
Gut-Brain Axis Cross-Talk and Limbic Disorders as Biological Basis of Secondary TMAU

 ,
, 
 ,
, 

Abstract
1. Introduction
2. Materials and Methods
2.1. Subjects
2.2. DNA Extraction and Sequencing
2.3. Statistical Analysis
2.4. Neurotransmission Pathway Analysis of Gut-Brain Axis
3. Results
3.1. Microbiota of Neuro-Disordered TMAU Patients Revealed Huge Differences in Composition and Relative Abundances If Compared with “Brain-Healthy” TMAU Affected Individuals
3.2. Altered Bacterial Families of Neuro-Disordered TMAU Patients’ Microbiomes Produce Neurotransmitters and/or a Wide Range of Metabolites Involved in Their Biochemical Pathways
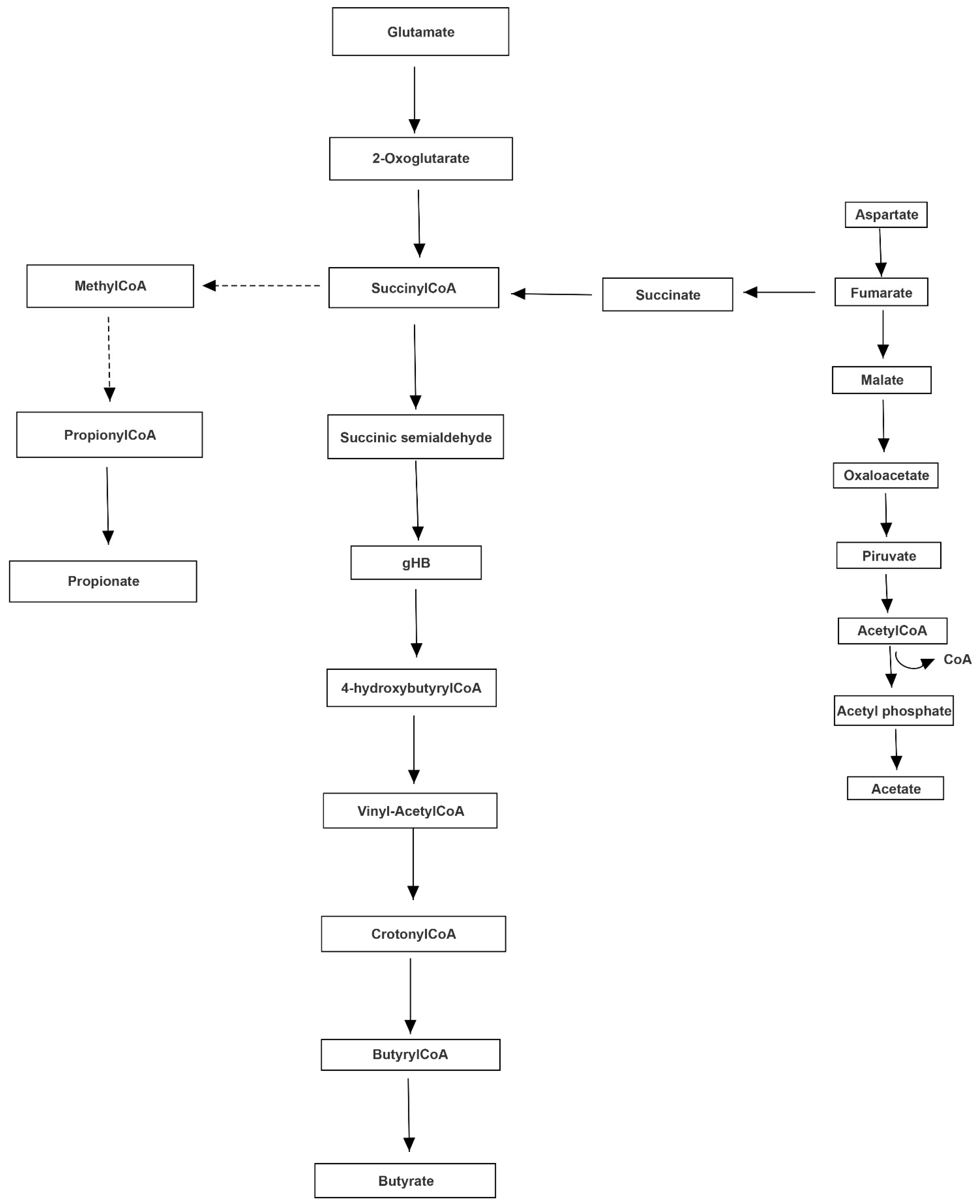
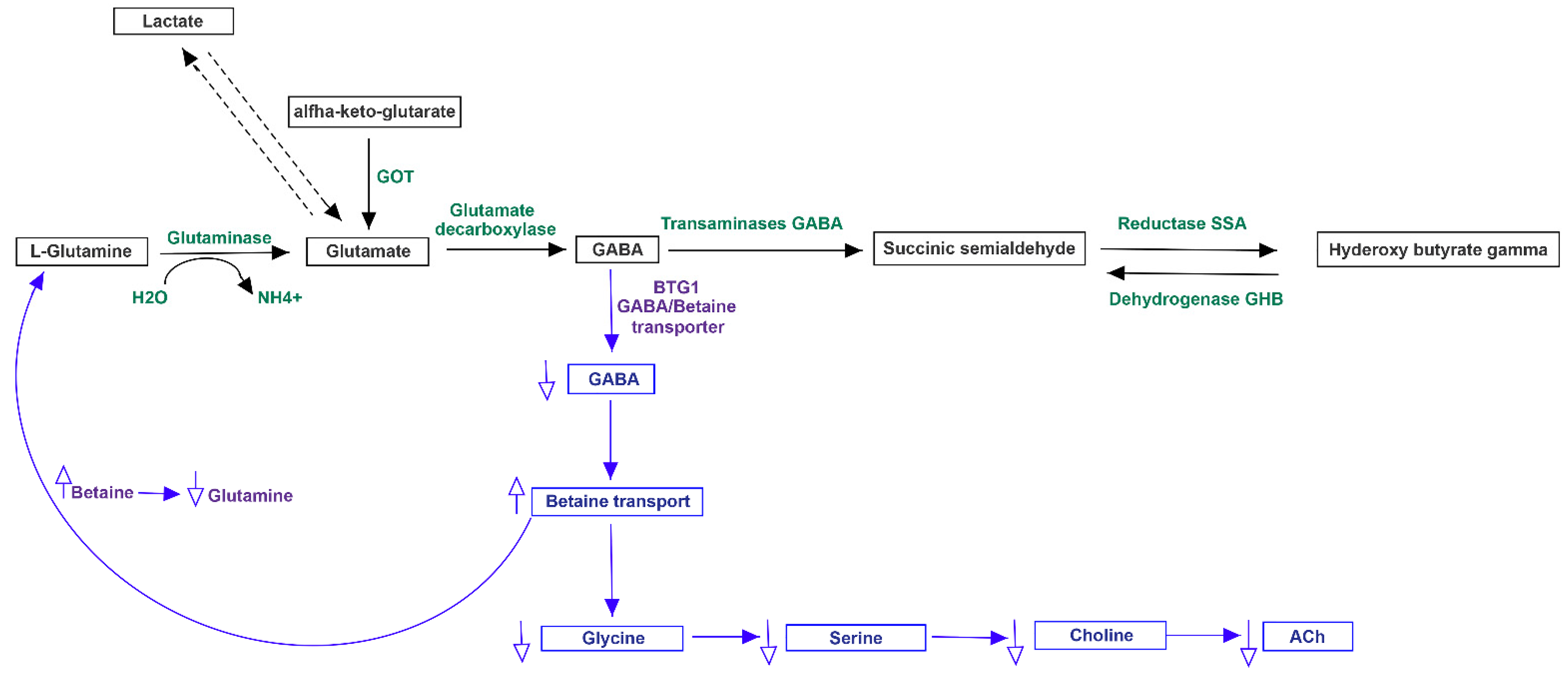
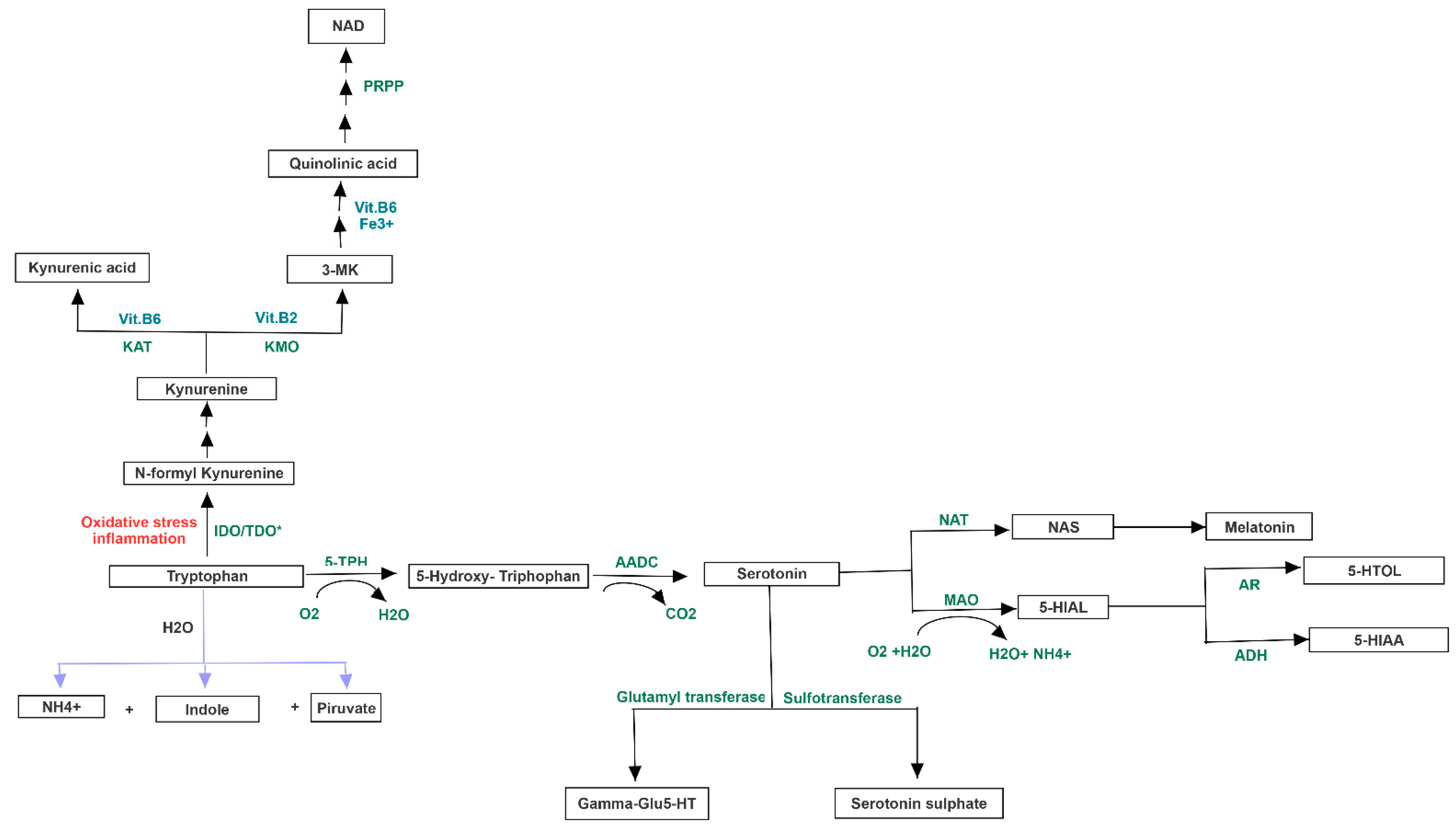
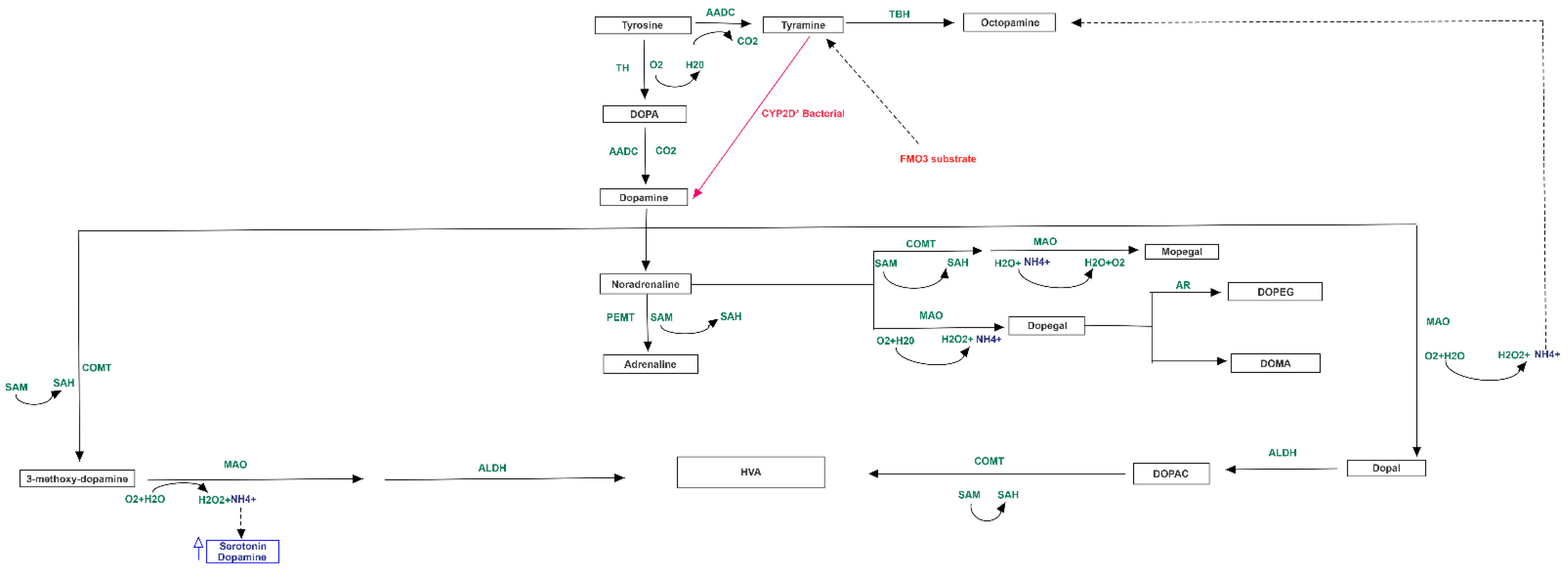
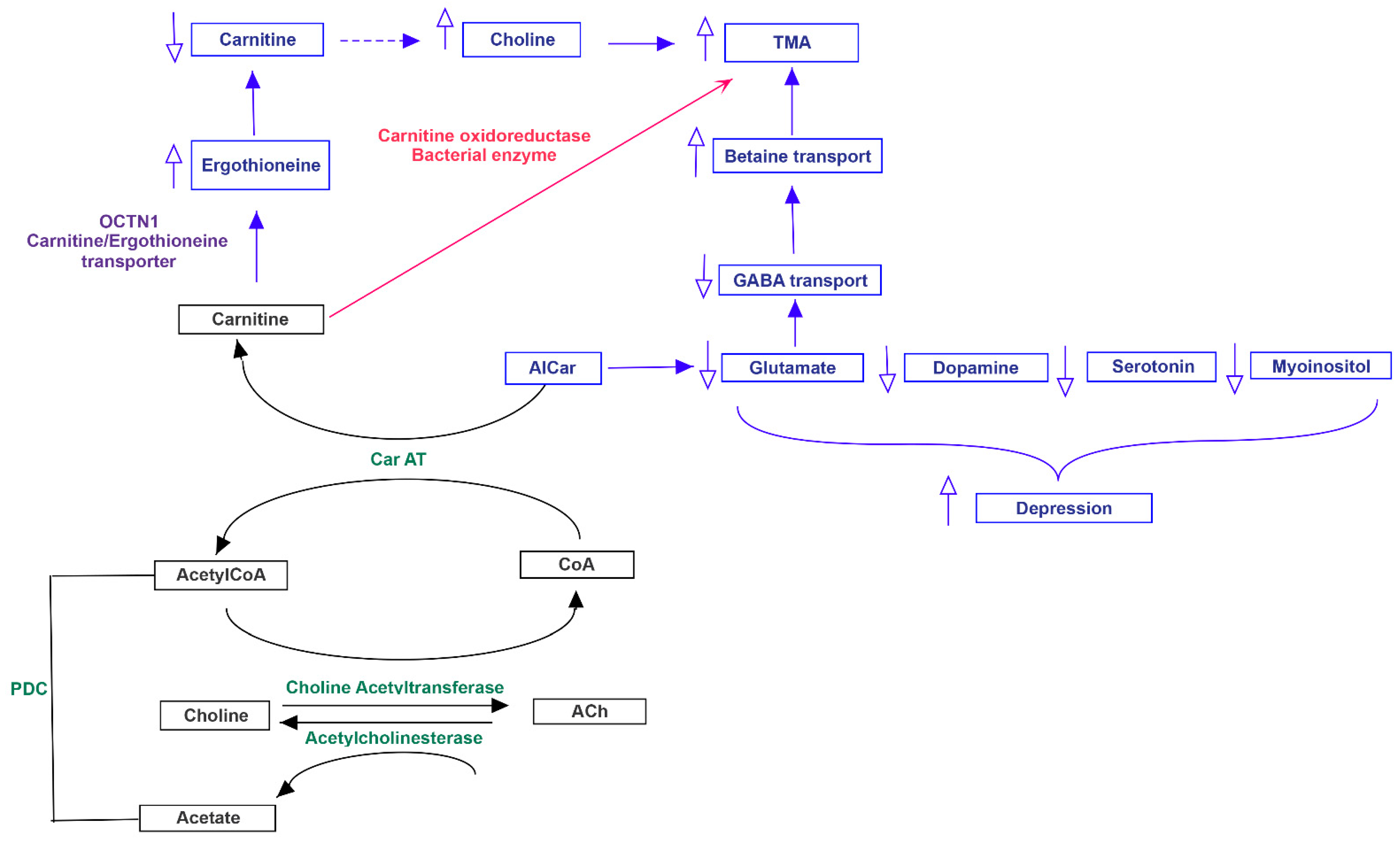
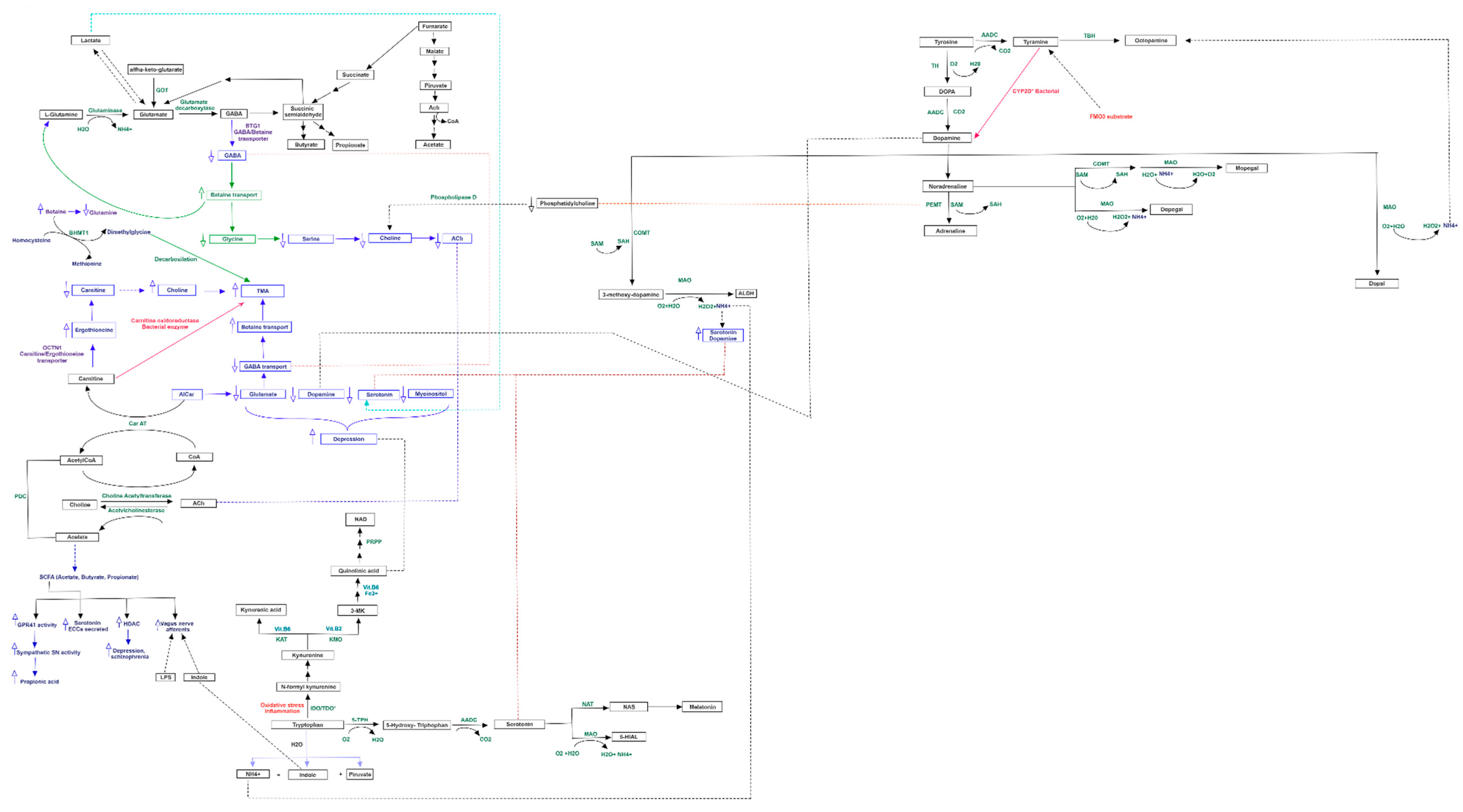
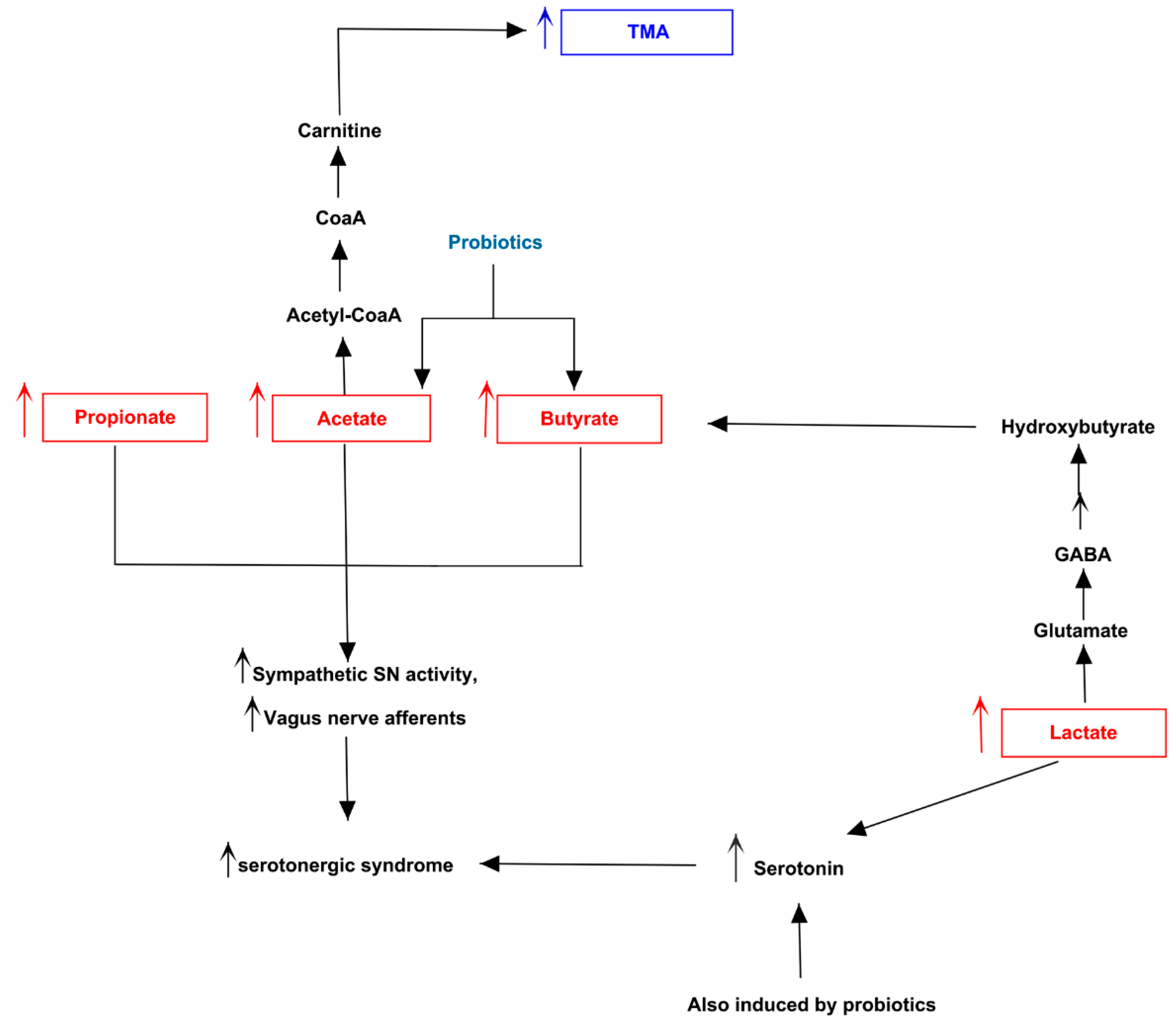
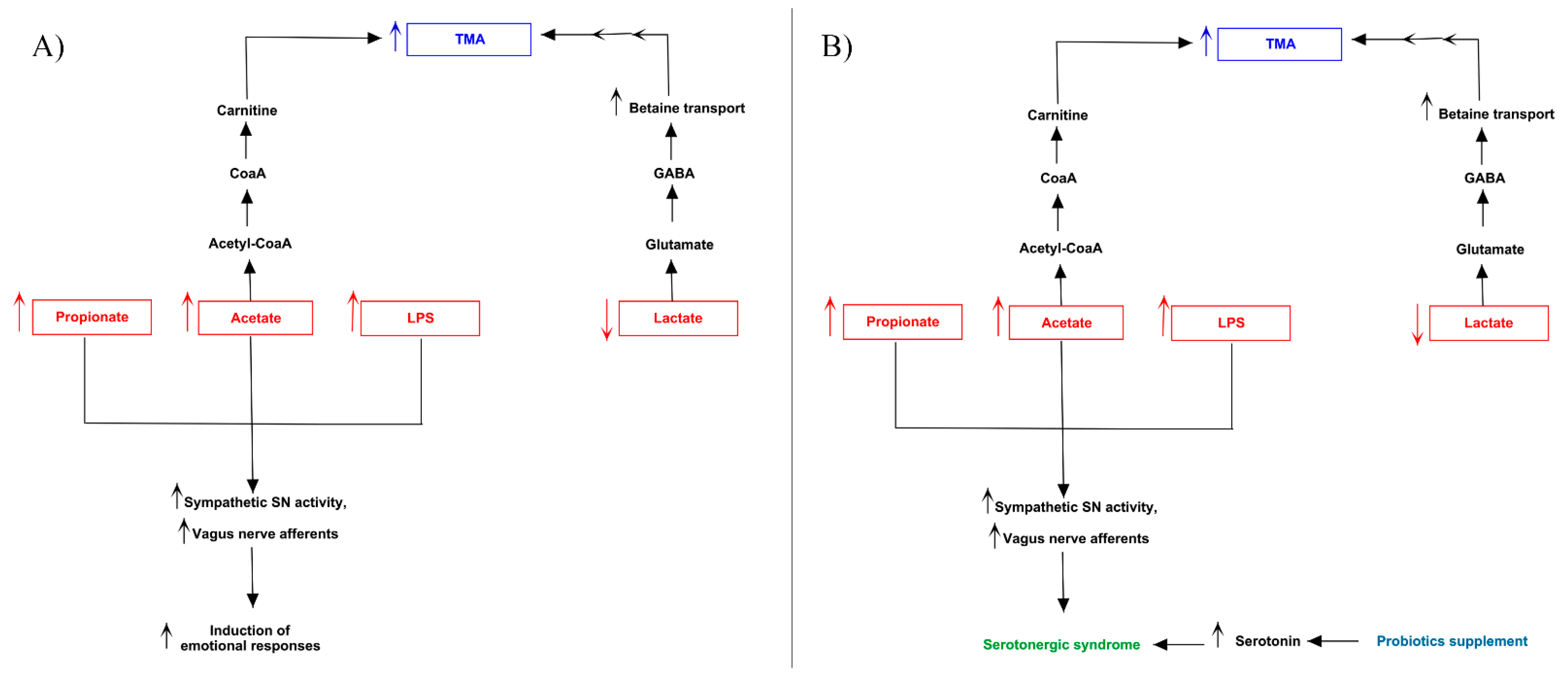
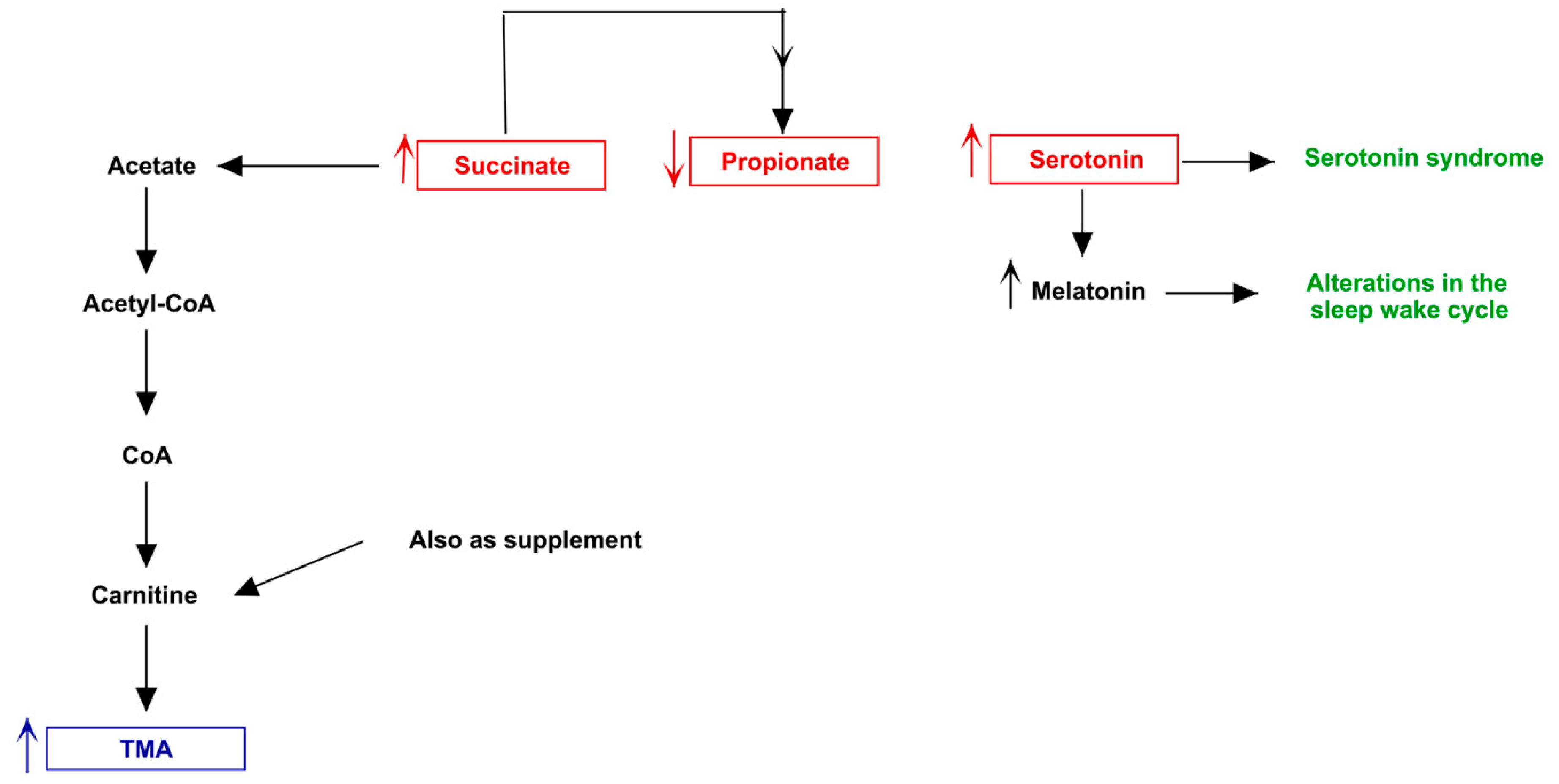
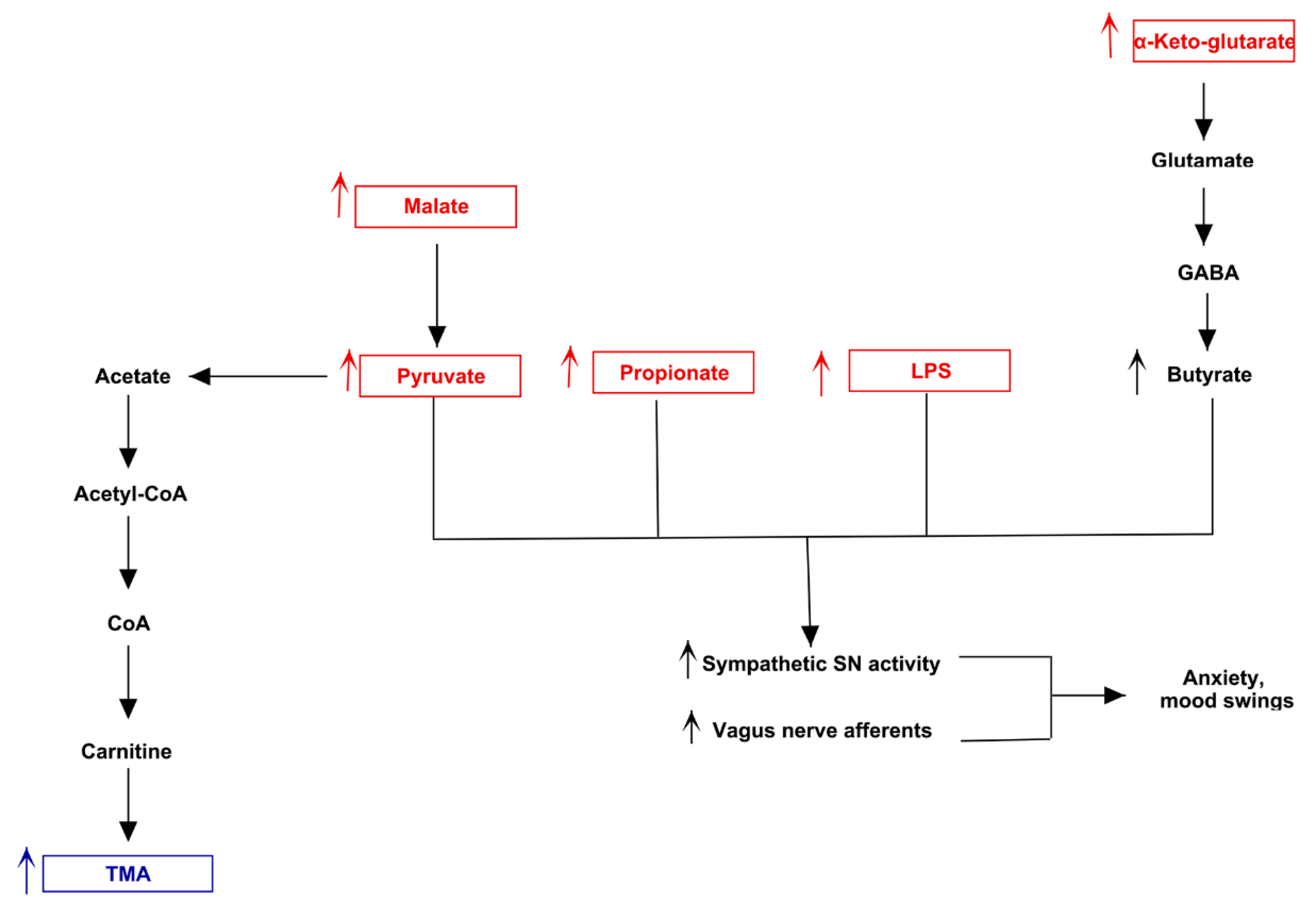
3.3. Pathway Analysis of Differential Abundances of Bacterial Families Suggested a Possible Biochemical Link between Microbiota Produced Metabolites, TMA Biosynthesis and Mood/Behavioral Disorders
4. Discussion
Limitations
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Conflicts of Interest
References
- Schmidt, A.C.; Leroux, J.C. Treatments of trimethylaminuria: Where we are and where we might be heading. Drug Discov. Today 2020, 25, 1710–1717. [Google Scholar] [CrossRef] [PubMed]
- Messenger, J.; Clark, S.; Massick, S.; Bechtel, M. A Review of Trimethylaminuria: (Fish Odor Syndrome). J. Clin. Aesthetic Dermatol. 2013, 6, 45–48. [Google Scholar]
- Fraser-Andrews, E.A.; Manning, N.J.; Ashton, G.H.; Eldridge, P.; McGrath, J.; Menage, H.D.P. Fish odour syndrome with features of both primary and secondary trimethylaminuria. Clin. Exp. Dermatol. 2003, 28, 203–205. [Google Scholar] [CrossRef] [PubMed]
- Esposito, T.; Varriale, B.; D’Angelo, R.; Amato, A.; Sidoti, A. Regulation of flavin-containing mono-oxygenase (Fmo3) gene expression by steroids in mice and humans. Horm. Mol. Biol. Clin. Investig. 2014, 20, 99–109. [Google Scholar] [CrossRef]
- Cruciani, G.; Valeri, A.; Goracci, L.; Pellegrino, R.M.; Buonerba, F.; Baroni, M. Flavin Monooxygenase Metabolism: Why Medicinal Chemists Should Matter. J. Med. Chem. 2014, 57, 6183–6196. [Google Scholar] [CrossRef]
- Shephard, E.A.; Treacy, E.P.; Phillips, I.R. Clinical utility gene card for: Trimethylaminuria. Eur. J. Hum. Genet. 2012, 20, 4–5. [Google Scholar] [CrossRef][Green Version]
- Scimone, C.; Donato, L.; Rinaldi, C.; Sidoti, A.; D’Angelo, R. First case of Currarino syndrome and trimethylaminuria: Two rare diseases for a complex clinical presentation. J. Dig. Dis. 2016, 17, 628–632. [Google Scholar] [CrossRef]
- Vogt, N.M.; Romano, K.A.; Darst, B.F.; Engelman, C.D.; Johnson, S.C.; Carlsson, C.M.; Asthana, S.; Blennow, K.; Zetterberg, H.; Bendlin, B.B.; et al. The gut microbiota-derived metabolite trimethylamine N-oxide is elevated in Alzheimer’s disease. Alzheimer’s Res. Ther. 2018, 10, 124. [Google Scholar] [CrossRef]
- Mackay, R.J.; McEntyre, C.J.; Henderson, C.; Lever, M.; George, P.M. Trimethylaminuria: Causes and Diagnosis of a Socially Distressing Condition. Clin. Biochem. Rev. 2011, 32, 33–43. [Google Scholar]
- Gu, M.; Mei, X.-L.; Zhao, Y.N. Sepsis and Cerebral Dysfunction: BBB Damage, Neuroinflammation, Oxidative Stress, Apoptosis and Autophagy as Key Mediators and the Potential Therapeutic Approaches. Neurotox. Res. 2020. [Google Scholar] [CrossRef]
- Abushik, P.A.; Karelina, T.V.; Sibarov, D.A.; Stepanenko, J.D.; Giniatullin, R.; Antonov, S.M. Homocysteine-induced membrane currents, calcium responses and changes of mitochondrial potential in rat cortical neurons. Zh. Evol. Biokhim Fiziol. 2015, 51, 258–265. [Google Scholar] [CrossRef] [PubMed]
- Machado, F.R.; Ferreira, A.G.; Da Cunha, A.A.; Tagliari, B.; Mussulini, B.H.; Wofchuk, S.; Wyse, A.T. Homocysteine alters glutamate uptake and Na+,K+-ATPase activity and oxidative status in rats hippocampus: Protection by vitamin C. Metab. Brain Dis. 2011, 26, 61–67. [Google Scholar] [CrossRef] [PubMed]
- Reas, D.L.; Pedersen, G.; Karterud, S.; Ro, O. Self-harm and suicidal behavior in borderline personality disorder with and without bulimia nervosa. J. Consult. Clin. Psychol. 2015, 83, 643–648. [Google Scholar] [CrossRef] [PubMed]
- Allaband, C.; McDonald, D.; Vazquez-Baeza, Y.; Minich, J.J.; Tripathi, A.; Brenner, D.A.; Loomba, R.; Smarr, L.; Sandborn, W.J.; Schnabl, B.; et al. Microbiome 101: Studying, Analyzing, and Interpreting Gut Microbiome Data for Clinicians. Clin. Gastroenterol. Hepatol. 2019, 17, 218–230. [Google Scholar] [CrossRef]
- Klindworth, A.; Pruesse, E.; Schweer, T.; Peplies, J.; Quast, C.; Horn, M.; Glockner, F.O. Evaluation of general 16S ribosomal RNA gene PCR primers for classical and next-generation sequencing-based diversity studies. Nucleic Acids Res. 2013, 41, e1. [Google Scholar] [CrossRef]
- Masella, A.P.; Bartram, A.K.; Truszkowski, J.M.; Brown, D.G.; Neufeld, J.D. PANDAseq: Paired-end assembler for illumina sequences. BMC Bioinform. 2012, 13, 31. [Google Scholar] [CrossRef]
- Caporaso, J.G.; Kuczynski, J.; Stombaugh, J.; Bittinger, K.; Bushman, F.D.; Costello, E.K.; Fierer, N.; Pena, A.G.; Goodrich, J.K.; Gordon, J.I.; et al. QIIME Allows Analysis of High-Throughput Community Sequencing data. Nat. Methods 2010, 7, 335–336. [Google Scholar] [CrossRef]
- Golob, J.L.; Margolis, E.; Hoffman, N.G.; Fredricks, D.N. Evaluating the accuracy of amplicon-based microbiome computational pipelines on simulated human gut microbial communities. BMC Bioinform. 2017, 18, 283. [Google Scholar] [CrossRef]
- Leong, L.E.X.; Taylor, S.L.; Shivasami, A.; Goldwater, P.N.; Rogers, G.B. Intestinal Microbiota Composition in Sudden Infant Death Syndrome and Age-Matched Controls. J. Pediatr. 2017, 191, 63–68. [Google Scholar] [CrossRef]
- Wandro, S.; Osborne, S.; Enriquez, C.; Bixby, C.; Arrieta, A.; Whiteson, K. The Microbiome and Metabolome of Preterm Infant Stool Are Personalized and Not Driven by Health Outcomes, Including Necrotizing Enterocolitis and Late-Onset Sepsis. mSphere 2018, 3. [Google Scholar] [CrossRef]
- Niu, J.; Xu, L.; Qian, Y.; Sun, Z.; Yu, D.; Huang, J.; Zhou, X.; Wang, Y.; Zhang, T.; Ren, R.; et al. Evolution of the Gut Microbiome in Early Childhood: A Cross-Sectional Study of Chinese Children. Front. Microbiol. 2020, 11, 439. [Google Scholar] [CrossRef] [PubMed]
- McNeill, T.W.; Sinkora, G.; Leavitt, F. Psychologic Classification of Low-Back Pain Patients: A Prognostic Tool. Spine 1986, 11, 955–959. [Google Scholar] [CrossRef] [PubMed]
- Le Boulch, M.; Dehais, P.; Combes, S.; Pascal, G. The MACADAM database: A MetAboliC pAthways DAtabase for Microbial taxonomic groups for mining potential metabolic capacities of archaeal and bacterial taxonomic groups. Database 2019, 2019. [Google Scholar] [CrossRef] [PubMed]
- Le Chatelier, E.; Nielsen, T.; Qin, J.; Prifti, E.; Hildebrand, F.; Falony, G.; Almeida, M.; Arumugam, M.; Batto, J.M.; Kennedy, S.; et al. Richness of human gut microbiome correlates with metabolic markers. Nature 2013, 500, 541–546. [Google Scholar] [CrossRef]
- Silva, Y.P.; Bernardi, A.; Frozza, R.L. The Role of Short-Chain Fatty Acids From Gut Microbiota in Gut-Brain Communication. Front. Endocrinol. 2020, 11, 25. [Google Scholar] [CrossRef]
- Jenkins, T.A.; Nguyen, J.C.; Polglaze, K.E.; Bertrand, P.P. Influence of Tryptophan and Serotonin on Mood and Cognition with a Possible Role of the Gut-Brain Axis. Nutrients 2016, 8, 56. [Google Scholar] [CrossRef]
- Murphy, M.M.; Gueant, J.L. B vitamins and one carbon metabolism micronutrients in health and disease. Biochime 2020, 173, 1–2. [Google Scholar] [CrossRef]
- Kutmon, M.; Van Iersel, M.P.; Bohler, A.; Kelder, T.; Nunes, N.; Pico, A.R.; Evelo, C.T. PathVisio 3: An Extendable Pathway Analysis Toolbox. PLoS Comput. Biol. 2015, 11, e1004085. [Google Scholar] [CrossRef]
- Vallianou, N.; Stratigou, T.; Christodoulatos, G.S.; Dalamaga, M. Understanding the Role of the Gut Microbiome and Microbial Metabolites in Obesity and Obesity-Associated Metabolic Disorders: Current Evidence and Perspectives. Curr. Obes. Rep. 2019, 8, 317–332. [Google Scholar] [CrossRef]
- Kim, Y.K.; Shin, C. The Microbiota-Gut-Brain Axis in Neuropsychiatric Disorders: Pathophysiological Mechanisms and Novel Treatments. Curr. Neuropharmacol. 2018, 16, 559–573. [Google Scholar] [CrossRef]
- Strandwitz, P. Neurotransmitter modulation by the gut microbiota. Brain Res. 2018, 1693, 128–133. [Google Scholar] [CrossRef] [PubMed]
- Tian, J.S.; Xia, X.T.; Wu, Y.F.; Zhao, L.; Xiang, H.; Du, G.H.; Zhang, X.; Qin, X.M. Discovery, screening and evaluation of a plasma biomarker panel for subjects with psychological suboptimal health state using (1)H-NMR-based metabolomics profiles. Sci. Rep. 2016, 6, 33820. [Google Scholar] [CrossRef] [PubMed]
- Nagalski, A.; Kozinski, K.; Wisniewska, M.B. Metabolic pathways in the periphery and brain: Contribution to mental disorders? Int. J. Biochem. Cell Biol. 2016, 80, 19–30. [Google Scholar] [CrossRef] [PubMed]
- McNiven, V.; Mamane, S.; Zai, G.; So, J. The Nose Knows… or Does it? Olfactory Reference Syndrome in Patients Presenting for Assessment of Unusual Body Odor. J. Nerv. Ment. Dis. 2019, 207, 145–151. [Google Scholar] [CrossRef] [PubMed]
- Phillips, I.R.; Shephard, E.A. Flavin-containing monooxygenase 3 (FMO3): Genetic variants and their consequences for drug metabolism and disease. Xenobiotica 2020, 50, 19–33. [Google Scholar] [CrossRef]
- Caspani, G.; Kennedy, S.; Foster, J.A.; Swann, J. Gut microbial metabolites in depression: Understanding the biochemical mechanisms. Microb. Cell 2019, 6, 454–481. [Google Scholar] [CrossRef]
- Carabotti, M.; Scirocco, A.; Maselli, M.A.; Severi, C. The gut-brain axis: Interactions between enteric microbiota, central and enteric nervous systems. Ann. Gastroenterol. 2015, 28, 203–209. [Google Scholar]
- Oliphant, K.; Allen-Vercoe, E. Macronutrient metabolism by the human gut microbiome: Major fermentation by-products and their impact on host health. Microbiome 2019, 7, 91. [Google Scholar] [CrossRef]
- Newington, J.T.; Harris, R.A.; Cumming, R.C. Reevaluating Metabolism in Alzheimer’s Disease from the Perspective of the Astrocyte-Neuron Lactate Shuttle Model. J. Neurodegener. Dis. 2013, 2013, 234572. [Google Scholar] [CrossRef]
- Jin, X.T.; Galvan, A.; Wichmann, T.; Smith, Y. Localization and Function of GABA Transporters GAT-1 and GAT-3 in the Basal Ganglia. Front. Syst. Neurosci. 2011, 5, 63. [Google Scholar] [CrossRef]
- Cheng, L.H.; Liu, Y.W.; Wu, C.C.; Wang, S.; Tsai, Y.C. Psychobiotics in mental health, neurodegenerative and neurodevelopmental disorders. J. Food Drug Anal. 2019, 27, 632–648. [Google Scholar] [CrossRef] [PubMed]
- Frick, A.; Ahs, F.; Engman, J.; Jonasson, M.; Alaie, I.; Bjorkstrand, J.; Frans, O.; Faria, V.; Linnman, C.; Appel, L.; et al. Serotonin Synthesis and Reuptake in Social Anxiety Disorder: A Positron Emission Tomography Study. JAMA Psychiatry 2015, 72, 794–802. [Google Scholar] [CrossRef] [PubMed]
- Fava, M.; Mischoulon, D. Folate in depression: Efficacy, safety, differences in formulations, and clinical issues. J. Clin. Psychiatry 2009, 70 (Suppl. 5), 12–17. [Google Scholar] [CrossRef] [PubMed]
- Spector, R.; Johanson, C.E. Vitamin transport and homeostasis in mammalian brain: Focus on Vitamins B and E. J. Neurochem. 2007, 103, 425–438. [Google Scholar] [CrossRef] [PubMed]
- Kennedy, P.J.; Cryan, J.F.; Dinan, T.G.; Clarke, G. Kynurenine pathway metabolism and the microbiota-gut-brain axis. Neuropharmacology 2017, 112, 399–412. [Google Scholar] [CrossRef]
- Baldani, J.I.; Rouws, L.; Cruz, L.M.; Olivares, F.L.; Schmid, M.; Hartmann, A. The Family Oxalobacteraceae. In The Prokaryotes: Alphaproteobacteria and Betaproteobacteria; Rosenberg, E., DeLong, E.F., Lory, S., Stackebrandt, E., Thompson, F., Eds.; Springer: Berlin/Heidelberg, Germany, 2014; pp. 919–974. [Google Scholar] [CrossRef]
- Yano, J.M.; Yu, K.; Donaldson, G.P.; Shastri, G.G.; Ann, P.; Ma, L.; Nagler, C.R.; Ismagilov, R.F.; Mazmanian, S.K.; Hsiao, E.Y. Indigenous Bacteria from the Gut Microbiota Regulate Host Serotonin Biosynthesis. Cell 2015, 161, 264–276. [Google Scholar] [CrossRef] [PubMed]













{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| ID | AGE | SEX | TMAU AGE of ONSET | DIET | ANTIBIOTIC MASSIVE USE | PROBIOTIC/FOOD SUPPLEMENTS | BEHAVIOR DISORDER | KIND OF BEHAVIOR DISORDER | OTHER |
|---|---|---|---|---|---|---|---|---|---|
| 1 | 30 | M | 17 | Chocolate, Eggs, Peas | NO | NO | YES | Anxiety, Fear, Suicidal instincts, Mood alteration | / |
| 2 | 40 | F | 14 | Fish, Vegetables | NO | NO | YES | Excessive emotionality, Anxiety | / |
| 3 | 54 | F | 6 | Dairy products, Meat, Fish | NO | L-carnitine, bromelain | YES | Migraine, Sleep disorders, Mood alteration, Sense of marginalization, Difficulties in social relations | / |
| 4 | 45 | F | 7 | Chocolate, Legumes, Eggs, Fish | YES | NO | YES | Chronic and rapid mental fatigue, Frequent headaches, Dizziness, Anxiety, Depression | Low levels of Folate, Plasmatic Vitamin B2 and D, Cu2+, Zn2+; High levels of PTH, homocysteine, Ca2+ |
| 5 | 44 | M | 34 | Coffee, Tea, White Meat, Vegetables, Fish | YES | L. acidophilus, Bifidobacterium lactis, L. rhamnosus, Streptococcus thermophilus and L. Paracasei | YES | Obsessive-compulsive disorder, Sense of marginalization | / |
| 6 | 36 | F | 9 | Vegetables, Coffee, Eggs | YES | Zinc, selenium, folic acid, iron, inulin, magnesium, L. Helveticus, B. longum spp.longum, Vitamin B6, Vitamin B1 and Vitamin D | YES | Mood alteration, Sense of marginalization, Suicidal instincts | / |
| 7 | 25 | F | 4 | Fish, Eggs, Chocolate, Legumes | NO | NO | YES | Depression, Obsessive-compulsive disorder, Sense of persecution | / |
| 1c | 47 | F | 8 | Gluten-free foods, Vegetables, Coffee | NO | NO | NO | NO | / |
| 2c | 26 | M | 10 | Fish, Chocolate, Red meat, Coffee, Alcohol | NO | Bifidobacterium lactis, L. acidophilus, L. plantarum, L. paracasei; Streptococcus salivarius subsp. thermophilus, Bifidobacterium brevis, Lactobacillus delbrueckii subsp. bulgaricus, Enterococcus faecium. | NO | NO | / |
| 3c | 20 | M | 16 | Gluten-free foods, Vegetables, Meat | NO | L. acidophilus, Bifidobacterium lactis, L. rhamnosus, Streptococcus thermophilus and L. Paracasei | NO | NO | / |
| 4c | 72 | F | 2 | Gluten-free and Lactose-free foods, Fish | NO | NO | NO | NO | High ROS and Arachidonic Acid |
| 5c | 35 | M | 35 | Red meat, Legumes, vegetables, Salmon | NO | NO | NO | NO | Use of alcohol |
| ID | 1 | 2 | 3 | 4 | 5 | 6 | 7 | 1c | 2c | 3c | 4c | 5c |
|---|---|---|---|---|---|---|---|---|---|---|---|---|
| Enterobacteriaceae [0.1–1.1] | 0.85 | 1.08 | 0.45 | 0.1 | 0.74 | 0.15 | 0.15 | 0.02 | 0.01 | 0.1 | 2.8 | 0.05 |
| Oxalobacteraceae [0.0–0.0] | 0 | 0.05 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0.08 | 0 |
| Enterococcaceae [0.0–0.0] | 0.02 | 0 | 0.02 | 0 | 0 | 0.68 | 0 | 0 | 0 | 0 | 0 | 0 |
| Erysipelotrichaceae [0.1–2.9] | 2.8 | 0.4 | 0.78 | 0.1 | 0.38 | 3.9 | 3.3 | 0.15 | 0.21 | 0.1 | 3.8 | 2.62 |
| Rikenellaceae [0.2–5.3] | 0.48 | 5.22 | 1.25 | 0.2 | 2.2 | 6.95 | 0.2 | 0.2 | 0.2 | 0.2 | 6.78 | 0.48 |
| Veilloneaceae [0.8–7.7] | 6.35 | 3.15 | 1.58 | 0.8 | 2.8 | 5.35 | 3.35 | 0.8 | 0.8 | 0.8 | 0.48 | 1.85 |
| Roseburia [0.0–0.9] | 0 | 0.15 | 0.25 | 0.85 | 0 | 0.04 | 1.03 | 3.09 | 4.4 | 0 | 1 | 1.53 |
| Streptococcaceae [0.1–1.8] | 0.28 | 0.22 | 3.48 | 0.01 | 0.15 | 2.62 | 0.15 | 0.1 | 0.1 | 0.03 | 0.32 | 0.08 |
| Clostridiaceae [0.1–1.4] | 0.28 | 1.45 | 1.25 | 287.8 | 134.1 | 0.28 | 1.6 | 0.1 | 0.1 | 0..23 | 0.32 | 0.18 |
| Lachnospiraceae [12.8–37.26] | 20.52 | 9.98 | 24.78 | 1.86 | 15.8 | 3.78 | 23.22 | 72.24 | 44.65 | 0.04 | 18.58 | 23.25 |
| Prevotellaceae [0.1–13.66] | 0.12 | 2.3 | 16.68 | 0.1 | 0.7 | 3.85 | 40.0 | 0.02 | 0.1 | 0.1 | 0.13 | 26.65 |
| Coriobacteriaceae [0.3–5.9] | 0.15 | 1.08 | 2.12 | 0.01 | 0.7 | 6.5 | 0.82 | 0.3 | 0.3 | 0.04 | 0.52 | 1.7 |
| Bacteroidaceae [3.2–35.36] | 55.62 | 17.5 | 9.98 | 3.2 | 9.2 | 25.38 | 1.4 | 3.2 | 3.2 | 3.2 | 14.58 | 9.45 |
| Ruminococcaceae [13.7–34.7] | 2.42 | 24.4 | 23.38 | 13.7 | 18.7 | 24.35 | 16.23 | 0.27 | 1.43 | 0.13 | 24.25 | 19.8 |
| Faecalibacterium [2.5–15.56] | 0 | 3.05 | 9.35 | 5.2 | 5.5 | 0.58 | 8.43 | 6.4 | 23.97 | 10.33 | 8.25 | 7.2 |
| Porphiromonodaceae [0.2–3.2] | 1.25 | 0.2 | 0.98 | 0.22 | 0.52 | 1.5 | 0.55 | 0.12 | 0.2 | 0.2 | 1.22 | 0.28 |
| Sutterellaceae [0.1–3.5] | 0.1 | 0.1 | 0.1 | 0.1 | 0.1 | 0.1 | 0.1 | 0.01 | 0.61 | 0.1 | 0.1 | 0.1 |
| Bifidobacteriaceae [0.1–7.96] | 4.38 | 1.82 | 0.38 | 0.39 | 3.55 | 3.88 | 0.1 | 0.1 | 0.003 | 0.11 | 0.1 | 1.05 |
| BACTERIA/METABOLITES | Lactate | Dopamine | Norepinephrine | Acetate | Serotonin | Succinate | Butyrate | Glycolate | Propionate | Pyruvate | α-ketoglutarate | LPS | Malate | Tryptophan | GABA |
|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|
| Enterobacteriaceae | X | X | X | X | X | X | X | ||||||||
| Oxalobacteraceae | X | X | X | X | X | X | |||||||||
| Enterococcaceae | X | X | X | ||||||||||||
| Erysipelotrichaceae | X | X | |||||||||||||
| Bifidobacteriaceae | X | X | X | ||||||||||||
| Rikenellaceae | X | X | X | ||||||||||||
| Sutterellaceae | X | X | |||||||||||||
| Veilloneaceae | X | X | X | X | |||||||||||
| Roseburia | X | X | X | X | |||||||||||
| Ruminococcaceae | X | X | X | ||||||||||||
| Streptococcaceae | X | X | X | ||||||||||||
| Clostridiaceae | X | X | X | X | |||||||||||
| Lachnospiraceae | X | X | X | X | |||||||||||
| Prevotellaceae | X | X | X | ||||||||||||
| Coriobacteriaceae | X | X | |||||||||||||
| Bacteroidaceae | X | X | X | X | X | ||||||||||
| Faecalibacteriaceae | X | ||||||||||||||
| Porphiromonodaceae | X | X | X |
| ID | 1 | 2 | 3 | 4 | 5 | 6 | 7 | 1c | 2c | 3c | 4c | 5c |
|---|---|---|---|---|---|---|---|---|---|---|---|---|
| Acetate | ↑ | ↓ | ↑ | ↑ | ↓ | ↓ | ↓ | ↓ | ↓ | |||
| Lactate | ↓ | ↓ | ↑ | ↓ | ↑ | ↓ | ↓ | ↑ | ||||
| Succinate | ↑ | ↑ | ↑ | ↓ | ↓ | ↓ | ||||||
| Dopamine | ↓ | ↑ | ↑ | |||||||||
| Norepinephrine | ↓ | ↓ | ↑ | ↑ | ||||||||
| Serotonin | ↑ | ↓ | ↑ | ↓ | ↓ | ↑ | ||||||
| α-ketoglutarate | ↑ | ↑ | ||||||||||
| Malate | ↑ | ↑ | ||||||||||
| Pyruvate | ↑ | ↑ | ||||||||||
| LPS | ↑ | ↑ | ↓ | ↑ | ||||||||
| Propionate | ↑ | ↑ | ↓ | ↑ | ↑ | ↓ | ↓ | |||||
| Butyrate | ↓ | ↓ | ↓ | ↑ | ↓ | ↑ | ↑ | ↑ | ↑ | ↑ | ||
| Tryptophan | ↓ | ↓ | ||||||||||
| GABA | ↓ |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Donato, L.; Alibrandi, S.; Scimone, C.; Castagnetti, A.; Rao, G.; Sidoti, A.; D’Angelo, R. Gut-Brain Axis Cross-Talk and Limbic Disorders as Biological Basis of Secondary TMAU. J. Pers. Med. 2021, 11, 87. https://doi.org/10.3390/jpm11020087
Donato L, Alibrandi S, Scimone C, Castagnetti A, Rao G, Sidoti A, D’Angelo R. Gut-Brain Axis Cross-Talk and Limbic Disorders as Biological Basis of Secondary TMAU. Journal of Personalized Medicine. 2021; 11(2):87. https://doi.org/10.3390/jpm11020087
Chicago/Turabian StyleDonato, Luigi, Simona Alibrandi, Concetta Scimone, Andrea Castagnetti, Giacomo Rao, Antonina Sidoti, and Rosalia D’Angelo. 2021. "Gut-Brain Axis Cross-Talk and Limbic Disorders as Biological Basis of Secondary TMAU" Journal of Personalized Medicine 11, no. 2: 87. https://doi.org/10.3390/jpm11020087
APA StyleDonato, L., Alibrandi, S., Scimone, C., Castagnetti, A., Rao, G., Sidoti, A., & D’Angelo, R. (2021). Gut-Brain Axis Cross-Talk and Limbic Disorders as Biological Basis of Secondary TMAU. Journal of Personalized Medicine, 11(2), 87. https://doi.org/10.3390/jpm11020087