
Altered Lnc-EGFR, SNHG1, and LincRNA-Cox2 Profiles in Patients with Relapsing-Remitting Multiple Sclerosis: Impact on Disease Activity and Progression
 ,
,
Abstract
1. Introduction
2. Subjects and Methods
2.1. Participants
2.2. Sample Collection and Biochemical Measurements
2.3. Total RNA Extraction and Reverse Transcription
2.4. Quantitative Real-Time Polymerase Chain Reaction (qRT-PCR)
2.5. TGF-β1 and IL-1β Levels Using Enzyme-Linked Immunosorbent Assay (ELISA)
3. Statistical Analyses
4. Results
4.1. Demographic and Clinical Characteristics of the Study Participants
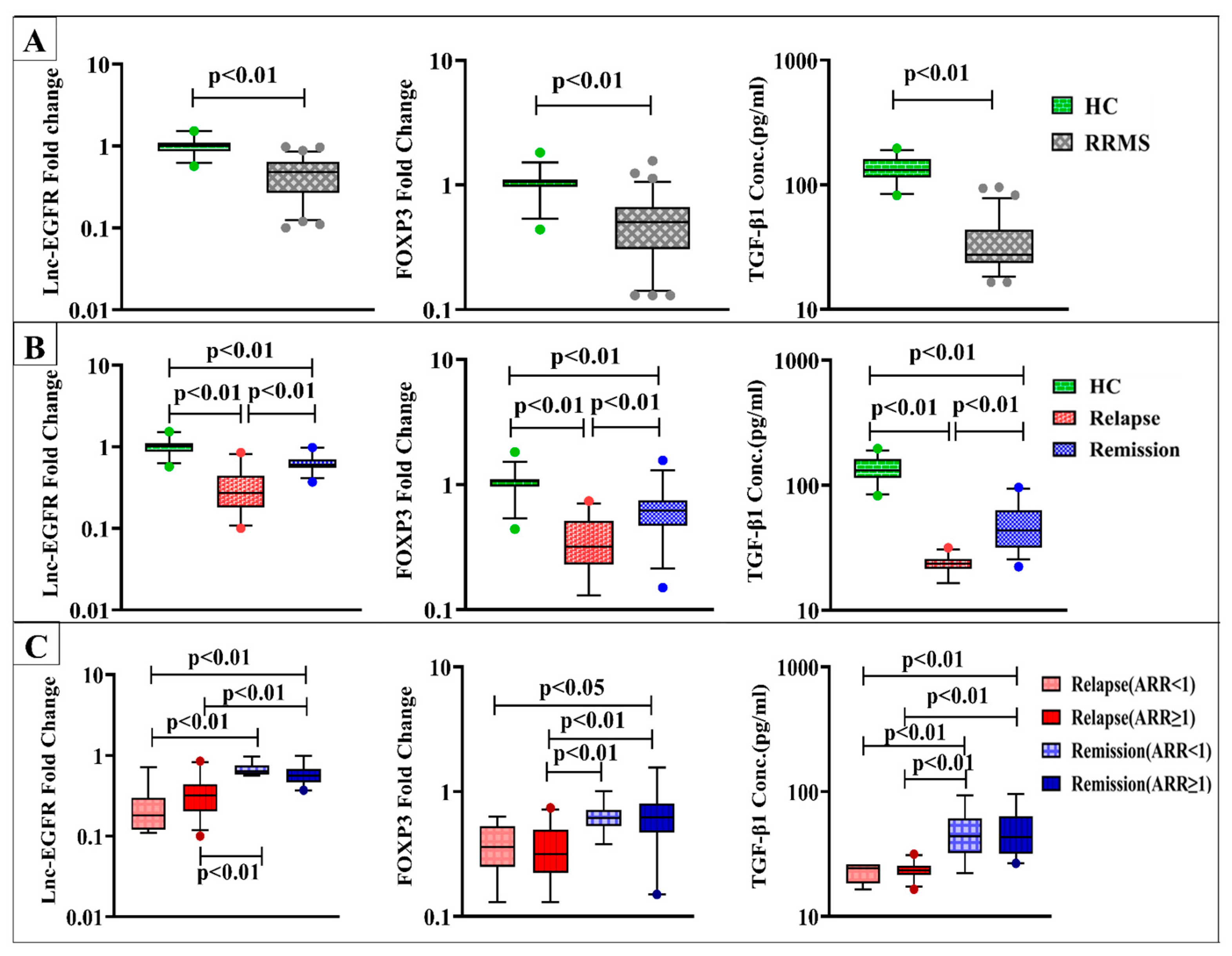
4.2. Levels of lnc-EGFR, FOXP3, and TGF-β1
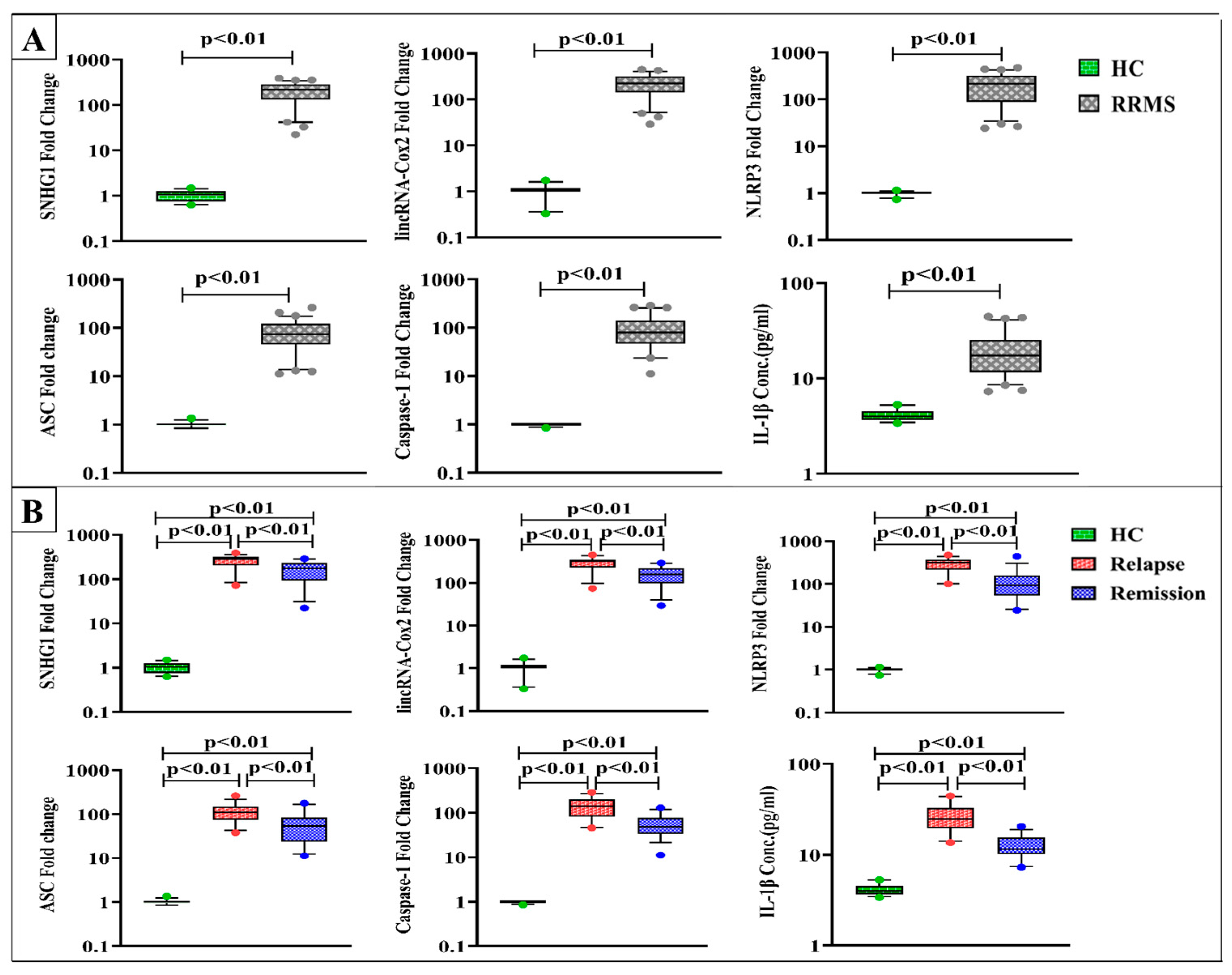
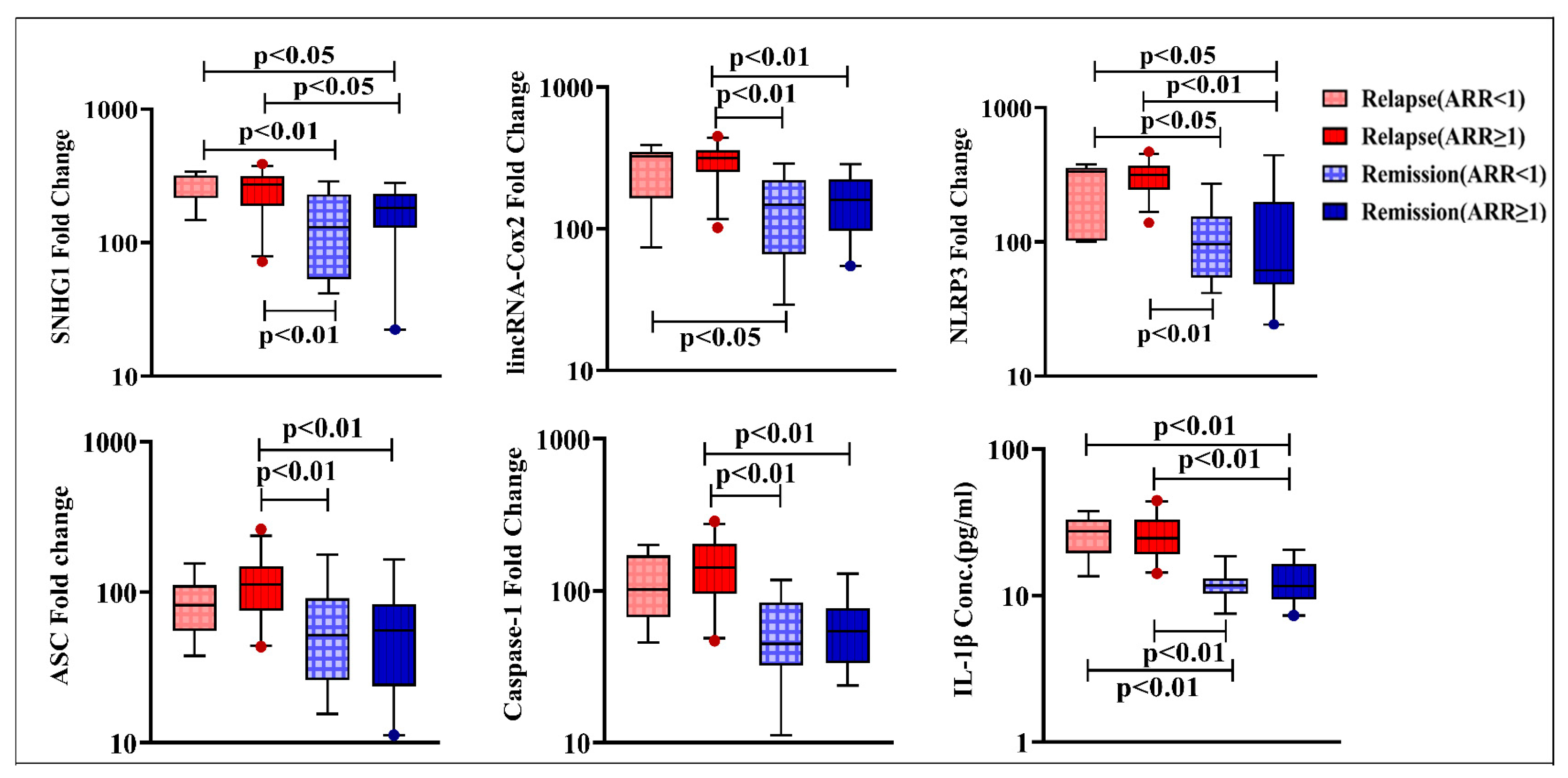
4.3. Levels of SNHG1, lincRNA-Cox2, NLRP3, ASC, Caspase-1, and IL-1β
4.4. Correlation Analyses of Lnc-EGFR, SNHG1, and LincRNA-Cox2 Levels
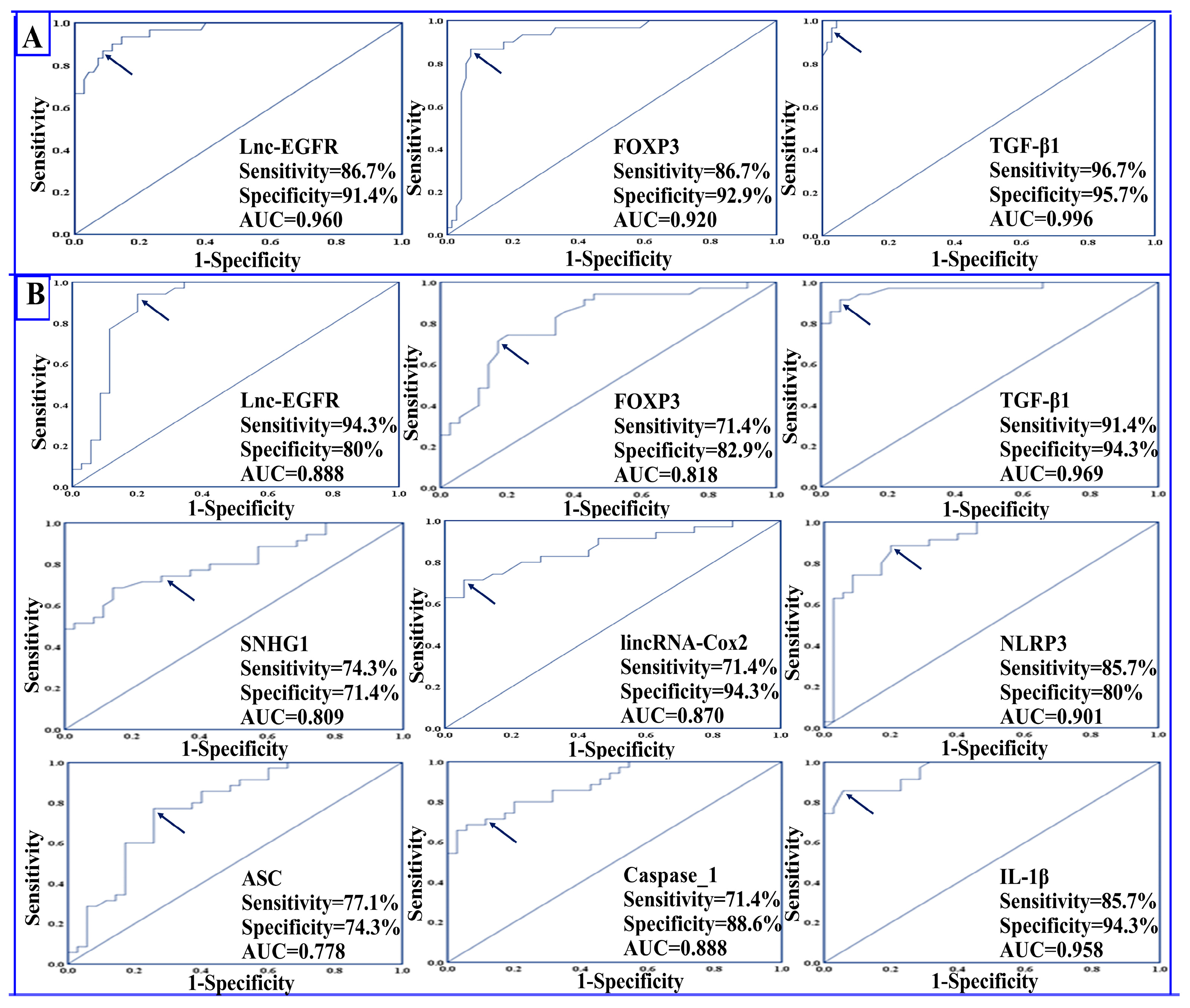
4.5. Diagnostic Potential of the Studied Parameters
4.6. Prognostic Potential of the Studied Parameters
4.7. Univariant Binary Logistic Regression Analysis
5. Discussion
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Tuosto, L. Targeting inflammatory T cells in multiple sclerosis: Current therapies and future challenges. Austin J. Mult. Scler. Neuroimmunol. 2015, 2, 1009. [Google Scholar] [CrossRef]
- Dobson, R.; Giovannoni, G. Multiple sclerosis—A review. Eur. J. Neurol. 2019, 26, 27–40. [Google Scholar] [CrossRef] [PubMed]
- Lublin, F.D.; Reingold, S.C.; Cohen, J.A.; Cutter, G.R.; Sørensen, P.S.; Thompson, A.J.; Wolinsky, J.S.; Balcer, L.J.; Banwell, B.; Barkhof, F.; et al. Defining the clinical course of multiple sclerosis: The 2013 revisions. Neurology 2014, 83, 278–286. [Google Scholar] [CrossRef] [PubMed]
- Kalincik, T. Multiple Sclerosis Relapses: Epidemiology, Outcomes and Management. A Systematic Review. Neuroepidemiology 2015, 44, 199–214. [Google Scholar] [CrossRef] [PubMed]
- Kimura, K. Regulatory T cells in multiple sclerosis. Clin. Exp. Immunol. 2020, 11, 148–155. [Google Scholar] [CrossRef]
- Calahorra, L.; Camacho-Toledano, C.; Serrano-Regal, M.P.; Ortega, M.C.; Clemente, D. Regulatory Cells in Multiple Sclerosis: From Blood to Brain. Biomedicines 2022, 10, 335. [Google Scholar] [CrossRef]
- Li, Z.; Li, D.; Tsun, A.; Li, B. FOXP3+ regulatory T cells and their functional regulation. Cell. Mol. Immunol. 2015, 12, 558–565. [Google Scholar] [CrossRef]
- Williams, L.M.; Rudensky, A.Y. Maintenance of the Foxp3-dependent developmental program in mature regulatory T cells requires continued expression of Foxp3. Nat. Immunol. 2007, 8, 277–284. [Google Scholar] [CrossRef]
- Zozulya, A.L.; Wiendl, H. The role of regulatory T cells in multiple sclerosis. Nat. Clin. Pract. Neurol. 2008, 4, 384–398. [Google Scholar] [CrossRef]
- Jamshidian, A.; Shaygannejad, V.; Pourazar, A.; Zarkesh-Esfahani, S.-H.; Gharagozloo, M. Biased Treg/Th17 balance away from regulatory toward inflammatory phenotype in relapsed multiple sclerosis and its correlation with severity of symptoms. J. Neuroimmunol. 2013, 262, 106–112. [Google Scholar] [CrossRef]
- Venken, K.; Hellings, N.; Thewissen, M.; Somers, V.; Hensen, K.; Rummens, J.L.; Medaer, R.; Hupperts, R.; Stinissen, P. Compromised CD4+ CD25(high) regulatory T-cell function in patients with relapsing-remitting multiple sclerosis is correlated with a reduced frequency of FOXP3-positive cells and reduced FOXP3 expression at the single-cell level. Immunology 2008, 123, 79–89. [Google Scholar] [CrossRef] [PubMed]
- Yavarpour-Bali, H.; Ghasemi-Kasman, M. The role of inflammasomes in multiple sclerosis. Mult. Scler. 2021, 27, 1323–1331. [Google Scholar] [CrossRef] [PubMed]
- Voet, S.; Srinivasan, S.; Lamkanfi, M.; van Loo, G. Inflammasomes in neuroinflammatory and neurodegenerative diseases. EMBO Mol. Med. 2019, 11, e10248. [Google Scholar] [CrossRef] [PubMed]
- Barclay, W.; Shinohara, M.L. Inflammasome activation in multiple sclerosis and experimental autoimmune encephalomyelitis (EAE). Brain Pathol. 2017, 27, 213–219. [Google Scholar] [CrossRef]
- Inoue, M.; Williams, K.L.; Gunn, M.D.; Shinohara, M.L. NLRP3 inflammasome induces chemotactic immune cell migration to the CNS in experimental autoimmune encephalomyelitis. Proc. Natl. Acad. Sci. USA 2012, 109, 10480–10485. [Google Scholar] [CrossRef]
- Gharagozloo, M.; Gris, K.V.; Mahvelati, T.; Amrani, A.; Lukens, J.R.; Gris, D. NLR-Dependent Regulation of Inflammation in Multiple Sclerosis. Front. Immunol. 2017, 8, 2012. [Google Scholar] [CrossRef]
- Shao, S.; Chen, C.; Shi, G.; Zhou, Y.; Wei, Y.; Fan, N.; Yang, Y.; Wu, L.; Zhang, T. Therapeutic potential of the target on NLRP3 inflammasome in multiple sclerosis. Pharmacol. Ther. 2021, 227, 107880. [Google Scholar] [CrossRef]
- Peelen, E.; Damoiseaux, J.; Muris, A.-H.; Knippenberg, S.; Smolders, J.; Hupperts, R.; Thewissen, M. Increased inflammasome related gene expression profile in PBMC may facilitate T helper 17 cell induction in multiple sclerosis. Mol. Immunol. 2015, 63, 521–529. [Google Scholar] [CrossRef]
- Mamik, M.K.; Power, C. Inflammasomes in neurological diseases: Emerging pathogenic and therapeutic concepts. Brain 2017, 140, 2273–2285. [Google Scholar] [CrossRef]
- Keane, R.W.; Dietrich, W.D.; de Rivero Vaccari, J.P. Inflammasome Proteins As Biomarkers of Multiple Sclerosis. Front. Neurol. 2018, 9, 135. [Google Scholar] [CrossRef]
- Yousuf, A.; Qurashi, A. Non-coding RNAs in the Pathogenesis of Multiple Sclerosis. Front. Genet. 2021, 12, 717922. [Google Scholar] [CrossRef] [PubMed]
- Haridy, S.F.; Shahin, N.N.; Shabayek, M.I.; Selim, M.M.; Abdelhafez, M.A.; Motawi, T.K. Diagnostic and prognostic value of the RUNXOR/RUNX1 axis in multiple sclerosis. Neurobiol. Dis. 2023, 178, 106032. [Google Scholar] [CrossRef] [PubMed]
- Pandey, R.; Prakash, V.; Verma, S.; Sankhwar, S.N.; Ahmad, M.K. Circulating serum levels of Fox P3, GATA-3 and IL-17 A as potential biomarkers in patients with symptomatic asthma. J. Commun. Dis. 2019, 51, 28–37. [Google Scholar]
- Bocchetti, M.; Scrima, M.; Melisi, F.; Luce, A.; Sperlongano, R.; Caraglia, M.; Zappavigna, S.; Cossu, A. LncRNAs and Immunity: Coding the Immune System with Noncoding Oligonucleotides. J. Neuroimmunol. 2021, 22, 1741. [Google Scholar] [CrossRef]
- Wang, W.; Yang, N.; Yang, Y.-H.; Wen, R.; Liu, C.-F.; Zhang, T.-N. Non-Coding RNAs: Master Regulators of Inflammasomes in Inflammatory Diseases. J. Inflamm. Res. 2021, 14, 5023–5050. [Google Scholar] [CrossRef] [PubMed]
- Han, Z.; Hua, J.; Xue, W.; Zhu, F. Integrating the Ribonucleic Acid Sequencing Data from Various Studies for Exploring the Multiple Sclerosis-Related Long Noncoding Ribonucleic Acids and Their Functions. Front. Genet. 2019, 10, 1136. [Google Scholar] [CrossRef]
- Jiang, R.; Tang, J.; Chen, Y.; Deng, L.; Ji, J.; Xie, Y.; Wang, K.; Jia, W.; Chu, W.-M.; Sun, B. The long noncoding RNA lnc-EGFR stimulates T-regulatory cells differentiation thus promoting hepatocellular carcinoma immune evasion. Nat. Commun. 2017, 8, 15129. [Google Scholar] [CrossRef]
- Kraus, T.F.J.; Haider, M.; Spanner, J.; Steinmaurer, M.; Dietinger, V.; Kretzschmar, H.A. Altered Long Noncoding RNA Expression Precedes the Course of Parkinson’s Disease-a Preliminary Report. Mol. Neurobiol. 2017, 54, 2869–2877. [Google Scholar] [CrossRef]
- Cao, B.; Wang, T.; Qu, Q.; Kang, T.; Yang, Q. Long Noncoding RNA SNHG1 Promotes Neuroinflammation in Parkinson’s Disease via Regulating miR-7/NLRP3 Pathway. J. Neurosci. 2018, 388, 118–127. [Google Scholar] [CrossRef]
- Dong, J.; Fu, T.; Yang, Y.; Mu, Z.; Li, X. Long Noncoding RNA SNHG1 Promotes Lipopolysaccharide-Induced Activation and Inflammation in Microglia via Targeting miR-181b. Neuroimmunomodulation 2021, 28, 255–265. [Google Scholar] [CrossRef]
- Sabaie, H.; Salkhordeh, Z.; Asadi, M.R.; Ghafouri-Fard, S.; Amirinejad, N.; Behzadi, M.A.; Hussen, B.M.; Taheri, M.; Rezazadeh, M. Long Non-Coding RNA- Associated Competing Endogenous RNA Axes in T-Cells in Multiple Sclerosis. Front. Immunol. 2021, 12, 770679. [Google Scholar] [CrossRef] [PubMed]
- Mathy, N.W.; Chen, X.-M. Long non-coding RNAs (lncRNAs) and their transcriptional control of inflammatory responses. J. Biol. Chem. 2017, 292, 12375–12382. [Google Scholar] [CrossRef] [PubMed]
- Hu, G.; Gong, A.Y.; Wang, Y.; Ma, S.; Chen, X.; Chen, J.; Su, C.-J.; Shibata, A.; Strauss-Soukup, J.K.; Drescher, K.M.; et al. LincRNA-Cox2 Promotes Late Inflammatory Gene Transcription in Macrophages through Modulating SWI/SNF-Mediated Chromatin Remodeling. J. Immunol. 2016, 196, 2799–2808. [Google Scholar] [CrossRef] [PubMed]
- Xue, Z.; Zhang, Z.; Liu, H.; Li, W.; Guo, X.; Zhang, Z.; Liu, Y.; Jia, L.; Li, Y.; Ren, Y.; et al. lincRNA-Cox2 regulates NLRP3 inflammasome and autophagy mediated neuroinflammation. Cell Death Differ. 2019, 26, 130–145. [Google Scholar] [CrossRef]
- Polman, C.H.; Reingold, S.C.; Banwell, B.; Clanet, M.; Cohen, J.A.; Filippi, M.; Fujihara, K.; Havrdova, E.; Hutchinson, M.; Kappos, L.; et al. Diagnostic criteria for multiple sclerosis: 2010 revisions to the McDonald criteria. Ann. Neurol. 2011, 69, 292–302. [Google Scholar] [CrossRef]
- Kurtzke, J.F. Rating neurologic impairment in multiple sclerosis: An expanded disability status scale (EDSS). Neurology 1983, 33, 1444–1452. [Google Scholar] [CrossRef]
- Belman, A.L.; Krupp, L.B.; Olsen, C.S.; Rose, J.W.; Aaen, G.; Benson, L.; Chitnis, T.; Gorman, M.; Graves, J.; Harris, Y.; et al. Characteristics of Children and Adolescents with Multiple Sclerosis. Pediatrics 2016, 138, e20160120. [Google Scholar] [CrossRef]
- Ewida, H.A.; Zayed, R.K.; Darwish, H.A.; Shaheen, A.A. Circulating lncRNAs HIF1A-AS2 and LINLK-A: Role and relation to hypoxia-inducible factor-1α in cerebral stroke patients. Mol. Neurobiol. 2021, 58, 4564–4574. [Google Scholar] [CrossRef]
- Zayed, A.A.; Seleem, M.M.; Darwish, H.A.; Shaheen, A.A. Role of long noncoding RNAs; BDNF-AS and 17A and their relation to GABAergic dysfunction in Egyptian epileptic patients. Metab. Brain Dis. 2023, 1–12. [Google Scholar] [CrossRef]
- Fleige, S.; Pfaffl, M.W. RNA integrity and the effect on the real-time qRT-PCR performance. Mol. Aspects Med. 2006, 27, 126–139. [Google Scholar] [CrossRef]
- Hauser, S.L.; Cree, B.A. Treatment of Multiple Sclerosis: A Review. Am. J. Med. 2020, 133, 1380–1390.e2. [Google Scholar] [CrossRef] [PubMed]
- Hur, K.; Kim, S.-H.; Kim, J.-M. Potential Implications of Long Noncoding RNAs in Autoimmune Diseases. Immune Netw. 2019, 19, e4. [Google Scholar] [CrossRef] [PubMed]
- Kong, K.-F.; Fu, G.; Zhang, Y.; Yokosuka, T.; Casas, J.; Canonigo-Balancio, A.J.; Bécart, S.; Kim, G.; Yates, J.R.; Kronenberg, M.; et al. Protein kinase C-η controls CTLA-4-mediated regulatory T cell function. Nat. Immunol. 2014, 15, 465–472. [Google Scholar] [CrossRef]
- Kleinewietfeld, M.; Hafler, D.A. Regulatory T cells in autoimmune neuroinflammation. Immunol. Rev. 2014, 259, 231–244. [Google Scholar] [CrossRef]
- Alatab, S.; Maghbooli, Z.; Hossein-Nezhad, A.; Khosrofar, M.; Mokhtari, F. Cytokine profile, Foxp3 and nuclear factor-kB ligand levels in multiple sclerosis subtypes. Minerva Med. 2011, 102, 461–468. [Google Scholar] [PubMed]
- Frisullo, G.; Nociti, V.; Iorio, R.; Plantone, D.; Patanella, A.K.; Tonali, P.A.; Batocchi, A.P. CD8(+)Foxp3(+) T cells in peripheral blood of relapsing-remitting multiple sclerosis patients. Hum. Immunol. 2010, 71, 437–441. [Google Scholar] [CrossRef]
- Frisullo, G.; Nociti, V.; Iorio, R.; Patanella, A.K.; Caggiula, M.; Marti, A.; Sancricca, C.; Angelucci, F.; Mirabella, M.; Tonnali, P.A.; et al. Regulatory T cells fail to suppress CD4T+-bet+ T cells in relapsing multiple sclerosis patients. Immunology 2009, 127, 418–428. [Google Scholar] [CrossRef]
- Ghadiri, N.; Emamnia, N.; Ganjalikhani-Hakemi, M.; Ghaedi, K.; Etemadifar, M.; Salehi, M.; Shirzad, H.; Nasr-Esfahani, M.H. Analysis of the expression of mir-34a, mir-199a, mir-30c and mir-19a in peripheral blood CD4+T lymphocytes of relapsing-remitting multiple sclerosis patients. Gene 2018, 659, 109–117. [Google Scholar] [CrossRef]
- Lee, P.W.; Severin, M.E.; Lovett-Racke, A.E. TGF-β regulation of encephalitogenic and regulatory T cells in multiple sclerosis. Eur. J. Immunol. 2017, 47, 446–453. [Google Scholar] [CrossRef]
- Rollnik, J.D.; Sindern, E.; Schweppe, C.; Malin, J.P. Biologically active TGF-beta 1 is increased in cerebrospinal fluid while it is reduced in serum in multiple sclerosis patients. Acta Neurol. Scand. 1997, 96, 101–105. [Google Scholar] [CrossRef]
- Balasa, R.; Maier, S.; Voidazan, S.; Hutanu, A.; Bajko, Z.; Motataianu, A.; Brandusa, T.; Tiu, C. Assessment of Interleukin-17A, Interleukin-10 and Transforming Growth Factor-Beta1 Serum Titers in Relapsing Remitting Multiple Sclerosis Patients Treated with Avonex, Possible Biomarkers for Treatment Response. CNS Neurol. Disord. Drug Targets 2017, 16, 93–101. [Google Scholar] [CrossRef] [PubMed]
- Dolati, S.; Babaloo, Z.; Ayromlou, H.; Ahmadi, M.; Rikhtegar, R.; Rostamzadeh, D.; Roshangar, L.; Nouri, M.; Mehdizadeh, A.; Younesi, V.; et al. Nanocurcumin improves regulatory T-cell frequency and function in patients with multiple sclerosis. J. Neuroimmunol. 2019, 327, 15–21. [Google Scholar] [CrossRef] [PubMed]
- Losy, J.; Michałowska-Wender, G. In vivo effect of interferon-beta 1a on interleukin-12 and TGF-beta(1) cytokines in patients with relapsing-remitting multiple sclerosis. Acta Neurol. Scand. 2002, 106, 44–46. [Google Scholar] [CrossRef] [PubMed]
- Ganji, A.; Monfared, M.E.; Shapoori, S.; Nourbakhsh, P.; Ghazavi, A.; Ghasami, K.; Mosayebi, G. Effects of interferon and glatiramer acetate on cytokine patterns in multiple sclerosis patients. Cytokine 2020, 126, 154911. [Google Scholar] [CrossRef]
- Nicoletti, F.; Di Marco, R.; Patti, F.; Reggio, E.; Zaccone, P.; Stivala, F.; Meroni, P.L.; Reggio, A. Blood levels of transforming growth factor-beta 1 (TGF-beta1) are elevated in both relapsing remitting and chronic progressive multiple sclerosis (MS) patients and are further augmented by treatment with interferon-beta 1b (IFN-beta1b). Clin. Exp. Immunol. 1998, 113, 96–99. [Google Scholar] [CrossRef]
- Hollifield, R.D.; Harbige, L.S.; Pham-Dinh, D.; Sharief, M.K. Evidence for cytokine dysregulation in multiple sclerosis: Peripheral blood mononuclear cell production of pro-inflammatory and anti-inflammatory cytokines during relapse and remission. J. Autoimmun. 2003, 36, 133–141. [Google Scholar] [CrossRef]
- Derfuss, T.; Ontaneda, D.; Nicholas, J.; Meng, X.; Hawker, K. Relapse rates in patients with multiple sclerosis treated with fingolimod: Subgroup analyses of pooled data from three phase 3 trials. Mult. Scler. Relat. Disord. 2016, 8, 124–130. [Google Scholar] [CrossRef]
- Tripathi, S.; Shree, B.; Mohapatra, S.; Swati; Basu, A.; Sharma, V. The Expanding Regulatory Mechanisms and Cellular Functions of Long Non-coding RNAs (lncRNAs) in Neuroinflammation. Mol. Neurobiol. 2021, 58, 2916–2939. [Google Scholar] [CrossRef]
- Wang, H.; Lu, B.; Chen, J. Knockdown of lncRNA SNHG1 attenuated Aβ(25-35)-inudced neuronal injury via regulating KREMEN1 by acting as a ceRNA of miR-137 in neuronal cells. Biochem. Biophys. Res. Commun. 2019, 518, 438–444. [Google Scholar] [CrossRef]
- Carpenter, S.; Aiello, D.; Atianand, M.K.; Ricci, E.P.; Gandhi, P.; Hall, L.L.; Byron, M.; Monks, B.; Henry-Bezy, M.; Lawrence, J.B.; et al. A long noncoding RNA mediates both activation and repression of immune response genes. Science 2013, 341, 789–792. [Google Scholar] [CrossRef]
- Kawana, N.; Yamamoto, Y.; Ishida, T.; Saito, Y.; Konno, H.; Arima, K.; Satoh, J.-I. Reactive astrocytes and perivascular macrophages express NLRP 3 inflammasome in active demyelinating lesions of multiple sclerosis and necrotic lesions of neuromyelitis optica and cerebral infarction. Clin. Exp. Immunol. 2013, 4, 296–304. [Google Scholar]
- Imani, D.; Azimi, A.; Salehi, Z.; Rezaei, N.; Emamnejad, R.; Sadr, M.; Izad, M. Association of nod-like receptor protein-3 single nucleotide gene polymorphisms and expression with the susceptibility to relapsing-remitting multiple sclerosis. Int. J. Immunogenet. 2018, 45, 329–336. [Google Scholar] [CrossRef] [PubMed]
- Park, S.-H.; Ham, S.; Lee, A.; Möller, A.; Kim, T.S. NLRP3 negatively regulates Treg differentiation through Kpna2-mediated nuclear translocation. J. Biol. Chem. 2019, 294, 17951–17961. [Google Scholar] [CrossRef] [PubMed]





{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Groups | Healthy Controls (n = 30) | RRMS Patients | p-Value | ||
|---|---|---|---|---|---|
| Parameters | Relapse (n = 35) | Remission (n = 35) | |||
| Age (Years) | |||||
| Range | 22–45 | 20–50 | 19–48 | 0.106 | |
| Median | 30 | 35 | 32 | ||
| Gender: | |||||
| -Male n (%) | 10 (33.3%) | 9 (25.7%) | 11 (31.4%) | 0.779 | |
| -Female n (%) | 20 (66.7%) | 26 (74.3%) | 24 (68.6%) | ||
| MS Family History | |||||
| -Negative | \\ | 33 (94.3%) | 33 (94.3%) | 0.693 | |
| -Positive | \\ | 2 (5.7%) | 2 (5.7%) | ||
| Disease Duration (years) | |||||
| -Range | \\ | 2–10 | 1–10 | 0.282 | |
| -Median | \\ | 5 | 5 | ||
| Relapses in Last 2 Years | |||||
| Range | \\ | 2–6 | 1–5 | 0.658 | |
| Median | \\ | 3 | 3 | ||
| ARR in Last 2 Years | |||||
| -Range | \\ | 1–3 | 0.5–2.5 | 0.672 | |
| -Median | \\ | 1.5 | 1.5 | ||
| EDSS Score | |||||
| -Range | \\ | 1.5–6 | 1–6 | 0.04 | |
| -Median | \\ | 3 | 2 | ||
| Treatment: n (%) | |||||
| -Interferon-β | \\ | 25 (71.4%) | 23 (65.7%) | 0.071 | |
| -Fingolimod | \\ | 10 (28.6%) | 12 (34.3%) | ||
| Lnc-EGFR | SNHG1 | lincRNA-Cox2 | |
|---|---|---|---|
| Age | −0.198 * | 0.262 ** | N.S. |
| Disease Duration | N.S. | N.S. | N.S. |
| Relapses in Last 2 Years | −0.331 ** | N.S. | 0.298 * |
| ARR | −0.321 ** | 0.245 * | 0.283 * |
| EDSS Score | −0.347 ** | N.S. | 0.322 ** |
| Lnc-EGFR | ----- | −0.705 ** | −0.648 ** |
| FOXP3 | 0.721 ** | −0.563 ** | −0.606 ** |
| TGF-β1 | 0.733 ** | −0.777 ** | −0.742 ** |
| SNHG1 | −0.705 ** | ----- | 0.706 ** |
| lincRNA-Cox2 | −0.648 ** | 0.706 ** | ----- |
| NLRP3 | −0.631 ** | 0.708 ** | 0.824 ** |
| ASC | −0.584 ** | 0.649 ** | 0.594 ** |
| Caspase-1 | −0.581 ** | 0.689 ** | 0.776 ** |
| IL-1β | −0.717 ** | 0.692 ** | 0.768 ** |
| I-Between RRMS Patients and Healthy Controls | Healthy Controls (n = 30) | RRMS Patients (n = 70) | Chi-Square X2 | p-Value | Sensitivity (%) | Specificity (%) | PPV (%) | NPV (%) | Accuracy (%) |
|---|---|---|---|---|---|---|---|---|---|
| Lnc-EGFR | |||||||||
| -No. of +ve cases (≥0.78) | 26 (86.7%) | 6 (8.6%) | |||||||
| -No. of −ve cases (<0.78) | 4 (13.3%) | 64 (91.4%) | 58.859 | 0.000 | 86.7 | 91.4 | 81.25 | 94.11 | 89.05 |
| FOXP3 | |||||||||
| -No. of +ve cases (≥0.83) | 26 (86.7%) | 5 (7.1%) | |||||||
| -No. of −ve cases (<0.83) | 4 (13.3%) | 65 (92.9%) | 62.087 | 0.000 | 86.7 | 92.9 | 83.87 | 94.2 | 89.8 |
| TGF-β1 | |||||||||
| -No. of +ve cases (≥82.45) | 29 (96.7%) | 3 (4.3%) | |||||||
| -No. of −ve cases (<82.45) | 1 (3.3%) | 67 (95.7%) | 82.362 | 0.000 | 96.7 | 95.7 | 90.6 | 98.5 | 96.2 |
| II-Between RRMS patients during relapse and in remission | Relapse (n = 35) | Remission (n = 35) | Chi-Square X2 | p-Value | Sensitivity (%) | Specificity (%) | PPV (%) | NPV (%) | Accuracy (%) |
| Lnc-EGFR | |||||||||
| -No. of +ve cases (≥0.44) | 7 (20%) | 33 (94.3%) | |||||||
| -No. of −ve cases (<0.44) | 28 (80%) | 2 (5.7%) | 39.433 | 0.000 | 94.3 | 80 | 82.5 | 93.33 | 87.15 |
| FOXP3 | |||||||||
| -No. of +ve cases (≥0.53) | 6 (17.1%) | 25 (71.4%) | |||||||
| -No. of −ve cases (<0.53) | 29 (82.9%) | 10 (28.6%) | 20.902 | 0.000 | 71.4 | 82.9 | 80.6 | 74.35 | 77.15 |
| TGF-β1 | |||||||||
| -No. of +ve cases (≥29) | 2 (5.7%) | 32 (91.4%) | |||||||
| -No. of −ve cases (<29) | 33 (94.3%) | 3 (8.6%) | 51.471 | 0.000 | 91.4 | 94.3 | 94.11 | 91.66 | 92.85 |
| SNHG1 | |||||||||
| -No. of +ve cases (≥214.5) | 26 (74.3%) | 10 (28.6%) | |||||||
| -No. of −ve cases (<214.5) | 9 (25.7%) | 25 (71.4%) | 14.641 | 0.000 | 74.3 | 71.4 | 72.22 | 73.53 | 72.85 |
| LincRNA-Cox2 | |||||||||
| -No. of +ve cases (≥261.3) | 25 (71.4) | 2 (5.7%) | |||||||
| -No. of −ve cases (<261.3) | 10 (28.6%) | 33 (94.3%) | 31.895 | 0.000 | 71.4 | 94.3 | 92.59 | 76.74 | 82.85 |
| NLRP3 | |||||||||
| -No. of +ve cases (≥207.3) | 30 (85.7%) | 7 (20%) | |||||||
| -No. of −ve cases (<207.3) | 5 (14.3%) | 28 (80%) | 30.328 | 0.000 | 85.7 | 80 | 81.08 | 84.85 | 82.85 |
| ASC | |||||||||
| -No. of +ve cases (≥73) | 27 (77.1%) | 9 (25.7%) | |||||||
| -No. of −ve cases (<73) | 8 (22.9%) | 26 (74.3%) | 18.529 | 0.000 | 77.1 | 74.3 | 75 | 76.47 | 75.7 |
| Caspase-1 | |||||||||
| -No. of +ve cases (≥98.4) | 25 (71.4%) | 4 (11.4%) | |||||||
| -No. of −ve cases (<98.4) | 10 (28.6%) | 31 (88.6%) | 25.963 | 0.000 | 71.4 | 88.6 | 86.2 | 75.6 | 80 |
| IL-1β | |||||||||
| -No. of +ve cases (≥18.4) | 30 (85.7%) | 2 (5.7%) | |||||||
| -No. of −ve cases (<18.4) | 5 (14.3%) | 33 (94.3%) | 45.132 | 0.000 | 85.7 | 94.3 | 93.75 | 86.84 | 90 |
| Parameters | B | S.E. | p-Value | Odds Ratio | 95% CI |
|---|---|---|---|---|---|
| Biomarkers for diagnosis of RRMS | |||||
| Lnc-EGFR | −11.64 | 2.46 | 0.000 | 0.000 | 0.000–0.001 |
| FOXP3 | −6.96 | 1.33 | 0.000 | 0.001 | 0.000–0.013 |
| TGF-β1 | −0.163 | 0.064 | 0.012 | 0.850 | 0.749–0.964 |
| Biomarkers that predict prognosis of relapse in RRMS patients | |||||
| Lnc-EGFR | −9.491 | 2.18 | 0.000 | 0.000 | 0.000–0.005 |
| FOXP3 | −6.62 | 1.703 | 0.000 | 0.001 | 0.000–0.038 |
| TGF-β1 | −0.529 | 0.143 | 0.000 | 0.589 | 0.445–0.780 |
| SNHG1 | 0.013 | 0.003 | 0.000 | 1.014 | 1.007–1.020 |
| LincRNA-Cox2 | 0.018 | 0.004 | 0.000 | 1.018 | 1.010–1.026 |
| NLRP3 | 0.017 | 0.004 | 0.000 | 1.017 | 1.010–1.024 |
| ASC | 0.021 | 0.006 | 0.001 | 1.022 | 1.009–1.034 |
| Caspase-1 | 0.038 | 0.009 | 0.000 | 1.038 | 1.020–1.057 |
| IL-1β | 0.480 | 0.122 | 0.000 | 1.616 | 1.272–2.053 |
| EDSS | 1.742 | 0.825 | 0.035 | 5.712 | 1.135–28.75 |
| Relapses in Last 2 Years | 0.737 | 0.248 | 0.003 | 2.089 | 1.284–3.409 |
| ARR | 1.474 | 0.497 | 0.003 | 4.365 | 1.65–11.56 |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Attia, M.S.; Ewida, H.A.; Abdel Hafez, M.A.; El-Maraghy, S.A.; El-Sawalhi, M.M. Altered Lnc-EGFR, SNHG1, and LincRNA-Cox2 Profiles in Patients with Relapsing-Remitting Multiple Sclerosis: Impact on Disease Activity and Progression. Diagnostics 2023, 13, 1448. https://doi.org/10.3390/diagnostics13081448
Attia MS, Ewida HA, Abdel Hafez MA, El-Maraghy SA, El-Sawalhi MM. Altered Lnc-EGFR, SNHG1, and LincRNA-Cox2 Profiles in Patients with Relapsing-Remitting Multiple Sclerosis: Impact on Disease Activity and Progression. Diagnostics. 2023; 13(8):1448. https://doi.org/10.3390/diagnostics13081448
Chicago/Turabian StyleAttia, Mohamed S., Heba A. Ewida, Mohamed Aly Abdel Hafez, Shohda A. El-Maraghy, and Maha M. El-Sawalhi. 2023. "Altered Lnc-EGFR, SNHG1, and LincRNA-Cox2 Profiles in Patients with Relapsing-Remitting Multiple Sclerosis: Impact on Disease Activity and Progression" Diagnostics 13, no. 8: 1448. https://doi.org/10.3390/diagnostics13081448
APA StyleAttia, M. S., Ewida, H. A., Abdel Hafez, M. A., El-Maraghy, S. A., & El-Sawalhi, M. M. (2023). Altered Lnc-EGFR, SNHG1, and LincRNA-Cox2 Profiles in Patients with Relapsing-Remitting Multiple Sclerosis: Impact on Disease Activity and Progression. Diagnostics, 13(8), 1448. https://doi.org/10.3390/diagnostics13081448