell cycles. Science 1996, 274, 1672–1677. [Google Scholar] [CrossRef]
- Zhou, M.; Wang, Y.; Shen, J.; Fei, G. Melatonin Induces Cell Cycle Arrest, Inhibits Cell Proliferation and Accelerates Apoptosis by Modulation of CDK4 in NSCLC. Res. Sq. 2022. [Google Scholar] [CrossRef]
- Shen, C.J.; Chang, C.C.; Chen, Y.T.; Lai, C.S.; Hsu, Y.C. Melatonin Suppresses the Growth of Ovarian Cancer Cell Lines (OVCAR-429 and PA-1) and Potentiates the Effect of G1 Arrest by Targeting CDKs. Int. J. Mol. Sci. 2016, 17, 176. [Google Scholar] [CrossRef] [PubMed]
- Song, J.; Ma, S.J.; Luo, J.H.; Zhang, H.; Wang, R.X.; Liu, H.; Li, L.; Zhang, Z.G.; Zhou, R.X. Melatonin induces the apoptosis and inhibits the proliferation of human gastric cancer cells via blockade of the AKT/MDM2 pathway. Oncol. Rep. 2018, 39, 1975–1983. [Google Scholar] [CrossRef] [PubMed]
- Liu, L.; Xu, Y.; Reiter, R.J.; Pan, Y.; Chen, D.; Liu, Y.; Pu, X.; Jiang, L.; Li, Z. Inhibition of ERK1/2 Signaling Pathway is Involved in Melatonin’s Antiproliferative Effect on Human MG-63 Osteosarcoma Cells. Cell. Physiol. Biochem. 2016, 39, 2297–2307. [Google Scholar] [CrossRef] [PubMed]
- Martín-Renedo, J.; Mauriz, J.L.; Jorquera, F.; Ruiz-Andrés, O.; González, P.; González-Gallego, J. Melatonin induces cell cycle arrest and apoptosis in hepatocarcinoma HepG2 cell line. J. Pineal Res. 2008, 45, 532–540. [Google Scholar] [CrossRef]
- García-Santos, G.; Antolín, I.; Herrera, F.; Martín, V.; Rodriguez-Blanco, J.; Carrera, M.D.P.; Rodriguez, C. Melatonin induces apoptosis in human neuroblastoma cancer cells. J. Pineal Res. 2006, 41, 130–135. [Google Scholar] [CrossRef]
- Kasi, R.; Yeo, P.L.; Yen, N.K.; Koh, R.Y.; Ponnudurai, G.; Tiong, Y.L.; Chye, S.M. Melatonin Induces Apoptosis and Inhibits the Proliferation of Cancer Cells via Reactive Oxygen Species-mediated MAPK and mTOR Pathways. Clin. Cancer Drugs 2020, 7, 44–56. [Google Scholar] [CrossRef]
- Koşar, P.A.; Naziroğlu, M.; Övey, İ.S.; Çiğ, B. Synergic Effects of Doxorubicin and Melatonin on Apoptosis and Mitochondrial Oxidative Stress in MCF-7 Breast Cancer Cells: Involvement of TRPV1 Channels. J. Membr. Biol. 2016, 249, 129–140. [Google Scholar] [CrossRef] [PubMed]
- Alonso-González, C.; Menéndez-Menéndez, J.; González-González, A.; González, A.; Cos, S.; Martínez-Campa, C. Melatonin enhances the apoptotic effects and modulates the changes in gene expression induced by docetaxel in MCF-7 human breast cancer cells. Int. J. Oncol. 2018, 52, 560–570. [Google Scholar] [CrossRef] [PubMed]
- Di Bella, G.; Colori, B.; Mascia, F. The Di Bella Method (DBM) improved survival, objective response and performance status in a retrospective observational clinical study on 55 cases of lymphomas. Neuroendocrinol. Lett. 2012, 33, 773–781. [Google Scholar]
- Granzotto, M.; Rapozzi, V.; Decorti, G.; Giraldi, T. Effects of melatonin on doxorubicin cytotoxicity in sensitive and pleiotropically resistant tumor cells. J. Pineal Res. 2001, 31, 206–213. [Google Scholar] [CrossRef] [PubMed]
- Rameshknia, V.; Movahhedi, M.; Akhavan, A.; Majidinia, M.; Yousefi, B. Effect of Melatonin on Reversing Multidrug Resistance by Targeting Phosphatase and Tensin Homolog in Leukemia Cancer Cells. Iran. Red Crescent Med. J. 2021, 23, e212. [Google Scholar] [CrossRef]
- Reiter, R.J.; Tan, D.X.; Sainz, R.M.; Mayo, J.C.; Lopez-Burillo, S. Melatonin: Reducing the toxicity and increasing the efficacy of drugs. J. Pharm. Pharmacol. 2002, 54, 1299–1321. [Google Scholar] [CrossRef] [PubMed]
- Gamen, S.; Anel, A.; Pérez-Galán, P.; Lasierra, P.; Johnson, D.; Piñeiro, A.; Naval, J. Doxorubicin treatment activates a Z-VAD-sensitive caspase, which causes deltapsim loss, caspase-9 activity, and apoptosis in Jurkat cells. Exp. Cell Res. 2000, 258, 223–235. [Google Scholar] [CrossRef] [PubMed]
- Liu, W.; Zhao, Z.; Na, Y.; Meng, C.; Wang, J.; Bai, R. Dexamethasone-induced production of reactive oxygen species promotes apoptosis via endoplasmic reticulum stress and autophagy in MC3T3-E1 cells. Int. J. Mol. Med. 2018, 41, 2028–2036. [Google Scholar] [CrossRef] [PubMed]

Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Mańka, S.; Smolewski, P.; Cebula-Obrzut, B.; Majchrzak, A.; Szmejda, K.; Witkowska, M. Cytotoxic Activity of Melatonin Alone and in Combination with Doxorubicin and/or Dexamethasone on Diffuse Large B-Cell Lymphoma Cells in In Vitro Conditions. J. Pers. Med. 2023, 13, 1314. https://doi.org/10.3390/jpm13091314
Mańka S, Smolewski P, Cebula-Obrzut B, Majchrzak A, Szmejda K, Witkowska M. Cytotoxic Activity of Melatonin Alone and in Combination with Doxorubicin and/or Dexamethasone on Diffuse Large B-Cell Lymphoma Cells in In Vitro Conditions. Journal of Personalized Medicine. 2023; 13(9):1314. https://doi.org/10.3390/jpm13091314
Chicago/Turabian StyleMańka, Sylwia, Piotr Smolewski, Barbara Cebula-Obrzut, Agata Majchrzak, Klaudia Szmejda, and Magdalena Witkowska. 2023. "Cytotoxic Activity of Melatonin Alone and in Combination with Doxorubicin and/or Dexamethasone on Diffuse Large B-Cell Lymphoma Cells in In Vitro Conditions" Journal of Personalized Medicine 13, no. 9: 1314. https://doi.org/10.3390/jpm13091314
APA StyleMańka, S., Smolewski, P., Cebula-Obrzut, B., Majchrzak, A., Szmejda, K., & Witkowska, M. (2023). Cytotoxic Activity of Melatonin Alone and in Combination with Doxorubicin and/or Dexamethasone on Diffuse Large B-Cell Lymphoma Cells in In Vitro Conditions. Journal of Personalized Medicine, 13(9), 1314. https://doi.org/10.3390/jpm13091314