
Ionizing Radiation Protein Biomarkers in Normal Tissue and Their Correlation to Radiosensitivity: A Systematic Review


Abstract
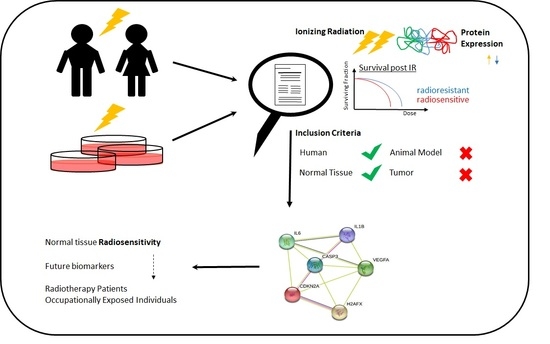
1. Introduction
1.1. Background and Rationale
1.2. Objectives
2. Methods
2.1. Protocol and Registration
2.2. Eligibility Criteria
2.3. Information Sources
2.4. Search
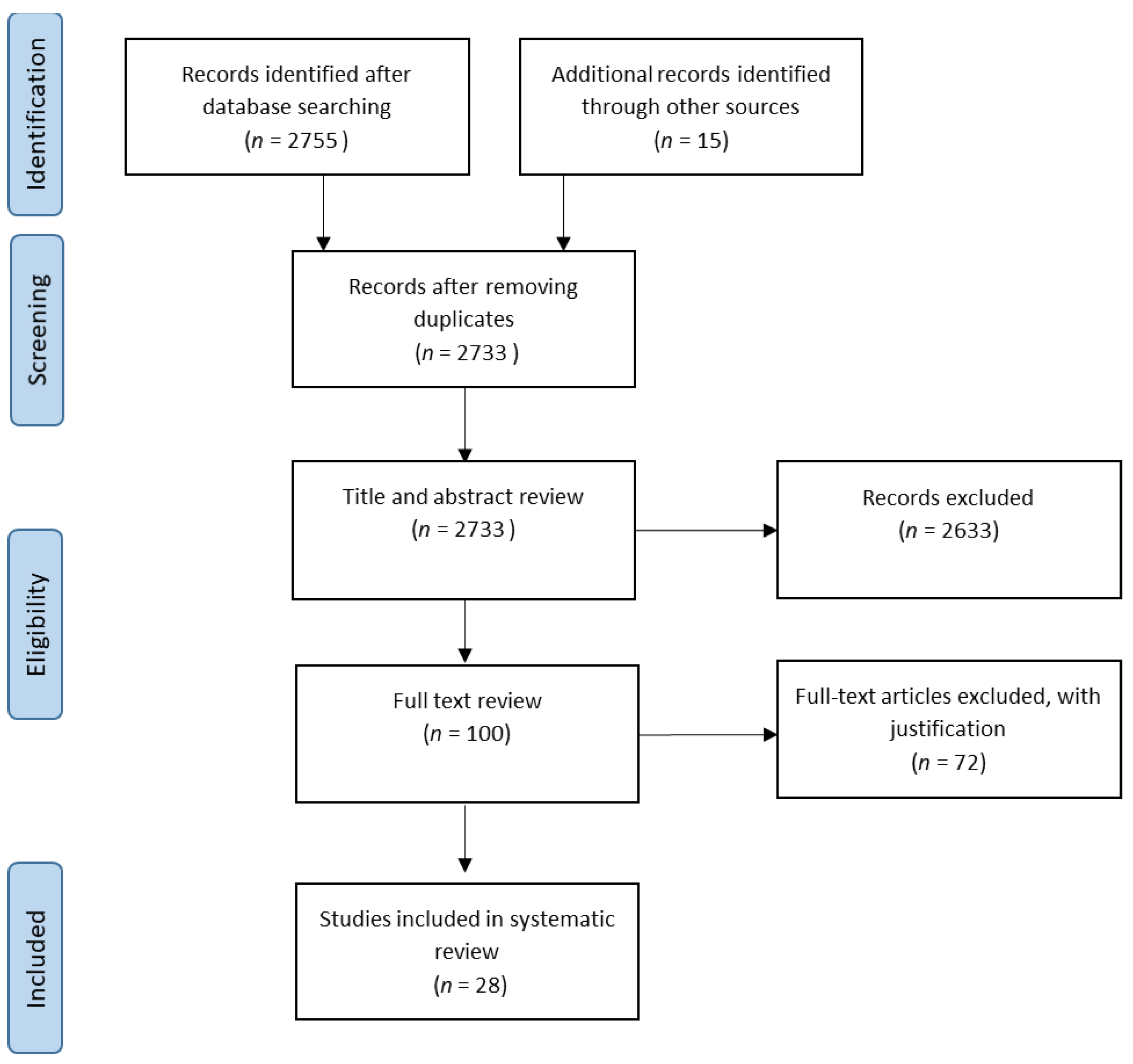
2.5. Study Selection
2.6. Data Collection Process
2.7. Grouping Studies for Synthesis
2.8. Standardised Metric and Transformation Used
2.9. Synthesis Methods
2.10. Certainity of Evidence
3. Results
3.1. Study Characteristics of the included Articles
3.2. IR-Induced Changes in Repair Foci Proteins
3.3. IR-Induced Deregulated Proteins Excluding Repair Foci and Risk of Biases
- Can we be confident in the exposure assessment?
- Can we be confident in the outcome assessment?
- Did the study design or analyses account for important confounding and modifying variables?
3.3.1. Vascular Endothelial Growth Factor (VEGF)
3.3.2. Caspase 3
3.3.3. p16INK4A (Cyclin-Dependent Kinase Inhibitor 2A, CDKN2A)
3.3.4. Interleukin-6 (IL-6)
3.3.5. Interleukin-1 Beta (IL-1β)
4. Outlook
5. Conclusions
6. Differences between Protocol and the Review
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Global Cancer Observatory. Available online: https://gco.iarc.fr/today/data/factsheets/cancers/39-All-cancers-fact-sheet.pdf (accessed on 11 December 2020).
- Rosenblatt, E.; Izewska, J.; Anacak, Y.; Pynda, Y.; Scalliet, P.; Boniol, M.; Autier, P. Radiotherapy capacity in European countries: An analysis of the Directory of Radiotherapy Centres (DIRAC) database. Lancet Oncol. 2013, 14, e79–e86. [Google Scholar] [CrossRef]
- Fazel, R.; Gerber, T.C.; Balter, S.; Brenner, D.J.; Carr, J.J.; Cerqueira, M.D.; Chen, J.; Einstein, A.J.; Krumholz, H.M.; Mahesh, M.; et al. Approaches to enhancing radiation safety in cardiovascular imaging: A scientific statement from the American Heart Association. Circulation 2014, 130, 1730–1748. [Google Scholar] [CrossRef]
- Salomaa, S.; Jung, T. Roadmap for research on individual radiosensitivity and radiosusceptibility—The MELODI view on research needs. Int. J. Radiat. Biol. 2020, 96, 277–279. [Google Scholar] [CrossRef]
- Gomolka, M.; Blyth, B.; Bourguignon, M.; Badie, C.; Schmitz, A.; Talbot, C.; Hoeschen, C.; Salomaa, S. Potential screening assays for individual radiation sensitivity and susceptibility and their current validation state. Int. J. Radiat. Biol. 2019, 96, 280–296. [Google Scholar] [CrossRef]
- Wojcik, A.; Bouffler, S.; Hauptmann, M.; Rajaraman, P. Considerations on the use of the terms radiosensitivity and radiosusceptibility. J. Radiol. Prot. 2018, 38, N25–N29. [Google Scholar] [CrossRef]
- Averbeck, D.; Salomaa, S.; Bouffler, S.; Ottolenghi, A.; Smyth, V.; Sabatier, L. Progress in low dose health risk research: Novel effects and new concepts in low dose radiobiology. Mutat. Res. 2018, 776, 46–69. [Google Scholar] [CrossRef]
- Hoeller, U.; Borgmann, K.; Bonacker, M.; Kuhlmey, A.; Bajrovic, A.; Jung, H.; Alberti, W.; Dikomey, E. Individual radiosensitivity measured with lymphocytes may be used to predict the risk of fibrosis after radiotherapy for breast cancer. Radiother. Oncol. 2003, 69, 137–144. [Google Scholar] [CrossRef] [PubMed]
- Heemsbergen, W.D.; Peeters, S.T.; Koper, P.C.; Hoogeman, M.S.; Lebesque, J.V. Acute and late gastrointestinal toxicity after radiotherapy in prostate cancer patients: Consequential late damage. Int. J. Radiat. Oncol. Biol. Phys. 2006, 66, 3–10. [Google Scholar] [CrossRef] [PubMed]
- Pollack, A.; Zagars, G.K.; Antolak, J.A.; Kuban, D.A.; Rosen, I.I. Prostate biopsy status and PSA nadir level as early surrogates for treatment failure: Analysis of a prostate cancer randomized radiation dose escalation trial. Int. J. Radiat. Oncol. Biol. Phys. 2002, 54, 677–685. [Google Scholar] [CrossRef]
- Nakanishi, K.; Taniguchi, T.; Ranganathan, V.; New, H.V.; Moreau, L.A.; Stotsky, M.; Mathew, C.G.; Kastan, M.B.; Weaver, D.T.; D’Andrea, A.D. Interaction of FANCD2 and NBS1 in the DNA damage response. Nat. Cell Biol. 2002, 4, 913–920. [Google Scholar] [CrossRef] [PubMed]
- Petrini, J.H. The mammalian Mre11-Rad50-nbs1 protein complex: Integration of functions in the cellular DNA-damage response. Am. J. Hum. Genet. 1999, 64, 1264–1269. [Google Scholar] [CrossRef]
- Digweed, M. Human genetic instability syndromes: Single gene defects with increased risk of cancer. Toxicol. Lett. 1993, 67, 259–281. [Google Scholar] [CrossRef]
- Pietrucha, B.M.; Heropolitanska-Pliszka, E.; Wakulinska, A.; Skopczynska, H.; Gatti, R.A.; Bernatowska, E. Ataxia-telangiectasia with hyper-IgM and Wilms tumor: Fatal reaction to irradiation. J. Pediatr. Hematol. Oncol. 2010, 32, e28–e30. [Google Scholar] [CrossRef]
- Mizutani, S.; Takagi, M. XCIND as a genetic disease of X-irradiation hypersensitivity and cancer susceptibility. Int. J. Hematol. 2013, 97, 37–42. [Google Scholar] [CrossRef]
- Andreassen, C.N.; Schack, L.M.; Laursen, L.V.; Alsner, J. Radiogenomics—Current status, challenges and future directions. Cancer Lett. 2016, 382, 127–136. [Google Scholar] [CrossRef]
- Chua, M.L.; Rothkamm, K. Biomarkers of radiation exposure: Can they predict normal tissue radiosensitivity? Clin. Oncol. 2013, 25, 610–616. [Google Scholar] [CrossRef] [PubMed]
- Hall, J.; Jeggo, P.A.; West, C.; Gomolka, M.; Quintens, R.; Badie, C.; Laurent, O.; Aerts, A.; Anastasov, N.; Azimzadeh, O.; et al. Ionizing radiation biomarkers in epidemiological studies—An update. Mutat. Res. 2017, 771, 59–84. [Google Scholar] [CrossRef]
- Tebaldi, T.; Re, A.; Viero, G.; Pegoretti, I.; Passerini, A.; Blanzieri, E.; Quattrone, A. Widespread uncoupling between transcriptome and translatome variations after a stimulus in mammalian cells. BMC Genomics 2012, 13, 220. [Google Scholar] [CrossRef] [PubMed]
- Dietz, A.; Gomolka, M.; Moertl, S.; Subedi, P. Ionizing Radiation Protein Biomarkers in Normal Tissue and Their Correlation to Radiosensitivity: Protocol for a Systematic Review. J. Pers. Med. 2020, 11, 3. [Google Scholar] [CrossRef]
- Pernot, E.; Hall, J.; Baatout, S.; Benotmane, M.A.; Blanchardon, E.; Bouffler, S.; El Saghire, H.; Gomolka, M.; Guertler, A.; Harms-Ringdahl, M.; et al. Ionizing radiation biomarkers for potential use in epidemiological studies. Mutat. Res. 2012, 751, 258–286. [Google Scholar] [CrossRef]
- National Library of Medicine, Pubmed. Available online: https://pubmed.ncbi.nlm.nih.gov/ (accessed on 28 December 2020).
- Clarivate Analytics, Web of Science. Available online: https://apps.webofknowledge.com/ (accessed on 28 December 2020).
- Van Oorschot, B.; Hovingh, S.E.; Moerland, P.D.; Medema, J.P.; Stalpers, L.J.; Vrieling, H.; Franken, N.A. Reduced activity of double-strand break repair genes in prostate cancer patients with late normal tissue radiation toxicity. Int. J. Radiat. Oncol. Biol. Phys. 2014, 88, 664–670. [Google Scholar] [CrossRef]
- Vasireddy, R.S.; Sprung, C.N.; Cempaka, N.L.; Chao, M.; McKay, M.J. H2AX phosphorylation screen of cells from radiosensitive cancer patients reveals a novel DNA double-strand break repair cellular phenotype. Br. J. Cancer 2010, 102, 1511–1518. [Google Scholar] [CrossRef] [PubMed]
- Bourton, E.C.; Plowman, P.N.; Smith, D.; Arlett, C.F.; Parris, C.N. Prolonged expression of the gamma-H2AX DNA repair biomarker correlates with excess acute and chronic toxicity from radiotherapy treatment. Int. J. Cancer 2011, 129, 2928–2934. [Google Scholar] [CrossRef]
- Mumbrekar, K.D.; Fernandes, D.J.; Goutham, H.V.; Sharan, K.; Vadhiraja, B.M.; Satyamoorthy, K.; Bola Sadashiva, S.R. Influence of double-strand break repair on radiation therapy-induced acute skin reactions in breast cancer patients. Int. J. Radiat. Oncol. Biol. Phys. 2014, 88, 671–676. [Google Scholar] [CrossRef]
- Pouliliou, S.E.; Lialiaris, T.S.; Dimitriou, T.; Giatromanolaki, A.; Papazoglou, D.; Pappa, A.; Pistevou, K.; Kalamida, D.; Koukourakis, M.I. Survival Fraction at 2 Gy and gammaH2AX Expression Kinetics in Peripheral Blood Lymphocytes From Cancer Patients: Relationship With Acute Radiation-Induced Toxicities. Int. J. Radiat. Oncol. Biol. Phys. 2015, 92, 667–674. [Google Scholar] [CrossRef] [PubMed]
- Lobachevsky, P.; Leong, T.; Daly, P.; Smith, J.; Best, N.; Tomaszewski, J.; Thompson, E.R.; Li, N.; Campbell, I.G.; Martin, R.F.; et al. Compromized DNA repair as a basis for identification of cancer radiotherapy patients with extreme radiosensitivity. Cancer Lett. 2016, 383, 212–219. [Google Scholar] [CrossRef]
- Buchbinder, D.; Smith, M.J.; Kawahara, M.; Cowan, M.J.; Buzby, J.S.; Abraham, R.S. Application of a radiosensitivity flow assay in a patient with DNA ligase 4 deficiency. Blood Adv. 2018, 2, 1828–1832. [Google Scholar] [CrossRef]
- COPERNIC Project Investigators; Granzotto, A.; Benadjaoud, M.A.; Vogin, G.; Devic, C.; Ferlazzo, M.L.; Bodgi, L.; Pereira, S.; Sonzogni, L.; Forcheron, F.; et al. Influence of Nucleoshuttling of the ATM Protein in the Healthy Tissues Response to Radiation Therapy: Toward a Molecular Classification of Human Radiosensitivity. Int. J. Radiat. Oncol. Biol. Phys. 2016, 94, 450–460. [Google Scholar] [CrossRef]
- Djuzenova, C.S.; Elsner, I.; Katzer, A.; Worschech, E.; Distel, L.V.; Flentje, M.; Polat, B. Radiosensitivity in breast cancer assessed by the histone gamma-H2AX and 53BP1 foci. Radiat. Oncol. 2013, 8, 98. [Google Scholar] [CrossRef]
- Goutham, H.V.; Mumbrekar, K.D.; Vadhiraja, B.M.; Fernandes, D.J.; Sharan, K.; Kanive Parashiva, G.; Kapaettu, S.; Bola Sadashiva, S.R. DNA double-strand break analysis by gamma-H2AX foci: A useful method for determining the overreactors to radiation-induced acute reactions among head-and-neck cancer patients. Int. J. Radiat. Oncol. Biol. Phys. 2012, 84, e607–e612. [Google Scholar] [CrossRef]
- Vandersickel, V.; Depuydt, J.; Van Bockstaele, B.; Perletti, G.; Philippe, J.; Thierens, H.; Vral, A. Early increase of radiation-induced gammaH2AX foci in a human Ku70/80 knockdown cell line characterized by an enhanced radiosensitivity. J. Radiat. Res. 2010, 51, 633–641. [Google Scholar] [CrossRef]
- Martin, N.T.; Nakamura, K.; Paila, U.; Woo, J.; Brown, C.; Wright, J.A.; Teraoka, S.N.; Haghayegh, S.; McCurdy, D.; Schneider, M.; et al. Homozygous mutation of MTPAP causes cellular radiosensitivity and persistent DNA double-strand breaks. Cell Death Dis. 2014, 5, e1130. [Google Scholar] [CrossRef]
- Martin, N.T.; Nahas, S.A.; Tunuguntla, R.; Fike, F.; Gatti, R.A. Assessing ‘radiosensitivity’ with kinetic profiles of gamma-H2AX, 53BP1 and BRCA1 foci. Radiother. Oncol. 2011, 101, 35–38. [Google Scholar] [CrossRef]
- Minafra, L.; Bravatà, V.; Russo, G.; Forte, G.I.; Cammarata, F.P.; Ripamonti, M.; Candiano, G.; Cervello, M.; Giallongo, A.; Perconti, G.; et al. Gene Expression Profiling of MCF10A Breast Epithelial Cells Exposed to IOERT. Anticancer Res. 2015, 35, 3223–3234. [Google Scholar]
- Miyake, T.; Shimada, M.; Matsumoto, Y.; Okino, A. DNA Damage Response After Ionizing Radiation Exposure in Skin Keratinocytes Derived from Human-Induced Pluripotent Stem Cells. Int. J. Radiat. Oncol. Biol. Phys. 2019, 105, 193–205. [Google Scholar] [CrossRef]
- Nguyen, H.Q.; Belkacemi, Y.; Mann, C.; Hoffschir, F.; Kerbrat, S.; Surenaud, M.; Zadigue, P.; de La Taille, A.; Romeo, P.H.; Le Gouvello, S. Human CCR6+ Th17 Lymphocytes Are Highly Sensitive to Radiation-Induced Senescence and Are a Potential Target for Prevention of Radiation-Induced Toxicity. Int. J. Radiat. Oncol. Biol. Phys. 2020, 108, 314–325. [Google Scholar] [CrossRef] [PubMed]
- Braicu, E.I.; Gasimli, K.; Richter, R.; Nassir, M.; Kümmel, S.; Blohmer, J.-U.; Yalcinkaya, I.; Chekerov, R.; Ignat, I.; Ionescu, A.; et al. Role of serum VEGFA, TIMP2, MMP2 and MMP9 in Monitoring Response to Adjuvant Radiochemotherapy in Patients with Primary Cervical Cancer—Results of a Companion Protocol of the Randomized NOGGO-AGO Phase III Clinical Trial. Anticancer Res. 2014, 34, 385–391. [Google Scholar]
- Rodriguez-Gil, J.L.; Takita, C.; Wright, J.; Reis, I.M.; Zhao, W.; Lally, B.E.; Hu, J.J. Inflammatory biomarker C-reactive protein and radiotherapy-induced early adverse skin reactions in patients with breast cancer. Cancer Epidemiol. Biomark. Prev. 2014, 23, 1873–1883. [Google Scholar] [CrossRef]
- Skiold, S.; Azimzadeh, O.; Merl-Pham, J.; Naslund, I.; Wersall, P.; Lidbrink, E.; Tapio, S.; Harms-Ringdahl, M.; Haghdoost, S. Unique proteomic signature for radiation sensitive patients; a comparative study between normo-sensitive and radiation sensitive breast cancer patients. Mutat. Res. 2015, 776, 128–135. [Google Scholar] [CrossRef]
- Yu, B.B.; Lin, G.X.; Li, L.; Qu, S.; Liang, Z.G.; Chen, K.H.; Zhou, L.; Lu, Q.T.; Sun, Y.C.; Zhu, X.D. Cofilin-2 Acts as a Marker for Predicting Radiotherapy Response and Is a Potential Therapeutic Target in Nasopharyngeal Carcinoma. Med. Sci. Monit. 2018, 24, 2317–2329. [Google Scholar] [CrossRef] [PubMed]
- Lacombe, J.; Brengues, M.; Mange, A.; Bourgier, C.; Gourgou, S.; Pelegrin, A.; Ozsahin, M.; Solassol, J.; Azria, D. Quantitative proteomic analysis reveals AK2 as potential biomarker for late normal tissue radiotoxicity. Radiat. Oncol. 2019, 14, 142. [Google Scholar] [CrossRef]
- Cao, M.; Cabrera, R.; Xu, Y.; Liu, C.; Nelson, D. Different radiosensitivity of CD4(+)CD25(+) regulatory T cells and effector T cells to low dose gamma irradiation in vitro. Int. J. Radiat. Biol. 2011, 87, 71–80. [Google Scholar] [CrossRef]
- Park, M.T.; Oh, E.T.; Song, M.J.; Lee, H.; Park, H.J. Radio-sensitivities and angiogenic signaling pathways of irradiated normal endothelial cells derived from diverse human organs. J. Radiat. Res. 2012, 53, 570–580. [Google Scholar] [CrossRef][Green Version]
- Fekete, N.; Erle, A.; Amann, E.M.; Furst, D.; Rojewski, M.T.; Langonne, A.; Sensebe, L.; Schrezenmeier, H.; Schmidtke-Schrezenmeier, G. Effect of high-dose irradiation on human bone-marrow-derived mesenchymal stromal cells. Tissue Eng. Part C Methods 2015, 21, 112–122. [Google Scholar] [CrossRef]
- Nishad, S.; Ghosh, A. Dynamic changes in the proteome of human peripheral blood mononuclear cells with low dose ionizing radiation. Mutat. Res. Genet. Toxicol. Environ. Mutagen. 2016, 797, 9–20. [Google Scholar] [CrossRef]
- Shimura, T.; Hamada, N.; Sasatani, M.; Kamiya, K.; Kunugita, N. Nuclear accumulation of cyclin D1 following long-term fractionated exposures to low-dose ionizing radiation in normal human diploid cells. Cell Cycle 2014, 13, 1248–1255. [Google Scholar] [CrossRef][Green Version]
- Yim, J.H.; Yun, J.M.; Kim, J.Y.; Lee, I.K.; Nam, S.Y.; Kim, C.S. Phosphoprotein profiles of candidate markers for early cellular responses to low-dose gamma-radiation in normal human fibroblast cells. J. Radiat. Res. 2017, 58, 329–340. [Google Scholar] [CrossRef] [PubMed]
- Moertl, S.; Buschmann, D.; Azimzadeh, O.; Schneider, M.; Kell, R.; Winkler, K.; Tapio, S.; Hornhardt, S.; Merl-Pham, J.; Pfaffl, M.W.; et al. Radiation Exposure of Peripheral Mononuclear Blood Cells Alters the Composition and Function of Secreted Extracellular Vesicles. Int. J. Mol. Sci. 2020, 21, 2336. [Google Scholar] [CrossRef]
- Rogakou, E.P.; Pilch, D.R.; Orr, A.H.; Ivanova, V.S.; Bonner, W.M. DNA double-stranded breaks induce histone H2AX phosphorylation on serine 139. J. Biol. Chem. 1998, 273, 5858–5868. [Google Scholar] [CrossRef]
- Rogakou, E.P.; Boon, C.; Redon, C.; Bonner, W.M. Megabase chromatin domains involved in DNA double-strand breaks in vivo. J. Cell Biol. 1999, 146, 905–916. [Google Scholar] [CrossRef]
- Paull, T.T.; Rogakou, E.P.; Yamazaki, V.; Kirchgessner, C.U.; Gellert, M.; Bonner, W.M. A critical role for histone H2AX in recruitment of repair factors to nuclear foci after DNA damage. Curr. Biol. 2000, 10, 886–895. [Google Scholar] [CrossRef]
- Burma, S.; Chen, D.J. Role of DNA-PK in the cellular response to DNA double-strand breaks. DNA Repair 2004, 3, 909–918. [Google Scholar] [CrossRef]
- Lobrich, M.; Shibata, A.; Beucher, A.; Fisher, A.; Ensminger, M.; Goodarzi, A.A.; Barton, O.; Jeggo, P.A. gammaH2AX foci analysis for monitoring DNA double-strand break repair: Strengths, limitations and optimization. Cell Cycle 2010, 9, 662–669. [Google Scholar] [CrossRef]
- Panier, S.; Boulton, S.J. Double-strand break repair: 53BP1 comes into focus. Nat. Rev. Mol. Cell. Biol. 2014, 15, 7–18. [Google Scholar] [CrossRef]
- Bourton, E.C.; Plowman, P.N.; Zahir, S.A.; Senguloglu, G.U.; Serrai, H.; Bottley, G.; Parris, C.N. Multispectral imaging flow cytometry reveals distinct frequencies of gamma-H2AX foci induction in DNA double strand break repair defective human cell lines. Cytom. A 2012, 81, 130–137. [Google Scholar] [CrossRef] [PubMed]
- Kuhne, M.; Riballo, E.; Rief, N.; Rothkamm, K.; Jeggo, P.A.; Lobrich, M. A double-strand break repair defect in ATM-deficient cells contributes to radiosensitivity. Cancer Res. 2004, 64, 500–508. [Google Scholar] [CrossRef]
- Valdiglesias, V.; Giunta, S.; Fenech, M.; Neri, M.; Bonassi, S. gammaH2AX as a marker of DNA double strand breaks and genomic instability in human population studies. Mutat. Res. 2013, 753, 24–40. [Google Scholar] [CrossRef]
- Vandevoorde, C.; Gomolka, M.; Roessler, U.; Samaga, D.; Lindholm, C.; Fernet, M.; Hall, J.; Pernot, E.; El-Saghire, H.; Baatout, S.; et al. EPI-CT: In vitro assessment of the applicability of the gamma-H2AX-foci assay as cellular biomarker for exposure in a multicentre study of children in diagnostic radiology. Int. J. Radiat. Biol. 2015, 91, 653–663. [Google Scholar] [CrossRef]
- National Toxicology Program. Handbook for Conducting a Literature-Based Health Assessment Using OHAT Approach for Systematic Review and Evidence Integration; U.S. Department of Health and Human Services; National Toxicology Program: Washington, DC, USA, 2019. [Google Scholar]
- Murphy, J.F.; Fitzgerald, D.J. Vascular endothelial growth factor induces cyclooxygenase-dependent proliferation of endothelial cells via the VEGF-2 receptor. FASEB J. 2001, 15, 1667–1669. [Google Scholar] [CrossRef]
- Dixelius, J.; Olsson, A.K.; Thulin, A.; Lee, C.; Johansson, I.; Claesson-Welsh, L. Minimal active domain and mechanism of action of the angiogenesis inhibitor histidine-rich glycoprotein. Cancer Res. 2006, 66, 2089–2097. [Google Scholar] [CrossRef]
- Yu, L.; Chen, Y.; Tooze, S.A. Autophagy pathway: Cellular and molecular mechanisms. Autophagy 2018, 14, 207–215. [Google Scholar] [CrossRef]
- Shintani, T.; Klionsky, D.J. Autophagy in health and disease: A double-edged sword. Science 2004, 306, 990–995. [Google Scholar] [CrossRef]
- Mizushima, N.; Levine, B.; Cuervo, A.M.; Klionsky, D.J. Autophagy fights disease through cellular self-digestion. Nature 2008, 451, 1069–1075. [Google Scholar] [CrossRef]
- Yuan, X.; Du, J.; Hua, S.; Zhang, H.; Gu, C.; Wang, J.; Yang, L.; Huang, J.; Yu, J.; Liu, F. Suppression of autophagy augments the radiosensitizing effects of STAT3 inhibition on human glioma cells. Exp. Cell Res. 2015, 330, 267–276. [Google Scholar] [CrossRef] [PubMed]
- Chang, L.; Graham, P.H.; Hao, J.; Ni, J.; Bucci, J.; Cozzi, P.J.; Kearsley, J.H.; Li, Y. PI3K/Akt/mTOR pathway inhibitors enhance radiosensitivity in radioresistant prostate cancer cells through inducing apoptosis, reducing autophagy, suppressing NHEJ and HR repair pathways. Cell Death Dis. 2014, 5, e1437. [Google Scholar] [CrossRef]
- Zois, C.E.; Koukourakis, M.I. Radiation-induced autophagy in normal and cancer cells: Towards novel cytoprotection and radio-sensitization policies? Autophagy 2009, 5, 442–450. [Google Scholar] [CrossRef]
- Ferrara, N.; Gerber, H.P.; LeCouter, J. The biology of VEGF and its receptors. Nat. Med. 2003, 9, 669–676. [Google Scholar] [CrossRef]
- Fu, Z.Z.; Sun, X.D.; Li, P.; Zhang, Z.; Li, G.Z.; Gu, T.; Shao, S.S. Relationship between serum VEGF level and radiosensitivity of patients with nonsmall cell lung cancer among asians: A meta-analysis. DNA Cell Biol. 2014, 33, 426–437. [Google Scholar] [CrossRef] [PubMed]
- Hu, X.; Xing, L.; Wei, X.; Liu, X.; Pang, R.; Qi, L.; Song, S. Nonangiogenic function of VEGF and enhanced radiosensitivity of HeLa cells by inhibition of VEGF expression. Oncol. Res. 2012, 20, 93–101. [Google Scholar] [CrossRef] [PubMed]
- Nicholson, D.W.; Ali, A.; Thornberry, N.A.; Vaillancourt, J.P.; Ding, C.K.; Gallant, M.; Gareau, Y.; Griffin, P.R.; Labelle, M.; Lazebnik, Y.A.; et al. Identification and inhibition of the ICE/CED-3 protease necessary for mammalian apoptosis. Nature 1995, 376, 37–43. [Google Scholar] [CrossRef]
- Santos, N.; Silva, R.F.; Pinto, M.; Silva, E.B.D.; Tasat, D.R.; Amaral, A. Active caspase-3 expression levels as bioindicator of individual radiosensitivity. Acad. Bras. Cienc. 2017, 89, 649–659. [Google Scholar] [CrossRef] [PubMed]
- Yang, X.H.; Edgerton, S.; Thor, A.D. Reconstitution of caspase-3 sensitizes MCF-7 breast cancer cells to radiation therapy. Int. J. Oncol. 2005, 26, 1675–1680. [Google Scholar] [CrossRef] [PubMed]
- Okamoto, A.; Demetrick, D.J.; Spillare, E.A.; Hagiwara, K.; Hussain, S.P.; Bennett, W.P.; Forrester, K.; Gerwin, B.; Serrano, M.; Beach, D.H.; et al. Mutations and altered expression of p16INK4 in human cancer. Proc. Natl. Acad. Sci. USA 1994, 91, 11045–11049. [Google Scholar] [CrossRef]
- Bockstaele, L.; Kooken, H.; Libert, F.; Paternot, S.; Dumont, J.E.; de Launoit, Y.; Roger, P.P.; Coulonval, K. Regulated activating Thr172 phosphorylation of cyclin-dependent kinase 4(CDK4): Its relationship with cyclins and CDK “inhibitors”. Mol. Cell Biol. 2006, 26, 5070–5085. [Google Scholar] [CrossRef]
- Narita, M.; Narita, M.; Krizhanovsky, V.; Nunez, S.; Chicas, A.; Hearn, S.A.; Myers, M.P.; Lowe, S.W. A novel role for high-mobility group a proteins in cellular senescence and heterochromatin formation. Cell 2006, 126, 503–514. [Google Scholar] [CrossRef] [PubMed]
- Matsumura, Y.; Yamagishi, N.; Miyakoshi, J.; Imamura, S.; Takebe, H. Increase in radiation sensitivity of human malignant melanoma cells by expression of wild-type p16 gene. Cancer Lett. 1997, 115, 91–96. [Google Scholar] [CrossRef]
- Lee, A.W.; Li, J.H.; Shi, W.; Li, A.; Ng, E.; Liu, T.J.; Klamut, H.J.; Liu, F.F. p16 gene therapy: A potentially efficacious modality for nasopharyngeal carcinoma. Mol. Cancer 2003, 2, 961–969. [Google Scholar]
- Dok, R.; Kalev, P.; Van Limbergen, E.J.; Asbagh, L.A.; Vazquez, I.; Hauben, E.; Sablina, A.; Nuyts, S. p16INK4a impairs homologous recombination-mediated DNA repair in human papillomavirus-positive head and neck tumors. Cancer Res. 2014, 74, 1739–1751. [Google Scholar] [CrossRef] [PubMed]
- Kang, S.; Tanaka, T.; Narazaki, M.; Kishimoto, T. Targeting Interleukin-6 Signaling in Clinic. Immunity 2019, 50, 1007–1023. [Google Scholar] [CrossRef]
- Chen, M.F.; Hsieh, C.C.; Chen, W.C.; Lai, C.H. Role of interleukin-6 in the radiation response of liver tumors. Int. J. Radiat. Oncol. Biol. Phys. 2012, 84, e621–e630. [Google Scholar] [CrossRef]
- Twillie, D.A.; Eisenberger, M.A.; Carducci, M.A.; Hseih, W.-S.; Kim, W.Y.; Simons, J.W. Interleukin-6: A candidate mediator of human prostate cancer morbidity. Urology 1995, 45, 542–549. [Google Scholar] [CrossRef]
- Shariat, S.F.; Andrews, B.; Kattan, M.W.; Kim, J.; Wheeler, T.M.; Slawin, K.M. Plasma levels of interleukin-6 and its soluble receptor are associated with prostate cancer progression and metastasis. Urology 2001, 58, 1008–1015. [Google Scholar] [CrossRef]
- George, D.J.; Halabi, S.; Shepard, T.F.; Sanford, B.; Vogelzang, N.J.; Small, E.J.; Kantoff, P.W. The prognostic significance of plasma interleukin-6 levels in patients with metastatic hormone-refractory prostate cancer: Results from cancer and leukemia group B 9480. Clin. Cancer Res. 2005, 11, 1815–1820. [Google Scholar] [CrossRef] [PubMed]
- Choi, Y.S.; Kim, S.; Oh, Y.S.; Cho, S.; Hoon Kim, S. Elevated serum interleukin-32 levels in patients with endometriosis: A cross-sectional study. Am. J. Reprod. Immunol. 2019, 82, e13149. [Google Scholar] [CrossRef]
- Berek, J.S.; Chung, C.; Kaldi, K.; Watson, J.M.; Knox, R.M.; Martínez-Maza, O. Serum interleukin-6 levels correlate with disease status in patients with epithelial ovarian cancer. Am. J. Obstet. Gynecol. 1991, 164, 1038–1043. [Google Scholar] [CrossRef]
- Scambia, G.; Testa, U.; Benedetti Panici, P.; Foti, E.; Martucci, R.; Gadducci, A.; Perillo, A.; Facchini, V.; Peschle, C.; Mancuso, S. Prognostic significance of interleukin 6 serum levels in patients with ovarian cancer. Br. J. Cancer 1995, 71, 354–356. [Google Scholar] [CrossRef] [PubMed]
- Tominaga, K.; Yoshimoto, T.; Torigoe, K.; Kurimoto, M.; Matsui, K.; Hada, T.; Okamura, H.; Nakanishi, K. IL-12 synergizes with IL-18 or IL-1beta for IFN-gamma production from human T cells. Int. Immunol. 2000, 12, 151–160. [Google Scholar] [CrossRef] [PubMed]
- Nakahara, H.; Song, J.; Sugimoto, M.; Hagihara, K.; Kishimoto, T.; Yoshizaki, K.; Nishimoto, N. Anti-interleukin-6 receptor antibody therapy reduces vascular endothelial growth factor production in rheumatoid arthritis. Arthritis Rheum. 2003, 48, 1521–1529. [Google Scholar] [CrossRef]
- Chen, M.F.; Lu, M.S.; Chen, P.T.; Chen, W.C.; Lin, P.Y.; Lee, K.D. Role of interleukin 1 beta in esophageal squamous cell carcinoma. J. Mol. Med. 2012, 90, 89–100. [Google Scholar] [CrossRef] [PubMed]
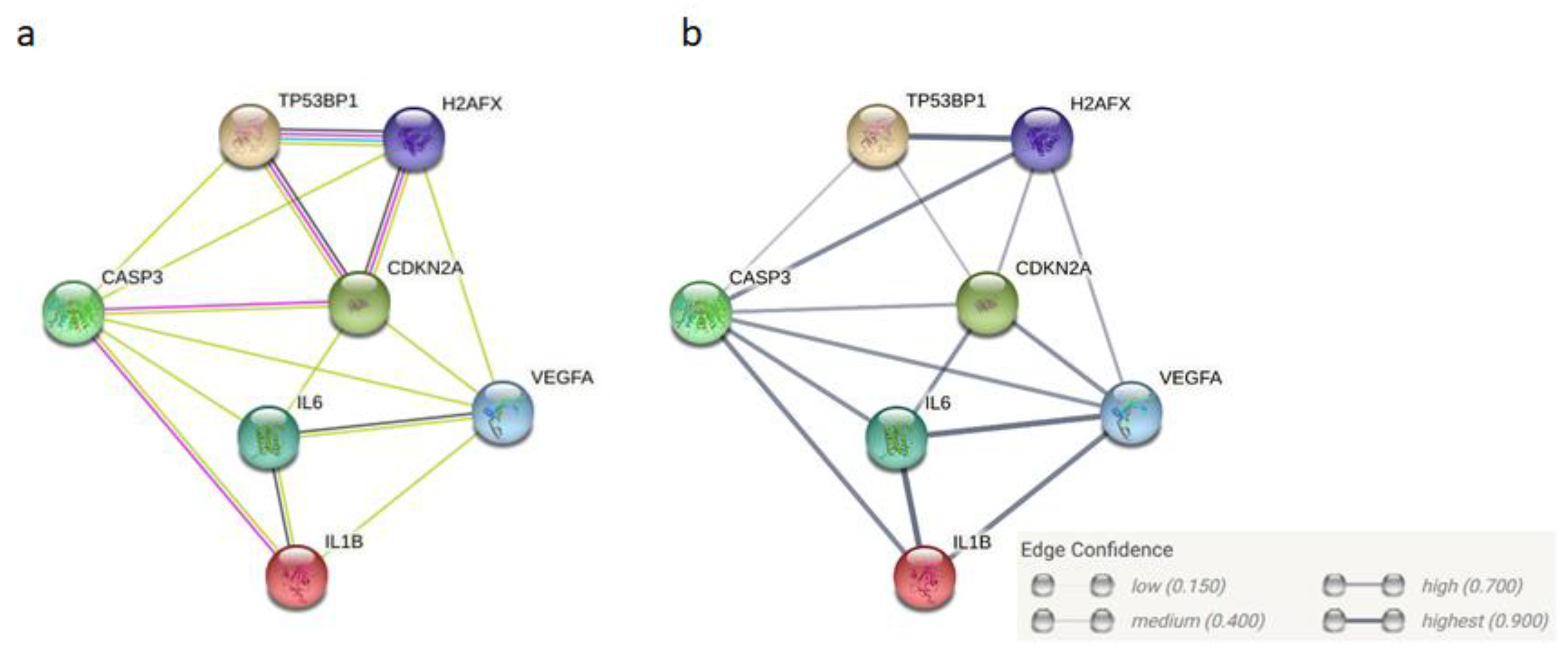
- Szklarczyk, D.; Franceschini, A.; Wyder, S.; Forslund, K.; Heller, D.; Huerta-Cepas, J.; Simonovic, M.; Roth, A.; Santos, A.; Tsafou, K.P.; et al. STRING v10: Protein-protein interaction networks, integrated over the tree of life. Nucleic Acids Res. 2015, 43, D447–D452. [Google Scholar] [CrossRef] [PubMed]
- Szklarczyk, D.; Gable, A.L.; Lyon, D.; Junge, A.; Wyder, S.; Huerta-Cepas, J.; Simonovic, M.; Doncheva, N.T.; Morris, J.H.; Bork, P.; et al. STRING v11: Protein-protein association networks with increased coverage, supporting functional discovery in genome-wide experimental datasets. Nucleic Acids Res. 2019, 47, D607–D613. [Google Scholar] [CrossRef]
- Wilhelm, M.; Schlegl, J.; Hahne, H.; Gholami, A.M.; Lieberenz, M.; Savitski, M.M.; Ziegler, E.; Butzmann, L.; Gessulat, S.; Marx, H.; et al. Mass-spectrometry-based draft of the human proteome. Nature 2014, 509, 582–587. [Google Scholar] [CrossRef]
- Gatti, R.A.; Boder, E.; Good, R.A. Immunodeficiency, radiosensitivity, and the XCIND syndrome. Immunol. Res. 2007, 38, 87–101. [Google Scholar] [CrossRef]
- Nahas, S.A.; Gatti, R.A. DNA double strand break repair defects, primary immunodeficiency disorders, and ‘radiosensitivity’. Curr. Opin. Allergy Clin. Immunol. 2009, 9, 510–516. [Google Scholar] [CrossRef]
- O’Driscoll, M.; Gennery, A.R.; Seidel, J.; Concannon, P.; Jeggo, P.A. An overview of three new disorders associated with genetic instability: LIG4 syndrome, RS-SCID and ATR-Seckel syndrome. DNA Repair 2004, 3, 1227–1235. [Google Scholar] [CrossRef]
- Royal, H.D. Effects of low level radiation-what’s new? Semin. Nucl. Med. 2008, 38, 392–402. [Google Scholar] [CrossRef] [PubMed]
- Preston, R.J. Children as a sensitive subpopulation for the risk assessment process. Toxicol. Appl. Pharm. 2004, 199, 132–141. [Google Scholar] [CrossRef] [PubMed]
- Kleinerman, R.A. Cancer risks following diagnostic and therapeutic radiation exposure in children. Pediatr. Radiol. 2006, 36 (Suppl. 2), 121–125. [Google Scholar] [CrossRef] [PubMed]
- Pearce, M.S.; Salotti, J.A.; Little, M.P.; McHugh, K.; Lee, C.; Kim, K.P.; Howe, N.L.; Ronckers, C.M.; Rajaraman, P.; Craft, A.W.; et al. Radiation exposure from CT scans in childhood and subsequent risk of leukaemia and brain tumours: A retrospective cohort study. Lancet 2012, 380, 499–505. [Google Scholar] [CrossRef]
- Mathews, J.D.; Forsythe, A.V.; Brady, Z.; Butler, M.W.; Goergen, S.K.; Byrnes, G.B.; Giles, G.G.; Wallace, A.B.; Anderson, P.R.; Guiver, T.A.; et al. Cancer risk in 680,000 people exposed to computed tomography scans in childhood or adolescence: Data linkage study of 11 million Australians. BMJ 2013, 346, f2360. [Google Scholar] [CrossRef]
- Brenner, D.J.; Doll, R.; Goodhead, D.T.; Hall, E.J.; Land, C.E.; Little, J.B.; Lubin, J.H.; Preston, D.L.; Preston, R.J.; Puskin, J.S.; et al. Cancer risks attributable to low doses of ionizing radiation: Assessing what we really know. Proc. Natl. Acad. Sci. USA 2003, 100, 13761–13766. [Google Scholar] [CrossRef]
- Narendran, N.; Luzhna, L.; Kovalchuk, O. Sex Difference of Radiation Response in Occupational and Accidental Exposure. Front. Genet. 2019, 10, 260. [Google Scholar] [CrossRef] [PubMed]
- Grant, E.J.; Brenner, A.; Sugiyama, H.; Sakata, R.; Sadakane, A.; Utada, M.; Cahoon, E.K.; Milder, C.M.; Soda, M.; Cullings, H.M.; et al. Solid Cancer Incidence among the Life Span Study of Atomic Bomb Survivors: 1958–2009. Radiat. Res. 2017, 187, 513–537. [Google Scholar] [CrossRef]
- Wakeford, R. Radiation effects: Modulating factors and risk assessment—An overview. Ann. ICRP 2012, 41, 98–107. [Google Scholar] [CrossRef] [PubMed]
- Preston, D.L.; Ron, E.; Tokuoka, S.; Funamoto, S.; Nishi, N.; Soda, M.; Mabuchi, K.; Kodama, K. Solid cancer incidence in atomic bomb survivors: 1958–1998. Radiat. Res. 2007, 168, 1–64. [Google Scholar] [CrossRef]
- Dreicer, M. Chernobyl: Consequences of the Catastrophe for People and the Environment. Environ. Health Perspect. 2010, 118, A500. [Google Scholar]
- Schmitz-Feuerhake, I.; Busby, C.; Pflugbeil, S. Genetic radiation risks: A neglected topic in the low dose debate. Environ. Health Toxicol. 2016, 31, e2016001. [Google Scholar] [CrossRef] [PubMed]
- Bentzen, S.M.; Overgaard, J. Patient-to-Patient Variability in the Expression of Radiation-Induced Normal Tissue Injury. Semin. Radiat. Oncol. 1994, 4, 68–80. [Google Scholar] [CrossRef]
- Fekrmandi, F.; Panzarella, T.; Dinniwell, R.E.; Helou, J.; Levin, W. Predictive factors for persistent and late radiation complications in breast cancer survivors. Clin. Transl. Oncol. 2020, 22, 360–369. [Google Scholar] [CrossRef] [PubMed]
- Barnett, G.C.; West, C.M.; Dunning, A.M.; Elliott, R.M.; Coles, C.E.; Pharoah, P.D.; Burnet, N.G. Normal tissue reactions to radiotherapy: Towards tailoring treatment dose by genotype. Nat. Rev. Cancer 2009, 9, 134–142. [Google Scholar] [CrossRef] [PubMed]
- Marsh, J.W.; Harrison, J.D.; Laurier, D.; Birchall, A.; Blanchardon, E.; Paquet, F.; Tirmarche, M. Doses and lung cancer risks from exposure to radon and plutonium. Int. J. Radiat. Biol. 2014, 90, 1080–1087. [Google Scholar] [CrossRef]
- Cardis, E.; Kesminiene, A.; Ivanov, V.; Malakhova, I.; Shibata, Y.; Khrouch, V.; Drozdovitch, V.; Maceika, E.; Zvonova, I.; Vlassov, O.; et al. Risk of thyroid cancer after exposure to 131I in childhood. J. Natl. Cancer Inst. 2005, 97, 724–732. [Google Scholar] [CrossRef]
- Belli, M.; Ottolenghi, A.; Weiss, W. The European strategy on low dose risk research and the role of radiation quality according to the recommendations of the “ad hoc” High Level and Expert Group (HLEG). Radiat. Environ. Biophys. 2010, 49, 463–468. [Google Scholar] [CrossRef] [PubMed]
- Kreuzer, M.; Auvinen, A.; Cardis, E.; Durante, M.; Harms-Ringdahl, M.; Jourdain, J.R.; Madas, B.G.; Ottolenghi, A.; Pazzaglia, S.; Prise, K.M.; et al. Multidisciplinary European Low Dose Initiative (MELODI): Strategic research agenda for low dose radiation risk research. Radiat. Environ. Biophys. 2018, 57, 5–15. [Google Scholar] [CrossRef] [PubMed]
- Kreuzer, M.; Sobotzki, C.; Schnelzer, M.; Fenske, N. Factors Modifying the Radon-Related Lung Cancer Risk at Low Exposures and Exposure Rates among German Uranium Miners. Radiat. Res. 2018, 189, 165–176. [Google Scholar] [CrossRef] [PubMed]
- Bassi, C.; Xavier, D.; Palomino, G.; Nicolucci, P.; Soares, C.; Sakamoto-Hojo, E.; Donadi, E. Efficiency of the DNA repair and polymorphisms of the XRCC1, XRCC3 and XRCC4 DNA repair genes in systemic lupus erythematosus. Lupus 2008, 17, 988–995. [Google Scholar] [CrossRef] [PubMed]
- Bashir, S.; Harris, G.; Denman, M.A.; Blake, D.R.; Winyard, P.G. Oxidative DNA damage and cellular sensitivity to oxidative stress in human autoimmune diseases. Ann. Rheum Dis. 1993, 52, 659–666. [Google Scholar] [CrossRef]
- Health Protection Agency. Human Radiosensitivity. Report of the Independent Advisory Group on Ionising Radiation. Available online: https://assets.publishing.service.gov.uk/government/uploads/system/uploads/attachment_data/file/333058/RCE-21_v2_for_website.pdf (accessed on 17 December 2020).
- Genetic susceptibility to cancer. International Commission on Radiological Protection (ICRP) publication 79. Approved by the Commission in May 1997. International Commission on Radiological Protection. Ann. ICRP 1998, 28, 1–157. [Google Scholar]
- Seibold, P.; Auvinen, A.; Averbeck, D.; Bourguignon, M.; Hartikainen, J.M.; Hoeschen, C.; Laurent, O.; Noël, G.; Sabatier, L.; Salomaa, S.; et al. Clinical and epidemiological observations on individual radiation sensitivity and susceptibility. Int. J. Radiat. Biol. 2019, 96, 324–339. [Google Scholar] [CrossRef]



{kind=link}
{kind=link}
{kind=link}
| (a) | ||||||||
|---|---|---|---|---|---|---|---|---|
| Author, Date | Title | Study Design | Sample-Size | Methods Used | Statistical Method | Repair Foci | Viability | Cell System |
| Vasireddy, 2010 [25] | H2AX phosphorylation screen of cells from radiosensitive cancer patients reveals a novel DNA double-strand break repair cellular phenotype | Cohort | 29 | IF | nr | γH2AX | (RTOG) | lymphoblastoid cell lines (LCLs) |
| Bourton, 2011 [26] | Prolonged expression of the γ-H2AX DNA repair biomarker correlates with excess acute and chronic toxicity from radiotherapy treatment | Cohort | 30 | FACS | unpaired t-test | γH2AX | (RTOG) | lymphocytes |
| Goutham, 2012 [33] | DNA double-strand break analysis by γ-H2AX foci: a useful method for determining the overreactors to radiation-induced acute reactions among head-and-neck cancer patients | Cohort | 54 | IF | nr | γH2AX | (RTOG) | lymphocytes |
| Djuzenova, 2013 [32] | Radiosensitivity in breast cancer assessed by the histone γ-H2AX and 53BP1 foci | Cohort | 69 | IF | Student’s t-test or one way ANOVA | γH2AX, 53BP1 | (RTOG) | PBMCs |
| Mumbrekar, 2013 [27] | Influence of double-strand break repair on radiation therapy-induced acute skin reactions in breast cancer patients | Cohort | 118 | IF | t test, ANOVA followed by Tukey multiple comparison tests and Pearson correlation test | γH2AX | (RTOG) | lymphocytes |
| Oorschot, 2013 [24] | Reduced Activity of Double-Strand Break Repair Genes in Prostate Cancer Patients With Late Normal Tissue Radiation Toxicity | Cohort | 61 | IF | Continuous variables: Shapiro-Wilk test, normal distributed data: unpaired Student t test, non-normal distributed data: Mann–Whitney test | γH2AX | Late toxicity using EORTC | lymphocytes |
| Granzotto, 2015 [31] | Influence of Nucleoshuttling of the ATM Protein in the Healthy Tissues Response to Radiation Therapy: Toward a Molecular Classification of Human Radiosensitivity | Cohort | 117 | IF | ANOVA | γH2AX, pATM | Common Terminology CTCAE, RTOG | fibroblasts |
| Pouliliou, 2015 [28] | Survival Fraction at 2 Gy and γH2AX Expression Kinetics in Peripheral Blood Lymphocytes From Cancer Patients: Relationship With Acute Radiation-Induced Toxicities | Cohort | 89 | WB | nr | γH2AX | Common Terminology CTCAE, Trypan Blue assay | PBMCs |
| Lobachevsky, 2016 [29] | Compromised DNA repair as a basis for identification of cancer radiotherapy patients with extreme radiosensitivity | Cohort | 28 | IF | Unpaired t-test, Mann–Whitney test | γH2AX | (RTOG) | lymphocytes, hair follicles |
| Buchbinder, 2018 [30] | Application of a radiosensitivity flow assay in a patient with DNA ligase 4 deficiency | Cohort | 11 | IF | nr | γH2AX | known sensitivity LIG4-SCID | T cells |
| Vandersickel, 2010 [34] | Early Increase in Radiation-induced γH2AX Foci in a HumanKu70/80 Knockdown Cell Line Characterised by an Enhanced Radiosensitivity | model system | 1 | IF | nr | γH2AX | known sensitivity Ku70i | LVTHM cells synchronised in the G0–G1 phase, Ku70i cells synchronised in the G0–G1 phase |
| Martin, 2011 [36] | Assessing ’radiosensitivity’ with kinetic profiles of γ-H2AX, 53BP1 and BRCA1 foci | model system | 15 | IF | unpaired t test | γH2AX, 53BP1 | clonogenic survival | LCL |
| Martin, 2014 [35] | Homozygous mutation of MTPAP causes cellular radiosensitivity and persistent DNA double-strand breaks | model system | 4 | IF | Student’s t-test | γH2AX | clonogenic survival | LCL |
| Minafra, 2015 [37] | Gene Expression Profiling of MCF10A Breast Epithelial Cells Exposed to IOERT | model system | 1 | IF | nr | γH2AX | clonogenic survival | MCF10A |
| Miyake, 2019 [38] | DNA Damage Response After Ionizing Radiation Exposure in Skin Keratinocytes Derived from Human-Induced Pluripotent Stem Cells | model system | 1 | IF | Student’s t test (1-tailed) | γH2AX, 53BP1 | Cell survival WST-8 assay; TUNEL assay | Normal human skin fibroblast NB1RGB, iPSCs NB1RGB C2, NB1RGB KCs 1stP, NB1RGB KCs 2ndP, NB1RGB KCs 3rdP |
| Nguyen, 2019 [39] | Human CCR6+ Th17 Lymphocytes Are Highly Sensitive to Radiation-Induced Senescence and Are a Potential Target for Prevention of Radiation-Induced Toxicity | model system | 32 | IF | two-tailed Mann–Whitney U-test, Kruskal–Wallis test | γH2AX | Annexin V-FITC; Senescence-associated β-Galactosidase | Treg, CCR6+Th17, CCR6negTh |
| (b) | ||||||||
| Author, Date | Title | Study Design | Sample-Size | Methods Used | Statistical Method | Results (Protein Name) | Viability | Cell System |
| Braicu, 2014 [40] | Role of serum VEGFA, TIMP2, MMP2, and MMP9 in Monitoring Response to Adjuvant Radiochemotherapy in Patients with Primary Cervical Cancer – Results of a Companion Protocol of the Randomised NOGGO-AGO Phase III Clinical Trial | Cohort | 72 | ELISA | Fisher’s exact test | VEGFA, TIMP2, MMP2, MMP9 | overall survival | Serum |
| Rodriguez-Gil, 2014 [41] | Inflammatory Biomarker C-Reactive Protein and Radiotherapy-Induced Early Adverse Skin Reactions in Patients with Breast Cancer | Cohort | 159 | ELISA | two-sided Student’s t-test | C-reactive protein (CRP) | EASR | plasma |
| Skiöld, 2014 [42] | Unique proteomic signature for radiation sensitive patients; a comparative study between normo-sensitive and radiation sensitive breast cancer patients | Cohort | 17 | LC-MS/MS | Student’s t-test | 8-oxo-dG, BLVRB, PRDX2, SOD1, CA1, PARK7, SH3BGRL3 | RTOG | blood/leukocytes (RTOG 0), blood/leukocytes (RTOG 4) |
| Yu, 2018 [43] | Cofilin-2 Acts as a Marker for Predicting Radiotherapy Response and Is a Potential Therapeutic Target in Nasopharyngeal Carcinoma | Cohort | 70 | ELISA | Wilcoxon rank-sum test, t test or one-way analysis of variance (ANOVA) | Cofilin-2 | Patients were divided into radiosensitivity and radio-resistance groups according to therapeutic effects | Serum |
| Lacombe, 2019 [44] | Quantitative proteomic analysis reveals AK2 as potential biomarker for late normal tissue radiotoxicity | Cohort | 5 | WB | Mann–Whitney test | adenylate kinase 2 (AK2), annexin A1 (ANXA1), isocitrate dehydrogenase 2 (IDH2), HSPA8, Nox4 | RILA | T lymphocytes (Grade > 2 breast fibrosis+), T lymphocytes (Grade < 2 breast fibrosis+) |
| Cao, 2011 [45] | Different radiosensitivity of CD4+CD25+ regulatory T cells and effector T cells to low dose gamma irradiation in vitro | model system | 5 | FACS, Luminex | Wilcoxon’s signed rank test | Caspase 3, Bax, IL-1 Beta, IL-2, IL-4, IL-6, IL-10, Interferon Gamma, TNF alpha | Annexin V-FITC | CD4+CD25+ regulatory T cells and effector T cells |
| Park, 2012 [46] | Radio-sensitivities and angiogenic signaling pathways of irradiated normal endothelial cells derived from diverse human organs | model system | 1 | ELISA | Student’s t-test | angiostatin | clonogenic survival | HHSEC, HDMEC |
| Fekete, 2015 [47] | Effect of High-Dose Irradiation on Human Bone-Marrow-Derived Mesenchymal Stromal Cells | model system | nr | Luminex | unpaired, two sided Student’s t-test | PDGF-AA, PDGF-AB/BB, GRO, IL-6, VEGF | CyQUANT Cell Proliferation Assay, Trypan blue staining, colony formation | MSCs |
| Minafra, 2015 [37] | Gene Expression Profiling of MCF10A Breast Epithelial Cells Exposed to IOERT | model system | 1 | WB | nr | PARP, FAS, Pro-Caspase 8, PLK1, P53, p-EGFR, EGFR, c-MYC, | clonogenic survival | MCF10A cell line |
| Nishad, 2015 [48] | Dynamic changes in the proteome of human peripheral blood mononuclear cells with low dose ionizing radiation Radiotherapy-Induced Early Adverse Skin Reactions in | model system | 8 | 2DE-MS, WB | Student’s t-test | GRP78, HSP90, PDIA3, PRDX6 | trypan blue, PI Staining, alkaline comet assay | PBMCs |
| Shimura, 2015 [49] | Nuclear accumulation of cyclin D1 following long-term fractionated exposures to low-dose ionizing radiation in normal human diploid cells | model system | 1 | WB | Student’s t-test | cyclin D1 | cell growth assay | WI-38 (detergent insoluble fraction) |
| Yim, 2017 [50] | Phosphoprotein profiles of candidate markers for early cellular responses to low-dose γ-radiation in normal human fibroblast cells | model system | 1 | WB, antibody microarray | Student’s t-test | Phospho-Gab2 (Tyr643), Phospho-P95/NBS (Ser343), Phospho-BTK (Tyr550), Phospho-Elk1 (Ser383), Phospho-ETK (Tyr40), Phospho-CaMK4 (Thr196/200), Phospho-MEK1 (Thr298), Phospho-PLCG1 (Tyr1253), Phospho-IRS-1 (Ser612), Phospho-TFII-I (Tyr248), Phospho-IKK-alpha/beta (Ser176/177), Phospho-MEK1 (Thr286), Phospho-Pyk2 (Tyr580), Phospho-Keratin 8 (Ser431), Phospho-ERK3 (Ser189), Phospho-Chk1 (Ser296), Phospho-CBL (Tyr700), Phospho-BTK (Tyr550), Phospho-LIMK1/2 (Thr508/505), p-BTK(Tyr550)/BTK, p-Gab2(Tyr643)/Gab2, p-BTK(Tyr550)/BTK, p-Gab2(Tyr643)/Gab2, | MTT | MRC5, NHDF |
| Miyake, 2019 [38] | DNA Damage Response After Ionizing Radiation Exposure in Skin Keratinocytes Derived from Human-Induced Pluripotent Stem Cells | model system | 1 | IF | Student’s t test (1-tailed) | p16 | Cell survival WST-8 assay, TUNEL assay | Skin keratinocytes were derived from iPSCs |
| Nguyen, 2019 [39] | Human CCR6+Th17 lymphocytes are highly sensitive to radiation-induced senescence and are a potential target for prevention of radiation-induced toxicity | model system | 32 | IF, Luminex | two-tailed Mann–Whitney U-test, Kruskal–Wallis test | Caspase 3, p16Ink4a, p21Cdkn1a, IL-1 Beta, VEGF-A, IL-8, H2A.J | Annexin V-FITC, Senescence-associated β-Galactosidase | CCR6+Th17 lymphocytes |
| Moertl, 2020 [51] | Radiation Exposure of Peripheral Mononuclear Blood Cells Alters the Composition and Function of Secreted Extracellular Vesicles | model system | 5 | LC-MS/MS | two-sided Student’s t-test | hemopexin (HPX), syntaxin-binding protein 3 (STXBP3), proteasome subunit alpha type-6 (PSMA6) | sub-G1 fraction, Caspase 3 activity | PBMC-derived EVs |
| Risk of Bias Domains and Ratings | Miyake, 2019 | Cao, 2011 | Nguyen, 2019 | Minafra, 2015 | Yim, 2017 | Braicu, 2014 | Park, 2012 | Fekete, 2015 | Shumura, 2015 | Lacombe, 2019 | Skiöld, 2014 | Nishad, 2015 | Rodruiguez-Gil, 2014 | Moertl, 2020 | Granzotto, 2016 | Yu, 2018 | |
|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|
| Key criteria | Can we be confident in the exposure characterisation? | -- | + | + | ++ | ++ | -- | -- | - | + | - | + | + | - | ++ | + | -- |
| Can we be confident in the outcome assessment? | - | -- | - | - | + | + | - | + | + | ++ | + | + | ++ | - | + | ++ | |
| Did the study design or analyses account for important confounding and modifying variables? | + | - | ++ | + | ++ | -- | - | + | - | + | - | - | ++ | - | - | + | |
| Other RoB criterion | Was administered dose or exposure level adequately randomised? | - | - | - | - | - | - | - | - | - | -- | - | + | - | - | - | + |
| Was allocation to study groups adequately concealed? | - | - | - | - | - | - | - | - | - | - | - | - | - | - | - | - | |
| Did selection of study participants result in appropriate comparison groups? | ++ | ++ | - | ++ | ++ | ++ | + | + | - | + | - | + | ++ | ++ | - | + | |
| Were experimental conditions identical across study groups? | - | ++ | - | - | - | - | - | -- | - | + | ++ | - | ++ | - | ++ | ||
| Were the research personnel and human subjects blinded to the study group in the study? | - | - | - | - | - | - | - | - | - | - | - | - | - | - | - | - | |
| Were outcome data complete without attrition or exclusion from analysis? | - | + | + | ++ | - | ++ | + | ++ | ++ | ++ | ++ | ++ | ++ | ++ | - | ++ | |
| Were all the measured outcomes reported? | + | ++ | ++ | ++ | ++ | ++ | ++ | ++ | - | ++ | ++ | ++ | ++ | ++ | ++ | ++ | |
| Were there no other potential threat to internal validity (e.g., statistical methods were appropriate and researchers adhered to study protocol? | -- | - | ++ | - | + | - | - | ++ | + | ++ | - | + | + | ++ | + | ++ | |
| Final Category | T2 | T2 | T2 | T2 | T2 | T2 | T3 | T2 | T2 | T2 | T2 | T2 | T2 | T2 | T2 | T2 | |
| Priority Group | Author, Date | Marker | Outcome | Cell System |
|---|---|---|---|---|
| 1 | Braicu, 2014 | VEGFA | decrease in VEGFA concentration leads to increase in survival, >500 pg/mL negative influence on survival | Serum |
| Fekete, 2015 | VEGFA | nr | MSCs | |
| Nguyen, 2019 | VEGFA | resistant compared to Treg | CCR6 + Th17 | |
| 2 | Cao, 2011 | Caspase 3 # | radiosensitive | CD4 + CD25 + Treg cells |
| Nguyen, 2019 | Caspase 3 | resistant compared to Treg | CCR6 + Th17 | |
| 2 | Miyake, 2019 | p16 | resistant compared to primary fibroblasts (IR-induced senescence) | NB1RGB KCs 1stP, 2ndP and 3rdP |
| Nguyen, 2019 | p16 | resistant compared to Treg (IR-induced senescence) | CCR6 + Th17 | |
| 2 | Cao, 2011 | IL-6 | radiosensitive | CD4 + CD25 + Treg cells |
| Fekete, 2015 | IL-6 | nr | MSCs | |
| 2 | Cao, 2011 | IL-1Beta | radiosensitive | CD4 + CD25 + Treg cells |
| Nguyen, 2019 | IL-1Beta | sensitive compared to CCR6 + Th17 | CCR6negTh |
| Author, Date | Initial Confidence by Features of Study Design | Initial Confidence Rating | Factors Decreasing Confidence | Factors Increasing Confidence | Final Confidence | |||
|---|---|---|---|---|---|---|---|---|
| Controlled Exposure | Exposure Prior to Outcome | Individual Outcome Data | Use of Comparison Groups | |||||
| Fekete, 2015 | × | √ | × | × | Very low | Risk of bias Unexplained consistency Indirectness Imprecision | Magnitude effect Dose Response Residual Confounding Consistency | Very low |
| Braicu, 2014 | × | √ | √ | √ | Moderate | Low | ||
| Nguyen, 2019 | √ | √ | √ | × | Moderate | Moderate | ||
| Cao, 2011 | √ | √ | √ | √ | High | High | ||
| Miyake, 2019 | × | √ | √ | × | Low | Low | ||
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Subedi, P.; Gomolka, M.; Moertl, S.; Dietz, A. Ionizing Radiation Protein Biomarkers in Normal Tissue and Their Correlation to Radiosensitivity: A Systematic Review. J. Pers. Med. 2021, 11, 140. https://doi.org/10.3390/jpm11020140
Subedi P, Gomolka M, Moertl S, Dietz A. Ionizing Radiation Protein Biomarkers in Normal Tissue and Their Correlation to Radiosensitivity: A Systematic Review. Journal of Personalized Medicine. 2021; 11(2):140. https://doi.org/10.3390/jpm11020140
Chicago/Turabian StyleSubedi, Prabal, Maria Gomolka, Simone Moertl, and Anne Dietz. 2021. "Ionizing Radiation Protein Biomarkers in Normal Tissue and Their Correlation to Radiosensitivity: A Systematic Review" Journal of Personalized Medicine 11, no. 2: 140. https://doi.org/10.3390/jpm11020140
APA StyleSubedi, P., Gomolka, M., Moertl, S., & Dietz, A. (2021). Ionizing Radiation Protein Biomarkers in Normal Tissue and Their Correlation to Radiosensitivity: A Systematic Review. Journal of Personalized Medicine, 11(2), 140. https://doi.org/10.3390/jpm11020140