
Theratyping of the Rare CFTR Variants E193K and R334W in Rectal Organoid-Derived Epithelial Monolayers

 ,
,  , ,
, ,  , , ,
, , , 

Abstract
:1. Introduction
2. Materials and Methods
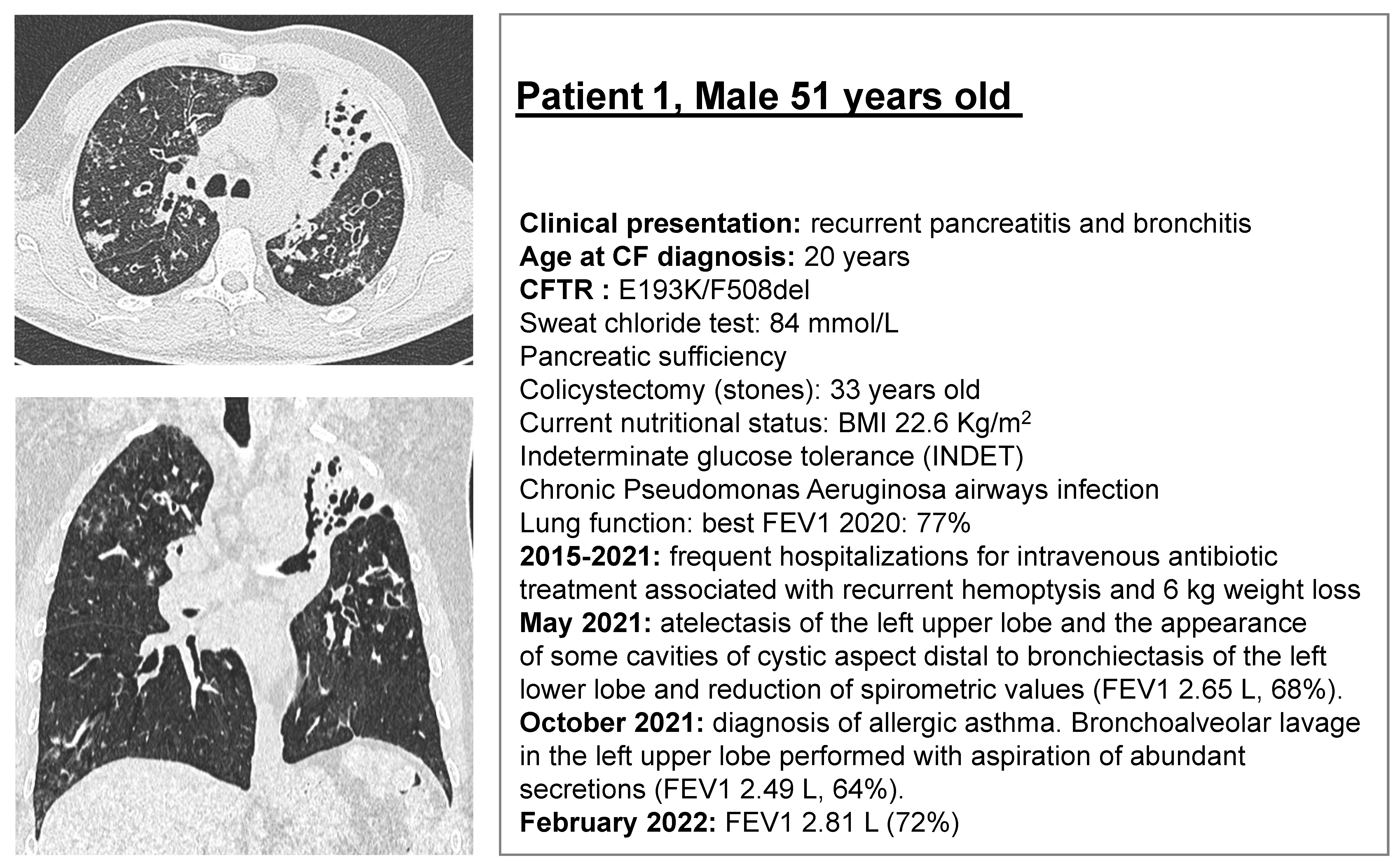
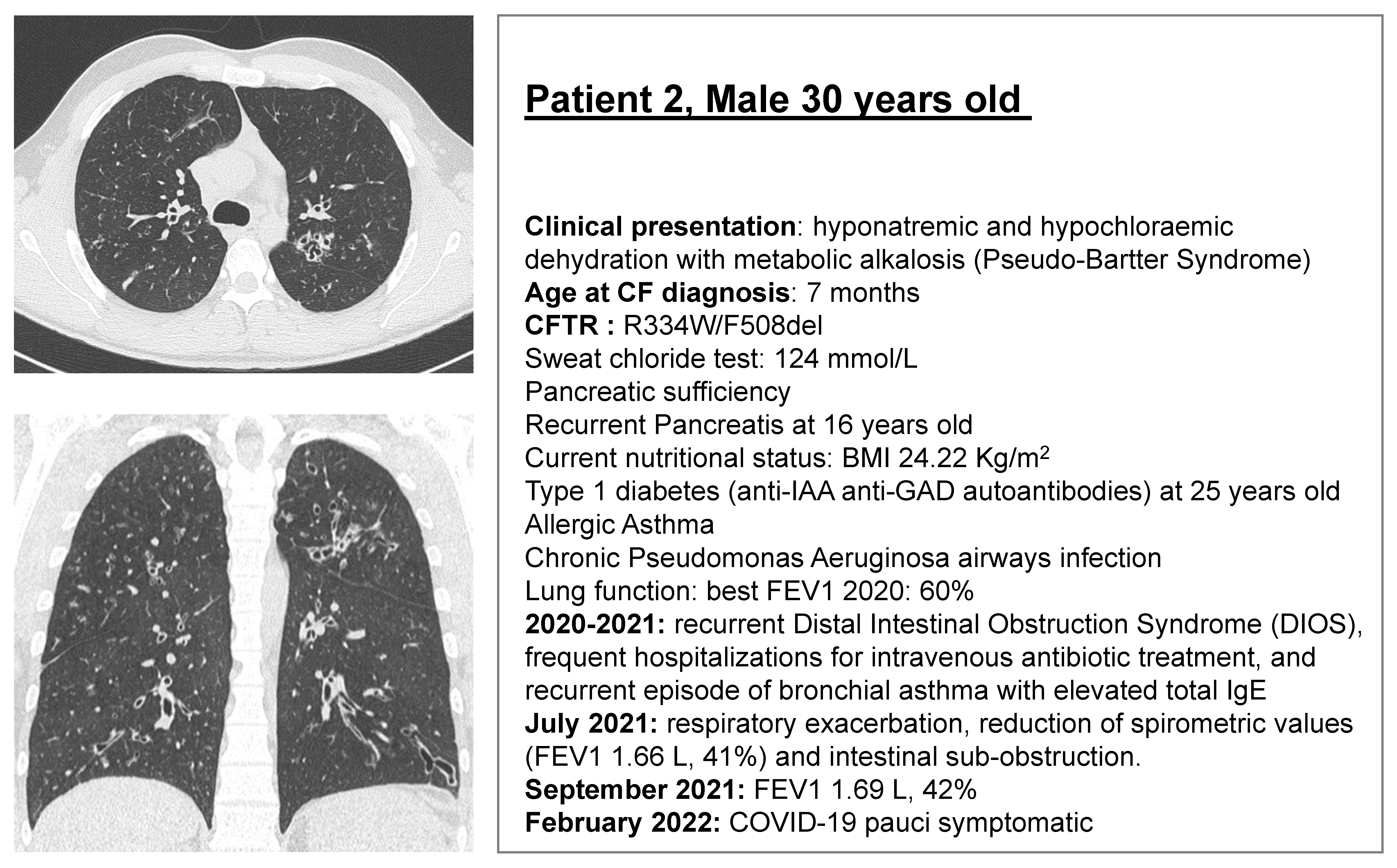
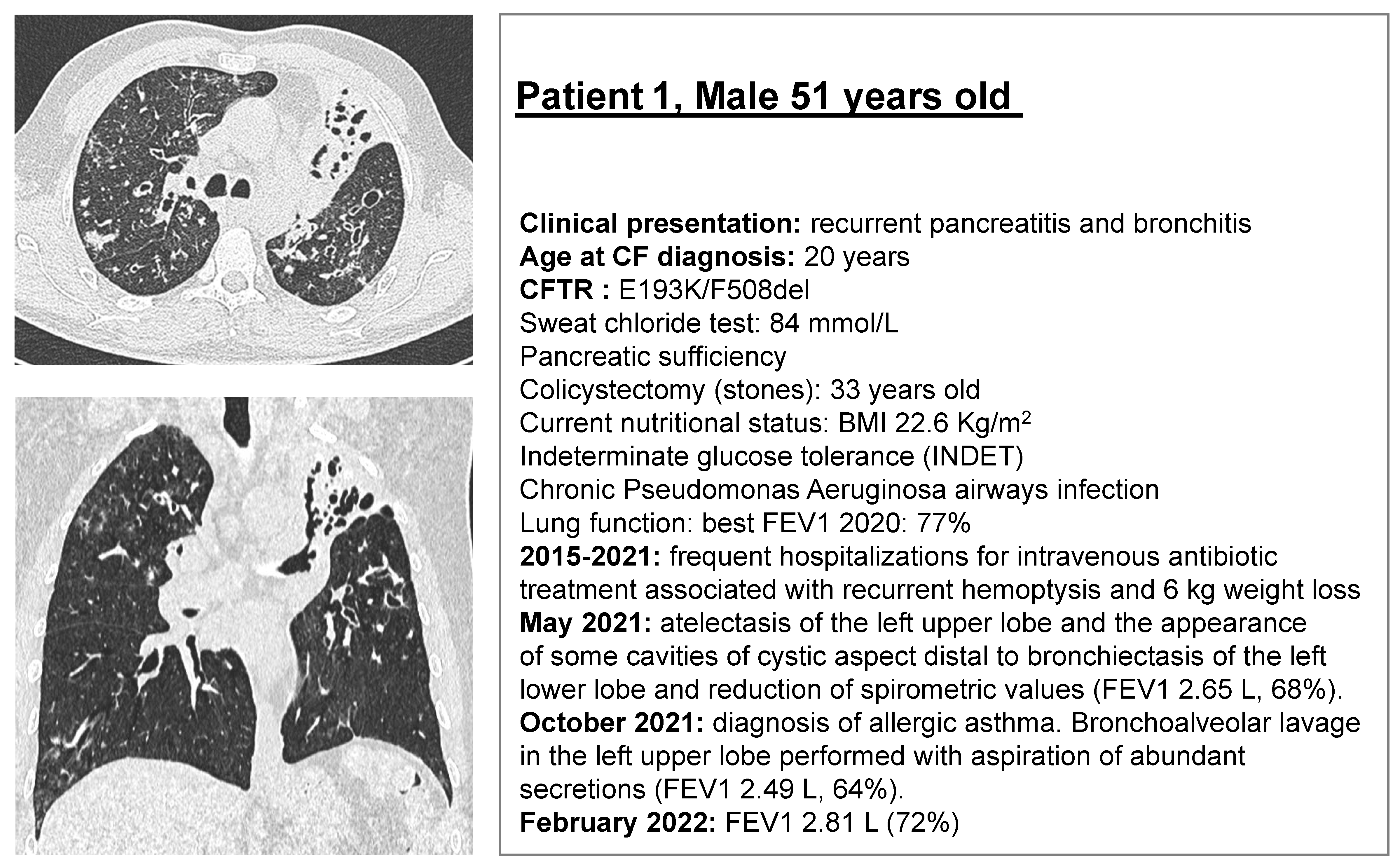
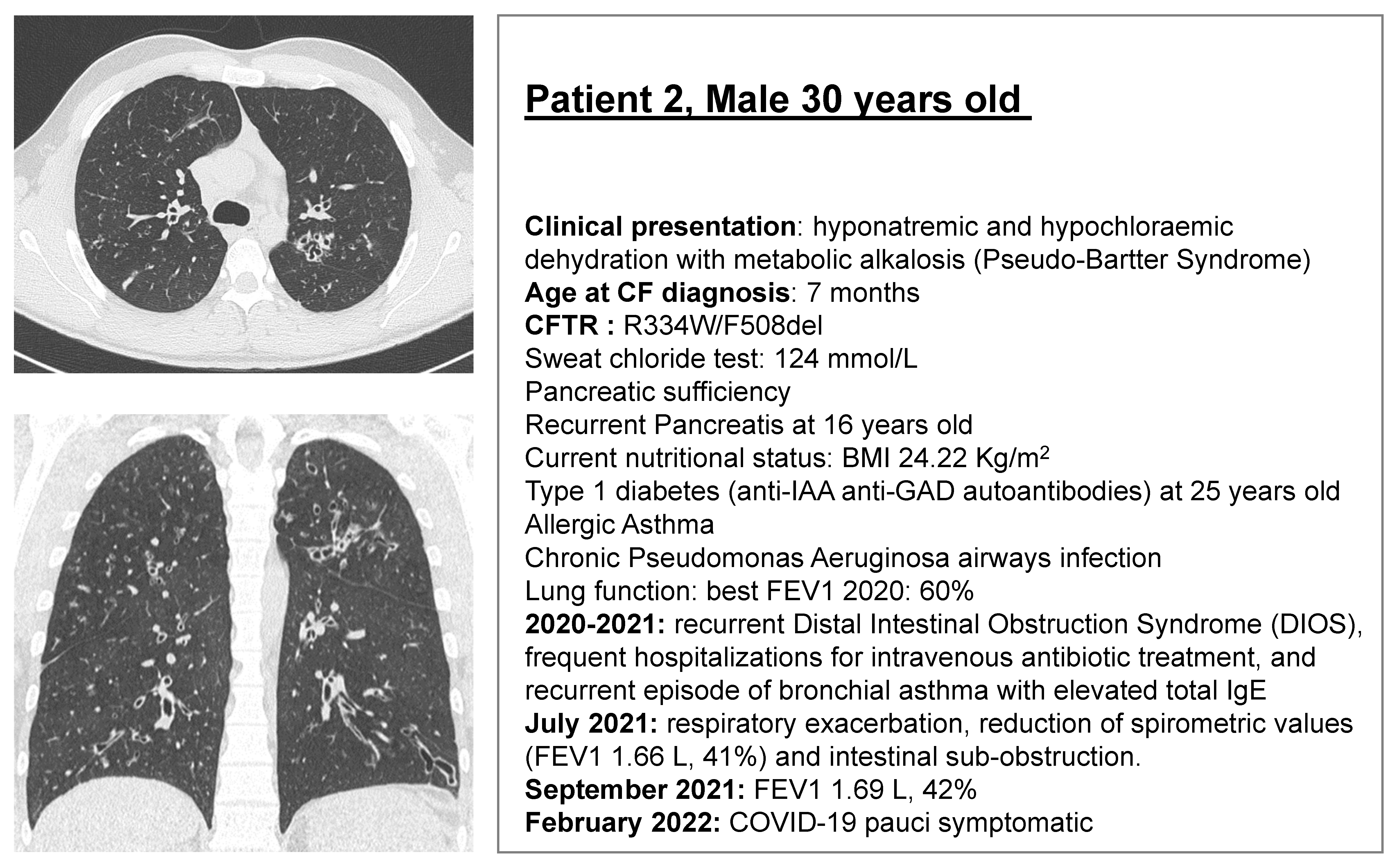
2.1. CF Subjects Selection
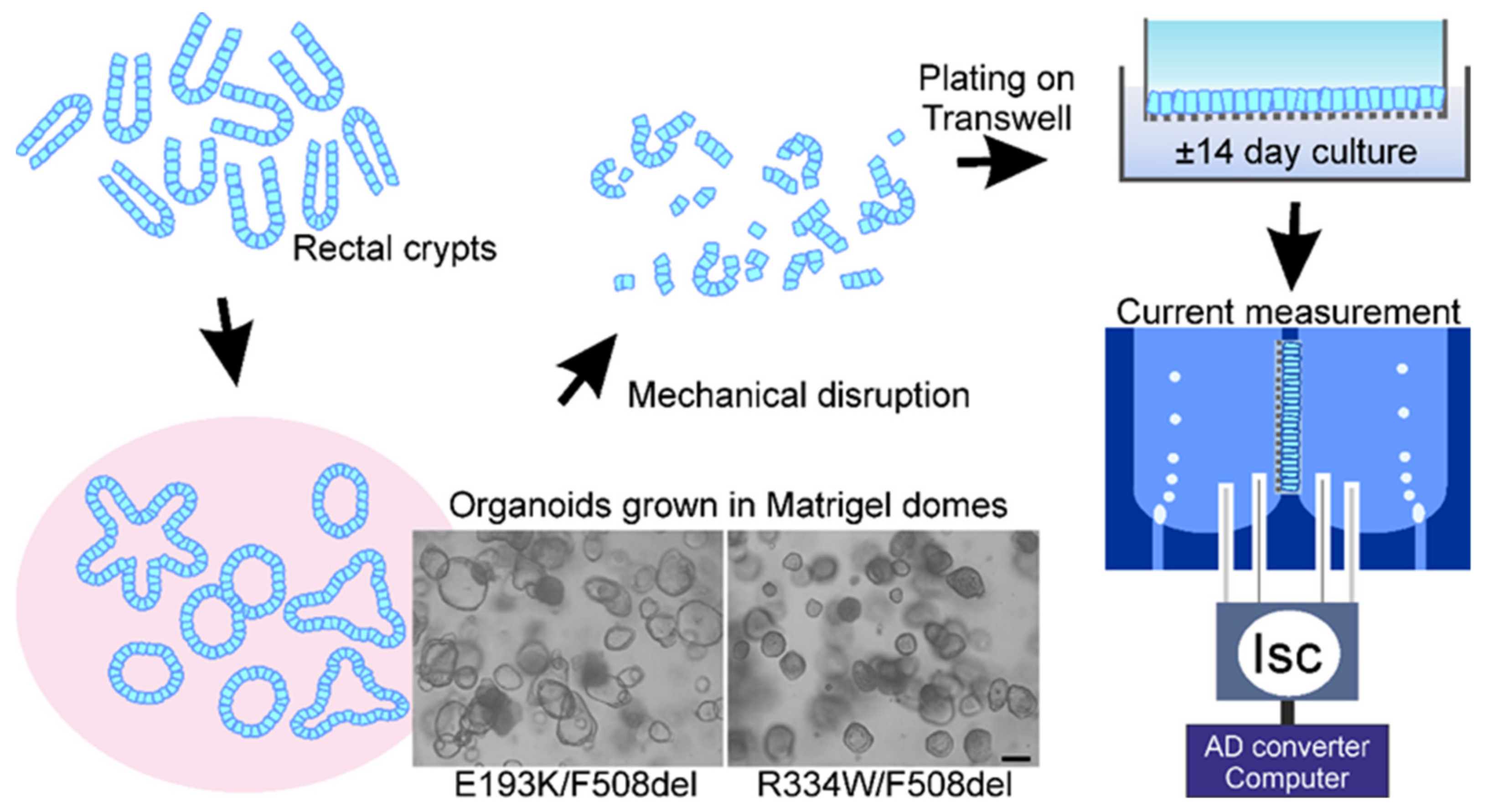
2.2. Rectal Biopsies
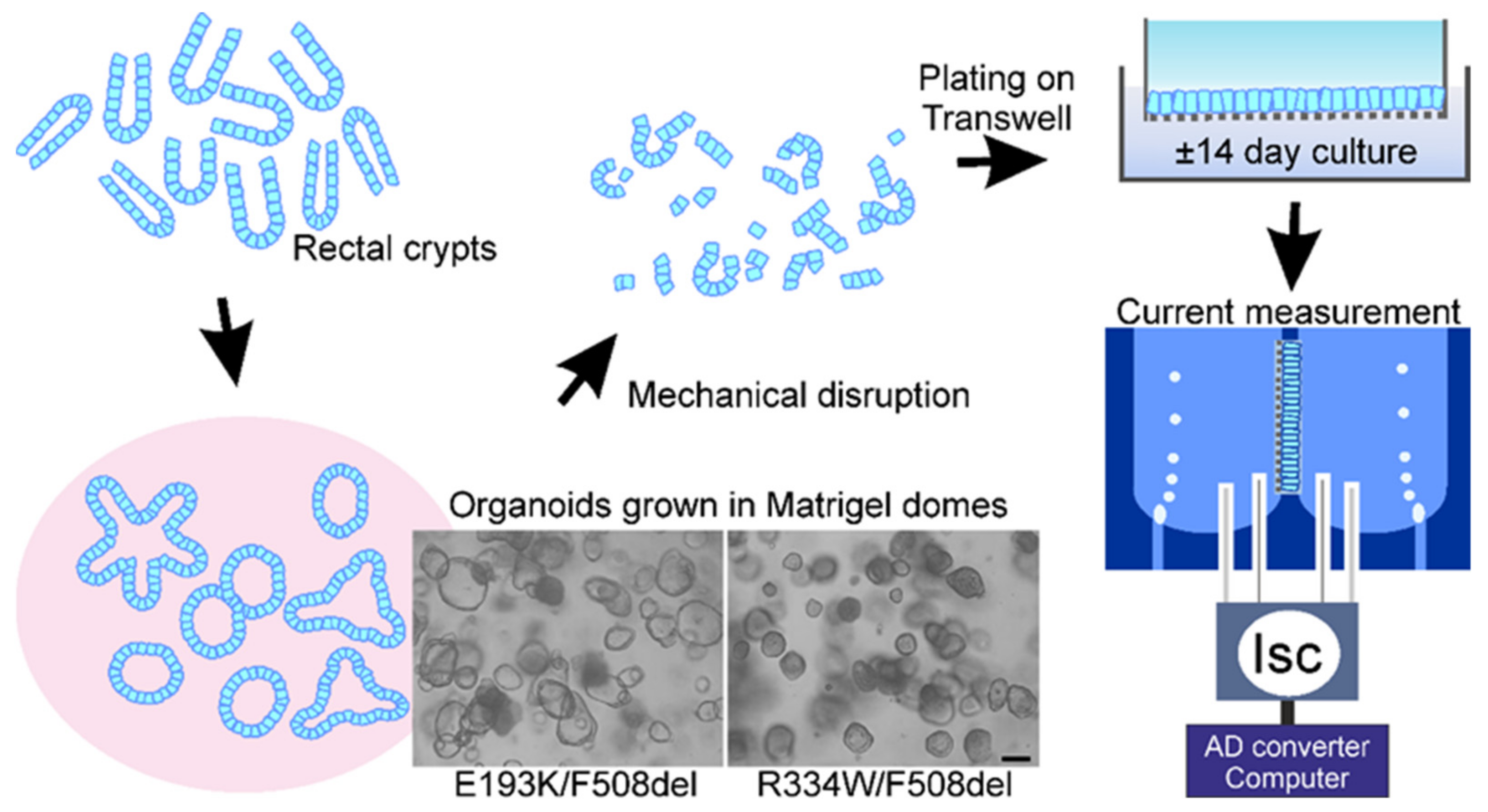
2.3. Organoid Cultures (3D)
2.4. Culture of Epithelial Monolayers of Intestinal Organoids (2D)
2.5. Measurements of Chloride and Bicarbonate Secretory Currents across Organoid Monolayers
2.6. Statistical Analysis
3. Results
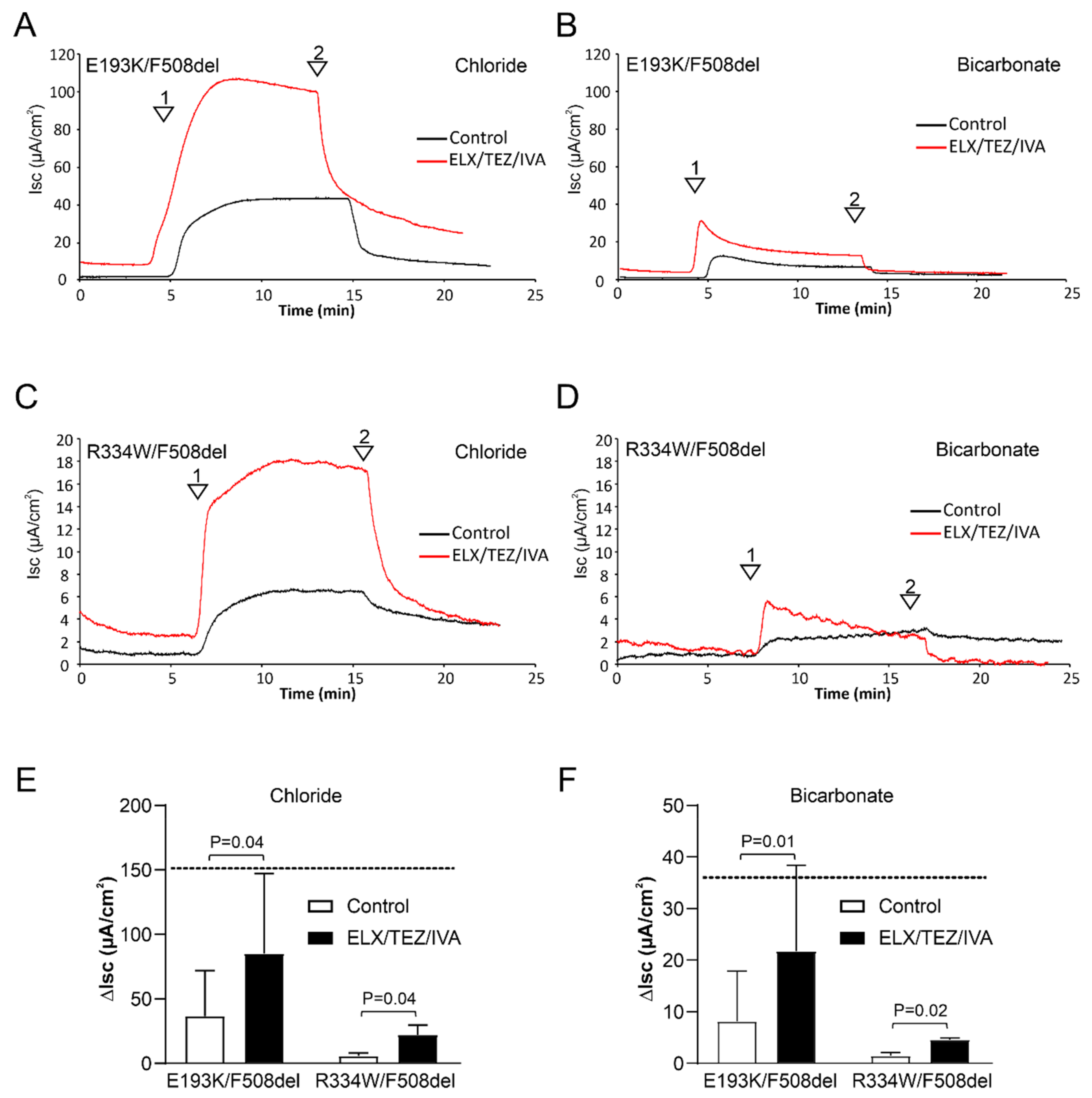
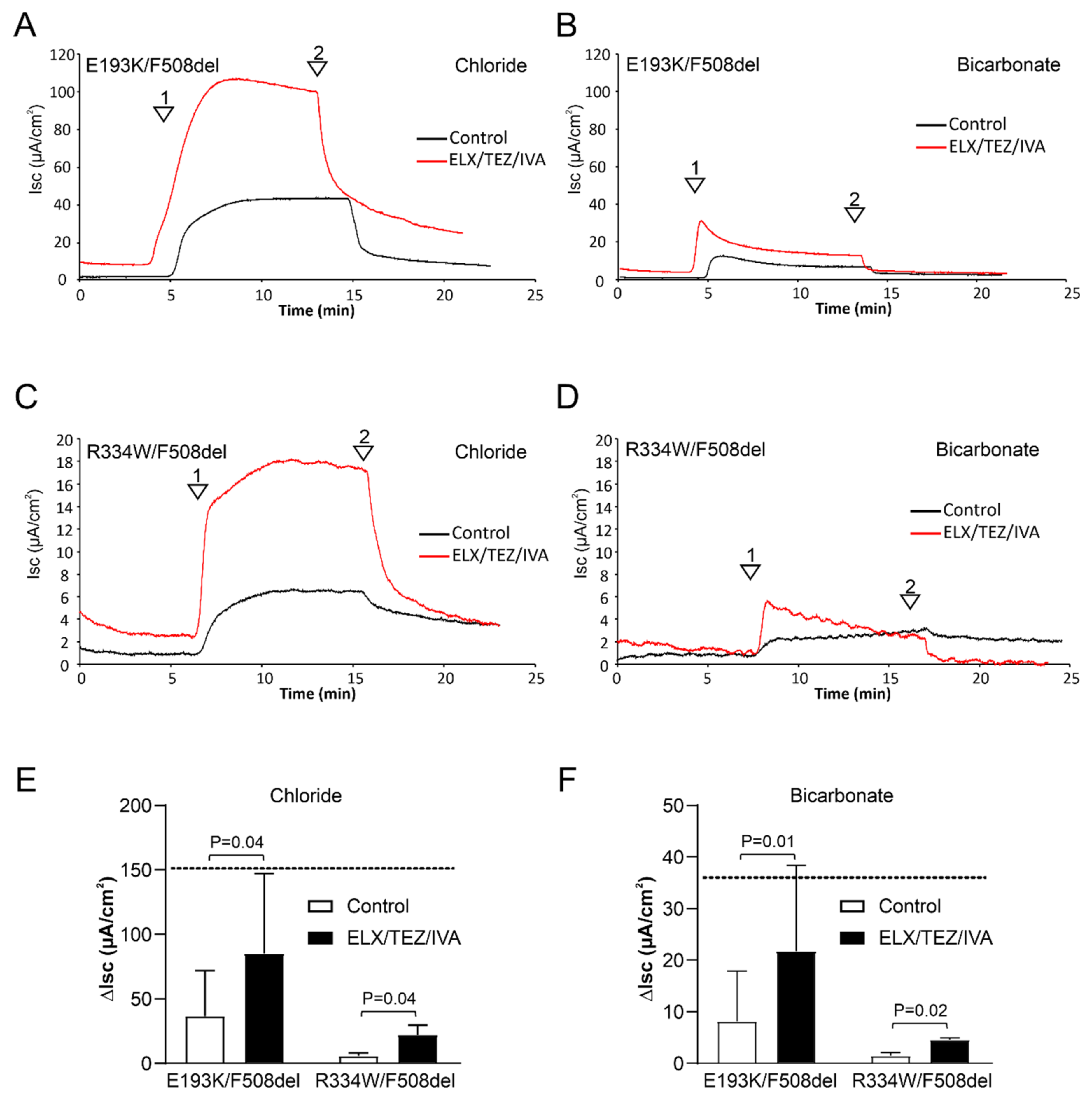
3.1. ELX/TEZ/IVA Rescues CFTR-Mediated Chloride and Bicarbonate Transport across E193K/F508del and R334W/F508del Monolayers
3.2. ELX/TEZ/IVA Is Efficacious in Patient 2 (R334W/F508del)
4. Discussion
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
Abbreviations
References
- Ratjen, F.; Döring, G. Cystic fibrosis. Lancet 2003, 361, 681–689. [Google Scholar] [CrossRef]
- Zolin, A.; Orenti, A.; Nahrlich, L.; Jung, A.; van Rens, J. ECFS Patient Registry Annual Report 2018. Available online: https://www.ecfs.eu/projects/ecfs-patientregistry/annual-reports (accessed on 1 January 2020).
- Lopes-Pacheco, M.; Pedemonte, N.; Veit, G. Discovery of CFTR modulators for the treatment of cystic fibrosis. Expert Opin. Drug Discov. 2021, 16, 897–913. [Google Scholar] [CrossRef]
- Middleton, P.G.; Mall, M.A.; Dřevínek, P.; Lands, L.C.; McKone, E.F.; Polineni, D.; Ramsey, B.W.; Taylor-Cousar, J.L.; Tullis, E.; Vermeulen, F.; et al. Elexacaftor-Tezacaftor-Ivacaftor for Cystic Fibrosis with a Single Phe508del Allele. N. Engl. J. Med. 2019, 381, 1809–1819. [Google Scholar] [CrossRef] [PubMed]
- Ratjen, F.; VanDevanter, D.R. Retracing changes in cystic fibrosis understanding and management over the past twenty years. J. Cyst. Fibros. 2021, 21, 3–9. [Google Scholar] [CrossRef] [PubMed]
- Veit, G.; Velkov, T.; Xu, H.; Vadeboncoeur, N.; Bilodeau, L.; Matouk, E.; Lukacs, G. A Precision Medicine Approach to Optimize Modulator Therapy for Rare CFTR Folding Mutants. J. Pers. Med. 2021, 11, 643. [Google Scholar] [CrossRef] [PubMed]
- Veit, G.; Roldan, A.; Hancock, M.A.; Da Fonte, D.F.; Xu, H.; Hussein, M.; Frenkiel, S.; Matouk, E.; Velkov, T.; Lukacs, G.L. Allosteric folding correction of F508del and rare CFTR mutants by elexacaftor-tezacaftor-ivacaftor (Trikafta) combination. JCI Insight 2020, 5, e139983. [Google Scholar] [CrossRef] [PubMed]
- Laselva, O.; Bartlett, C.; Gunawardena, T.N.; Ouyang, H.; Eckford, P.D.; Moraes, T.J.; Bear, C.E.; Gonska, T. Rescue of multiple class II CFTR mutations by elexacaftor+tezacaftor+ivacaftor mediated in part by the dual activities of elexacaftor as both corrector and potentiator. Eur. Respir. J. 2021, 57, 2002774. [Google Scholar] [CrossRef] [PubMed]
- Laselva, O.; McCormack, J.; Bartlett, C.; Ip, W.; Gunawardena, T.; Ouyang, H.; Eckford, P.; Gonska, T.; Moraes, T.; Bear, C. Preclinical Studies of a Rare CF-Causing Mutation in the Second Nucleotide Binding Domain (c.3700A>G) Show Robust Functional Rescue in Primary Nasal Cultures by Novel CFTR Modulators. J. Pers. Med. 2020, 10, 209. [Google Scholar] [CrossRef]
- Capurro, V.; Tomati, V.; Sondo, E.; Renda, M.; Borrelli, A.; Pastorino, C.; Guidone, D.; Venturini, A.; Giraudo, A.; Bertozzi, S.M.; et al. Partial Rescue of F508del-CFTR Stability and Trafficking Defects by Double Corrector Treatment. Int. J. Mol. Sci. 2021, 22, 5262. [Google Scholar] [CrossRef]
- Shaughnessy, C.A.; Zeitlin, P.L.; Bratcher, P.E. Elexacaftor is a CFTR potentiator and acts synergistically with ivacaftor during acute and chronic treatment. Sci. Rep. 2021, 11, 19810, Erratum in Sci. Rep. 2021, 11, 21295. [Google Scholar] [CrossRef]
- Barry, P.J.; Mall, M.A.; Álvarez, A.; Colombo, C.; Groot, K.M.D.W.-D.; Fajac, I.; McBennett, K.A.; McKone, E.F.; Ramsey, B.W.; Sutharsan, S.; et al. Triple Therapy for Cystic Fibrosis Phe508del–Gating and –Residual Function Genotypes. N. Engl. J. Med. 2021, 385, 815–825. [Google Scholar] [CrossRef]
- de Poel, E.; Lefferts, J.; Beekman, J. Intestinal organoids for Cystic Fibrosis research. J. Cyst. Fibros. 2020, 19, S60–S64. [Google Scholar] [CrossRef] [Green Version]
- Pranke, I.; Hatton, A.; Masson, A.; Flament, T.; Le Bourgeois, M.; Chedevergne, F.; Bailly, C.; Urbach, V.; Hinzpeter, A.; Edelman, A.; et al. Might Brushed Nasal Cells Be a Surrogate for CFTR Modulator Clinical Response? Am. J. Respir. Crit. Care Med. 2019, 199, 123–126. [Google Scholar] [CrossRef]
- Scudieri, P.; Musante, I.; Venturini, A.; Guidone, D.; Genovese, M.; Cresta, F.; Caci, E.; Palleschi, A.; Poeta, M.; Santamaria, F.; et al. Ionocytes and CFTR Chloride Channel Expression in Normal and Cystic Fibrosis Nasal and Bronchial Epithelial Cells. Cells 2020, 9, 2090. [Google Scholar] [CrossRef]
- Barker, N.; Van Es, J.H.; Kuipers, J.; Kujala, P.; Van Den Born, M.; Cozijnsen, M.; Haegebarth, A.; Korving, J.; Begthel, H.; Peters, P.J.; et al. Identification of stem cells in small intestine and colon by marker gene Lgr5. Nature 2007, 449, 1003–1007. [Google Scholar] [CrossRef]
- Dekkers, J.F.; Wiegerinck, C.L.; De Jonge, H.R.; Bronsveld, I.; Janssens, H.M.; De Winter-de Groot, K.M.; Brandsma, A.M.; de Jong, N.W.; Bijvelds, M.J.; Scholte, B.J.; et al. A functional CFTR assay using primary cystic fibrosis intestinal organoids. Nat. Med. 2013, 19, 939–945. [Google Scholar] [CrossRef]
- Dekkers, J.F.; Berkers, G.; Kruisselbrink, E.; Vonk, A.; de Jonge, H.R.; Janssens, H.M.; Bronsveld, I.; van de Graaf, E.A.; Nieuwenhuis, E.E.S.; Houwen, R.H.J.; et al. Characterizing responses to CFTR-modulating drugs using rectal organoids derived from subjects with cystic fibrosis. Sci. Transl. Med. 2016, 8, 344ra84. [Google Scholar] [CrossRef]
- Vonk, A.M.; van Mourik, P.; Ramalho, A.S.; Silva, I.; Statia, M.; Kruisselbrink, E.; Suen, S.W.; Dekkers, J.F.; Vleggaar, F.P.; Houwen, R.H.; et al. Protocol for Application, Standardization and Validation of the Forskolin-Induced Swelling Assay in Cystic Fibrosis Human Colon Organoids. STAR Protoc. 2020, 1, 100019. [Google Scholar] [CrossRef]
- Kozuka, K.; He, Y.; Koo-McCoy, S.; Kumaraswamy, P.; Nie, B.; Shaw, K.; Chan, P.; Leadbetter, M.; He, L.; Lewis, J.G.; et al. Development and Characterization of a Human and Mouse Intestinal Epithelial Cell Monolayer Platform. Stem Cell Rep. 2017, 9, 1976–1990. [Google Scholar] [CrossRef] [Green Version]
- Foulke-Abel, J.; In, J.; Yin, J.; Zachos, N.C.; Kovbasnjuk, O.; Estes, M.K.; de Jonge, H.; Donowitz, M. Human Enteroids as a Model of Upper Small Intestinal Ion Transport Physiology and Pathophysiology. Gastroenterology 2016, 150, 638–649.e8. [Google Scholar] [CrossRef] [Green Version]
- De Jonge, H.R.; Bijvelds, M.J.C.; Strubberg, A.M.; Liu, J.; Clarke, L.L. Organoids as a Model for Intestinal Ion Transport Physiology. In Ion Transport Across Epithelial Tissues and Disease—Ion Channels and Transporters of Epithelia in Health and Disease, 2nd ed.; Hamilton, K.L., Devor, D.C., Eds.; Physiology in Health and Disease Series; Springer: London, UK, 2020; Chapter 1; Volume 2, pp. 1–39. ISBN 9783030553098. [Google Scholar]
- Caputo, A.; Hinzpeter, A.; Caci, E.; Pedemonte, N.; Arous, N.; Di Duca, M.; Zegarra-Moran, O.; Fanen, P.; Galietta, L.J.V. Mutation-Specific Potency and Efficacy of Cystic Fibrosis Transmembrane Conductance Regulator Chloride Channel Potentiators. J. Pharmacol. Exp. Ther. 2009, 330, 783–791. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Van Goor, F.; Yu, H.; Burton, B.; Hoffman, B.J. Effect of ivacaftor on CFTR forms with missense mutations associated with defects in protein processing or function. J. Cyst. Fibros. 2014, 13, 29–36. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Gong, X.; Linsdell, P. Maximization of the rate of chloride conduction in the CFTR channel pore by ion–ion interactions. Arch. Biochem. Biophys. 2004, 426, 78–82. [Google Scholar] [CrossRef] [PubMed]
- Phuan, P.W.; Tan, J.A.; Rivera, A.A.; Zlock, L.; Nielson, D.W.; Finkbeiner, W.E.; Haggie, P.M.; Verkman, A.S. Nanomolar-potency ‘co-potentiator’ therapy for cystic fibrosis caused by a defined subset of minimal function CFTR mutants. Sci. Rep. 2019, 9, 17640. [Google Scholar] [CrossRef]
- Vogel, G.F.; van Rijn, J.; Krainer, I.M.; Janecke, A.R.; Posovszky, C.; Cohen, M.; Searle, C.; Jantchou, P.; Escher, J.C.; Patey, N.; et al. Disrupted apical exocytosis of cargo vesicles causes enteropathy in FHL5 patients with Munc18-2 mutations. JCI Insight 2017, 2, 94564. [Google Scholar] [CrossRef] [Green Version]
- Ramsey, M.L.; Gokun, Y.; Sobotka, L.A.; Wellner, M.R.; Porter, K.; Kirkby, S.E.; Li, S.S.; Papachristou, G.I.; Krishna, S.G.; Stanich, P.P.; et al. Cystic Fibrosis Transmembrane Conductance Regulator Modulator Use Is Associated With Reduced Pancreatitis Hospitalizations in Patients With Cystic Fibrosis. Am. J. Gastroenterol. 2021, 116, 2446–2454. [Google Scholar] [CrossRef]
- Keating, D.; Marigowda, G.; Burr, L.D.; Daines, C.; Mall, M.A.; McKone, E.F.; Ramsey, B.W.; Rowe, S.M.; Sass, L.A.; Tullis, E.; et al. VX-445–Tezacaftor–Ivacaftor in Patients with Cystic Fibrosis and One or Two Phe508del Alleles. N. Engl. J. Med. 2018, 379, 1612–1620. [Google Scholar] [CrossRef]
- Veit, G.; Vaccarin, C.; Lukacs, G.L. Elexacaftor co-potentiates the activity of F508del and gating mutants of CFTR. J. Cyst. Fibros. 2021, 20, 895–898. [Google Scholar] [CrossRef]
- Griese, M.; Costa, S.; Linnemann, R.W.; Mall, M.A.; McKone, E.F.; Polineni, D.; Quon, B.S.; Ringshausen, F.C.; Taylor-Cousar, J.L.; Withers, N.J.; et al. Safety and Efficacy of Elexacaftor/Tezacaftor/Ivacaftor for 24 Weeks or Longer in People with Cystic Fibrosis and One or More F508del Alleles: Interim Results of an Open-Label Phase 3 Clinical Trial. Am. J. Respir. Crit. Care Med. 2021, 203, 381–385. [Google Scholar] [CrossRef]
- Heijerman, H.G.M.; McKone, E.F.; Downey, D.G.; Van Braeckel, E.; Rowe, S.M.; Tullis, E.; Mall, M.A.; Welter, J.J.; Ramsey, B.W.; McKee, C.M.; et al. Efficacy and safety of the elexacaftor plus tezacaftor plus ivacaftor combination regimen in people with cystic fibrosis homozygous for the F508del mutation: A double-blind, randomised, phase 3 trial. Lancet 2019, 394, 1940–1948. [Google Scholar] [CrossRef]
- Clancy, J.P.; Cotton, C.U.; Donaldson, S.H.; Solomon, G.M.; VanDevanter, D.R.; Boyle, M.P.; Gentzsch, M.; Nick, J.A.; Illek, B.; Wallenburg, J.C.; et al. CFTR modulator theratyping: Current status, gaps and future directions. J. Cyst. Fibros. 2019, 18, 22–34. [Google Scholar] [CrossRef] [Green Version]
- Van Mourik, P.; Michel, S.; Vonk, A.M.; Beekman, J.M.; van der Ent, C.K.; De Keyser, H.; Lammertyn, E.; van Koningsbruggen-Rietschel, S.; Naehrlich, L.; Pool, J.; et al. Rationale and design of the HIT-CF organoid study: Stratifying cystic fibrosis patients based on intestinal organoid response to different CFTR-modulators. Transl. Med. Commun. 2020, 5, 1–8. [Google Scholar] [CrossRef]
- Aalbers, B.L.; Brunsveld, J.E.; van der Ent, C.K.; van den Eijnden, J.C.; Beekman, J.M.; Heijerman, H.G.M. Forskolin induced swelling (FIS) assay in intestinal organoids to guide eligibility for compassionate use treatment in a CF patient with a rare genotype. J. Cyst. Fibros. 2022, 21, 254–257. [Google Scholar] [CrossRef]
- Pedemonte, N. Nasal epithelial cells as a gold-standard predictive model for personalized medicine in cystic fibrosis. J. Physiol. 2022, 600, 1285–1286. [Google Scholar] [CrossRef]
- Fajac, I.; Sermet, I. Therapeutic Approaches for Patients with Cystic Fibrosis Not Eligible for Current CFTR Modulators. Cells 2021, 10, 2793. [Google Scholar] [CrossRef]
- Bijvelds, M.J.; Roos, F.J.; Meijsen, K.F.; Roest, H.P.; Verstegen, M.M.; Janssens, H.M.; van der Laan, L.J.; de Jonge, H.R. Rescue of chloride and bicarbonate transport by elexacaftor-ivacaftor-tezacaftor in organoid-derived CF intestinal and cholangiocyte monolayers. J. Cyst. Fibros. 2021, in press. [Google Scholar] [CrossRef]
- Hanssens, L.S.; Duchateau, J.; Casimir, G.J. CFTR Protein: Not Just a Chloride Channel? Cells 2021, 10, 2844. [Google Scholar] [CrossRef]

{kind=link}
{kind=link}
{kind=link}
{kind=link}
| A | Non-CF | |||||||||
| Meyler | Bicarbonate | Chloride | ||||||||
| n | Mean | SD | n | Mean | SD | n | Mean | SD | ||
| DMSO | ΔIsc Forskolin | 3 | 189.3 | 73.7 | 3 | 36.8 | 28.2 | 3 | 151.6 | 60.6 |
| B | E193K/F508del | |||||||||
| Meyler | Bicarbonate | Chloride | ||||||||
| n | Mean | SD | n | Mean | SD | n | Mean | SD | ||
| DMSO | ΔIsc Forskolin | 6 | 50.3 | 54.7 | 6 | 8.2 | 9.7 | 6 | 36.6 | 35.3 |
| ΔIsc CFTRinh172 | 6 | −35.3 | 39.2 | 6 | −3.0 | 2.2 | 6 | −25.3 | 22.9 | |
| ELX/TEZ/IVA | ΔIsc Forskolin | 6 | 84.7 | 50.6 | 6 | 21.7 | 16.7 | 6 | 85.0 | 62.1 |
| ΔIsc CFTRinh172 | 6 | −71.6 | 43.6 | 6 | −11.1 | 6.4 | 6 | −67.9 | 51.4 | |
| C | R334W/F508del | |||||||||
| Meyler | Bicarbonate | Chloride | ||||||||
| n | Mean | SD | n | Mean | SD | n | Mean | SD | ||
| DMSO | ΔIsc Forskolin | 3 | 6.1 | 2.5 | 4 | 1.3 | 1.4 | 4 | 6.1 | 1.9 |
| ΔIsc CFTRinh172 | 3 | −2.9 | 1.3 | 4 | −0.7 | 0.5 | 4 | −3.9 | 1.5 | |
| ELX/TEZ/IVA | ΔIsc Forskolin | 3 | 16.6 | 10.6 | 3 | 4.6 | 0.3 | 3 | 22.1 | 7.6 |
| ΔIsc CFTRinh172 | 3 | −17.1 | 8.1 | 3 | −3.2 | 2.2 | 3 | −19.4 | 6.4 | |
| D | G542X/F508del | |||||||||
| Meyler | Bicarbonate | Chloride | ||||||||
| n | Mean | SD | n | Mean | SD | n | Mean | SD | ||
| DMSO | ΔIsc Forskolin | 4 | 0.4 | 0.6 | 4 | 0.6 | 0.7 | 4 | 2.2 | 3.2 |
| ΔIsc CFTRinh172 | 4 | 0.1 | 0.4 | 4 | 0.3 | 1.0 | 4 | −1.9 | 3.2 | |
| ELX/TEZ/IVA | ΔIsc Forskolin | 4 | 8.0 | 6.2 | 4 | 3.1 | 2.0 | 4 | 11.4 | 6.9 |
| ΔIsc CFTRinh172 | 4 | −8.0 | 5.6 | 4 | −1.7 | 0.9 | 4 | −10.7 | 5.9 | |
| Patient 2 (R334W/F508del) | Chloride % of Non-CF in HIOs | Bicarbonate % of Non-CF in HIOs | Sweat Chloride (mmol/L) | FEV1 |
|---|---|---|---|---|
| Baseline | 4 | 4 | 124 | 1.69 L, 42% |
| + ELX/TEZ/IVA (Trikafta) | 13 | 15 | 77 | 2.37 L, 58% |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Ciciriello, F.; Bijvelds, M.J.C.; Alghisi, F.; Meijsen, K.F.; Cristiani, L.; Sorio, C.; Melotti, P.; Fiocchi, A.G.; Lucidi, V.; De Jonge, H.R. Theratyping of the Rare CFTR Variants E193K and R334W in Rectal Organoid-Derived Epithelial Monolayers. J. Pers. Med. 2022, 12, 632. https://doi.org/10.3390/jpm12040632
Ciciriello F, Bijvelds MJC, Alghisi F, Meijsen KF, Cristiani L, Sorio C, Melotti P, Fiocchi AG, Lucidi V, De Jonge HR. Theratyping of the Rare CFTR Variants E193K and R334W in Rectal Organoid-Derived Epithelial Monolayers. Journal of Personalized Medicine. 2022; 12(4):632. https://doi.org/10.3390/jpm12040632
Chicago/Turabian StyleCiciriello, Fabiana, Marcel J. C. Bijvelds, Federico Alghisi, Kelly F. Meijsen, Luca Cristiani, Claudio Sorio, Paola Melotti, Alessandro G. Fiocchi, Vincenzina Lucidi, and Hugo R. De Jonge. 2022. "Theratyping of the Rare CFTR Variants E193K and R334W in Rectal Organoid-Derived Epithelial Monolayers" Journal of Personalized Medicine 12, no. 4: 632. https://doi.org/10.3390/jpm12040632
APA StyleCiciriello, F., Bijvelds, M. J. C., Alghisi, F., Meijsen, K. F., Cristiani, L., Sorio, C., Melotti, P., Fiocchi, A. G., Lucidi, V., & De Jonge, H. R. (2022). Theratyping of the Rare CFTR Variants E193K and R334W in Rectal Organoid-Derived Epithelial Monolayers. Journal of Personalized Medicine, 12(4), 632. https://doi.org/10.3390/jpm12040632