
Olfactory Dysfunction as a Clinical Marker of Early Glymphatic Failure in Neurodegenerative Diseases

 , and
, and {kind=link}
{kind=link}
Abstract
1. Introduction
2. Olfactory Dysfunction in Neurodegenerative Diseases
2.1. OD in Preclinical Alzheimer’s Disease
2.2. OD in Prodromal Lewy Body Diseases
3. Glymphatic System and Its Failure in Neurodegenerative Diseases
4. Menopause, Olfactory Dysfunction, and Glymphatic Failure
5. Olfactory Dysfunction as a Clinical Marker of Glymphatic Failure in Neurodegenerative Disease
- (1)
- Olfactory structures show early protein aggregation. Early Aβ, tau and α-synuclein protein aggregates have been observed in the olfactory bulb, olfactory epithelium, piriform cortex, the glomerular layer, anterior olfactory nucleus, and olfactory tubercle [74,75,76]. In LBD, the olfactory bulbs are an initial site of α-synuclein aggregation [34]. Neuropil threads and neurofibrillary tangles of tau protein have been observed in the olfactory bulb and olfactory nerve in all cases of definite AD, as well as in many cases of probable AD, MCI, and even cognitively normal aging [77,78]. Moreover, in a healthy elderly population, primary age-related tauopathy typically concentrates in the medial temporal lobe and olfactory regions, including the olfactory bulb, transentorhinal region, and entorhinal cortex [79,80].
- (2)
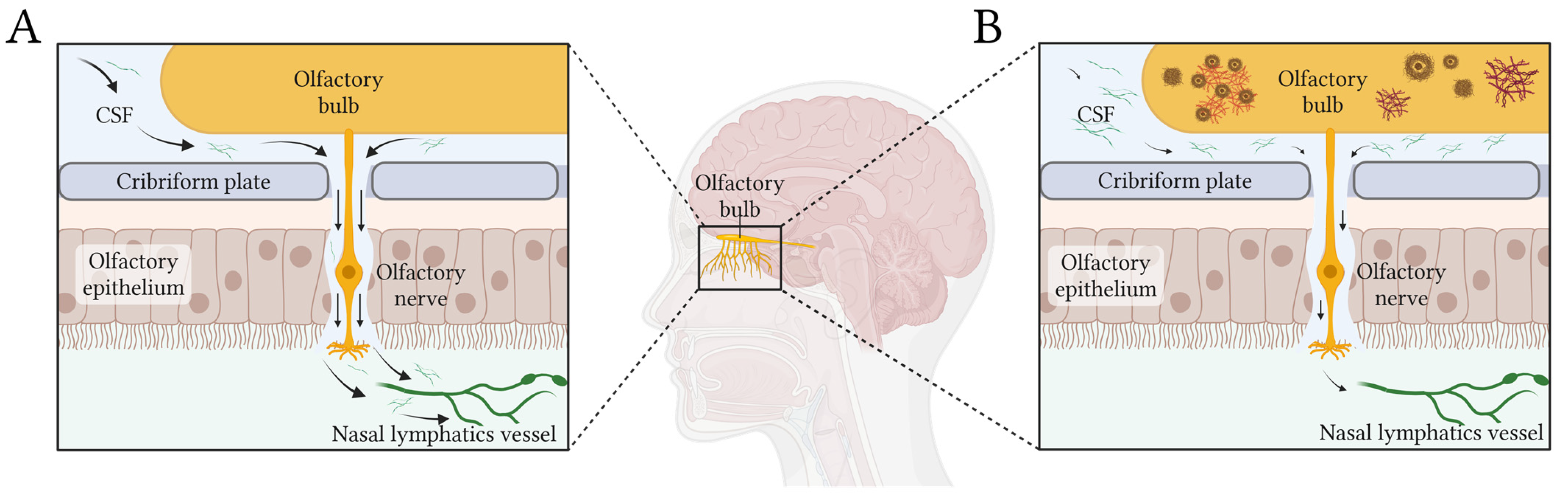
- Nasal pathway is the main CSF egress from the brain. CSF is primarily cleared along olfactory nerves that traverse the cribriform plate, draining into lymphatic vessels in the nasal mucosa [81]. De Leon and colleagues demonstrated, using dynamic PET, that, as in other mammals, human nasal turbinates are part of the CSF egress system [82]. They also observed that such clearance measures were 66% lower in AD patients. Using a different technique, serial MRI after an intrathecal contrast injection, Zhou et al. found concordant results [4]. They observed drainage of CSF to the turbinates via the cribriform plate along the olfactory nerve, this being the main egress pathway. Moreover, clearance function through the peri-olfactory inferior turbinate pathway was diminished with aging and was associated with cognitive function and reported sleep quality [4]. Figure 1 depicts the physiological CSF egress through the perineural olfactory pathway (Figure 1A) and the pathological protein aggregation associated with diminished clearance (Figure 1B).
- (3)
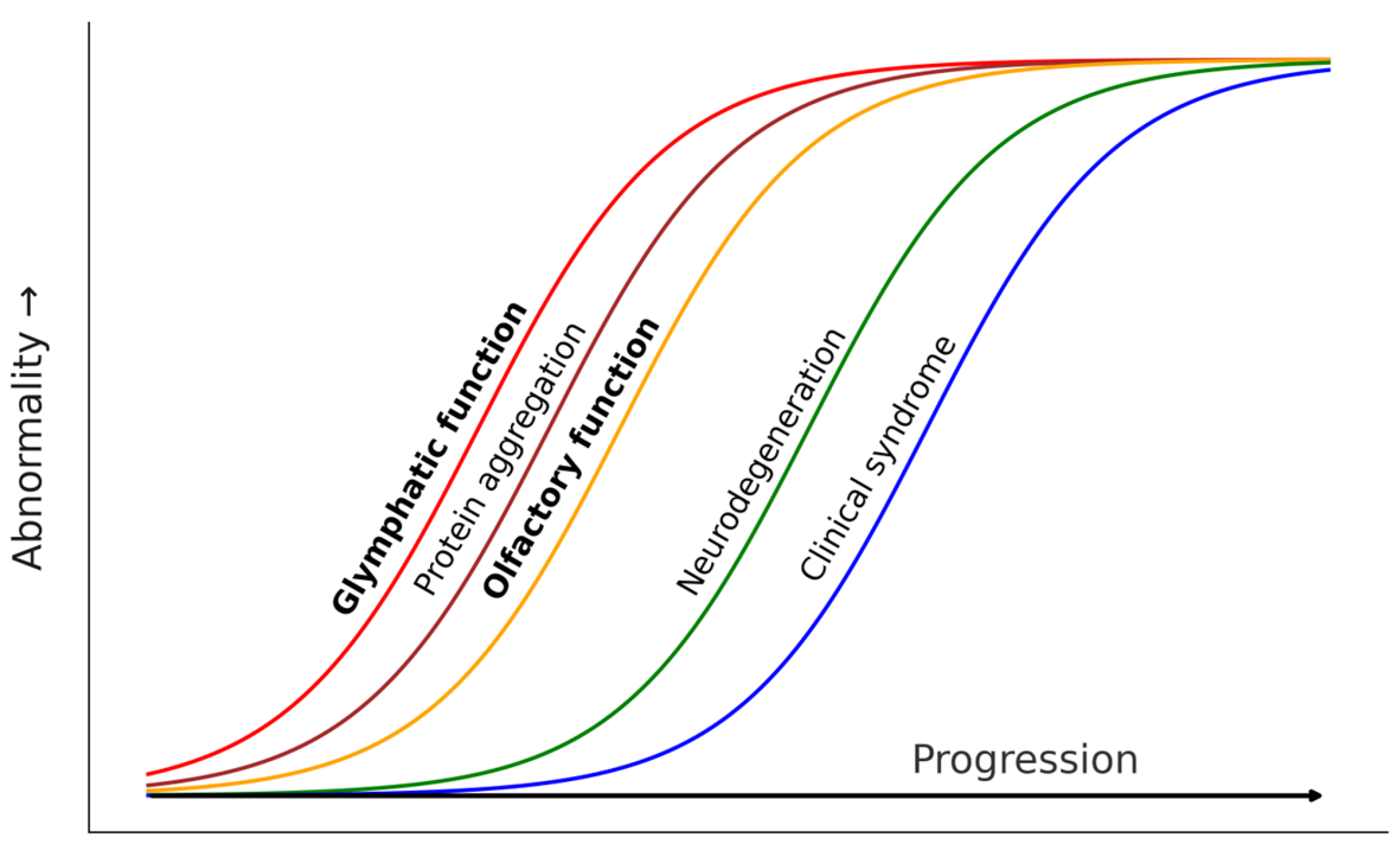
- Glymphatic failure seems to be a very early event in neurodegenerative diseases. Recent evidence suggested that glymphatic function is affected very early in AD. Using diffusion tensor imaging (DTI) along perivascular spaces (ALPS), a neuroimaging proxy of glymphatic function, two large longitudinal studies found that the ALPS index becomes abnormal in individuals with subjective cognitive decline (SCD), before objective cognitive impairment can be detected [83,84]. Moreover, the lower ALPS index predicts accelerated Aβ PET burden and AD signature ROI thinning, higher risk of amyloid-positive transition, and faster cognitive decline [83]. Based on such evidence, Huang et al. proposed a hypothetical cascade model of pathological events in AD, in which reduction in the ALPS index would be the first biomarker to change. The evidence in LBD is similar. Using the DTI-ALPS index as well, Bae et al. found that glymphatic function was diminished in RBD patients and that the lower ALPS index predicted phenoconversion to PD [85]. Accordingly, they concluded that glymphatic impairment is presumed to start at the preclinical stage of PD.
- (4)
- Olfactory dysfunction is associated with loss of estrogen after menopause and parallels glymphatic-related events. Olfactory dysfunction in postmenopausal women seems linked to estrogen loss, paralleling glymphatic dysfunction. Estrogen influences LC-NA regulation, sleep, and vascular function, all of which are critical for glymphatic clearance [59]. Sleep disturbances and abnormal BP patterns, both prevalent after menopause, further compromise glymphatic efficiency and may contribute to AD risk. Experimental and neuroimaging studies support estrogen’s role in olfactory function, suggesting that its decline may accelerate neurodegenerative processes.
6. Concluding Remarks
Author Contributions
Funding
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Iliff, J.J.; Wang, M.; Liao, Y.; Plogg, B.A.; Peng, W.; Gundersen, G.A.; Benveniste, H.; Vates, G.E.; Deane, R.; Goldman, S.A.; et al. A Paravascular Pathway Facilitates CSF Flow through the Brain Parenchyma and the Clearance of Interstitial Solutes, Including Amyloid β. Sci. Transl. Med. 2012, 4, 147ra111. [Google Scholar] [CrossRef] [PubMed]
- Nedergaard, M.; Goldman, S.A. Glymphatic Failure as a Final Common Pathway to Dementia. Science 2020, 370, 50–56. [Google Scholar] [CrossRef] [PubMed]
- Fatuzzo, I.; Niccolini, G.F.; Zoccali, F.; Cavalcanti, L.; Bellizzi, M.G.; Riccardi, G.; de Vincentiis, M.; Fiore, M.; Petrella, C.; Minni, A.; et al. Neurons, Nose, and Neurodegenerative Diseases: Olfactory Function and Cognitive Impairment. Int. J. Mol. Sci. 2023, 24, 2117. [Google Scholar] [CrossRef] [PubMed]
- Zhou, Y.; Ran, W.; Luo, Z.; Wang, J.; Fang, M.; Wei, K.; Sun, J.; Lou, M. Impaired Peri-Olfactory Cerebrospinal Fluid Clearance Is Associated with Ageing, Cognitive Decline and Dyssomnia. EBioMedicine 2022, 86, 104381. [Google Scholar] [CrossRef]
- Doty, R.L.; Kamath, V. The Influences of Age on Olfaction: A Review. Front. Psychol. 2014, 5, 20. [Google Scholar] [CrossRef]
- Schubert, C.R.; Cruickshanks, K.J.; Fischer, M.E.; Huang, G.-H.; Klein, B.E.K.; Klein, R.; Pankow, J.S.; Nondahl, D.M. Olfactory Impairment in an Adult Population: The Beaver Dam Offspring Study. Chem. Senses 2012, 37, 325–334. [Google Scholar] [CrossRef]
- Man, R.E.K.; Wong, C.M.J.; Gupta, P.; Fenwick, E.K.; Aravindhan, A.; Teo, N.W.Y.; Charn, T.C.; Forde, C.; Lamoureux, E.L. Understanding the Prevalence and Risk Factor Profile of Olfactory Impairment and Its Impact on Patient Health Indicators and Economic Outcomes in Community-Dwelling Older Asian Adults. Innov. Aging 2024, 8, igae088. [Google Scholar] [CrossRef]
- Murphy, C.; Schubert, C.R.; Cruickshanks, K.J.; Klein, B.E.K.; Klein, R.; Nondahl, D.M. Prevalence of Olfactory Impairment in Older Adults. JAMA 2002, 288, 2307–2312. [Google Scholar] [CrossRef]
- Attems, J.; Walker, L.; Jellinger, K.A. Olfaction and Aging: A Mini-Review. Gerontology 2015, 61, 485–490. [Google Scholar] [CrossRef]
- Uchida, S.; Shimada, C.; Sakuma, N.; Kagitani, F.; Kan, A.; Awata, S. The Relationship between Olfaction and Cognitive Function in the Elderly. J. Physiol. Sci. 2020, 70, 48. [Google Scholar] [CrossRef]
- Doty, R.L.; Hawkes, C.H.; Good, K.P.; Duda, J.E. Odor Perception and Neuropathology in Neurodegenerative Diseases and Schizophrenia. In Handbook of Olfaction and Gustation; John Wiley & Sons, Inc.: Hoboken, NJ, USA, 2015. [Google Scholar]
- Ross, G.W.; Petrovitch, H.; Abbott, R.D.; Tanner, C.M.; Popper, J.; Masaki, K.; Launer, L.; White, L.R. Association of Olfactory Dysfunction with Risk for Future Parkinson’s Disease. Ann. Neurol. 2008, 63, 167–173. [Google Scholar] [CrossRef] [PubMed]
- Doty, R.L. Olfactory Dysfunction in Neurodegenerative Diseases: Is There a Common Pathological Substrate? Lancet Neurol. 2017, 16, 478–488. [Google Scholar] [CrossRef] [PubMed]
- Murphy, C. Olfactory and Other Sensory Impairments in Alzheimer Disease. Nat. Rev. Neurol. 2019, 15, 11–24. [Google Scholar] [CrossRef] [PubMed]
- Jack, C.R.; Bennett, D.A.; Blennow, K.; Carrillo, M.C.; Dunn, B.; Haeberlein, S.B.; Holtzman, D.M.; Jagust, W.; Jessen, F.; Karlawish, J.; et al. NIA-AA Research Framework: Toward a Biological Definition of Alzheimer’s Disease. Alzheimer’s Dement. 2018, 14, 535–562. [Google Scholar] [CrossRef]
- Hassenstab, J.; Ruvolo, D.; Jasielec, M.; Xiong, C.; Grant, E.; Morris, C.; Hassenstab, J.; Ruvolo, D.; Jasielec, M.; Xiong, C.; et al. Absence of Practice Effects in Preclinical Alzheimer’ s Disease. Neuropsychology 2015, 29, 940. [Google Scholar] [CrossRef]
- Mowrey, W.B.; Lipton, R.B.; Katz, M.J.; Ramratan, W.S.; Loewenstein, D.A.; Zimmerman, M.E.; Buschke, H. Memory Binding Test Predicts Incident Amnestic Mild Cognitive Impairment. J. Alzheimer’s Dis. 2016, 53, 1585–1595. [Google Scholar] [CrossRef]
- Graves, A.B.; Larson, E.B.; Edland, S.D.; Bowen, J.D.; McCormick, W.C.; McCurry, S.M.; Rice, M.M.; Wenzlow, A.; Uomoto, J.M. Prevalence of Dementia and Its Subtypes in the Japanese American Population of King County, Washington State. The Kame Project. Am. J. Epidemiol. 1996, 144, 760–771. [Google Scholar] [CrossRef]
- Calhoun-Haney, R.; Murphy, C. Apolipoprotein Epsilon4 Is Associated with More Rapid Decline in Odor Identification than in Odor Threshold or Dementia Rating Scale Scores. Brain Cogn. 2005, 58, 178–182. [Google Scholar] [CrossRef]
- Devanand, D.P.; Lee, S.; Luchsinger, J.A.; Knopman, D.; Vassilaki, M.; Motter, J.N. Comparison of Brief Olfactory and Cognitive Assessments to Neuroimaging Biomarkers in the Prediction of Cognitive Decline and Dementia in the MCSA Cohort. Alzheimer’s Dement. 2024, 20, 8346–8358. [Google Scholar] [CrossRef]
- Palta, P.; Chen, H.; Deal, J.A.; Sharrett, A.R.; Gross, A.; Knopman, D.; Griswold, M.; Heiss, G.; Mosley, T.H. Olfactory Function and Neurocognitive Outcomes in Old Age: The Atherosclerosis Risk in Communities Neurocognitive Study. Alzheimer’s Dement. 2018, 14, 1015–1021. [Google Scholar] [CrossRef]
- Roberts, R.O.; Christianson, T.J.H.; Kremers, W.K.; Mielke, M.M.; Machulda, M.M.; Vassilaki, M.; Alhurani, R.E.; Geda, Y.E.; Knopman, D.S.; Petersen, R.C. Association Between Olfactory Dysfunction and Amnestic Mild Cognitive Impairment and Alzheimer Disease Dementia. JAMA Neurol. 2016, 73, 93–101. [Google Scholar] [CrossRef] [PubMed]
- Laukka, E.J.; Ekström, I.; Larsson, M.; Grande, G.; Fratiglioni, L.; Rizzuto, D. Markers of Olfactory Dysfunction and Progression to Dementia: A 12-Year Population-Based Study. Alzheimer’s Dement. 2023, 19, 3019–3027. [Google Scholar] [CrossRef] [PubMed]
- Guo, J.; Dove, A.; Wang, J.; Laukka, E.J.; Ekström, I.; Dunk, M.M.; Bennett, D.A.; Xu, W. Trajectories of Olfactory Identification Preceding Incident Mild Cognitive Impairment and Dementia: A Longitudinal Study. EBioMedicine 2023, 98, 104862. [Google Scholar] [CrossRef] [PubMed]
- Tu, L.; Lv, X.; Fan, Z.; Zhang, M.; Wang, H.; Yu, X. Association of Odor Identification Ability With Amyloid-β and Tau Burden: A Systematic Review and Meta-Analysis. Front. Neurosci. 2020, 14, 586330. [Google Scholar] [CrossRef]
- Bothwell, A.R.; Resnick, S.M.; Ferrucci, L.; Tian, Q. Associations of Olfactory Function with Brain Structural and Functional Outcomes. A Systematic Review. Ageing Res. Rev. 2023, 92, 102095. [Google Scholar] [CrossRef]
- Dong, Y.; Li, Y.; Liu, K.; Han, X.; Liu, R.; Ren, Y.; Cong, L.; Zhang, Q.; Hou, T.; Song, L.; et al. Anosmia, Mild Cognitive Impairment, and Biomarkers of Brain Aging in Older Adults. Alzheimer’s Dement. 2023, 19, 589–601. [Google Scholar] [CrossRef]
- Iranzo, A.; Molinuevo, J.L.; Santamaría, J.; Serradell, M.; Martí, M.J.; Valldeoriola, F.; Tolosa, E. Rapid-Eye-Movement Sleep Behaviour Disorder as an Early Marker for a Neurodegenerative Disorder: A Descriptive Study. Lancet Neurol. 2006, 5, 572–577. [Google Scholar] [CrossRef]
- Iranzo, A.; Tolosa, E.; Gelpi, E.; Molinuevo, J.L.; Valldeoriola, F.; Serradell, M.; Sanchez-Valle, R.; Vilaseca, I.; Lomeña, F.; Vilas, D.; et al. Neurodegenerative Disease Status and Post-Mortem Pathology in Idiopathic Rapid-Eye-Movement Sleep Behaviour Disorder: An Observational Cohort Study. Lancet Neurol. 2013, 12, 443–453. [Google Scholar] [CrossRef]
- Schenck, C.H.; Boeve, B.F.; Mahowald, M.W. Delayed Emergence of a Parkinsonian Disorder or Dementia in 81% of Older Men Initially Diagnosed with Idiopathic Rapid Eye Movement Sleep Behavior Disorder: A 16-Year Update on a Previously Reported Series. Sleep Med. 2013, 14, 744–748. [Google Scholar] [CrossRef]
- Mahlknecht, P.; Iranzo, A.; Högl, B.; Frauscher, B.; Müller, C.; Santamaría, J.; Tolosa, E.; Serradell, M.; Mitterling, T.; Gschliesser, V.; et al. Olfactory Dysfunction Predicts Early Transition to a Lewy Body Disease in Idiopathic RBD. Neurology 2015, 84, 654–658. [Google Scholar] [CrossRef]
- Iranzo, A.; Marrero-González, P.; Serradell, M.; Gaig, C.; Santamaria, J.; Vilaseca, I. Significance of Hyposmia in Isolated REM Sleep Behavior Disorder. J. Neurol. 2021, 268, 963–966. [Google Scholar] [CrossRef] [PubMed]
- Postuma, R.B.; Iranzo, A.; Hu, M.; Högl, B.; Boeve, B.F.; Manni, R.; Oertel, W.H.; Arnulf, I.; Ferini-Strambi, L.; Puligheddu, M.; et al. Risk and Predictors of Dementia and Parkinsonism in Idiopathic REM Sleep Behaviour Disorder: A Multicentre Study. Brain 2019, 142, 744–759. [Google Scholar] [CrossRef] [PubMed]
- Braak, H.; Del Tredici, K.; Rüb, U.; de Vos, R.A.I.; Jansen Steur, E.N.H.; Braak, E. Staging of Brain Pathology Related to Sporadic Parkinson’s Disease. Neurobiol. Aging 2003, 24, 197–211. [Google Scholar] [CrossRef] [PubMed]
- Hawkes, C.H.; Del Tredici, K.; Braak, H. Parkinson’s Disease: A Dual-hit Hypothesis. Neuropathol. Appl. Neurobiol. 2007, 33, 599–614. [Google Scholar] [CrossRef]
- Braak, H.; Rüb, U.; Gai, W.P.; Del Tredici, K. Idiopathic Parkinson’s Disease: Possible Routes by Which Vulnerable Neuronal Types May Be Subject to Neuroinvasion by an Unknown Pathogen. J. Neural Transm. 2003, 110, 517–536. [Google Scholar] [CrossRef]
- Sánchez-Ferro, Á.; Rábano, A.; Catalán, M.J.; Rodríguez-Valcárcel, F.C.; Díez, S.F.; Herreros-Rodríguez, J.; García-Cobos, E.; Álvarez-Santullano, M.M.; López-Manzanares, L.; Mosqueira, A.J.; et al. In Vivo Gastric Detection of A-synuclein Inclusions in Parkinson’s Disease. Mov. Disord. 2015, 30, 517–524. [Google Scholar] [CrossRef]
- Kim, C.; Lv, G.; Lee, J.S.; Jung, B.C.; Masuda-Suzukake, M.; Hong, C.-S.; Valera, E.; Lee, H.-J.; Paik, S.R.; Hasegawa, M.; et al. Exposure to Bacterial Endotoxin Generates a Distinct Strain of α-Synuclein Fibril. Sci. Rep. 2016, 6, 30891. [Google Scholar] [CrossRef]
- Forsyth, C.B.; Shannon, K.M.; Kordower, J.H.; Voigt, R.M.; Shaikh, M.; Jaglin, J.A.; Estes, J.D.; Dodiya, H.B.; Keshavarzian, A. Increased Intestinal Permeability Correlates with Sigmoid Mucosa Alpha-Synuclein Staining and Endotoxin Exposure Markers in Early Parkinson’s Disease. PLoS ONE 2011, 6, e28032. [Google Scholar] [CrossRef]
- Knudsen, K.; Fedorova, T.D.; Hansen, A.K.; Sommerauer, M.; Otto, M.; Svendsen, K.B.; Nahimi, A.; Stokholm, M.G.; Pavese, N.; Beier, C.P.; et al. In-Vivo Staging of Pathology in REM Sleep Behaviour Disorder: A Multimodality Imaging Case-Control Study. Lancet Neurol. 2018, 17, 618–628. [Google Scholar] [CrossRef]
- Buongiorno, M.; Marzal, C.; Fernandez, M.; Cullell, N.; de Mena, L.; Sánchez-Benavides, G.; de la Sierra, A.; Krupinski, J.; Compta, Y. Altered Sleep and Neurovascular Dysfunction in Alpha-Synucleinopathies: The Perfect Storm for Glymphatic Failure. Front. Aging Neurosci. 2023, 15, 1251755. [Google Scholar] [CrossRef]
- Rasmussen, M.K.; Mestre, H.; Nedergaard, M. Fluid Transport in the Brain. Physiol. Rev. 2022, 102, 1025–1151. [Google Scholar] [CrossRef] [PubMed]
- Mestre, H.; Hablitz, L.M.; Xavier, A.L.; Feng, W.; Zou, W.; Pu, T.; Monai, H.; Murlidharan, G.; Castellanos Rivera, R.M.; Simon, M.J.; et al. Aquaporin-4-Dependent Glymphatic Solute Transport in the Rodent Brain. Elife 2018, 7, e40070. [Google Scholar] [CrossRef] [PubMed]
- Nedergaard, M. Neuroscience. Garbage Truck of the Brain. Science 2013, 340, 1529–1530. [Google Scholar] [CrossRef]
- Koundal, S.; Elkin, R.; Nadeem, S.; Xue, Y.; Constantinou, S.; Sanggaard, S.; Liu, X.; Monte, B.; Xu, F.; Van Nostrand, W.; et al. Optimal Mass Transport with Lagrangian Workflow Reveals Advective and Diffusion Driven Solute Transport in the Glymphatic System. Sci. Rep. 2020, 10, 1990. [Google Scholar] [CrossRef]
- Iliff, J.J.; Wang, M.; Zeppenfeld, D.M.; Venkataraman, A.; Plog, B.A.; Liao, Y.; Deane, R.; Nedergaard, M. Cerebral Arterial Pulsation Drives Paravascular CSF-Interstitial Fluid Exchange in the Murine Brain. J. Neurosci. 2013, 33, 18190–18199. [Google Scholar] [CrossRef]
- Mestre, H.; Tithof, J.; Du, T.; Song, W.; Peng, W.; Sweeney, A.M.; Olveda, G.; Thomas, J.H.; Nedergaard, M.; Kelley, D.H. Flow of Cerebrospinal Fluid Is Driven by Arterial Pulsations and Is Reduced in Hypertension. Nat. Commun. 2018, 9, 4878. [Google Scholar] [CrossRef]
- Xie, L.; Kang, H.; Xu, Q.; Chen, M.J.; Liao, Y.; Thiyagarajan, M.; O’Donnell, J.; Christensen, D.J.; Nicholson, C.; Iliff, J.J.; et al. Sleep Drives Metabolite Clearance from the Adult Brain. Science 2013, 342, 373–377. [Google Scholar] [CrossRef]
- Louveau, A.; Smirnov, I.; Keyes, T.J.; Eccles, J.D.; Rouhani, S.J.; Peske, J.D.; Derecki, N.C.; Castle, D.; Mandell, J.W.; Lee, K.S.; et al. Structural and Functional Features of Central Nervous System Lymphatic Vessels. Nature 2015, 523, 337–341. [Google Scholar] [CrossRef]
- Henderson, M.X.; Cornblath, E.J.; Darwich, A.; Zhang, B.; Brown, H.; Gathagan, R.J.; Sandler, R.M.; Bassett, D.S.; Trojanowski, J.Q.; Lee, V.M.Y. Spread of α-Synuclein Pathology through the Brain Connectome Is Modulated by Selective Vulnerability and Predicted by Network Analysis. Nat. Neurosci. 2019, 22, 1248–1257. [Google Scholar] [CrossRef]
- Ossenkoppele, R.; Smith, R.; Mattsson-Carlgren, N.; Groot, C.; Leuzy, A.; Strandberg, O.; Palmqvist, S.; Olsson, T.; Jögi, J.; Stormrud, E.; et al. Accuracy of Tau Positron Emission Tomography as a Prognostic Marker in Preclinical and Prodromal Alzheimer Disease: A Head-to-Head Comparison Against Amyloid Positron Emission Tomography and Magnetic Resonance Imaging. JAMA Neurol. 2021, 78, 961–971. [Google Scholar] [CrossRef]
- Ringstad, G.; Vatnehol, S.A.S.; Eide, P.K. Glymphatic MRI in Idiopathic Normal Pressure Hydrocephalus. Brain 2017, 140, 2691–2705. [Google Scholar] [CrossRef] [PubMed]
- Ringstad, G.; Valnes, L.M.; Dale, A.M.; Pripp, A.H.; Vatnehol, S.-A.S.; Emblem, K.E.; Mardal, K.-A.; Eide, P.K. Brain-Wide Glymphatic Enhancement and Clearance in Humans Assessed with MRI. JCI Insight 2018, 3, e121537. [Google Scholar] [CrossRef] [PubMed]
- Hablitz, L.M.; Vinitsky, H.S.; Sun, Q.; Stæger, F.F.; Sigurdsson, B.; Mortensen, K.N.; Lilius, T.O.; Nedergaard, M. Increased Glymphatic Influx Is Correlated with High EEG Delta Power and Low Heart Rate in Mice under Anesthesia. Sci. Adv. 2019, 5, eaav5447. [Google Scholar] [CrossRef]
- Malhotra, R.K. Neurodegenerative Disorders and Sleep. Sleep Med. Clin. 2018, 13, 63–70. [Google Scholar] [CrossRef]
- Mielke, M.M.; Teresa Ferretti, M.; Florencia Iulita, M.; Hayden, K.; Khachaturian, A.S. Sex and Gender in Alzheimer’s Disease—Does It Matter? Alzheimer’s Dement. 2018, 14, 1101–1103. [Google Scholar] [CrossRef]
- Luckey, A.M.; Robertson, I.H.; Lawlor, B.; Mohan, A.; Vanneste, S. Sex Differences in Locus Coeruleus: A Heuristic Approach That May Explain the Increased Risk of Alzheimer’s Disease in Females. J. Alzheimer’s Dis. 2021, 83, 505–522. [Google Scholar] [CrossRef]
- Cui, J.; Shen, Y.; Li, R. Estrogen Synthesis and Signaling Pathways during Aging: From Periphery to Brain. Trends Mol. Med. 2013, 19, 197–209. [Google Scholar] [CrossRef]
- Giorgi, F.S.; Galgani, A.; Puglisi-Allegra, S.; Limanaqi, F.; Busceti, C.L.; Fornai, F. Locus Coeruleus and Neurovascular Unit: From Its Role in Physiology to Its Potential Role in Alzheimer’s Disease Pathogenesis. J. Neurosci. Res. 2020, 98, 2406–2434. [Google Scholar] [CrossRef]
- Peter-Derex, L.; Yammine, P.; Bastuji, H.; Croisile, B. Sleep and Alzheimer’s Disease. Sleep Med. Rev. 2015, 19, 29–38. [Google Scholar] [CrossRef]
- Shi, L.; Chen, S.-J.; Ma, M.-Y.; Bao, Y.-P.; Han, Y.; Wang, Y.-M.; Shi, J.; Vitiello, M.V.; Lu, L. Sleep Disturbances Increase the Risk of Dementia: A Systematic Review and Meta-Analysis. Sleep Med. Rev. 2018, 40, 4–16. [Google Scholar] [CrossRef]
- Geiger, P.J.; Eisenlohr-Moul, T.; Gordon, J.L.; Rubinow, D.R.; Girdler, S.S. Effects of Perimenopausal Transdermal Estradiol on Self-Reported Sleep, Independent of Its Effect on Vasomotor Symptom Bother and Depressive Symptoms. Menopause 2019, 26, 1318–1323. [Google Scholar] [CrossRef] [PubMed]
- Lee, K.; Choi, I.H.; Lee, S.H.; Kim, T.H. Association between Subjective Olfactory Dysfunction and Female Hormone-Related Factors in South Korea. Sci. Rep. 2019, 9, 20007. [Google Scholar] [CrossRef] [PubMed]
- Doty, R.L.; Tourbier, I.; Ng, V.; Neff, J.; Armstrong, D.; Battistini, M.; Sammel, M.D.; Gettes, D.; Evans, D.L.; Mirza, N.; et al. Influences of Hormone Replacement Therapy on Olfactory and Cognitive Function in Postmenopausal Women. Neurobiol. Aging 2015, 36, 2053–2059. [Google Scholar] [CrossRef]
- Bernal-Mondragón, C.; Rivas-Arancibia, S.; Kendrick, K.M.; Guevara-Guzmán, R. Estradiol Prevents Olfactory Dysfunction Induced by A-β 25–35 Injection in Hippocampus. BMC Neurosci. 2013, 14, 104. [Google Scholar] [CrossRef]
- Martinez, B.; Karunanayaka, P.; Wang, J.; Tobia, M.J.; Vasavada, M.; Eslinger, P.J.; Yang, Q.X. Different Patterns of Age-Related Central Olfactory Decline in Men and Women as Quantified by Olfactory FMRI. Oncotarget 2017, 8, 79212–79222. [Google Scholar] [CrossRef]
- Ekanayake, A.; Peiris, S.; Ahmed, B.; Kanekar, S.; Grove, C.; Kalra, D.; Eslinger, P.; Yang, Q.; Karunanayaka, P. A Review of the Role of Estrogens in Olfaction, Sleep and Glymphatic Functionality in Relation to Sex Disparity in Alzheimer’s Disease. Am. J. Alzheimer’s Dis. Other Demen. 2024, 39, 15333175241272025. [Google Scholar] [CrossRef]
- Wali, M.A.; Raparelli, V.; Pilote, L.; Daskalopoulou, S.S. Blood Pressure Variability in Normotensive Perimenopausal Women: Non-Dipping Status, Maximum Blood Pressure and Arterial Stiffness. Int. J. Cardiol. 2021, 325, 149–154. [Google Scholar] [CrossRef]
- Gavriilaki, M.; Anyfanti, P.; Mastrogiannis, K.; Gavriilaki, E.; Lazaridis, A.; Kimiskidis, V.; Gkaliagkousi, E. Association between Ambulatory Blood Pressure Monitoring Patterns with Cognitive Function and Risk of Dementia: A Systematic Review and Meta-Analysis. Aging Clin. Exp. Res. 2023, 35, 745–761. [Google Scholar] [CrossRef]
- Tarumi, T.; Harris, T.S.; Hill, C.; German, Z.; Riley, J.; Turner, M.; Womack, K.B.; Kerwin, D.R.; Monson, N.L.; Stowe, A.M.; et al. Amyloid Burden and Sleep Blood Pressure in Amnestic Mild Cognitive Impairment. Neurology 2015, 85, 1922–1929. [Google Scholar] [CrossRef]
- Dorsey, E.R.; Elbaz, A.; Nichols, E.; Abbasi, N.; Abd-Allah, F.; Abdelalim, A.; Adsuar, J.C.; Ansha, M.G.; Brayne, C.; Choi, J.-Y.J.; et al. Global, Regional, and National Burden of Parkinson’s Disease, 1990–2016: A Systematic Analysis for the Global Burden of Disease Study 2016. Lancet Neurol. 2018, 17, 939–953. [Google Scholar] [CrossRef]
- Zirra, A.; Rao, S.C.; Bestwick, J.; Rajalingam, R.; Marras, C.; Blauwendraat, C.; Mata, I.F.; Noyce, A.J. Gender Differences in the Prevalence of Parkinson’s Disease. Mov. Disord. Clin. Pract. 2023, 10, 86–93. [Google Scholar] [CrossRef] [PubMed]
- Pringsheim, T.; Jette, N.; Frolkis, A.; Steeves, T.D.L. The Prevalence of Parkinson’s Disease: A Systematic Review and Meta-Analysis. Mov. Disord. 2014, 29, 1583–1590. [Google Scholar] [CrossRef] [PubMed]
- Bathini, P.; Mottas, A.; Jaquet, M.; Brai, E.; Alberi, L. Progressive Signaling Changes in the Olfactory Nerve of Patients with Alzheimer’s Disease. Neurobiol. Aging 2019, 76, 80–95. [Google Scholar] [CrossRef] [PubMed]
- Arnold, S.E.; Lee, E.B.; Moberg, P.J.; Stutzbach, L.; Kazi, H.; Han, L.-Y.; Lee, V.M.Y.; Trojanowski, J.Q. Olfactory Epithelium Amyloid-Beta and Paired Helical Filament-Tau Pathology in Alzheimer Disease. Ann. Neurol. 2010, 67, 462–469. [Google Scholar] [CrossRef]
- Saiz-Sanchez, D.; De la Rosa-Prieto, C.; Ubeda-Banon, I.; Martinez-Marcos, A. Interneurons, Tau and Amyloid-β in the Piriform Cortex in Alzheimer’s Disease. Brain Struct. Funct. 2015, 220, 2011–2025. [Google Scholar] [CrossRef]
- Christen-Zaech, S.; Kraftsik, R.; Pillevuit, O.; Kiraly, M.; Martins, R.; Khalili, K.; Miklossy, J. Early Olfactory Involvement in Alzheimer’s Disease. Can. J. Neurol. Sci. 2003, 30, 20–25. [Google Scholar] [CrossRef]
- Attems, J.; Jellinger, K.A. Olfactory Tau Pathology in Alzheimer Disease and Mild Cognitive Impairment. Clin. Neuropathol. 2006, 25, 265–271. [Google Scholar]
- Wuestefeld, A.; Pichet Binette, A.; Berron, D.; Spotorno, N.; van Westen, D.; Stomrud, E.; Mattsson-Carlgren, N.; Strandberg, O.; Smith, R.; Palmqvist, S.; et al. Age-Related and Amyloid-Beta-Independent Tau Deposition and Its Downstream Effects. Brain 2023, 146, 3192–3205. [Google Scholar] [CrossRef]
- Crary, J.F.; Trojanowski, J.Q.; Schneider, J.A.; Abisambra, J.F.; Abner, E.L.; Alafuzoff, I.; Arnold, S.E.; Attems, J.; Beach, T.G.; Bigio, E.H.; et al. Primary Age-Related Tauopathy (PART): A Common Pathology Associated with Human Aging. Acta Neuropathol. 2014, 128, 755–766. [Google Scholar] [CrossRef]
- Chae, J.; Choi, M.; Choi, J.; Yoo, S.-J. The Nasal Lymphatic Route of CSF Outflow: Implications for Neurodegenerative Disease Diagnosis and Monitoring. Anim. Cells Syst. 2024, 28, 45–54. [Google Scholar] [CrossRef]
- de Leon, M.J.; Li, Y.; Okamura, N.; Tsui, W.H.; Saint-Louis, L.A.; Glodzik, L.; Osorio, R.S.; Fortea, J.; Butler, T.; Pirraglia, E.; et al. Cerebrospinal Fluid Clearance in Alzheimer Disease Measured with Dynamic PET. J. Nucl. Med. 2017, 58, 1471–1476. [Google Scholar] [CrossRef] [PubMed]
- Huang, S.-Y.; Zhang, Y.-R.; Guo, Y.; Du, J.; Ren, P.; Wu, B.-S.; Feng, J.-F.; Alzheimer’s Disease Neuroimaging Initiative; Cheng, W.; Yu, J.-T. Glymphatic System Dysfunction Predicts Amyloid Deposition, Neurodegeneration, and Clinical Progression in Alzheimer’s Disease. Alzheimer’s Dement. 2024, 20, 3251–3269. [Google Scholar] [CrossRef]
- Li, Y.; Wang, L.; Zhong, J.; Xu, H.; Han, Y.; Alzheimer’s Disease Neuroimaging Initiative; Zuo, C.; Jiang, J. Impaired Glymphatic Function as a Biomarker for Subjective Cognitive Decline: An Exploratory Dual Cohort Study. Alzheimer’s Dement. 2024, 20, 6542–6555. [Google Scholar] [CrossRef] [PubMed]
- Bae, Y.J.; Kim, J.-M.; Choi, B.S.; Ryoo, N.; Song, Y.S.; Nam, Y.; Yoon, I.-Y.; Cho, S.J.; Kim, J.H. Altered Brain Glymphatic Flow at Diffusion-Tensor MRI in Rapid Eye Movement Sleep Behavior Disorder. Radiology 2023, 307, e221848. [Google Scholar] [CrossRef] [PubMed]
- Ethell, D.W. Disruption of Cerebrospinal Fluid Flow through the Olfactory System May Contribute to Alzheimer’s Disease Pathogenesis. J. Alzheimer’s Dis. 2014, 41, 1021–1030. [Google Scholar] [CrossRef]
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2025 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Sánchez-Benavides, G.; Iranzo, A.; Grau-Rivera, O.; Giraldo, D.M.; Buongiorno, M. Olfactory Dysfunction as a Clinical Marker of Early Glymphatic Failure in Neurodegenerative Diseases. Diagnostics 2025, 15, 719. https://doi.org/10.3390/diagnostics15060719
Sánchez-Benavides G, Iranzo A, Grau-Rivera O, Giraldo DM, Buongiorno M. Olfactory Dysfunction as a Clinical Marker of Early Glymphatic Failure in Neurodegenerative Diseases. Diagnostics. 2025; 15(6):719. https://doi.org/10.3390/diagnostics15060719
Chicago/Turabian StyleSánchez-Benavides, Gonzalo, Alex Iranzo, Oriol Grau-Rivera, Darly Milena Giraldo, and Mariateresa Buongiorno. 2025. "Olfactory Dysfunction as a Clinical Marker of Early Glymphatic Failure in Neurodegenerative Diseases" Diagnostics 15, no. 6: 719. https://doi.org/10.3390/diagnostics15060719
APA StyleSánchez-Benavides, G., Iranzo, A., Grau-Rivera, O., Giraldo, D. M., & Buongiorno, M. (2025). Olfactory Dysfunction as a Clinical Marker of Early Glymphatic Failure in Neurodegenerative Diseases. Diagnostics, 15(6), 719. https://doi.org/10.3390/diagnostics15060719