
Influence of Genetic Polymorphisms on Clinical Outcomes of Glatiramer Acetate in Multiple Sclerosis Patients

 ,
,  and
and
Abstract
1. Introduction
2. Materials and Methods
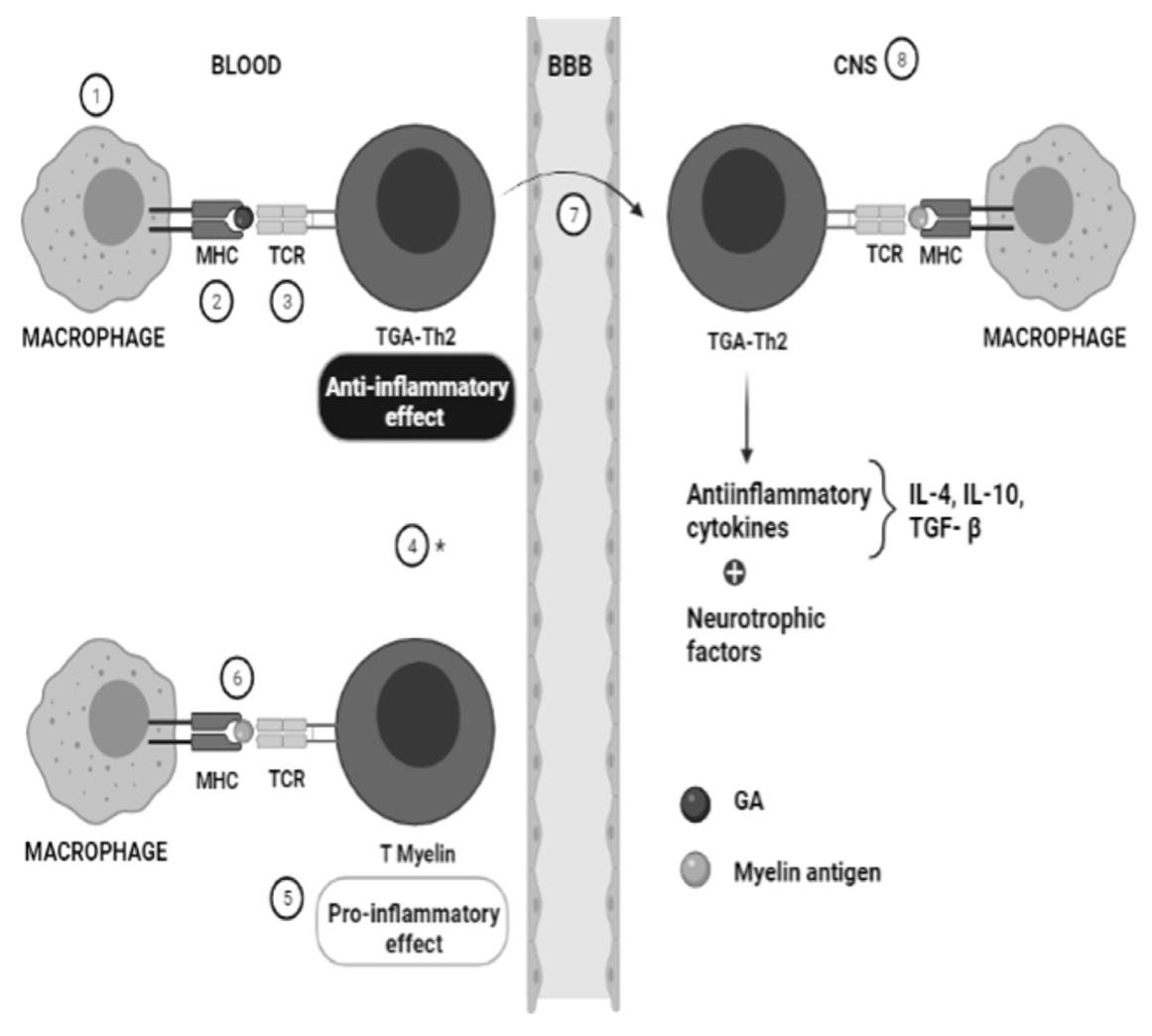
3. Pharmacogenetics of Glatiramer Acetate in MS
3.1. Multiple Sclerosis Related Genes
3.1.1. Arachidonate 5-Lipoxygenase-Activating Protein; ALOX5AP
3.1.2. CD86 Antigen; CD86
3.1.3. Eomesodermin; EOMES
3.1.4. Interleukin 1 Receptor-like 1; IL1RL1
3.1.5. Interleukin 12 Receptor, Beta-2; IL12RB2
3.1.6. Interleukin 22 Receptor, Alpha-2; IL22RA2
3.1.7. Membrane-Associated Guanylate Kinase, WW and PDZ Domains-Containing, 2; MAGI2
3.1.8. Oncogene PVT1; PVT1
3.1.9. RET Finger Protein-like 3; RFPL3
3.1.10. Solute Carrier Family 1 (Glutamate/Neutral Amino Acid Transporter), Member 4; SLC1A4
3.1.11. T-Cell Receptor Beta Chain Constant Region 1; TRBC1
3.2. Glatiramer Acetate Related Genes
3.2.1. Cathepsin S; CTSS
3.2.2. C-C Motif Chemokine Receptor 5; CCR5
3.2.3. C-Type Lectin Domain Family 16, Member A; CLEC16A
3.2.4. Fas Cell Surface Death Receptor; FAS (CD95)
3.2.5. Leucine Zipper- and Sterile Alpha Motif-Containing Kinase; ZAK
3.2.6. Major Histocompatibility Complex, Class II, DR Beta-1; HLA-DRB1
3.2.7. Myelin Basic Protein; MBP
3.2.8. Protein-Tyrosine Phosphatase, Receptor-Type, T; PTPRT
3.2.9. UV Radiation Resistance-Associated Gene; UVRAG
4. Conclusions
Author Contributions
Funding
Informed Consent Statement
Conflicts of Interest
Abbreviations
| ALOX5AP | Arachidonate 5-lipoxygenase-activating protein gene |
| APC | Antigen-presenting cell |
| CCR5 | C-C motif chemokine receptor 5 gene |
| CIS | Clinically isolated syndrome |
| CLEC16A | C-type lectin domain family 16 member A gene |
| CNS | Central nervous system |
| CTSS | Cathepsin S gene |
| EOMES | Eomesodermin gene |
| FORTE | Forty mg Efficacy of glatiramer acetate |
| GA | Glatiramer acetate |
| GALA | Glatiramer Acetate Low-frequency Administration |
| HLA-DRB1 | Major histocompatibility complex class II DR beta-1 gene |
| IL1RL1 | Interleukin 1 receptor-like 1 gene |
| IL22RA2 | Interleukin 22 receptor alpha-2 gene |
| JNK/SAPK1 | C-Jun N-terminal kinase 1/stress-activated protein kinase |
| MAGI2 | Membrane-associated guanylate kinase WW and PDZ domains-containing 2 gene |
| MBP | Myelin basic protein |
| MS | Multiple sclerosis |
| NFĸΒ | Nuclear factor kappa light chain in β cells |
| PPMS | Primary progressive multiple sclerosis |
| PTP | Protein tyrosine phosphatase |
| PTPRT | Protein-tyrosine phosphatase receptor-type T gene |
| PVT1 | Plasmacytoma variant translocation oncogene |
| RFPL3 | RET finger protein-like 3 gene |
| RRMS | Relapsing-remitting multiple sclerosis |
| SLC1A4 | Solute carrier family 1 member 4 gene |
| SPMS | Secondary progressive multiple sclerosis |
| Th2 | T helper type 2 |
| TRBC1 | T-cell receptor beta chain constant region 1 gene |
| UVRAG | UV radiation resistance-associated gene |
| ZAK | Leucine zipper- and sterile alpha motif-containing kinase gene |
References
- Freal, J.E.; Kraft, G.H.; Coryell, J.K. Symptomatic fatigue in multiple sclerosis. Arch. Phys. Med. Rehabil. 1984, 65, 135–138. [Google Scholar]
- Hauser, S.L.; Oksenberg, J.R. The neurobiology of multiple sclerosis: Genes, inflammation, and neurodegeneration. Neuron 2006, 52, 61–76. [Google Scholar] [CrossRef]
- Krupp, L.B.; Alvarez, L.A.; LaRocca, N.G.; Scheinberg, L.C. Fatigue in multiple sclerosis. Arch. Neurol. 1988, 45, 435–437. [Google Scholar] [CrossRef]
- Lucchinetti, C.; Bruck, W.; Parisi, J.; Scheithauer, B.; Rodriguez, M.; Lassmann, H. Heterogeneity of multiple sclerosis lesions: Implications for the pathogenesis of demyelination. Ann. Neurol. 2000, 47, 707–717. [Google Scholar] [CrossRef]
- Hestvik, A.L. The double-edged sword of autoimmunity: Lessons from multiple sclerosis. Toxins 2010, 2, 856–877. [Google Scholar] [CrossRef] [PubMed]
- Tanaka, M.; Vecsei, L. Monitoring the Redox Status in Multiple Sclerosis. Biomedicines 2020, 8, 406. [Google Scholar] [CrossRef] [PubMed]
- Tanaka, M.; Toldi, J.; Vecsei, L. Exploring the Etiological Links behind Neurodegenerative Diseases: Inflammatory Cytokines and Bioactive Kynurenines. Int. J. Mol. Sci. 2020, 21, 2431. [Google Scholar] [CrossRef] [PubMed]
- Arslan, B.; Arslan, G.A.; Tuncer, A.; Karabudak, R.; Dinçel, A.S. Evaluation of Thiol Homeostasis in Multiple Sclerosis and Neuromyelitis Optica Spectrum Disorders. Front. Neurol. 2021, 12, 716195. [Google Scholar] [CrossRef] [PubMed]
- Ömerhoca, S.; Akkaş, S.Y.; İçen, N.K. Multiple Sclerosis: Diagnosis and Differential Diagnosis. Arch. Neuropsychiatry 2018, 55, S1–S9. [Google Scholar] [CrossRef] [PubMed]
- Brownlee, W.J. Differential diagnosis of multiple sclerosis. Better Explan. Clin. Pract. 2019, 92, 1037–1038. [Google Scholar] [CrossRef]
- Luca, M.; Chisari, C.G.; Zanghì, A.; Patti, F. Early-Onset Alcohol Dependence and Multiple Sclerosis: Diagnostic Challenges. Int. J. Environ. Res. Public Health 2021, 18, 5588. [Google Scholar] [CrossRef] [PubMed]
- Toscano, S.; Patti, F. CSF biomarkers in multiple sclerosis: Beyond neuroinflammation. Neuroimmunol. Neuroinflamm. 2021, 8, 14–41. [Google Scholar] [CrossRef]
- Browne, P.; Chandraratna, D.; Angood, C.; Tremlett, H.; Baker, C.; Taylor, B.V.; Thompson, A.J. Atlas of Multiple Sclerosis 2013: A growing global problem with widespread inequity. Neurology 2014, 83, 1022–1024. [Google Scholar] [CrossRef] [PubMed]
- Pugliatti, M.; Rosati, G.; Carton, H.; Riise, T.; Drulovic, J.; Vecsei, L.; Milanov, I. The epidemiology of multiple sclerosis in Europe. Eur. J. Neurol. 2006, 13, 700–722. [Google Scholar] [CrossRef] [PubMed]
- Compston, A.; Coles, A. Multiple sclerosis. Lancet 2008, 372, 1502–1517. [Google Scholar] [CrossRef]
- Lublin, F.D.; Reingold, S.C.; Cohen, J.A.; Cutter, G.R.; Sorensen, P.S.; Thompson, A.J.; Wolinsky, J.S.; Balcer, L.J.; Banwell, B.; Barkhof, F.; et al. Defining the clinical course of multiple sclerosis: The 2013 revisions. Neurology 2014, 83, 278–286. [Google Scholar] [CrossRef] [PubMed]
- Grossman, I.; Avidan, N.; Singer, C.; Goldstaub, D.; Hayardeny, L.; Eyal, E.; Ben-Asher, E.; Paperna, T.; Pe’er, I.; Lancet, D.; et al. Pharmacogenetics of glatiramer acetate therapy for multiple sclerosis reveals drug-response markers. Pharm. Genom. 2007, 17, 657–666. [Google Scholar] [CrossRef]
- Grossman, I.; Knappertz, V.; Laifenfeld, D.; Ross, C.; Zeskind, B.; Kolitz, S.; Ladkani, D.; Hayardeny, L.; Loupe, P.; Laufer, R.; et al. Pharmacogenomics strategies to optimize treatments for multiple sclerosis: Insights from clinical research. Prog. Neurobiol. 2017, 152, 114–130. [Google Scholar] [CrossRef]
- Castro-Borrero, W.; Graves, D.; Frohman, T.C.; Flores, A.B.; Hardeman, P.; Logan, D.; Orchard, M.; Greenberg, B.; Frohman, E.M. Current and emerging therapies in multiple sclerosis: A systematic review. Adv. Neurol. Disord. 2012, 5, 205–220. [Google Scholar] [CrossRef]
- Huisman, E.; Papadimitropoulou, K.; Jarrett, J.; Bending, M.; Firth, Z.; Allen, F.; Adlard, N. Systematic literature review and network meta-analysis in highly active relapsing-remitting multiple sclerosis and rapidly evolving severe multiple sclerosis. BMJ Open 2017, 7, e013430. [Google Scholar] [CrossRef]
- Findling, O.; Sellner, J. Second-generation immunotherapeutics in multiple sclerosis: Can we discard their precursors? Drug Discov. Today 2021, 26, 416–428. [Google Scholar] [CrossRef]
- Doshi, A.; Chataway, J. Multiple sclerosis, a treatable disease. Clin. Med. 2017, 17, 530–536. [Google Scholar] [CrossRef] [PubMed]
- Coyle, P.K. Pharmacogenetic Biomarkers to Predict Treatment Response in Multiple Sclerosis: Current and Future Perspectives. Mult. Scler. Int. 2017, 2017, 6198530. [Google Scholar] [CrossRef]
- Aharoni, R. The mechanism of action of glatiramer acetate in multiple sclerosis and beyond. Autoimmun. Rev. 2013, 12, 543–553. [Google Scholar] [CrossRef] [PubMed]
- Tennakoon, D.K.; Mehta, R.S.; Ortega, S.B.; Bhoj, V.; Racke, M.K.; Karandikar, N.J. Therapeutic induction of regulatory, cytotoxic CD8+ T cells in multiple sclerosis. J. Immunol. 2006, 176, 7119–7129. [Google Scholar] [CrossRef] [PubMed]
- Tsareva, E.; Kulakova, O.; Boyko, A.; Favorova, O. Pharmacogenetics of multiple sclerosis: Personalized therapy with immunomodulatory drugs. Pharm. Genom. 2016, 26, 103–115. [Google Scholar] [CrossRef] [PubMed]
- Montalban, X.; Gold, R.; Thompson, A.J.; Otero-Romero, S.; Amato, M.P.; Chandraratna, D.; Clanet, M.; Comi, G.; Derfuss, T.; Fazekas, F.; et al. ECTRIMS/EAN Guideline on the pharmacological treatment of people with multiple sclerosis. Mult. Scler. 2018, 24, 96–120. [Google Scholar] [CrossRef] [PubMed]
- Boster, A.L.; Ford, C.C.; Neudorfer, O.; Gilgun-Sherki, Y. Glatiramer acetate: Long-term safety and efficacy in relapsing-remitting multiple sclerosis. Expert Rev. Neurother. 2015, 15, 575–586. [Google Scholar] [CrossRef] [PubMed]
- Weinstock-Guttman, B.; Nair, K.V.; Glajch, J.L.; Ganguly, T.C.; Kantor, D. Two decades of glatiramer acetate: From initial discovery to the current development of generics. J. Neurol. Sci. 2017, 376, 255–259. [Google Scholar] [CrossRef]
- Ganji, A.; Monfared, M.E.; Shapoori, S.; Nourbakhsh, P.; Ghazavi, A.; Ghasami, K.; Mosayebi, G. Effects of interferon and glatiramer acetate on cytokine patterns in multiple sclerosis patients. Cytokine 2019, 126, 154911. [Google Scholar] [CrossRef]
- Johnson, K.P.; Brooks, B.R.; Cohen, J.A.; Ford, C.C.; Goldstein, J.; Lisak, R.P.; Myers, L.W.; Panitch, H.S.; Rose, J.W.; Schiffer, R.B. Copolymer 1 reduces relapse rate and improves disability in relapsing-remitting multiple sclerosis: Results of a phase III multicenter, double-blind placebo-controlled trial. The Copolymer 1 Multiple Sclerosis Study Group. Neurology 1995, 45, 1268–1276. [Google Scholar] [CrossRef]
- Comi, G.; Filippi, M.; Wolinsky, J.S. European/Canadian multicenter, double-blind, randomized, placebo-controlled study of the effects of glatiramer acetate on magnetic resonance imaging--measured disease activity and burden in patients with relapsing multiple sclerosis. European/Canadian Glatiramer Acetate Study Group. Ann. Neurol. 2001, 49, 290–297. [Google Scholar]
- Fusco, C.; Andreone, V.; Coppola, G.; Luongo, V.; Guerini, F.; Pace, E.; Florio, C.; Pirozzi, G.; Lanzillo, R.; Ferrante, P.; et al. HLA-DRB1*1501 and response to copolymer-1 therapy in relapsing-remitting multiple sclerosis. Neurology 2001, 57, 1976–1979. [Google Scholar] [CrossRef]
- Bovis, F.; Kalincik, T.; Lublin, F.; Cutter, G.; Malpas, C.; Horakova, D.; Havrdova, E.K.; Trojano, M.; Prat, A.; Girard, M.; et al. Treatment Response Score to Glatiramer Acetate or Interferon Beta-1a. Neurology 2021, 96, e214–e227. [Google Scholar] [CrossRef]
- Drew, L. Pharmacogenetics: The right drug for you. Nature 2016, 537, S60–S62. [Google Scholar] [CrossRef] [PubMed]
- Zarzuelo-Romero, M.J.; Pérez-Ramírez, C.; Carrasco-Campos, M.I.; Sánchez-Martín, A.; Calleja Hernández, M.A.; Ramírez-Tortosa, M.C.; Jiménez-Morales, A. Therapeutic Value of Single Nucleotide Polymorphisms on the Efficacy of New Therapies in Patients with Multiple Sclerosis. J. Pers. Med. 2021, 11, 335. [Google Scholar] [CrossRef] [PubMed]
- Carrasco-Campos, M.I.; Pérez-Ramírez, C.; Macías-Cortés, E.; Puerta-García, E.; Sánchez-Pozo, A.; Arnal-García, C.; Barrero-Hernández, F.J.; Calleja-Hernández, M.; Jiménez-Morales, A.; Cañadas-Garre, M. Pharmacogenetic Predictors of Response to Interferon Beta Therapy in Multiple Sclerosis. Mol. Neurobiol. 2021, 58, 4716–4726. [Google Scholar] [CrossRef]
- Martínez-Aguilar, L.; Pérez-Ramírez, C.; Maldonado-Montoro, M.D.M.; Carrasco-Campos, M.I.; Membrive-Jiménez, C.; Martínez-Martínez, F.; García-Collado, C.; Calleja-Hernández, M.; Ramírez-Tortosa, M.C.; Jiménez-Morales, A. Effect of genetic polymorphisms on therapeutic response in multiple sclerosis relapsing-remitting patients treated with interferon-beta. Mutat. Res. Rev. Mutat. Res. 2020, 785, 108322. [Google Scholar] [CrossRef] [PubMed]
- Carter, N.J.; Keating, G.M. Glatiramer acetate: A review of its use in relapsing-remitting multiple sclerosis and in delaying the onset of clinically definite multiple sclerosis. Drugs 2010, 70, 1545–1577. [Google Scholar] [CrossRef]
- McGraw, C.A.; Lublin, F.D. Interferon beta and glatiramer acetate therapy. Neurotherapeutics 2013, 10, 2–18. [Google Scholar] [CrossRef]
- Aharoni, R. Immunomodulation neuroprotection and remyelination—The fundamental therapeutic effects of glatiramer acetate: A critical review. J. Autoimmun. 2014, 54, 81–92. [Google Scholar] [CrossRef]
- From, R.; Eilam, R.; Bar-Lev, D.D.; Levin-Zaidman, S.; Tsoory, M.; LoPresti, P.; Sela, M.; Arnon, R.; Aharoni, R. Oligodendrogenesis and myelinogenesis during postnatal development effect of glatiramer acetate. Glia 2014, 62, 649–665. [Google Scholar] [CrossRef] [PubMed]
- Hocevar, K.; Ristic, S.; Peterlin, B. Pharmacogenomics of Multiple Sclerosis: A Systematic Review. Front. Neurol. 2019, 10, 134. [Google Scholar] [CrossRef] [PubMed]
- Mahurkar, S.; Suppiah, V.; O’Doherty, C. Pharmacogenomics of interferon beta and glatiramer acetate response: A review of the literature. Autoimmun. Rev. 2014, 13, 178–186. [Google Scholar] [CrossRef] [PubMed]
- Comabella, M.; Vandenbroeck, K. Pharmacogenomics and multiple sclerosis: Moving toward individualized medicine. Curr. Neurol. Neurosci. Rep. 2011, 11, 484–491. [Google Scholar] [CrossRef]
- Yandava, C.N.; Kennedy, B.P.; Pillari, A.; Duncan, A.M.; Drazen, J.M. Cytogenetic and radiation hybrid mapping of human arachidonate 5-lipoxygenase-activating protein (ALOX5AP) to chromosome 13q12. Genomics 1999, 56, 131–133. [Google Scholar] [CrossRef]
- Safizadeh, B.; Hoshyar, R.; Mehrpour, M.; Eftekhar, M.; Salimi, V.; Yazdani, S.; Bijari, B.; Khodakhah, F.; Tavakoli-Yaraki, M. The role of expression and activity of 15-Lipoxygenase isoforms and related cytokines in patients with Multiple Sclerosis and healthy controls. J. Neuroimmunol. 2018, 325, 32–42. [Google Scholar] [CrossRef] [PubMed]
- Achiron, A.; Feldman, A.; Gurevich, M. Molecular profiling of glatiramer acetate early treatment effects in multiple sclerosis. Dis. Markers 2009, 27, 63–73. [Google Scholar] [CrossRef]
- Ross, C.J.; Towfic, F.; Shankar, J.; Laifenfeld, D.; Thoma, M.; Davis, M.; Weiner, B.; Kusko, R.; Zeskind, B.; Knappertz, V.; et al. A pharmacogenetic signature of high response to Copaxone in late-phase clinical-trial cohorts of multiple sclerosis. Genome Med. 2017, 9, 50. [Google Scholar] [CrossRef]
- Reeves, R.H.; Patch, D.; Sharpe, A.H.; Borriello, F.; Freeman, G.J.; Edelhoff, S.; Disteche, C. The costimulatory genes Cd80 and Cd86 are linked on mouse chromosome 16 and human chromosome 3. Mamm Genome 1997, 8, 581–582. [Google Scholar] [CrossRef]
- Freeman, G.J.; Borriello, F.; Hodes, R.J.; Reiser, H.; Gribben, J.G.; Ng, J.W.; Kim, J.; Goldberg, J.M.; Hathcock, K.; Laszlo, G. Murine B7-2, an alternative CTLA4 counter-receptor that costimulates T cell proliferation and interleukin 2 production. J. Exp. Med. 1993, 178, 2185–2192. [Google Scholar] [CrossRef] [PubMed]
- Fraussen, J.; Claes, N.; Van Wijmeersch, B.; van Horssen, J.; Stinissen, P.; Hupperts, R.; Somers, V. B cells of multiple sclerosis patients induce autoreactive proinflammatory T cell responses. Clin. Immunol. 2016, 173, 124–132. [Google Scholar] [CrossRef] [PubMed]
- Hussien, Y.; Sanna, A.; Soderstrom, M.; Link, H.; Huang, Y.M. Glatiramer acetate and IFN-beta act on dendritic cells in multiple sclerosis. J. Neuroimmunol. 2001, 121, 102–110. [Google Scholar] [CrossRef]
- Yi, C.H.; Terrett, J.A.; Li, Q.Y.; Ellington, K.; Packham, E.A.; Armstrong-Buisseret, L.; McClure, P.; Slingsby, T.; Brook, J.D. Identification, mapping, and phylogenomic analysis of four new human members of the T-box gene family: EOMES, TBX6, TBX18, and TBX19. Genomics 1999, 55, 10–20. [Google Scholar] [CrossRef] [PubMed]
- Parnell, G.P.; Gatt, P.N.; Krupa, M.; Nickles, D.; McKay, F.C.; Schibeci, S.D.; Batten, M.; Baranzini, S.; Henderson, A.; Barnett, M.; et al. The autoimmune disease-associated transcription factors EOMES and TBX21 are dysregulated in multiple sclerosis and define a molecular subtype of disease. Clin. Immunol. 2014, 151, 16–24. [Google Scholar] [CrossRef] [PubMed]
- Raveney, B.J.E.; Sato, W.; Takewaki, D.; Zhang, C.; Kanazawa, T.; Lin, Y.; Okamoto, T.; Araki, M.; Kimura, Y.; Sato, N.; et al. Involvement of cytotoxic Eomes-expressing CD4(+) T cells in secondary progressive multiple sclerosis. Proc. Natl. Acad. Sci. USA 2021, 118. [Google Scholar] [CrossRef]
- Kulakova, O.; Bashinskaya, V.; Kiselev, I.; Baulina, N.; Tsareva, E.; Nikolaev, R.; Kozin, M.; Shchur, S.; Favorov, A.; Boyko, A.; et al. Pharmacogenetics of glatiramer acetate therapy for multiple sclerosis: The impact of genome-wide association studies identified disease risk loci. Pharmacogenomics 2017, 18, 1563–1574. [Google Scholar] [CrossRef] [PubMed]
- Dale, M.; Nicklin, M.J. Interleukin-1 receptor cluster: Gene organization of IL1R2, IL1R1, IL1RL2 (IL-1Rrp2), IL1RL1 (T1/ST2), and IL18R1 (IL-1Rrp) on human chromosome 2q. Genomics 1999, 57, 177–179. [Google Scholar] [CrossRef] [PubMed]
- Yagami, A.; Orihara, K.; Morita, H.; Futamura, K.; Hashimoto, N.; Matsumoto, K.; Saito, H.; Matsuda, A. IL-33 mediates inflammatory responses in human lung tissue cells. J. Immunol. 2010, 185, 5743–5750. [Google Scholar] [CrossRef]
- Ahmadi, M.; Fathi, F.; Fouladi, S.; Alsahebfosul, F.; Manian, M.; Eskandari, N. Serum IL-33 Level and IL-33, IL1RL1 Gene Polymorphisms in Asthma and Multiple Sclerosis Patients. Curr. Mol. Med. 2019, 19, 357–363. [Google Scholar] [CrossRef]
- Carpintero, R.; Burger, D. IFNbeta and glatiramer acetate trigger different signaling pathways to regulate the IL-1 system in multiple sclerosis. Commun. Integr. Biol. 2011, 4, 112–114. [Google Scholar] [CrossRef][Green Version]
- Morton, S.M.; Bocaccio, I.; Depetris, D.; Mattei, M.; Dessein, A. Assignment of IL12RB2 to human chromosome 1p31.3-->p31.2 between D1S230 and D1S198. Cytogenet. Genome Res. 1997, 79, 282–283. [Google Scholar] [CrossRef]
- Koch, M.A.; Thomas, K.R.; Perdue, N.R.; Smigiel, K.S.; Srivastava, S.; Campbell, D.J. T-bet(+) Treg cells undergo abortive Th1 cell differentiation due to impaired expression of IL-12 receptor beta2. Immunity 2012, 37, 501–510. [Google Scholar] [CrossRef] [PubMed]
- Jana, M.; Mondal, S.; Jana, A.; Pahan, K. Interleukin-12 (IL-12), but not IL-23, induces the expression of IL-7 in microglia and macrophages: Implications for multiple sclerosis. Immunology 2014, 141, 549–563. [Google Scholar] [CrossRef] [PubMed]
- Xu, W.; Presnell, S.R.; Parrish-Novak, J.; Kindsvogel, W.; Jaspers, S.; Chen, Z.; Dillon, S.R.; Gao, Z.; Gilbert, T.; Madden, K.; et al. A soluble class II cytokine receptor, IL-22RA2, is a naturally occurring IL-22 antagonist. Proc. Natl. Acad. Sci. USA 2001, 98, 9511–9516. [Google Scholar] [CrossRef] [PubMed]
- Lindahl, H.; Guerreiro-Cacais, A.O.; Bedri, S.K.; Linnerbauer, M.; Linden, M.; Abdelmagid, N.; Tandre, K.; Hollins, C.; Irving, L.; Glover, C.; et al. IL-22 Binding Protein Promotes the Disease Process in Multiple Sclerosis. J. Immunol. 2019, 203, 888–898. [Google Scholar] [CrossRef]
- Wood, J.D.; Yuan, J.; Margolis, R.L.; Colomer, V.; Duan, K.; Kushi, J.; Kaminsky, Z.; Kleiderlein, J.J.; Sharp, A.H.; Ross, C.A. Atrophin-1, the DRPLA gene product, interacts with two families of WW domain-containing proteins. Mol. Cell. Neurosci. 1998, 11, 149–160. [Google Scholar] [CrossRef]
- Bierzynska, A.; Soderquest, K.; Dean, P.; Colby, E.; Rollason, R.; Jones, C.; Inward, C.D.; McCarthy, H.J.; Simpson, M.A.; Lord, G.M.; et al. MAGI2 Mutations Cause Congenital Nephrotic Syndrome. J. Am. Soc. Nephrol. 2017, 28, 1614–1621. [Google Scholar] [CrossRef]
- Wu, X.; Hepner, K.; Castelino-Prabhu, S.; Do, D.; Kaye, M.B.; Yuan, X.J.; Wood, J.; Ross, C.; Sawyers, C.L.; Whang, Y.E. Evidence for regulation of the PTEN tumor suppressor by a membrane-localized multi-PDZ domain containing scaffold protein MAGI-2. Proc. Natl. Acad. Sci. USA 2000, 97, 4233–4238. [Google Scholar] [CrossRef] [PubMed]
- Graham, M.; Adams, J.M. Chromosome 8 breakpoint far 3′ of the c-myc oncogene in a Burkitt’s lymphoma 2;8 variant translocation is equivalent to the murine pvt-1 locus. EMBO J. 1986, 5, 2845–2851. [Google Scholar] [CrossRef] [PubMed]
- Eftekharian, M.M.; Ghafouri-Fard, S.; Soudyab, M.; Omrani, M.D.; Rahimi, M.; Sayad, A.; Komaki, A.; Mazdeh, M.; Taheri, M. Expression Analysis of Long Non-coding RNAs in the Blood of Multiple Sclerosis Patients. J. Mol. Neurosci. 2017, 63, 333–341. [Google Scholar] [CrossRef]
- Zeni, P.F.; Mraz, M. LncRNAs in adaptive immunity: Role in physiological and pathological conditions. RNA Biol. 2021, 18, 619–632. [Google Scholar] [CrossRef]
- Timasheva, Y.R.; Nasibullin, T.R.; Tuktarova, I.A.; Erdman, V.V.; Galiullin, T.R.; Zaplakhova, O.V.; Bakhtiiarova, K.Z.; Mustafina, O.E. The analysis of association between multiple sclerosis and genetic markers identified in genome-wide association studies. Zhurnal Nevrol Psikhiatr Im S S Korsakova 2020, 120, 54–60. [Google Scholar] [CrossRef] [PubMed]
- Bonnefont, J.; Nikolaev, S.I.; Perrier, A.L.; Guo, S.; Cartier, L.; Sorce, S.; Laforge, T.; Aubry, L.; Khaitovich, P.; Peschanski, M.; et al. Evolutionary forces shape the human RFPL1,2,3 genes toward a role in neocortex development. Am. J. Hum. Genet. 2008, 83, 208–218. [Google Scholar] [CrossRef] [PubMed]
- Damseh, N.; Simonin, A.; Jalas, C.; Picoraro, J.A.; Shaag, A.; Cho, M.T.; Yaacov, B.; Neidich, J.; Al-Ashhab, M.; Juusola, J.; et al. Mutations in SLC1A4, encoding the brain serine transporter, are associated with developmental delay, microcephaly and hypomyelination. J. Med. Genet. 2015, 52, 541–547. [Google Scholar] [CrossRef]
- Heimer, G.; Marek-Yagel, D.; Eyal, E.; Barel, O.; Oz Levi, D.; Hoffmann, C.; Ruzzo, E.K.; Ganelin-Cohen, E.; Lancet, D.; Pras, E.; et al. SLC1A4 mutations cause a novel disorder of intellectual disability, progressive microcephaly, spasticity and thin corpus callosum. Clin. Genet. 2015, 88, 327–335. [Google Scholar] [CrossRef]
- Beall, S.S.; Biddison, W.E.; McFarlin, D.E.; McFarland, H.F.; Hood, L.E. Susceptibility for multiple sclerosis is determined, in part, by inheritance of a 175-kb region of the TcR V beta chain locus and HLA class II genes. J. Neuroimmunol. 1993, 45, 53–60. [Google Scholar] [CrossRef]
- Hockertz, M.K.; Paty, D.W.; Beall, S.S. Susceptibility to relapsing-progressive multiple sclerosis is associated with inheritance of genes linked to the variable region of the TcR beta locus: Use of affected family-based controls. Am. J. Hum. Genet. 1998, 62, 373–385. [Google Scholar] [CrossRef][Green Version]
- Deussing, J.; Roth, W.; Rommerskirch, W.; Wiederanders, B.; von Figura, K.; Peters, C. The genes of the lysosomal cysteine proteinases cathepsin B, H, L, and S map to different mouse chromosomes. Mamm. Genome 1997, 8, 241–245. [Google Scholar] [CrossRef] [PubMed]
- Foti Cuzzola, V.; Palella, E.; Celi, D.; Barresi, M.; Giacoppo, S.; Bramanti, P.; Marino, S. Pharmacogenomic update on multiple sclerosis: A focus on actual and new therapeutic strategies. Pharm. J 2012, 12, 453–461. [Google Scholar] [CrossRef] [PubMed][Green Version]
- Liu, R.; Paxton, W.A.; Choe, S.; Ceradini, D.; Martin, S.R.; Horuk, R.; MacDonald, M.E.; Stuhlmann, H.; Koup, R.A.; Landau, N.R. Homozygous defect in HIV-1 coreceptor accounts for resistance of some multiply-exposed individuals to HIV-1 infection. Cell 1996, 86, 367–377. [Google Scholar] [CrossRef]
- Rottman, J.B.; Ganley, K.P.; Williams, K.; Wu, L.; Mackay, C.R.; Ringler, D.J. Cellular localization of the chemokine receptor CCR5. Correlation to cellular targets of HIV-1 infection. Am. J. Pathol. 1997, 151, 1341–1351. [Google Scholar]
- Lynch, E.A.; Heijens, C.A.W.; Horst, N.F.; Center, D.M.; Cruikshank, W.W. Cutting edge: IL-16/CD4 preferentially induces Th1 cell migration: Requirement of CCR5. J. Immunol. 2003, 171, 4965–4968. [Google Scholar] [CrossRef] [PubMed]
- Nakajima, H.; Fukuda, K.; Doi, Y.; Sugino, M.; Kimura, F.; Hanafusa, T.; Ikemoto, T.; Shimizu, A. Expression of TH1/TH2-related chemokine receptors on peripheral T cells and correlation with clinical disease activity in patients with multiple sclerosis. Eur. Neurol. 2004, 52, 162–168. [Google Scholar] [CrossRef] [PubMed]
- Hoglund, R.A.; Hestvik, A.L.; Holmoy, T.; Maghazachi, A.A. Expression and functional activity of chemokine receptors in glatiramer acetate-specific T cells isolated from multiple sclerosis patient receiving the drug glatiramer acetate. Hum. Immunol. 2011, 72, 124–134. [Google Scholar] [CrossRef] [PubMed]
- Tsareva, E.Y.; Kulakova, O.G.; Boyko, A.N.; Shchur, S.G.; Lvovs, D.; Favorov, A.V.; Gusev, E.I.; Vandenbroeck, K.; Favorova, O.O. Allelic combinations of immune-response genes associated with glatiramer acetate treatment response in Russian multiple sclerosis patients. Pharmacogenomics 2012, 13, 43–53. [Google Scholar] [CrossRef] [PubMed]
- Hakonarson, H.; Grant, S.F.; Bradfield, J.P.; Marchand, L.; Kim, C.E.; Glessner, J.T.; Grabs, R.; Casalunovo, T.; Taback, S.P.; Frackelton, E.C.; et al. A genome-wide association study identifies KIAA0350 as a type 1 diabetes gene. Nature 2007, 448, 591–594. [Google Scholar] [CrossRef]
- Geijtenbeek, T.B.; Gringhuis, S.I. Signalling through C-type lectin receptors: Shaping immune responses. Nat. Rev. Immunol. 2009, 9, 465–479. [Google Scholar] [CrossRef]
- Rijvers, L.; Melief, M.J.; van Langelaar, J.; van der Vuurst de Vries, R.M.; Wierenga-Wolf, A.F.; Koetzier, S.C.; Priatel, J.J.; Jorritsma, T.; van Ham, S.M.; Hintzen, R.Q.; et al. The Role of Autoimmunity-Related Gene CLEC16A in the B Cell Receptor-Mediated HLA Class II Pathway. J. Immunol. 2020, 205, 945–956. [Google Scholar] [CrossRef]
- Zuvich, R.L.; Bush, W.S.; McCauley, J.L.; Beecham, A.H.; De Jager, P.L.; the International Multiple Sclerosis Genetics Consortium; Ivinson, A.J.; Compston, A.; Hafler, D.A.; Hauser, S.L.; et al. Interrogating the complex role of chromosome 16p13.13 in multiple sclerosis susceptibility: Independent genetic signals in the CIITA-CLEC16A-SOCS1 gene complex. Hum. Mol. Genet. 2011, 20, 3517–3524. [Google Scholar] [CrossRef]
- Inazawa, J.; Itoh, N.; Abe, T.; Nagata, S. Assignment of the human Fas antigen gene (Fas) to 10q24.1. Genomics 1992, 14, 821–822. [Google Scholar] [CrossRef]
- Haas, J.; Fritzsching, B.; Trubswetter, P.; Korporal, M.; Milkova, L.; Fritz, B.; Vobis, D.; Krammer, P.H.; Suri-Payer, E.; Wildemann, B. Prevalence of newly generated naive regulatory T cells (Treg) is critical for Treg suppressive function and determines Treg dysfunction in multiple sclerosis. J. Immunol. 2007, 179, 1322–1330. [Google Scholar] [CrossRef] [PubMed]
- Schwarz, A.; Schumacher, M.; Pfaff, D.; Schumacher, K.; Jarius, S.; Balint, B.; Wiendl, H.; Haas, J.; Wildemann, B. Fine-tuning of regulatory T cell function: The role of calcium signals and naive regulatory T cells for regulatory T cell deficiency in multiple sclerosis. J. Immunol. 2013, 190, 4965–4970. [Google Scholar] [CrossRef] [PubMed]
- Häusler, D.; Hajiyeva, Z.; Traub, J.W.; Zamvil, S.S.; Lalive, P.H.; Bruck, W.; Weber, M.S. Glatiramer acetate immune modulates B-cell antigen presentation in treatment of MS. Neurol.-Neuroimmunol. Neuroinflamm. 2020, 7, e698. [Google Scholar] [CrossRef] [PubMed]
- Gross, E.A.; Callow, M.G.; Waldbaum, L.; Thomas, S.; Ruggieri, R. MRK, a mixed lineage kinase-related molecule that plays a role in gamma-radiation-induced cell cycle arrest. J. Biol. Chem. 2002, 277, 13873–13882. [Google Scholar] [CrossRef]
- Liu, T.C.; Huang, C.J.; Chu, Y.C.; Wei, C.C.; Chou, C.C.; Chou, M.Y.; Chou, C.K.; Yang, J.J. Cloning and expression of ZAK, a mixed lineage kinase-like protein containing a leucine-zipper and a sterile-alpha motif. Biochem. Biophys. Res. Commun. 2000, 274, 811–816. [Google Scholar] [CrossRef]
- Hasson, T.; Kolitz, S.; Towfic, F.; Laifenfeld, D.; Bakshi, S.; Beriozkin, O.; Shacham-Abramson, M.; Timan, B.; Fowler, K.D.; Birnberg, T.; et al. Functional effects of the antigen glatiramer acetate are complex and tightly associated with its composition. J. Neuroimmunol. 2016, 290, 84–95. [Google Scholar] [CrossRef]
- Von Salome, J.; Gyllensten, U.; Bergstrom, T.F. Full-length sequence analysis of the HLA-DRB1 locus suggests a recent origin of alleles. Immunogenetics 2007, 59, 261–271. [Google Scholar] [CrossRef]
- Lundberg, A.S.; McDevitt, H.O. Evolution of major histocompatibility complex class II allelic diversity: Direct descent in mice and humans. Proc. Natl. Acad. Sci. USA 1992, 89, 6545–6549. [Google Scholar] [CrossRef]
- Gross, R.; Healy, B.C.; Cepok, S.; Chitnis, T.; Khoury, S.J.; Hemmer, B.; Weiner, H.L.; Hafler, D.A.; De Jager, P.L. Population structure and HLA DRB1 1501 in the response of subjects with multiple sclerosis to first-line treatments. J. Neuroimmunol. 2011, 233, 168–174. [Google Scholar] [CrossRef]
- Saxe, D.F.; Takahashi, N.; Hood, L.; Simon, M.I. Localization of the human myelin basic protein gene (MBP) to region 18q22→qter by in situ hybridization. Cytogenet. Genome Res. 1985, 39, 246–249. [Google Scholar] [CrossRef]
- Boggs, J.M. Myelin basic protein: A multifunctional protein. Cell. Mol. Life Sci. 2006, 63, 1945–1961. [Google Scholar] [CrossRef]
- Glatiramer Acetate. In LiverTox: Clinical and Research Information on Drug-Induced Liver Injury; National Institute of Diabetes and Digestive and Kidney Diseases: Bethesda, MD, USA, 2012.
- McAndrew, P.E.; Frostholm, A.; White, R.A.; Rotter, A.; Burghes, A.H. Identification and characterization of RPTP rho, a novel RPTP mu/kappa-like receptor protein tyrosine phosphatase whose expression is restricted to the central nervous system. Mol. Brain Res. 1998, 56, 9–21. [Google Scholar] [CrossRef]
- Wang, Z.; Shen, D.; Parsons, D.W.; Bardelli, A.; Sager, J.; Szabo, S.; Ptak, J.; Silliman, N.; Peters, B.A.; van der Heijden, M.S.; et al. Mutational analysis of the tyrosine phosphatome in colorectal cancers. Science 2004, 304, 1164–1166. [Google Scholar] [CrossRef] [PubMed]
- Perelman, B.; Dafni, N.; Naiman, T.; Eli, D.; Yaakov, M.; Feng, T.L.; Sinha, S.; Weber, G.; Khodaei, S.; Sancar, A.; et al. Molecular cloning of a novel human gene encoding a 63-kDa protein and its sublocalization within the 11q13 locus. Genomics 1997, 41, 397–405. [Google Scholar] [CrossRef] [PubMed]
- Afzal, S.; Hao, Z.; Itsumi, M.; Abouelkheer, Y.; Brenner, D.; Gao, Y.; Wakeham, A.; Hong, C.; Li, W.Y.; Sylvester, J.; et al. Autophagy-independent functions of UVRAG are essential for peripheral naive T-cell homeostasis. Proc. Natl. Acad. Sci. USA 2015, 112, 1119–1124. [Google Scholar] [CrossRef] [PubMed]



{kind=link}
{kind=link}
| Gene | Year | N | Ethnicity | Polymorphism | Overall Response Rate | PMID | ||
|---|---|---|---|---|---|---|---|---|
| p-Value | OR (95% CI) | Genotype Associated | ||||||
| ALOX5AP | 2017 | 639 | Multinational * | rs10162089 | 0.008 | 1.56 | T | 28569182 |
| 532 | Multinational * | rs10162089 | 0.032 | 1.58 | T | |||
| CD86 | 2007 | 35 | Belgium, Canada, The Netherlands, Italy, and the UK | rs1129055 | 0.022 | 6.28 (1.3–30.3) | C | 17622942 |
| 48 | Belgium, Canada, The Netherlands, Italy, and the UK | rs2001791 | 0.062 | 8.3 (0.9–77.0) | T | |||
| CLEC16A | 2017 | 296 | Russian | rs6498169 | 0.025 | 2.38 (1.08–5.27) | A | 29095108 |
| CTSS | 2007 | 43 | Belgium, Canada, The Netherlands, Italy, and the UK | rs2275235 | 0.014 | 11.59 (1.6–81.9) | G | 17622942 |
| 47 | Belgium, Canada, The Netherlands, Italy, and the UK | rs1415148 | 0.009 | 6.85 (1.6–29.2) | A | |||
| EOMES | 2017 | 296 | Russian | rs2371108 | 0.018 | 2.00 (1.09–3.66) | T | 29095108 |
| FAS | 2007 | 47 | Belgium, Canada, The Netherlands, Italy, and the UK | rs982764 | 0.050 | 2.97 (1.0–8.8) | C | 17622942 |
| IL1RL1 | 2007 | 48 | Belgium, Canada, The Netherlands, Italy, and the UK | rs956730 | 0.025 | 5.81 (1.2–27.1) | A | 17622942 |
| IL12RB2 | 2007 | 34 | United States of America | rs946685 | 0.027 | 0.24 (0.07–0.85) | G | 17622942 |
| IL22RA2 | 2017 | 296 | Russian | rs202573 | 0.008 | 2.08 (1.18–7.41) | GG | 29095108 |
| HLA-DRB1 | 2011 | 332 | United States of America | rs3135388 | 0.015 | 2.7 (1.2–6.0) | AA | 21115201 |
| MAGI2 | 2017 | 639 | Multinational * | rs16886004 | 0.002 | 2.15 | A | 28569182 |
| 532 | Multinational * | rs16886004 | <0.001 | 5.56 | A | |||
| MBP | 2007 | 32 | Belgium, Canada, The Netherlands, Italy, and the UK | rs470929 | 0.040 | 5.3(1.1–25.9) | T | 17622942 |
| 2017 | 639 | Multinational * | rs1789084 | 0.036 | 0.7 | T | 28569182 | |
| PTPRT | 2017 | 639 | Multinational * | rs1117602254 | 0.004 | 0.21 | C | 28569182 |
| 532 | Multinational * | rs1117602254 | 0.016 | 0.28 | C | |||
| PVT1 | 2017 | 296 | Russian | rs2114358 | 0.005 | 2.77 (1.33–5.77) | A | 29095108 |
| RFPL3 | 2017 | 532 | Multinational * | rs1789084 | 0.010 | 0.57 | C | 28569182 |
| 532 | Multinational * | rs73166319 | <0.001 | 0.12 | C | |||
| SLC1A4 | 2017 | 639 | Multinational * | rs759458 | <0.001 | 3.31 | G | 28569182 |
| 532 | Multinational * | rs759458 | 0.049 | 1.86 | G | |||
| TRBC | 2007 | 31 | Belgium, Canada, The Netherlands, Italy, and the UK | rs71878 | 0.015 | 6.8 (1.45–31.9) | C | 17622942 |
| UVRAG | 2017 | 639 | Multinational * | rs80191572 | 0.002 | 0.20 | A | 28569182 |
| 532 | Multinational * | rs80191572 | <0.001 | 0.12 | A | |||
| ZAK | 2017 | 639 | Multinational * | rs139890339 | <0.001 | 0.05 | C | 28569182 |
| 532 | Multinational * | rs139890339 | 0.011 | 0.14 | C | |||
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Zarzuelo-Romero, M.J.; Pérez-Ramírez, C.; Cura, Y.; Carrasco-Campos, M.I.; Marangoni-Iglecias, L.M.; Ramírez-Tortosa, M.C.; Jiménez-Morales, A. Influence of Genetic Polymorphisms on Clinical Outcomes of Glatiramer Acetate in Multiple Sclerosis Patients. J. Pers. Med. 2021, 11, 1032. https://doi.org/10.3390/jpm11101032
Zarzuelo-Romero MJ, Pérez-Ramírez C, Cura Y, Carrasco-Campos MI, Marangoni-Iglecias LM, Ramírez-Tortosa MC, Jiménez-Morales A. Influence of Genetic Polymorphisms on Clinical Outcomes of Glatiramer Acetate in Multiple Sclerosis Patients. Journal of Personalized Medicine. 2021; 11(10):1032. https://doi.org/10.3390/jpm11101032
Chicago/Turabian StyleZarzuelo-Romero, María José, Cristina Pérez-Ramírez, Yasmín Cura, María Isabel Carrasco-Campos, Luciana María Marangoni-Iglecias, María Carmen Ramírez-Tortosa, and Alberto Jiménez-Morales. 2021. "Influence of Genetic Polymorphisms on Clinical Outcomes of Glatiramer Acetate in Multiple Sclerosis Patients" Journal of Personalized Medicine 11, no. 10: 1032. https://doi.org/10.3390/jpm11101032
APA StyleZarzuelo-Romero, M. J., Pérez-Ramírez, C., Cura, Y., Carrasco-Campos, M. I., Marangoni-Iglecias, L. M., Ramírez-Tortosa, M. C., & Jiménez-Morales, A. (2021). Influence of Genetic Polymorphisms on Clinical Outcomes of Glatiramer Acetate in Multiple Sclerosis Patients. Journal of Personalized Medicine, 11(10), 1032. https://doi.org/10.3390/jpm11101032