
Vascular Contribution to Cerebral Waste Clearance Affected by Aging or Diabetes

 , ,
, , {kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
1. Introduction
2. Materials and Methods
2.1. Animals and Experimental Procedures
2.2. Imaging Data Analysis
2.3. Statistical Analysis
3. Results
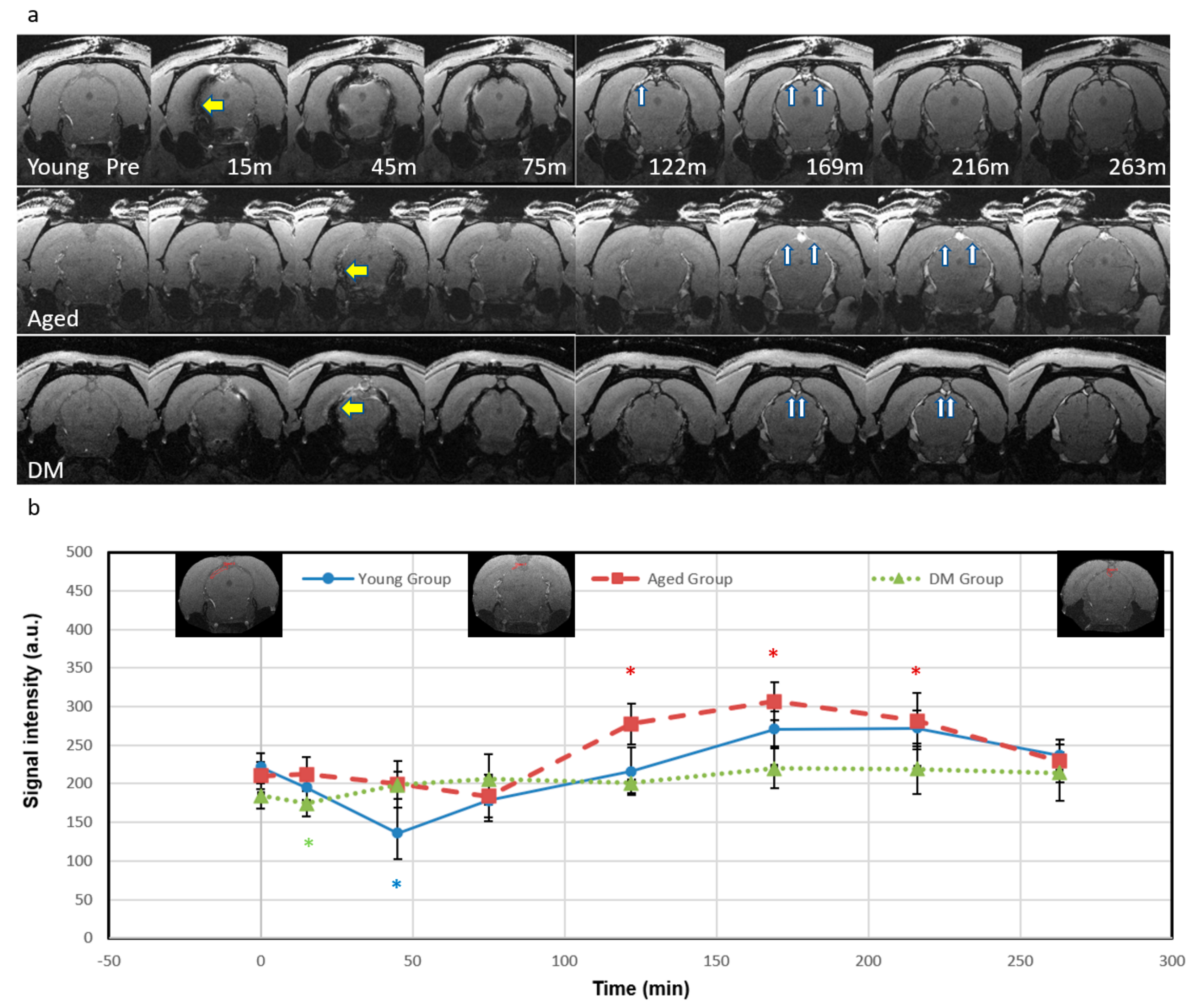
3.1. Visualization of Signal Intensity (SI) Changes After ICM Injection of Ferumoxytol
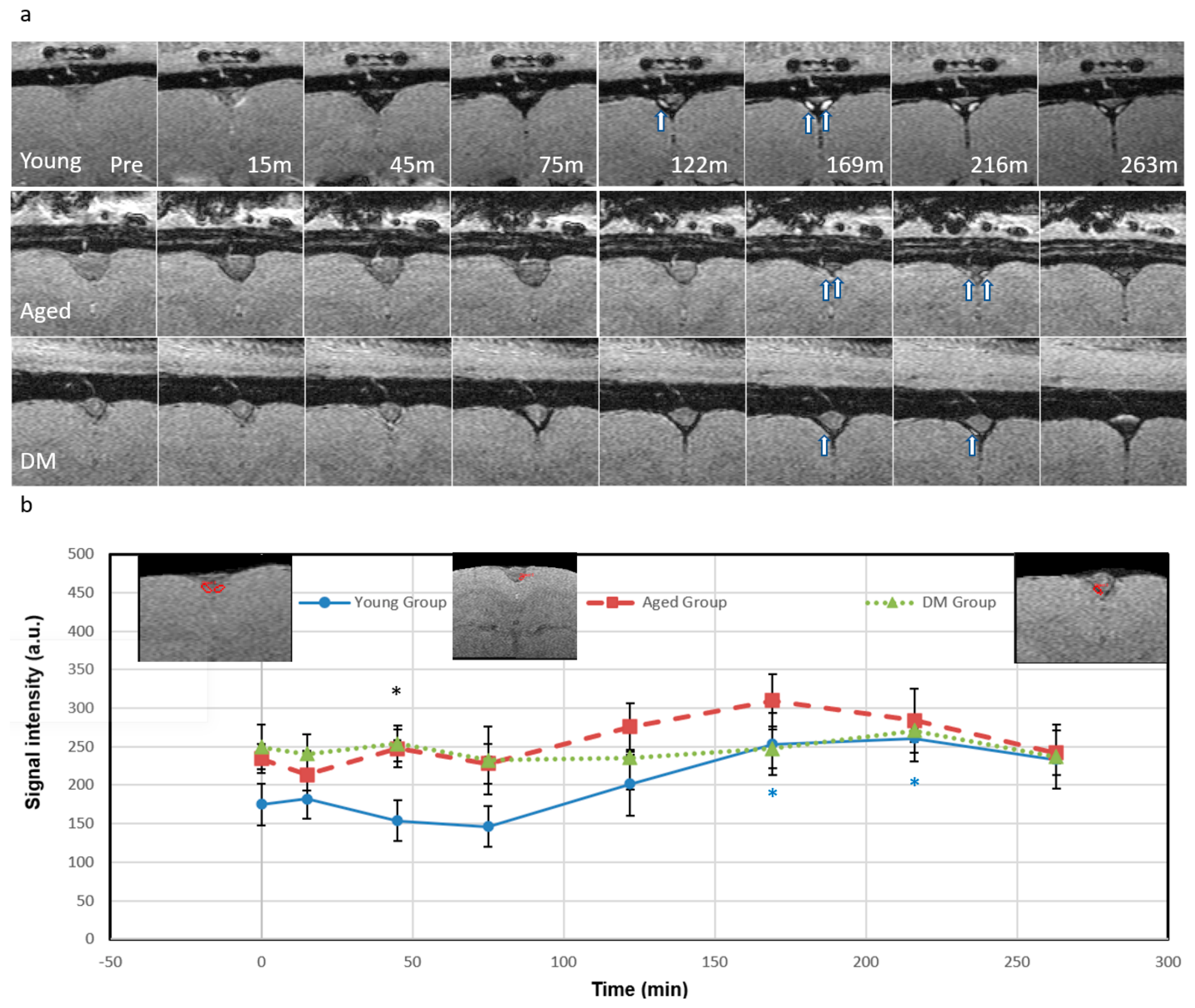
3.2. CSF Tracer Entry into the Parenchymal Veins Measured by QSM
4. Discussion
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
Abbreviations
| azicv | azygos internal cerebral vein |
| BBB | blood–brain barrier |
| CNS | central nervous system |
| CSF | cerebrospinal fluid |
| CWC | cerebral waste clearance |
| ICM | intracisternal magna |
| IV | intravenous |
| MRI | magnetic resonance imaging |
| PVS | peri-venous space |
| QSM | quantitative susceptibility mapping |
| ROI | region of interest |
| SI | signal intensity |
| SPIO | superparamagnetic iron oxide |
| SWI | susceptibility-weighted imaging |
| SPIO-SWI | superparamagnetic iron oxide-enhanced susceptibility-weighted imaging |
References
- Xie, L.; Kang, H.; Xu, Q.; Chen, M.J.; Liao, Y.; Thiyagarajan, M.; O’Donnell, J.; Christensen, D.J.; Nicholson, C.; Iliff, J.J.; et al. Sleep drives metabolite clearance from the adult brain. Science 2013, 342, 373–377. [Google Scholar] [CrossRef] [PubMed]
- Iliff, J.J.; Wang, M.; Zeppenfeld, D.M.; Venkataraman, A.; Plog, B.A.; Liao, Y.; Deane, R.; Nedergaard, M. Cerebral arterial pulsation drives paravascular CSF-interstitial fluid exchange in the murine brain. J. Neurosci. Off. J. Soc. Neurosci. 2013, 33, 18190–18199. [Google Scholar] [CrossRef] [PubMed]
- Iliff, J.J.; Wang, M.; Liao, Y.; Plogg, B.A.; Peng, W.; Gundersen, G.A.; Benveniste, H.; Vates, G.E.; Deane, R.; Goldman, S.A.; et al. A paravascular pathway facilitates CSF flow through the brain parenchyma and the clearance of interstitial solutes, including amyloid beta. Sci. Transl. Med. 2012, 4, 147ra111. [Google Scholar] [CrossRef] [PubMed]
- Rangroo Thrane, V.; Thrane, A.S.; Plog, B.A.; Thiyagarajan, M.; Iliff, J.J.; Deane, R.; Nagelhus, E.A.; Nedergaard, M. Paravascular microcirculation facilitates rapid lipid transport and astrocyte signaling in the brain. Sci. Rep. 2013, 3, 2582. [Google Scholar] [CrossRef]
- Plog, B.A.; Dashnaw, M.L.; Hitomi, E.; Peng, W.; Liao, Y.; Lou, N.; Deane, R.; Nedergaard, M. Biomarkers of traumatic injury are transported from brain to blood via the glymphatic system. J. Neurosci. Off. J. Soc. Neurosci. 2015, 35, 518–526. [Google Scholar] [CrossRef]
- Peng, W.; Achariyar, T.M.; Li, B.; Liao, Y.; Mestre, H.; Hitomi, E.; Regan, S.; Kasper, T.; Peng, S.; Ding, F.; et al. Suppression of glymphatic fluid transport in a mouse model of Alzheimer’s disease. Neurobiol. Dis. 2016, 93, 215–225. [Google Scholar] [CrossRef]
- Hladky, S.B.; Barrand, M.A. Elimination of substances from the brain parenchyma: Efflux via perivascular pathways and via the blood-brain barrier. Fluids Barriers CNS 2018, 15, 30. [Google Scholar] [CrossRef]
- Benveniste, H.; Liu, X.; Koundal, S.; Sanggaard, S.; Lee, H.; Wardlaw, J. The Glymphatic System and Waste Clearance with Brain Aging: A Review. Gerontology 2018, 65, 106–119. [Google Scholar] [CrossRef]
- Louveau, A.; Plog, B.A.; Antila, S.; Alitalo, K.; Nedergaard, M.; Kipnis, J. Understanding the functions and relationships of the glymphatic system and meningeal lymphatics. J. Clin. Investig. 2017, 127, 3210–3219. [Google Scholar] [CrossRef]
- Louveau, A.; Smirnov, I.; Keyes, T.J.; Eccles, J.D.; Rouhani, S.J.; Peske, J.D.; Derecki, N.C.; Castle, D.; Mandell, J.W.; Lee, K.S.; et al. Structural and functional features of central nervous system lymphatic vessels. Nature 2015, 523, 337–341. [Google Scholar] [CrossRef]
- Albargothy, N.J.; Johnston, D.A.; MacGregor-Sharp, M.; Weller, R.O.; Verma, A.; Hawkes, C.A.; Carare, R.O. Convective influx/glymphatic system: Tracers injected into the CSF enter and leave the brain along separate periarterial basement membrane pathways. Acta Neuropathol. 2018, 136, 139–152. [Google Scholar] [CrossRef] [PubMed]
- Carare, R.O.; Bernardes-Silva, M.; Newman, T.A.; Page, A.M.; Nicoll, J.A.; Perry, V.H.; Weller, R.O. Solutes, but not cells, drain from the brain parenchyma along basement membranes of capillaries and arteries: Significance for cerebral amyloid angiopathy and neuroimmunology. Neuropathol. Appl. Neurobiol. 2008, 34, 131–144. [Google Scholar] [CrossRef] [PubMed]
- Arbel-Ornath, M.; Hudry, E.; Eikermann-Haerter, K.; Hou, S.; Gregory, J.L.; Zhao, L.; Betensky, R.A.; Frosch, M.P.; Greenberg, S.M.; Bacskai, B.J. Interstitial fluid drainage is impaired in ischemic stroke and Alzheimer’s disease mouse models. Acta Neuropathol. 2013, 126, 353–364. [Google Scholar] [CrossRef]
- Fahmy, L.M.; Chen, Y.; Xuan, S.; Haacke, E.M.; Hu, J.; Jiang, Q. All Central Nervous System Neuro- and Vascular-Communication Channels Are Surrounded with Cerebrospinal Fluid. Front. Neurol. 2021, 12, 614636. [Google Scholar] [CrossRef]
- Hu, J.; Shen, Y.; Fahmy, L.M.; Krishnamurthy, S.; Li, J.; Zhang, L.; Chen, Y.; Haacke, E.M.; Jiang, Q. The role of the parenchymal vascular system in cerebrospinal fluid tracer clearance. Eur. Radiol. 2023, 33, 656–665. [Google Scholar] [CrossRef]
- Krishnamurthy, S.; Li, J.; Shen, Y.; Duncan, T.M.; Jenrow, K.A.; Haacke, E.M. Normal macromolecular clearance out of the ventricles is delayed in hydrocephalus. Brain Res. 2018, 1678, 337–355. [Google Scholar] [CrossRef]
- Weed, L.H. Studies on Cerebro-Spinal Fluid. No. III: The pathways of escape from the Subarachnoid Spaces with particular reference to the Arachnoid Villi. J. Med. Res. 1914, 31, 51–91. [Google Scholar]
- Brinker, T.; Stopa, E.; Morrison, J.; Klinge, P. A new look at cerebrospinal fluid circulation. Fluids Barriers CNS 2014, 11, 10. [Google Scholar] [CrossRef]
- Shah, T.; Leurgans, S.E.; Mehta, R.I.; Yang, J.; Galloway, C.A.; de Mesy Bentley, K.L.; Schneider, J.A.; Mehta, R.I. Arachnoid granulations are lymphatic conduits that communicate with bone marrow and dura-arachnoid stroma. J. Exp. Med. 2023, 220, e20220618. [Google Scholar] [CrossRef]
- McComb, J.G. Recent research into the nature of cerebrospinal fluid formation and absorption. J. Neurosurg. 1983, 59, 369–383. [Google Scholar] [CrossRef]
- Courtice, F.C.; Simmonds, W.J. The removal of protein from the subarachnoid space. Aust. J. Exp. Biol. Med. Sci. 1951, 29, 255–263. [Google Scholar] [CrossRef] [PubMed]
- Eide, P.K.; Mariussen, E.; Uggerud, H.; Pripp, A.H.; Lashkarivand, A.; Hassel, B.; Christensen, H.; Hovd, M.H.; Ringstad, G. Clinical application of intrathecal gadobutrol for assessment of cerebrospinal fluid tracer clearance to blood. JCI Insight 2021, 6, e147063. [Google Scholar] [CrossRef] [PubMed]
- Liu, S.; Buch, S.; Chen, Y.; Choi, H.S.; Dai, Y.; Habib, C.; Hu, J.; Jung, J.Y.; Luo, Y.; Utriainen, D.; et al. Susceptibility-weighted imaging: Current status and future directions. NMR Biomed. 2017, 30, e3552. [Google Scholar] [CrossRef] [PubMed]
- Jiang, Q.; Zhang, L.; Ding, G.; Davoodi-Bojd, E.; Li, Q.; Li, L.; Sadry, N.; Nedergaard, M.; Chopp, M.; Zhang, Z. Impairment of the glymphatic system after diabetes. J. Cereb. Blood Flow Metab. Off. J. Int. Soc. Cereb. Blood Flow Metab. 2017, 37, 1326–1337. [Google Scholar] [CrossRef]
- Masiello, P.; Broca, C.; Gross, R.; Roye, M.; Manteghetti, M.; Hillaire-Buys, D.; Novelli, M.; Ribes, G. Experimental NIDDM: Development of a new model in adult rats administered streptozotocin and nicotinamide. Diabetes 1998, 47, 224–229. [Google Scholar] [CrossRef]
- Su, H.C.; Hung, L.M.; Chen, J.K. Resveratrol, a red wine antioxidant, possesses an insulin-like effect in streptozotocin-induced diabetic rats. Am. J. Physiol. Endocrinol. Metab. 2006, 290, E1339–E1346. [Google Scholar] [CrossRef]
- Ding, G.; Chopp, M.; Li, L.; Zhang, L.; Davoodi-Bojd, E.; Li, Q.; Zhang, Z.; Jiang, Q. MRI investigation of glymphatic responses to Gd-DTPA infusion rates. J. Neurosci. Res. 2018, 96, 1876–1886. [Google Scholar] [CrossRef]
- Haacke, E.M. Magnetic Resonance Imaging: Principles and Sequence Design; John Wiley & Son: New York, NY, USA, 1999. [Google Scholar]
- Gossuin, Y.; Gillis, P.; Hocq, A.; Vuong, Q.L.; Roch, A. Magnetic resonance relaxation properties of superparamagnetic particles. Wiley Interdiscip Rev. Nanomed. Nanobiotechnol. 2009, 1, 299–310. [Google Scholar] [CrossRef]
- Wei, H.; Bruns, O.T.; Kaul, M.G.; Hansen, E.C.; Barch, M.; Wisniowska, A.; Chen, O.; Chen, Y.; Li, N.; Okada, S.; et al. Exceedingly small iron oxide nanoparticles as positive MRI contrast agents. Proc. Natl. Acad. Sci. USA 2017, 114, 2325–2330. [Google Scholar] [CrossRef]
- Haacke, E.M.; Liu, S.; Buch, S.; Zheng, W.; Wu, D.; Ye, Y. Quantitative susceptibility mapping: Current status and future directions. Magn. Reson. Imaging 2015, 33, 1–25. [Google Scholar] [CrossRef]
- Smith, S.M. Fast robust automated brain extraction. Hum. Brain Mapp. 2002, 17, 143–155. [Google Scholar] [CrossRef] [PubMed]
- Abdul-Rahman, H.S.; Gdeisat, M.A.; Burton, D.R.; Lalor, M.J.; Lilley, F.; Moore, C.J. Fast and robust three-dimensional best path phase unwrapping algorithm. Appl. Opt. 2007, 46, 6623–6635. [Google Scholar] [CrossRef] [PubMed]
- Schweser, F.; Deistung, A.; Lehr, B.W.; Reichenbach, J.R. Quantitative imaging of intrinsic magnetic tissue properties using MRI signal phase: An approach to in vivo brain iron metabolism? NeuroImage 2011, 54, 2789–2807. [Google Scholar] [CrossRef] [PubMed]
- Tang, J.; Liu, S.; Neelavalli, J.; Cheng, Y.C.; Buch, S.; Haacke, E.M. Improving susceptibility mapping using a threshold-based K-space/image domain iterative reconstruction approach. Magn. Reson. Med. 2013, 69, 1396–1407. [Google Scholar] [CrossRef]
- Arrighin, G.P.; Maestro, M.; Moccia, R. Magnetic properties of polyatomic molecules. I. Magnetic susceptibility of H2O, NH3, CH4, H2O2. J. Chem. Phys. 1968, 49, 882. [Google Scholar] [CrossRef]
- Zhou, Y.; Cai, J.; Zhang, W.; Gong, X.; Yan, S.; Zhang, K.; Luo, Z.; Sun, J.; Jiang, Q.; Lou, M. Impairment of the Glymphatic Pathway and Putative Meningeal Lymphatic Vessels in the Aging Human. Ann. Neurol. 2020, 87, 357–369. [Google Scholar] [CrossRef]
- Jani, R.H.; Sekula, R.F., Jr. Magnetic Resonance Imaging of Human Dural Meningeal Lymphatics. Neurosurgery 2018, 83, E10–E12. [Google Scholar] [CrossRef]
- Ha, S.K.; Nair, G.; Absinta, M.; Luciano, N.J.; Reich, D.S. Magnetic Resonance Imaging and Histopathological Visualization of Human Dural Lymphatic Vessels. Bio Protoc. 2018, 8, e2819. [Google Scholar] [CrossRef]
- Absinta, M.; Ha, S.K.; Nair, G.; Sati, P.; Luciano, N.J.; Palisoc, M.; Louveau, A.; Zaghloul, K.A.; Pittaluga, S.; Kipnis, J.; et al. Human and nonhuman primate meninges harbor lymphatic vessels that can be visualized noninvasively by MRI. eLife 2017, 6, e29738. [Google Scholar] [CrossRef]
- Li, L.; Ding, G.; Zhang, L.; Davoodi-Bojd, E.; Chopp, M.; Li, Q.; Zhang, Z.G.; Jiang, Q. Aging-Related Alterations of Glymphatic Transport in Rat: In vivo Magnetic Resonance Imaging and Kinetic Study. Front. Aging Neurosci. 2022, 14, 841798. [Google Scholar] [CrossRef]
- Kress, B.T.; Iliff, J.J.; Xia, M.; Wang, M.; Wei, H.S.; Zeppenfeld, D.; Xie, L.; Kang, H.; Xu, Q.; Liew, J.A.; et al. Impairment of paravascular clearance pathways in the aging brain. Ann. Neurol. 2014, 76, 845–861. [Google Scholar] [CrossRef] [PubMed]
- Taoka, T.; Masutani, Y.; Kawai, H.; Nakane, T.; Matsuoka, K.; Yasuno, F.; Kishimoto, T.; Naganawa, S. Evaluation of glymphatic system activity with the diffusion MR technique: Diffusion tensor image analysis along the perivascular space (DTI-ALPS) in Alzheimer’s disease cases. Jpn. J. Radiol. 2017, 35, 172–178. [Google Scholar] [CrossRef] [PubMed]
- Taoka, T.; Ito, R.; Nakamichi, R.; Nakane, T.; Kawai, H.; Naganawa, S. Diffusion Tensor Image Analysis ALong the Perivascular Space (DTI-ALPS): Revisiting the Meaning and Significance of the Method. Magn. Reson. Med. Sci. MRMS Off. J. Jpn. Soc. Magn. Reson. Med. 2024, 23, 268–290. [Google Scholar] [CrossRef] [PubMed]
- Ringstad, G. Glymphatic imaging: A critical look at the DTI-ALPS index. Neuroradiology 2024, 66, 157–160. [Google Scholar] [CrossRef]
- Yu, S.; Jiang, H.; Yu, L.; Liu, T.; Yang, C.; Cao, J.; Song, Q.; Miao, Y.; Wang, W. DTI-ALPS index decreased in patients with Type 2 Diabetes Mellitus. Front. Neurosci. 2024, 18, 1383780. [Google Scholar] [CrossRef]
- Wheaton, A.J.; Miyazaki, M. Non-contrast enhanced MR angiography: Physical principles. J. Magn. Reson. Imaging 2012, 36, 286–304. [Google Scholar] [CrossRef]
- Lowe, K.L.; Finney, B.A.; Deppermann, C.; Hagerling, R.; Gazit, S.L.; Frampton, J.; Buckley, C.; Camerer, E.; Nieswandt, B.; Kiefer, F.; et al. Podoplanin and CLEC-2 drive cerebrovascular patterning and integrity during development. Blood 2015, 125, 3769–3777. [Google Scholar] [CrossRef]
- Aspelund, A.; Tammela, T.; Antila, S.; Nurmi, H.; Leppanen, V.M.; Zarkada, G.; Stanczuk, L.; Francois, M.; Makinen, T.; Saharinen, P.; et al. The Schlemm’s canal is a VEGF-C/VEGFR-3-responsive lymphatic-like vessel. J. Clin. Investig. 2014, 124, 3975–3986. [Google Scholar] [CrossRef]
- Hsu, M.; Laaker, C.; Sandor, M.; Fabry, Z. Neuroinflammation-Driven Lymphangiogenesis in CNS Diseases. Front. Cell Neurosci. 2021, 15, 683676. [Google Scholar] [CrossRef]
- Ahn, J.H.; Cho, H.; Kim, J.H.; Kim, S.H.; Ham, J.S.; Park, I.; Suh, S.H.; Hong, S.P.; Song, J.H.; Hong, Y.K.; et al. Meningeal lymphatic vessels at the skull base drain cerebrospinal fluid. Nature 2019, 572, 62–66. [Google Scholar] [CrossRef]
- Mäkinen, T. Lymphatic vessels at the base of the mouse brain provide direct drainage to the periphery. Nature 2019, 572, 34–35. [Google Scholar] [CrossRef] [PubMed]
- Chiu, C.; Miller, M.C.; Caralopoulos, I.N.; Worden, M.S.; Brinker, T.; Gordon, Z.N.; Johanson, C.E.; Silverberg, G.D. Temporal course of cerebrospinal fluid dynamics and amyloid accumulation in the aging rat brain from three to thirty months. Fluids Barriers CNS 2012, 9, 3. [Google Scholar] [CrossRef] [PubMed]
- Serot, J.M.; Bene, M.C.; Faure, G.C. Choroid plexus, aging of the brain, and Alzheimer’s disease. Front. Biosci. A J. Virtual Libr. 2003, 8, s515–s521. [Google Scholar]
- Preston, J.E. Ageing choroid plexus-cerebrospinal fluid system. Microsc. Res. Tech. 2001, 52, 31–37. [Google Scholar] [CrossRef]
- Czosnyka, M.; Pickard, J.D. Monitoring and interpretation of intracranial pressure. J. Neurol. Neurosurg. Psychiatry 2004, 75, 813–821. [Google Scholar] [CrossRef]
- May, C.; Kaye, J.A.; Atack, J.R.; Schapiro, M.B.; Friedland, R.P.; Rapoport, S.I. Cerebrospinal fluid production is reduced in healthy aging. Neurology 1990, 40, 500–503. [Google Scholar] [CrossRef]
- Ma, Q.; Ineichen, B.V.; Detmar, M.; Proulx, S.T. Outflow of cerebrospinal fluid is predominantly through lymphatic vessels and is reduced in aged mice. Nat. Commun. 2017, 8, 1434. [Google Scholar] [CrossRef]
- Nagra, G.; Johnston, M.G. Impact of ageing on lymphatic cerebrospinal fluid absorption in the rat. Neuropathol. Appl. Neurobiol. 2007, 33, 684–691. [Google Scholar] [CrossRef]
- Park, M.; Kim, J.W.; Ahn, S.J.; Cha, Y.J.; Suh, S.H. Aging Is Positively Associated with Peri-Sinus Lymphatic Space Volume: Assessment Using 3T Black-Blood MRI. J. Clin. Med. 2020, 9, 3353. [Google Scholar] [CrossRef]
- Jessen, N.A.; Munk, A.S.; Lundgaard, I.; Nedergaard, M. The Glymphatic System: A Beginner’s Guide. Neurochem. Res. 2015, 40, 2583–2599. [Google Scholar] [CrossRef]
- Kyrtsos, C.R.; Baras, J.S. Modeling the Role of the Glymphatic Pathway and Cerebral Blood Vessel Properties in Alzheimer’s Disease Pathogenesis. PLoS ONE 2015, 10, e0139574. [Google Scholar] [CrossRef] [PubMed]
- Zeppenfeld, D.M.; Simon, M.; Haswell, J.D.; D’Abreo, D.; Murchison, C.; Quinn, J.F.; Grafe, M.R.; Woltjer, R.L.; Kaye, J.; Iliff, J.J. Association of Perivascular Localization of Aquaporin-4 With Cognition and Alzheimer Disease in Aging Brains. JAMA Neurol. 2017, 74, 91–99. [Google Scholar] [CrossRef] [PubMed]
- Zong, X.; Lian, C.; Jimenez, J.; Yamashita, K.; Shen, D.; Lin, W. Morphology of perivascular spaces and enclosed blood vessels in young to middle-aged healthy adults at 7T: Dependences on age, brain region, and breathing gas. NeuroImage 2020, 218, 116978. [Google Scholar] [CrossRef] [PubMed]
- Laveskog, A.; Wang, R.; Vetrano, D.L.; Bronge, L.; Wahlund, L.O.; Qiu, C. Associations of Vascular Risk Factors and APOE Genotype with Perivascular Spaces Among Community-Dwelling Older Adults. J. Am. Heart Assoc. 2020, 9, e015229. [Google Scholar] [CrossRef]
- Watanabe, N.; Noda, Y.; Nemoto, T.; Iimura, K.; Shimizu, T.; Hotta, H. Cerebral artery dilation during transient ischemia is impaired by amyloid beta deposition around the cerebral artery in Alzheimer’s disease model mice. J. Physiol. Sci. 2020, 70, 57. [Google Scholar] [CrossRef]
- Murugesan, N.; Demarest, T.G.; Madri, J.A.; Pachter, J.S. Brain regional angiogenic potential at the neurovascular unit during normal aging. Neurobiol. Aging 2012, 33, 1004-e1. [Google Scholar] [CrossRef]
- Toth, P.; Tarantini, S.; Csiszar, A.; Ungvari, Z. Functional vascular contributions to cognitive impairment and dementia: Mechanisms and consequences of cerebral autoregulatory dysfunction, endothelial impairment, and neurovascular uncoupling in aging. Am. J. Physiol. Heart Circ. Physiol. 2017, 312, H1–H20. [Google Scholar] [CrossRef]
- Venkat, P.; Chopp, M.; Chen, J. New insights into coupling and uncoupling of cerebral blood flow and metabolism in the brain. Croat. Med. J. 2016, 57, 223–228. [Google Scholar] [CrossRef]
- Biondetti, E.; Cho, J.; Lee, H. Cerebral oxygen metabolism from MRI susceptibility. NeuroImage 2023, 276, 120189. [Google Scholar] [CrossRef]
- Braz, I.D.; Fisher, J.P. The impact of age on cerebral perfusion, oxygenation and metabolism during exercise in humans. J. Physiol. 2016, 594, 4471–4483. [Google Scholar] [CrossRef]
- Uchida, Y.; Kan, H.; Sakurai, K.; Oishi, K.; Matsukawa, N. Quantitative susceptibility mapping as an imaging biomarker for Alzheimer’s disease: The expectations and limitations. Front. Neurosci. 2022, 16, 938092. [Google Scholar] [CrossRef] [PubMed]
- Wang, Q.; Zennadi, R. Oxidative Stress and Thrombosis during Aging: The Roles of Oxidative Stress in RBCs in Venous Thrombosis. Int. J. Mol. Sci. 2020, 21, 4259. [Google Scholar] [CrossRef] [PubMed]
- Hommer, N.; Kallab, M.; Schlatter, A.; Howorka, K.; Werkmeister, R.M.; Schmidl, D.; Schmetterer, L.; Garhofer, G. Retinal Oxygen Metabolism in Patients with Type 2 Diabetes and Different Stages of Diabetic Retinopathy. Diabetes 2022, 71, 2677–2684. [Google Scholar] [CrossRef]
- Kan, H.; Arai, N.; Takizawa, M.; Kasai, H.; Kunitomo, H.; Hirose, Y.; Shibamoto, Y. Improvement of Signal Inhomogeneity Induced by Radio-frequency Transmit-related Phase Error for Single-step Quantitative Susceptibility Mapping Reconstruction. Magn. Reson. Med. Sci. MRMS Off. J. Jpn. Soc. Magn. Reson. Med. 2019, 18, 276–285. [Google Scholar] [CrossRef]
- Kan, H.; Arai, N.; Kasai, H.; Kunitomo, H.; Hirose, Y.; Shibamoto, Y. Quantitative susceptibility mapping using principles of echo shifting with a train of observations sequence on 1.5T MRI. Magn. Reson. Imaging 2017, 42, 37–42. [Google Scholar] [CrossRef]
- Duchnowski, P.; Śmigielski, W. Usefulness of myocardial damage biomarkers in predicting cardiogenic shock in patients undergoing heart valve surgery. Kardiol. Pol. 2024, 82, 423–426. [Google Scholar] [CrossRef]
- Kritsilis, M.; Vanherle, L.; Rosenholm, M.; In ‘t Zandt, R.; Yao, Y.; Swanberg, K.M.; Weikop, P.; Gottschalk, M.; Shanbhag, N.C.; Luo, J.; et al. Loss of glymphatic homeostasis in heart failure. Brain A J. Neurol. 2025, 148, 985–1000. [Google Scholar] [CrossRef]



Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2025 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Shen, Y.; Zhang, L.; Ding, G.; Boyd, E.; Kaur, J.; Li, Q.; Haacke, E.M.; Hu, J.; Jiang, Q. Vascular Contribution to Cerebral Waste Clearance Affected by Aging or Diabetes. Diagnostics 2025, 15, 1019. https://doi.org/10.3390/diagnostics15081019
Shen Y, Zhang L, Ding G, Boyd E, Kaur J, Li Q, Haacke EM, Hu J, Jiang Q. Vascular Contribution to Cerebral Waste Clearance Affected by Aging or Diabetes. Diagnostics. 2025; 15(8):1019. https://doi.org/10.3390/diagnostics15081019
Chicago/Turabian StyleShen, Yimin, Li Zhang, Guangliang Ding, Edward Boyd, Jasleen Kaur, Qingjiang Li, E. Mark Haacke, Jiani Hu, and Quan Jiang. 2025. "Vascular Contribution to Cerebral Waste Clearance Affected by Aging or Diabetes" Diagnostics 15, no. 8: 1019. https://doi.org/10.3390/diagnostics15081019
APA StyleShen, Y., Zhang, L., Ding, G., Boyd, E., Kaur, J., Li, Q., Haacke, E. M., Hu, J., & Jiang, Q. (2025). Vascular Contribution to Cerebral Waste Clearance Affected by Aging or Diabetes. Diagnostics, 15(8), 1019. https://doi.org/10.3390/diagnostics15081019