
Association of Blood NK Cell Phenotype with the Severity of Liver Fibrosis in Patients with Chronic Viral Hepatitis C with Genotype 1 or 3

 ,
,  ,
,  and
and
Abstract
1. Introduction
2. Materials and Methods
2.1. Patients
2.2. Diagnosis of CVHC
2.3. Assessing the Degree of Fibrosis
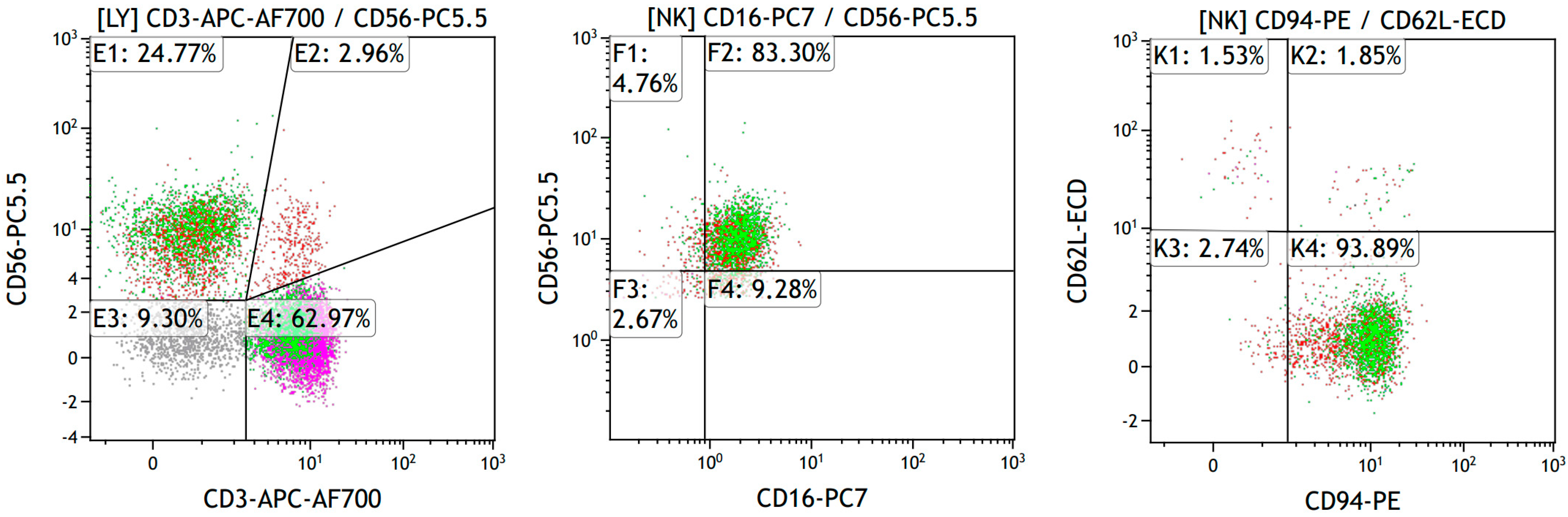
2.4. Assessment of NK Cell Phenotype by Flow Cytometry
2.5. Funding
2.6. Statistical Analysis
3. Results
3.1. Anamnestic, Clinical and Laboratory Parameters of the General Group of Patients with CVHC and Those Who Underwent Immunological Examination
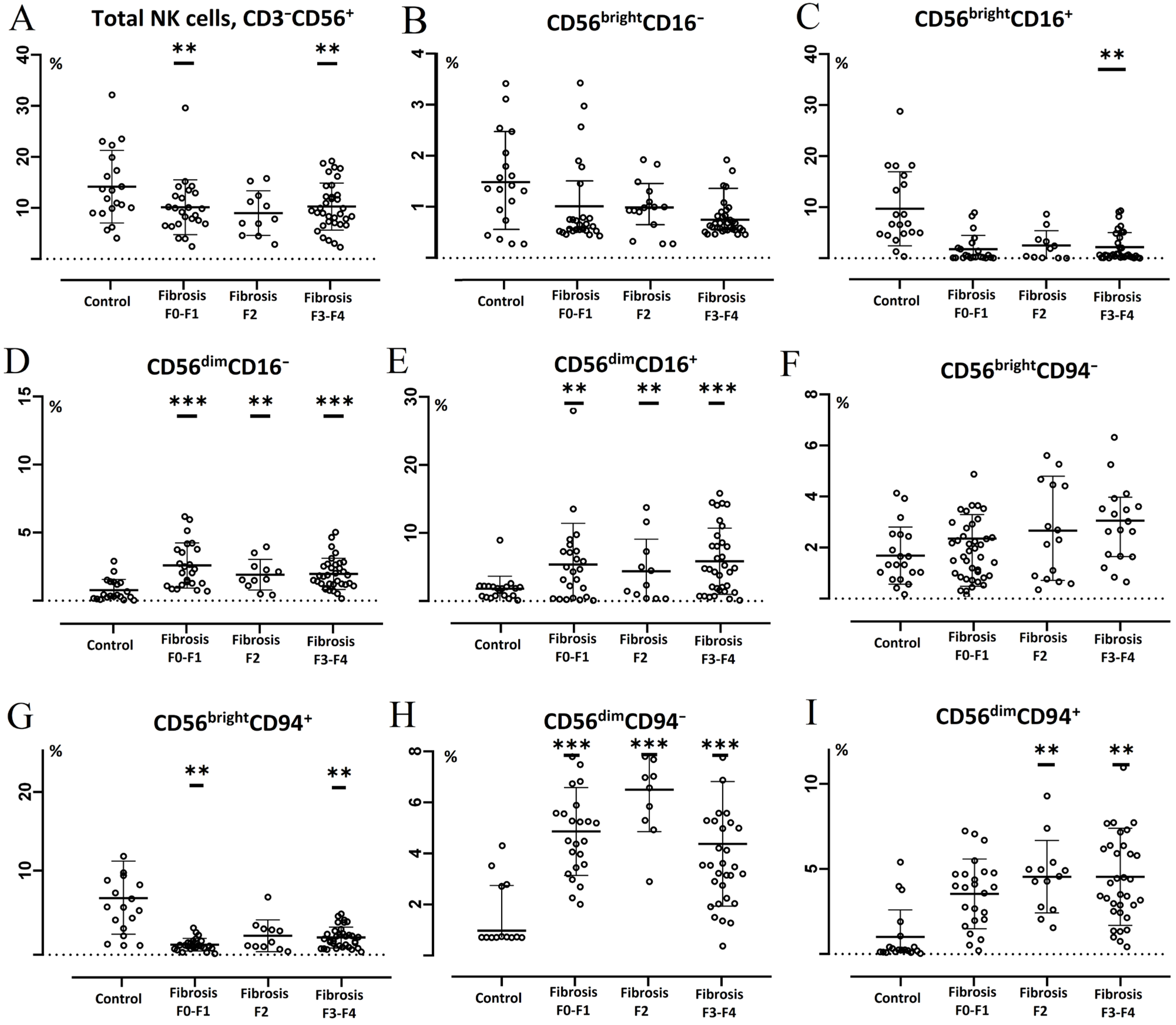
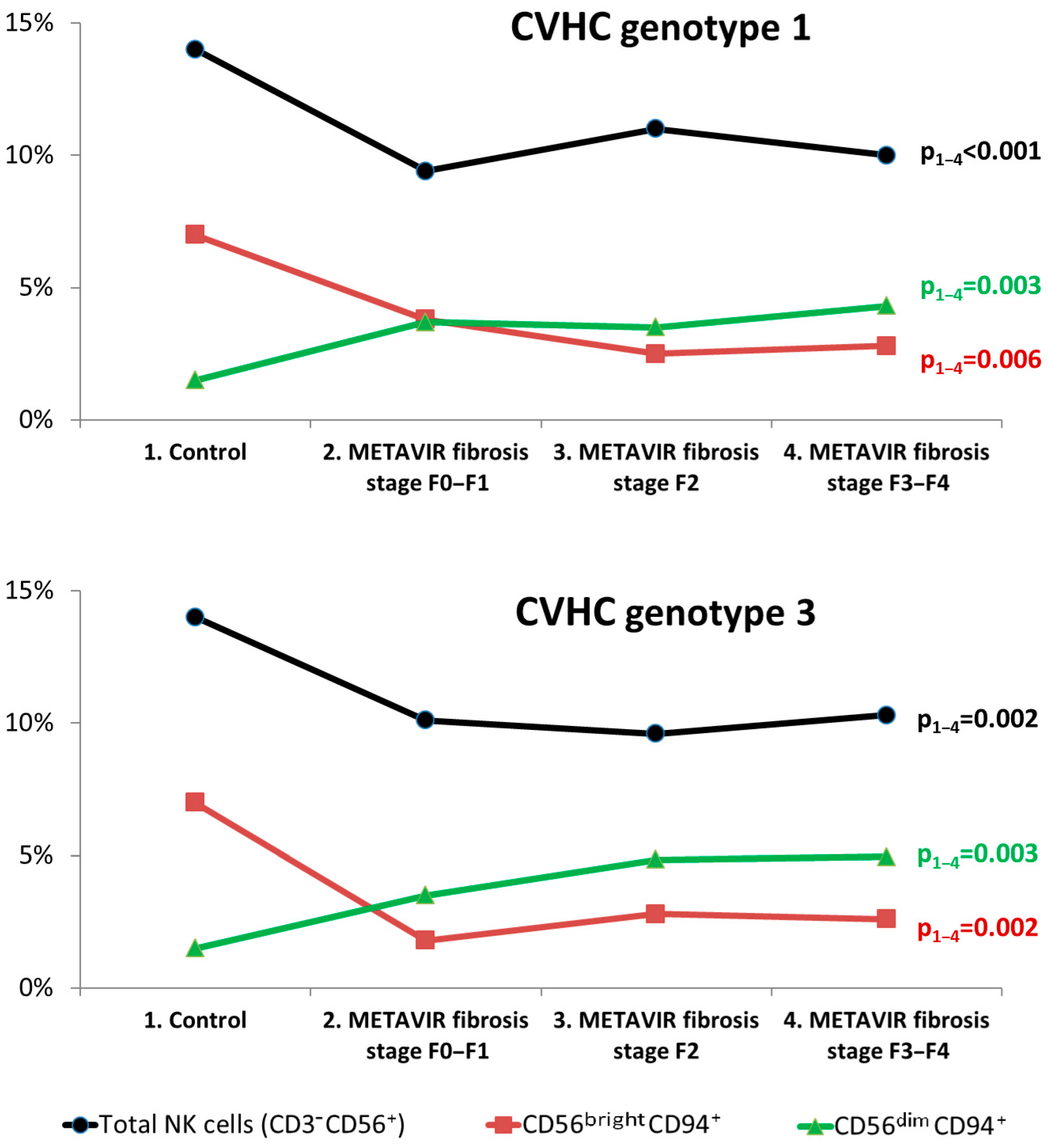
3.2. Subset Composition and Phenotype of NK Cells at Different Liver Fibrosis Degrees in Patients with CVHC
4. Discussion
5. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Bartenschlager, R.; Baumert, T.F.; Bukh, J.; Houghton, M.; Lemon, S.M.; Lindenbach, B.D.; Lohmann, V.; Moradpour, D.; Pietschmann, T.; Rice, C.M.; et al. Critical challenges and emerging opportunities in hepatitis C virus research in an era of potent antiviral therapy: Considerations for scientists and funding agencies. Virus Res. 2018, 248, 53–62. [Google Scholar] [CrossRef]
- de Oliveria Andrade, L.J.; D’Oliveira, A.; Melo, R.C.; De Souza, E.C.; Costa Silva, C.A.; Paraná, R. Association between hepatitis C and hepatocellular carcinoma. J. Glob. Infect. Dis. 2009, 1, 33–37. [Google Scholar] [CrossRef] [PubMed]
- Hayes, C.N.; Imamura, M.; Tanaka, J.; Chayama, K. Road to elimination of HCV: Clinical challenges in HCV management. Liver Int. 2022, 42, 1935–1944. [Google Scholar] [CrossRef]
- Isakov, V.; Nikityuk, D. Elimination of HCV in Russia: Barriers and Perspective. Viruses 2022, 14, 790. [Google Scholar] [CrossRef] [PubMed]
- Pimenov, N.; Kostyushev, D.; Komarova, S.; Fomicheva, A.; Urtikov, A.; Belaia, O.; Umbetova, K.; Darvina, O.; Tsapkova, N.; Chulanov, V. Epidemiology and Genotype Distribution of Hepatitis C Virus in Russia. Pathogens 2022, 11, 1482. [Google Scholar] [CrossRef] [PubMed]
- Shahnazarian, V.; Ramai, D.; Reddy, M.; Mohanty, S. Hepatitis C virus genotype 3: Clinical features, current and emerging viral inhibitors, future challenges. Ann. Gastroenterol. 2018, 31, 541–551. [Google Scholar] [CrossRef]
- Elsheikh, M.E.A.; McClure, C.P.; Tarr, A.W.; Irving, W.L. Sero-reactivity to three distinct regions within the hepatitis C virus alternative reading frame protein (ARFP/core+1) in patients with chronic HCV genotype-3 infection. J. Gen. Virol. 2022, 103, 001727. [Google Scholar] [CrossRef]
- Abulitifu, Y.; Lian, J.; Adilijiang, M.; Liu, L.; Zhao, F.; Qian, W.; Zhang, Y. Effectiveness and Safety of Sofosbuvir-Velpatasvir in Patients with Cirrhosis Associated with Genotype 3 Hepatitis C Infection in Xinjiang, China. Infect. Drug Resist. 2022, 15, 6463–6470. [Google Scholar] [CrossRef]
- Loo, J.H.; Xu, W.X.F.; Low, J.T.; Tay, W.X.; Ang, L.S.; Tam, Y.C.; Thurairajah, P.H.; Kumar, R.; Wong, Y.J. Efficacy and safety of sofosbuvir/velpatasvir with or without ribavirin in hepatitis C genotype 3 compensated cirrhosis: A meta-analysis. World J. Hepatol. 2022, 14, 1248–1257. [Google Scholar] [CrossRef]
- Njiomegnie, G.F.; Read, S.A.; Fewings, N.; George, J.; McKay, F.; Ahlenstiel, G. Immunomodulation of the Natural Killer Cell Phenotype and Response during HCV Infection. J. Clin. Med. 2020, 9, 1030. [Google Scholar] [CrossRef]
- Herberman, R.B.; Nunn, M.E.; Lavrin, D.H. Natural cytotoxic reactivity of mouse lymphoid cells against syngeneic acid allogeneic tumors. I. Distribution of reactivity and specificity. Int. J. Cancer 1975, 16, 216–229. [Google Scholar] [CrossRef] [PubMed]
- Kiessling, R.; Klein, E.; Wigzell, H. “Natural” killer cells in the mouse. I. Cytotoxic cells with specificity for mouse Moloney leukemia cells. Specificity and distribution according to genotype. Eur. J. Immunol. 1975, 5, 112–117. [Google Scholar] [CrossRef] [PubMed]
- Goyos, A.; Fort, M.; Sharma, A.; Lebrec, H. Current Concepts in Natural Killer Cell Biology and Application to Drug Safety Assessments. Toxicol. Sci. 2019, 170, 10–19. [Google Scholar] [CrossRef]
- Mandal, A.; Viswanathan, C. Natural killer cells: In health and disease. Hematol. Oncol. Stem Cell Ther. 2015, 8, 47–55. [Google Scholar] [CrossRef] [PubMed]
- Howell, J.; Visvanathan, K. The role of natural killer cells in hepatitis C infection. Antivir. Ther. 2013, 18, 853–865. [Google Scholar] [CrossRef] [PubMed]
- Björkström, N.K.; Strunz, B.; Ljunggren, H.G. Natural killer cells in antiviral immunity. Nat. Rev. Immunol. 2022, 22, 112–123. [Google Scholar] [CrossRef]
- Mele, D.; Oliviero, B.; Mantovani, S.; Ludovisi, S.; Lombardi, A.; Genco, F.; Gulminetti, R.; Novati, S.; Mondelli, M.U.; Varchetta, S. Adaptive Natural Killer Cell Functional Recovery in Hepatitis C Virus Cured Patients. Hepatology 2021, 73, 79–90. [Google Scholar] [CrossRef]
- Kleczka, A.; Mazur, B.; Tomaszek, K.; Gabriel, A.; Dzik, R.; Kabała-Dzik, A. Association of NK Cells with the Severity of Fibrosis in Patients with Chronic Hepatitis C. Diagnostics 2023, 13, 2187. [Google Scholar] [CrossRef]
- Wei, Y.; Bingyu, W.; Lei, Y.; Xingxing, Y. The antifibrotic role of natural killer cells in liver fibrosis. Exp. Biol. Med. 2022, 247, 1235–1243. [Google Scholar] [CrossRef]
- Yoon, J.C.; Yang, C.M.; Song, Y.; Lee, J.M. Natural killer cells in hepatitis C: Current progress. World J. Gastroenterol. 2016, 22, 1449–1460. [Google Scholar] [CrossRef]
- European Association for the Study of the Liver. EASL Recommendations on Treatment of Hepatitis C 2016. J. Hepatol. 2017, 66, 153–194. [Google Scholar] [CrossRef]
- European Association for the Study of the Liver. EASL Recommendations on Treatment of Hepatitis C 2018. J. Hepatol. 2018, 69, 461–511. [Google Scholar] [CrossRef]
- International Working Party (party leader Zudwig J.). Terminology of chronic hepatitis, hepatic allograft rejection, and nodular lesions of the liver: Summary of recommendations developed by an International Working Party, supported by the World Congresses of Gastroenterology, Los Angeles, 1994. Am. J. Gastroenterol. 1994, 89, S177–S181. [Google Scholar]
- Poynard, T.; Bedossa, P.; Opolon, P. Natural history of liver fibrosis progression in patients with chronic hepatitis C. The OBSVIRC, METAVIR, CLINIVIR, and DOSVIRC groups. Lancet 1997, 349, 825–832. [Google Scholar] [CrossRef] [PubMed]
- Quatrini, L.; Della Chiesa, M.; Sivori, S.; Mingari, M.C.; Pende, D.; Moretta, L. Human NK cells, their receptors and function. Eur. J. Immunol. 2021, 51, 1566–1579. [Google Scholar] [CrossRef] [PubMed]
- Cocker, A.T.H.; Guethlein, L.A.; Parham, P. The CD56-CD16+ NK cell subset in chronic infections. Biochem. Soc. Trans. 2023, 51, 1201–1212. [Google Scholar] [CrossRef] [PubMed]
- Gonzalez, J.C.; Chakraborty, S.; Thulin, N.K.; Wang, T.T. Heterogeneity in IgG-CD16 signaling in infectious disease outcomes. Immunol. Rev. 2022, 309, 64–74. [Google Scholar] [CrossRef] [PubMed]
- Melsen, J.E.; Lugthart, G.; Lankester, A.C.; Schilham, M.W. Human Circulating and Tissue-Resident CD56(bright) Natural Killer Cell Populations. Front. Immunol. 2016, 7, 262. [Google Scholar] [CrossRef]
- Van Acker, H.H.; Capsomidis, A.; Smits, E.L.; Van Tendeloo, V.F. CD56 in the Immune System: More Than a Marker for Cytotoxicity? Front. Immunol. 2017, 8, 892. [Google Scholar] [CrossRef] [PubMed]
- Bahadorian, D.; Mollazadeh, S.; Mirazi, H.; Faraj, T.A.; Kheder, R.K.; Esmaeili, S.A. Regulatory NK cells in autoimmune disease. Iran. J. Basic Med. Sci. 2023, 26, 609–616. [Google Scholar] [CrossRef] [PubMed]
- Lugli, E.; Marcenaro, E.; Mavilio, D. NK Cell Subset Redistribution during the Course of Viral Infections. Front. Immunol. 2014, 5, 390. [Google Scholar] [CrossRef]
- Xie, J.; Xu, B.; Wei, L.; Huang, C.; Liu, W. Effectiveness and Safety of Sofosbuvir/Velpatasvir/Voxilaprevir as a Hepatitis C Virus Infection Salvage Therapy in the Real World: A Systematic Review and Meta-analysis. Infect. Dis. Ther. 2022, 11, 1661–1682. [Google Scholar] [CrossRef] [PubMed]
- Perera Molligoda Arachchige, A.S. Human NK cells: From development to effector functions. Innate Immun. 2021, 27, 212–229. [Google Scholar] [CrossRef] [PubMed]
- Viola, D.; Dona, A.; Caserta, E.; Troadec, E.; Besi, F.; McDonald, T.; Ghoda, L.; Gunes, E.G.; Sanchez, J.F.; Khalife, J.; et al. Daratumumab induces mechanisms of immune activation through CD38+ NK cell targeting. Leukemia 2021, 35, 189–200. [Google Scholar] [CrossRef]
- Kaulfuss, M.; Mietz, J.; Fabri, A.; Vom Berg, J.; Münz, C.; Chijioke, O. The NK cell checkpoint NKG2A maintains expansion capacity of human NK cells. Sci. Rep. 2023, 13, 10555. [Google Scholar] [CrossRef]
- Sivori, S.; Vacca, P.; Del Zotto, G.; Munari, E.; Mingari, M.C.; Moretta, L. Human NK cells: Surface receptors, inhibitory checkpoints, and translational applications. Cell. Mol. Immunol. 2019, 16, 430–441. [Google Scholar] [CrossRef]
- Wu, H.T.; Zhao, X.Y. Regulation of CD38 on Multiple Myeloma and NK Cells by Monoclonal Antibodies. Int. J. Biol. Sci. 2022, 18, 1974–1988. [Google Scholar] [CrossRef]
- Bisht, K.; Fukao, T.; Chiron, M.; Richardson, P.; Atanackovic, D.; Chini, E.; Chng, W.J.; Van De Velde, H.; Malavasi, F. Immunomodulatory properties of CD38 antibodies and their effect on anticancer efficacy in multiple myeloma. Cancer Med. 2023, 12, 20332–20352. [Google Scholar] [CrossRef]
- Di Matteo, S.; Avanzini, M.A.; Pelizzo, G.; Calcaterra, V.; Croce, S.; Spaggiari, G.M.; Theuer, C.; Zuccotti, G.; Moretta, L.; Pelosi, A.; et al. Neuroblastoma Tumor-Associated Mesenchymal Stromal Cells Regulate the Cytolytic Functions of NK Cells. Cancers 2022, 15, 19. [Google Scholar] [CrossRef] [PubMed]
- Neo, S.Y.; Yang, Y.; Record, J.; Ma, R.; Chen, X.; Chen, Z.; Tobin, N.P.; Blake, E.; Seitz, C.; Thomas, R.; et al. CD73 immune checkpoint defines regulatory NK cells within the tumor microenvironment. J. Clin. Invest. 2020, 130, 1185–1198. [Google Scholar] [CrossRef]
- Shafaghat, Z.; Ghomi, A.K.; Khorramdelazad, H.; Safari, E. Purinergic signaling: Decoding its role in COVID-19 pathogenesis and promising treatment strategies. Inflammopharmacology 2023, 31, 3005–3020. [Google Scholar] [CrossRef] [PubMed]


{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Antibodies | Fluorochrome | Protein Clone | Catalog Number |
|---|---|---|---|
| Panel 1 | |||
| CD3 | Alexa Fluor 700 | UCHT1 | B10823 |
| CD16 | Phycoerythrin-cyanin 7 | 3G8 | 6607118 |
| CD45 | Alexa Fluor 750 | J33 | A79392 |
| CD56 | R-Phycoerythrin-cyanine 5.1 | N901 | A07789 |
| CD94 | Phycoerythrin | R34.34 | IM1980U |
| Panel 2 | |||
| CD3 | Alexa Fluor 700 | UCHT1 | B10823 |
| CD38 | Fluorescein isothiocyanate | T16 | A07778 |
| CD45 | Alexa Fluor 750 | J33 | A79392 |
| CD56 | R-Phycoerythrin-cyanine 5.1 | N901 | A07789 |
| CD73 | Phycoerythrin | AD-2 | B68176 |
| Parameters | CVHC with Genotype 1 Total (n = 297) | CVHC with Genotype 3 Total (n = 231) | p |
|---|---|---|---|
| Men/women, n | 164/133 | 125/106 | 0.800 |
| Age (years), Me (IQR) | 41 (38−46) | 41 (38−45) | 0.623 |
| BMI, kg/m2 Me (IQR) | 24 (22−25) | 25 (22−27) | 0.518 |
| Tobacco smoking, n (%) | 17 (5.7%) | 14 (6.1%) | 0.871 |
| Alcohol consumption, n (%) | 33 (11.1%) | 24 (10.4%) | 0.791 |
| White blood cells, 109/L Me (IQR) | 6.40 (5.26−8.33) | 6.58 (5.40−8.36) | 0.234 |
| Neutrophils, % Me (IQR) | 54.4 (48.8−60.0) | 55.4 (48.8−60.0) | 0.932 |
| Lymphocytes, % Me (IQR) | 33.3 (28.7−39.1) | 33.3 (29.7−38.7) | 0.702 |
| Platelets, 109/L Me (IQR) | 230 (202−280) | 239 (201−283) | 0.906 |
| ALT, IU Me (IQR) | 68 (30−85) | 70 (39.2−142) | 0.033 |
| Albumin, g/L Me (IQR) | 42 (38−45) | 41 (38−44) | 0.212 |
| Bilirubin, µmol/L Me (IQR) | 15.4 (11.5−25.6) | 15.0 (12.0−23.4) | 0.504 |
| Parameters | CVHC with Genotype 1 Total (n = 74) | CVHC with Genotype 3 Total (n = 69) | Control Group (n = 20) |
|---|---|---|---|
| Age (years), Me (IQR) | 43 (38−47) | 42 (38−46) | 42 (37−47) |
| p1–2 = 0.849 | p1–3 = 0.868 p2–3 = 0.934 | ||
| BMI, kg/m2 Me (IQR) | 24 (22−26) | 25 (22−27) p1–2 = 0.831 | 24 (21−27) p1–3 = 0.883 p2–3 = 0.911 |
| Tobacco smoking, n (%) | 4 (5.4%) | 4 (5.8%) p1–2 = 0.919 | 1 (5.0%) p1–3 > 0.943 p2–3 = 0.892 |
| Alcohol consumption, n (%) | 8 (10.8%) | 7 (10.1%) p1–2 = 0.897 | 0 (0.0%) p1–3 = 0.125 p2–3 = 0.138 |
| White blood cells, 109/L Me (IQR) | 6.42 (5.14−8.32) | 6.60 (5.31−8.39) p1–2 = 0.623 | 6.23 (4.98−7.56) p1–3 = 0.541 p2–3 = 0.460 |
| Neutrophils, % Me (IQR) | 54.7 (48.2−60.5) | 55.2 (49.1−59.8) p1–2 = 0.973 | 57.4 (50.9−63.1) p1–3 = 0.243 p2–3 = 0.319 |
| Lymphocytes, % Me (IQR) | 33.5 (28.5−39.4) | 33.2 (29.3−39.1) p1–2 = 0.962 | 29.8 (23.7−34.5) p1–3 = 0.168 p2–3 = 0.194 |
| Platelets, 109/L Me (IQR) | 232 (202−282) | 238 (200−284) p1–2 = 0.926 | 247 (205−297) p1–3 = 0.788 p2–3 = 0.823 |
| ALT, IU Me (IQR) | 67 (29−86) | 70 (38−145) p1–2 = 0.217 | 22 (13−30) p1–3 = 0.032 p2–3 = 0.008 |
| Albumin, g/L Me (IQR) | 42 (38−46) | 40 (37−44) p1–2 = 0.576 | 44 (39−48) p1–3 = 0.361 p2–3 = 0.122 |
| Bilirubin, µmol/L Me (IQR) | 15.3 (11.5−25.2) | 15.1 (11.8−23.9) p1–2 = 0.681 | 10.8 (5.3−15.6) p1–3 = 0.031 p2–3 = 0.038 |
| Liver Fibrosis | METAVIR Fibrosis Stage F0–F1 | METAVIR Fibrosis Stage F2 | METAVIR Fibrosis Stage F3–F4 |
|---|---|---|---|
| CVHC patients with HCV genotype 1 (n = 297) | 201 (67.7%) | 35 (11.8%) | 61 (20.5%) |
| CVHC patients with HCV genotype 3 (n = 231) | 107 (46.3%) | 49 (21.2%) | 75 (32.5%) |
| OR (CI) | 0.41 (0.29–0.59) p < 0.001 | 2.02 (1.26–3.23) p = 0.004 | 1.86 (1.25–2.76) p = 0.003 |
| Parameters | Control | Patients with CVHC | ||
|---|---|---|---|---|
| Fibrosis F0–F1 | Fibrosis F2 | Fibrosis F3–F4 | ||
| CD56 + CD38−CD73− | 1.46 (1.19–3.01) | 0.75 (0.26–1.23) p1 < 0.001 | 0.84 (0.52–1.00) p1 = 0.016 | 0.77 (0.55–1.34) p1 = 0.021 |
| CD56+CD38−CD73+ | 0.001 (0.0–0.011) | 0.065 (0.030–0.125) p1 < 0.001 | 0.121 (0.064–0.144) p1 < 0.001 | 0.059 (0.051–0.121) p1 < 0.001 |
| CD56+CD38+CD73− | 10.56 (5.51–14.82) | 5.95 (3.14–9.49) p1 = 0.020 | 4.63 (1.42–6.96) p1 = 0.011 | 3.45 (1.82–10.64) p1 = 0.023 |
| CD56+CD38+CD73+ | 0.001 (0.0–0.102) | 0.631 (0.191–0.845) p1 < 0.001 | 0.234 (0.075–0.330) p1 < 0.001 p2 = 0.024 | 0.809 (0.386–1.300) p1 < 0.001 p3 = 0.007 |
| Parameters | Control | Patients with CVHC | ||
|---|---|---|---|---|
| Fibrosis F0–F1 | Fibrosis F2 | Fibrosis F3–F4 | ||
| CD56+CD38−CD73− | 1.46 (1.19–3.01) | 0.82 (0.26–1.36) p1 = 0.008 | 0.63 (0.28–1.21) p1 = 0.004 | 0.62 (0.37–1.39) p1 = 0.002 |
| CD56+CD38−CD73+ | 0.001 (0.0–0.011) | 0.053 (0.044–0.122) p1 < 0.001 | 0.045 (0.042–0.057) p1 < 0.001 | 0.068 (0.028–0.104) p1 < 0.001 |
| CD56+CD38+CD73− | 10.56 (5.51–14.82) | 5.13 (3.39–6.45) p1 = 0.004 | 1.51 (0.94–5.84) p1 = 0.004 | 5.62 (2.58–7.63) p1 = 0.004 |
| CD56+CD38+CD73+ | 0.001 (0.0–0.102) | 0.498 (0.128–0.768) p1 < 0.001 | 0.310 (0.152–0.532) p1 < 0.001 | 0.674 (0.421–1.163) p1 < 0.001 p3 = 0.004 |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2024 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Tsukanov, V.V.; Savchenko, A.A.; Cherepnin, M.A.; Kasparov, E.V.; Tikhonova, E.P.; Vasyutin, A.V.; Tonkikh, J.L.; Anisimova, A.A.; Belenyuk, V.D.; Borisov, A.G. Association of Blood NK Cell Phenotype with the Severity of Liver Fibrosis in Patients with Chronic Viral Hepatitis C with Genotype 1 or 3. Diagnostics 2024, 14, 472. https://doi.org/10.3390/diagnostics14050472
Tsukanov VV, Savchenko AA, Cherepnin MA, Kasparov EV, Tikhonova EP, Vasyutin AV, Tonkikh JL, Anisimova AA, Belenyuk VD, Borisov AG. Association of Blood NK Cell Phenotype with the Severity of Liver Fibrosis in Patients with Chronic Viral Hepatitis C with Genotype 1 or 3. Diagnostics. 2024; 14(5):472. https://doi.org/10.3390/diagnostics14050472
Chicago/Turabian StyleTsukanov, Vladislav Vladimirovich, Andrei Anatolyevich Savchenko, Mikhail Aleksandrovich Cherepnin, Eduard Vilyamovich Kasparov, Elena Petrovna Tikhonova, Alexander Viktorovich Vasyutin, Julia Leongardovna Tonkikh, Anna Alexandrovna Anisimova, Vasily Dmitrievich Belenyuk, and Alexandr Gennadyevich Borisov. 2024. "Association of Blood NK Cell Phenotype with the Severity of Liver Fibrosis in Patients with Chronic Viral Hepatitis C with Genotype 1 or 3" Diagnostics 14, no. 5: 472. https://doi.org/10.3390/diagnostics14050472
APA StyleTsukanov, V. V., Savchenko, A. A., Cherepnin, M. A., Kasparov, E. V., Tikhonova, E. P., Vasyutin, A. V., Tonkikh, J. L., Anisimova, A. A., Belenyuk, V. D., & Borisov, A. G. (2024). Association of Blood NK Cell Phenotype with the Severity of Liver Fibrosis in Patients with Chronic Viral Hepatitis C with Genotype 1 or 3. Diagnostics, 14(5), 472. https://doi.org/10.3390/diagnostics14050472