

Targeted Sequencing of Germline Breast Cancer Susceptibility Genes for Discovering Pathogenic/Likely Pathogenic Variants in the Jakarta Population

 ,
,
Abstract
1. Introduction
2. Materials and Methods
2.1. Patients
2.2. DNA Extraction, Library Preparation, and Sequencing
2.3. Bioinformatics Analysis
2.4. Clinical Correlation and Statistical Analysis
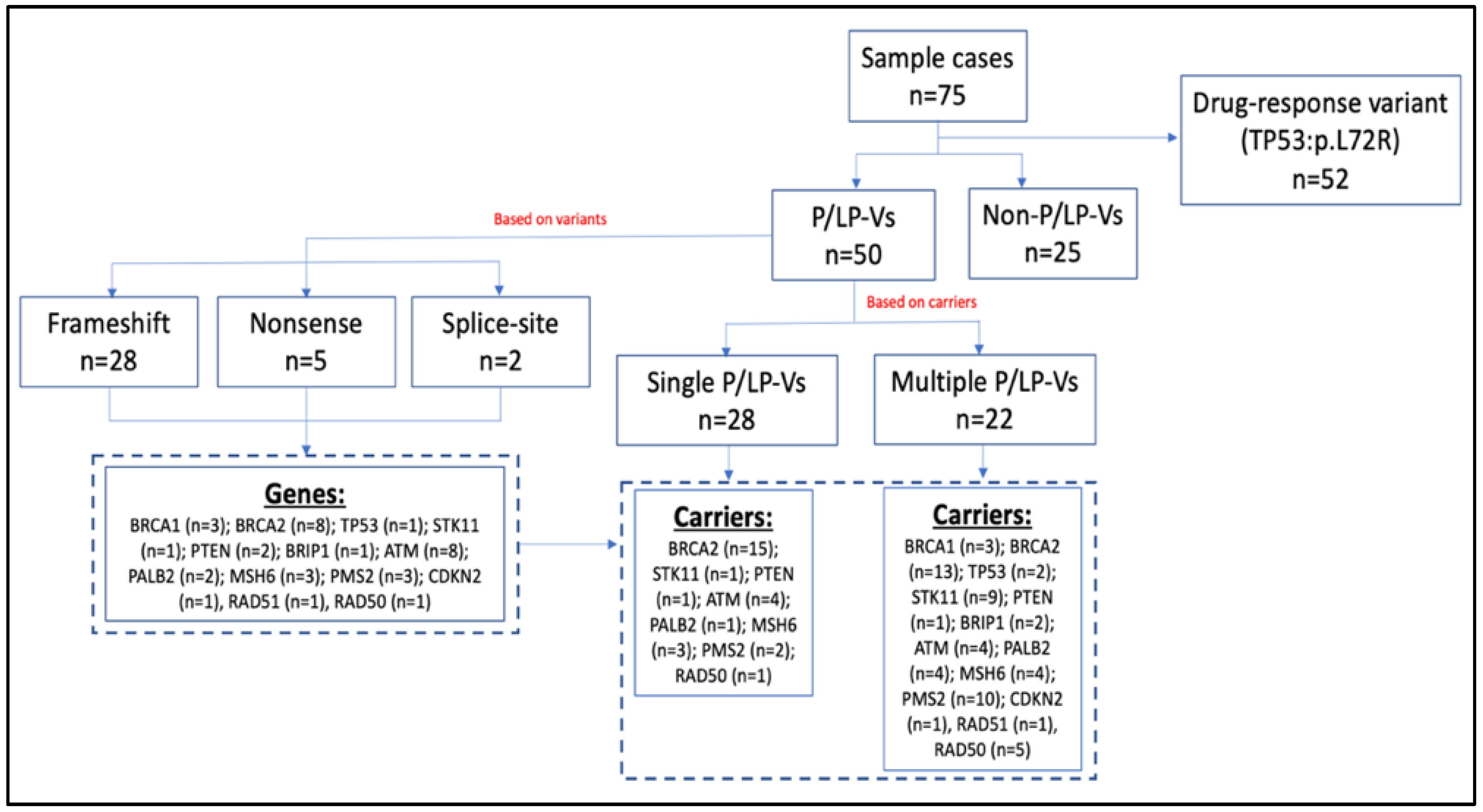
3. Results
3.1. Patients’ Characteristics
3.2. Characteristics of the Germline Variants
3.3. Prediction of Drug Response Alteration Associated with Gene Variants
3.4. Correlation of Variants with Tumor Characteristics
4. Discussion
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Observatory, G.C. Breast Fact Sheet. 2020, Volume 419. Available online: https://gco.iarc.fr/today/data/factsheets/cancers/20-Breast-fact-sheet.pdf (accessed on 4 March 2022).
- Chan, W.; Lee, M.; Yeo, Z.X.; Ying, D.; Grimaldi, K.A.; Pickering, C.; Yang, M.M.S.; Sundaram, S.K.; Tzang, L.C.H. Development and validation of next generation sequencing based 35-gene hereditary cancer panel. Hered. Cancer Clin. Pract. 2020, 18, 9. [Google Scholar] [CrossRef] [PubMed]
- Desmond, A.; Kurian, A.W.; Gabree, M.; Mills, M.A.; Anderson, M.J.; Kobayashi, Y.; Horick, N.; Yang, S.; Shannon, K.M.; Tung, N.; et al. Clinical Actionability of Multigene Panel Testing for Hereditary Breast and Ovarian Cancer Risk Assessment. JAMA Oncol. 2015, 1, 943–951. [Google Scholar] [CrossRef]
- Inagaki-Kawata, Y.; Yoshida, K.; Kawaguchi-Sakita, N.; Kawashima, M.; Nishimura, T.; Senda, N.; Shiozawa, Y.; Takeuchi, Y.; Inoue, Y.; Sato-Otsubo, A.; et al. Genetic and clinical landscape of breast cancers with germline BRCA1/2 variants. Commun Biol. 2020, 3, 578. [Google Scholar] [CrossRef] [PubMed]
- Wang, Y.A.; Jian, J.-W.; Hung, C.-F.; Peng, H.-P.; Yang, C.-F.; Cheng, H.-C.S.; Yang, A.-S. Germline breast cancer susceptibility gene mutations and breast cancer outcomes. BMC Cancer 2018, 18, 315. [Google Scholar] [CrossRef] [PubMed]
- Easton, D.F.; Pharoah, P.D.P.; Antoniou, A.C.; Tischkowitz, M.; Tavtigian, S.V.; Nathanson, K.L.; Devilee, P.; Meindl, A.; Couch, F.J.; Southey, M.; et al. Gene-Panel Se-quencing and the Prediction of Breast-Cancer Risk. N. Engl. J. Med. 2015, 372, 2243–2257. [Google Scholar] [CrossRef] [PubMed]
- Couch, F.J.; Shimelis, H.; Hu, C.; Hart, S.N.; Polley, E.C.; Na, J.; Hallberg, E.; Moore, R.; Thomas, A.; Lilyquist, J.; et al. Associations Between Cancer Predisposition Testing Panel Genes and Breast Cancer. JAMA Oncol. 2017, 3, 1190–1196. [Google Scholar] [CrossRef]
- Sato, K.; Koyasu, M.; Nomura, S.; Sato, Y.; Kita, M.; Ashihara, Y.; Adachi, Y.; Ohno, S.; Iwase, T.; Kitagawa, D.; et al. Mutation status of RAD 51C, PALB 2 and BRIP 1 in 100 Japanese familial breast cancer cases without BRCA 1 and BRCA 2 mutations. Cancer Sci. 2017, 108, 2287–2294. [Google Scholar] [CrossRef]
- Ripperger, T.; Gadzicki, D.; Meindl, A.; Schlegelberger, B. Breast cancer susceptibility: Current knowledge and implications for genetic counselling. Eur. J. Hum. Genet. 2009, 17, 722–731. [Google Scholar] [CrossRef]
- Dong, H.; Chandratre, K.; Qin, Y.; Zhang, J.; Tian, X.; Rong, C.; Wang, N.; Guo, M.; Zhao, G.; Wang, S.M. Prevalence of BRCA1/BRCA2 pathogenic variation in Chinese Han population. J. Med. Genet. 2020, 58, 565–569. [Google Scholar] [CrossRef]
- Kharel, S.; Shrestha, S.; Yadav, S.; Shakya, P.; Baidya, S.; Hirachan, S. BRCA1/BRCA2 mutation spectrum analysis in South Asia: A systematic review. J. Int. Med. Res. 2022, 50, 3000605211070757. [Google Scholar] [CrossRef]
- Kim, H.; Choi, D.H. Distribution of BRCA1 and BRCA2 mutations in asian patients with breast cancer. J. Breast Cancer 2013, 16, 357–365. [Google Scholar] [CrossRef]
- Edge, S.B.; Byrd, D.R.; Compton, C.C.; Fritz, A.G.; Greene, F.L.; Trotti, A. Breast. In AJCC Cancer Staging Handbook, 7th ed; Springer: New York, NY, USA, 2010; pp. 345–376. Available online: http://link.springer.com/10.1007/978-0-387-88443-1_29 (accessed on 4 March 2022).
- Panigoro, S.S.; Siswiandari, K.M.; Paramita, R.I.; Fadilah, F.; Erlina, L. Targeted genome sequencing data of young women breast cancer patients in Cipto Mangunkusumo national hospital, Jakarta. Data Brief 2020, 32, 106138. [Google Scholar] [CrossRef]
- Andrews, S. FastQC: A Quality Control Tool for High Throughput Sequence Data; Babraham Institute: Cambridge, UK, 2010; Available online: http://www.bioinformatics.babraham.ac.uk/projects/fastqc/ (accessed on 25 January 2022).
- Bolger, A.M.; Lohse, M.; Usadel, B. Trimmomatic: A flexible trimmer for Illumina sequence data. Bioinformatics 2014, 30, 2114–2120. [Google Scholar] [CrossRef]
- Li, H. Aligning sequence reads, clone sequences and assembly contigs with BWA-MEM. arXiv 2013, arXiv:1303.3997. [Google Scholar]
- Poplin, R.; Ruano-Rubio, V.; DePristo, M.A.; Fennell, T.J.; Carneiro, M.O.; Van der Auwera, G.A.; Kling, D.E.; Gauthier, L.D.; Levy-Moonshine, A.; Roazen, D.; et al. Scaling accurate genetic variant discovery to tens of thousands of samples. bioRxiv 2017. preprint. [Google Scholar]
- Cingolani, P.; Platts, A.; Wang, L.L.; Coon, M.; Nguyen, T.; Wang, L.; Land, S.J.; Lu, X.; Ruden, D.M. A program for annotating and predicting the effects of single nucleotide polymorphisms, SnpEff: SNPs in the genome of Drosophila melanogaster strain w1118; iso-2; iso-3. Fly 2012, 6, 80–92. [Google Scholar] [CrossRef] [PubMed]
- Kopanos, C.; Tsiolkas, V.; Kouris, A.; Chapple, C.E.; Aguilera, M.A.; Meyer, R.; Massouras, A. VarSome: The human genomic variant search engine. Bioinformatics 2019, 35, 1978–1980. [Google Scholar] [CrossRef] [PubMed]
- Gao, J.; Aksoy, B.A.; Dogrusoz, U.; Dresdner, G.; Gross, B.; Sumer, S.O.; Sun, Y.; Jacobsen, A.; Sinha, R.; Larsson, E.; et al. Integrative Analysis of Complex Cancer Ge-nomics and Clinical Profiles Using the cBioPortal. Sci Signal. 2013, 6, pl1. [Google Scholar] [CrossRef]
- Cerami, E.; Gao, J.; Dogrusoz, U.; Gross, B.E.; Sumer, S.O.; Aksoy, B.A.; Jacobsen, A.; Byrne, C.J.; Heuer, M.L.; Larsson, E.; et al. The cBio Cancer Genomics Portal: An Open Platform for Exploring Multidimensional Cancer Genomics Data: Figure 1. Cancer Discov. 2012, 2, 401–404. [Google Scholar] [CrossRef]
- Finn, R.S.; Dering, J.; Conklin, D.; Kalous, O.; Cohen, D.J.; Desai, A.J.; Ginther, C.; Atefi, M.; Chen, I.; Fowst, C.; et al. PD 0332991, a selective cyclin D kinase 4/6 inhibitor, preferentially inhibits proliferation of luminal estrogen receptor-positive human breast cancer cell lines in vitro. Breast Cancer Res. 2009, 11, R77. [Google Scholar] [CrossRef]
- Van Arsdale, T.; Boshoff, C.; Arndt, K.T.; Abraham, R.T. Molecular pathways: Targeting the cyclin D-CDK4/6 axis for cancer treatment. Clin. Cancer Res. 2015, 21, 2905–2910. [Google Scholar]
- Mateo, J.; Ganji, G.; Lemech, C.; Burris, H.A.; Han, S.-W.; Swales, K.; Decordova, S.; Deyoung, M.P.; Smith, D.A.; Kalyana-Sundaram, S.; et al. A First-Time-in-Human Study of GSK2636771, a Phosphoinositide 3 Kinase Beta-Selective Inhibitor, in Patients with Advanced Solid Tumors. Clin. Cancer Res. 2017, 23, 5981–5992. [Google Scholar] [CrossRef] [PubMed]
- Baretta, Z.; Mocellin, S.; Goldin, E.; Olopade, O.I.; Huo, D. Effect of BRCA germline mutations on breast cancer prognosis. Medicine 2016, 95, e4975. [Google Scholar] [CrossRef] [PubMed]
- Tung, N.; Battelli, C.; Allen, B.; Kaldate, R.; Bhatnagar, S.; Bowles, K.; Timms, K.; Garber, J.E.; Herold, C.; Ellisen, L.; et al. Frequency of mutations in individuals with breast cancer referred for BRCA 1 and BRCA 2 testing using next-generation sequencing with a 25-gene panel. Cancer 2015, 121, 25–33. [Google Scholar] [CrossRef] [PubMed]
- Shin, H.C.; Lee, H.B.; Yoo, T.K.; Lee, E.S.; Kim, R.N.; Park, B.; Yoon, K.A.; Park, C.; Lee, E.S.; Moon, H.G.; et al. Detection of Germline Mutations in Breast Cancer Pa-tients with Clinical Features of Hereditary Cancer Syndrome Using a Multi-Gene Panel Test. Cancer Res. Treat. 2020, 52, 697–713. [Google Scholar] [CrossRef] [PubMed]
- Yao, K.A.; Clifford, J.; Li, S.; LaDuca, H.; Hulick, P.; Gutierrez, S.; Black, M.H. Prevalence of Germline Pathogenic and Likely Pathogenic Variants in Patients with Second Breast Cancers. JNCI Cancer Spectr. 2020, 4, pkaa094. [Google Scholar] [CrossRef] [PubMed]
- Valachis, A.; Nearchou, A.D.; Lind, P. Surgical management of breast cancer in BRCA-mutation carriers: A systematic review and meta-analysis. Breast Cancer Res. Treat. 2014, 144, 443–455. [Google Scholar] [CrossRef]
- Jiang, Y.; Tian, T.; Yu, C.; Zhou, W.; Yang, J.; Wang, Y.; Wen, Y.; Chen, J.; Dai, J.; Jin, G.; et al. Identification of Recurrent Variants in BRCA1 and BRCA2 across Multiple Cancers in the Chinese Population. BioMed Res. Int. 2020, 2020, 6739823. [Google Scholar] [CrossRef] [PubMed]
- Jouali, F.; Laarabi, F.Z.; Marchoudi, N.; Ratbi, I.; Elalaoui, S.C.; Rhaissi, H.; Fekkak, J.; Sefiani, A. First application of next-generation se-quencing in Moroccan breast/ovarian cancer families and report of a novel frameshift mutation of the BRCA1 gene. Oncol Lett. 2016, 12, 1192–1196. [Google Scholar] [CrossRef][Green Version]
- Bakkach, J.; Mansouri, M.; Derkaoui, T.; Loudiyi, A.; El Fahime, E.; Barakat, A.; Nourouti, N.G.; De Villarreal, J.M.; Bringas, C.C.; Mechita, M.B. Contribution of BRCA1 and BRCA2 germline mutations to early onset breast cancer: A series from north of Morocco. BMC Cancer 2020, 20, 859. [Google Scholar] [CrossRef]
- Lolas Hamameh, S.; Renbaum, P.; Kamal, L.; Dweik, D.; Salahat, M.; Jaraysa, T.; Abu Rayyan, A.; Casadei, S.; Mandell, J.B.; Gulsuner, S.; et al. Genomic analysis of inherited breast cancer among Palestinian women: Genetic heterogeneity and a founder mutation in TP53. Int. J. Cancer 2017, 141, 750–756. [Google Scholar] [CrossRef] [PubMed]
- Akcay, I.M.; Celik, E.; Agaoglu, N.B.; Alkurt, G.; Msc, T.K.A.; Yildiz, J.; Enc, F.; Kir, G.; Canbek, S.; Kilic, A.; et al. Germline pathogenic variant spectrum in 25 cancer susceptibility genes in Turkish breast and colorectal cancer patients and elderly controls. Int. J. Cancer 2021, 148, 285–295. [Google Scholar] [CrossRef] [PubMed]
- Incorvaia, L.; Fanale, D.; Bono, M.; Calò, V.; Fiorino, A.; Brando, C.; Corsini, L.R.; Cutaia, S.; Cancelliere, D.; Pivetti, A.; et al. BRCA1/2 pathogenic variants in triple-negative versus luminal-like breast cancers: Genotype–phenotype correlation in a cohort of 531 patients. Ther. Adv. Med. Oncol. 2020, 12, 175883592097532. Available online: http://www.ncbi.nlm.nih.gov/pubmed/28086035 (accessed on 15 March 2022). [CrossRef]
- Larsen, M.J.; Kruse, T.A.; Tan, Q.; Lænkholm, A.V.; Bak, M.; Lykkesfeldt, A.E.; Sørensen, K.P.; Hansen, T.V.O.; Ejlertsen, B.; Gerdes, A.M.; et al. Classifications within Molecular Sub-types Enables Identification of BRCA1/BRCA2 Mutation Carriers by RNA Tumor Profiling. PLoS ONE 2013, 8, e64268. [Google Scholar]
- Shah, P.D.; Patil, S.; Dickler, M.N.; Offit, K.; Hudis, C.A.; Robson, M.E. Twenty-one-gene recurrence score assay in BRCA-associated versus sporadic breast cancers: Differences based on germline mutation status. Cancer 2016, 122, 1178–1184. [Google Scholar] [CrossRef] [PubMed]
- Litton, J.K.; Rugo, H.S.; Ettl, J.; Hurvitz, S.A.; Gonçalves, A.; Lee, K.-H.; Fehrenbacher, L.; Yerushalmi, R.; Mina, L.A.; Martin, M.; et al. Talazoparib in Patients with Advanced Breast Cancer and a Germline BRCA Mutation. N. Engl. J. Med. 2018, 379, 753–763. [Google Scholar] [CrossRef] [PubMed]
- Mateo, J.; Ganji, G.; Lemech, C.; Burris, H.A.; Han, S.W.; Swales, K.; Kim, M.K.; Lee, K.H.; Hyun, M.S.; Lee, W.S.; et al. TP53 codon 72 polymorphism associated with prognosis in patients with advanced gastric cancer treated with paclitaxel and cisplatin. Clin. Cancer Res. 2009, 11, 355–360. [Google Scholar]
- Xu, Y.; Yao, L.; Ouyang, T.; Li, J.; Wang, T.; Fan, Z.; Lin, B.; Lu, Y.; Xie, Y. p53 Codon 72 Polymorphism Predicts the Pathologic Response to Neoadjuvant Chemotherapy in Patients with Breast Cancer. Clin. Cancer Res. 2005, 11, 7328–7333. [Google Scholar] [CrossRef]





{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Characteristics | Patients (n = 75) |
|---|---|
| Age of onset (years) | 34 |
| Breast cancer family history; n (%) | 18 (24%) |
| Molecular subtype; n (%) | |
| Triple negative | 14 (18.7%) |
| HER2/ER positive | 10 (13.3%) |
| Luminal B | 31 (41.3%) |
| Luminal A | 20 (26.7%) |
| Metastatic status; n (%) | 16 (21.3%) |
| Stage of breast cancer; n (%) | |
| I | 13 (17.3%) |
| II | 27 (36%) |
| III | 19 (25.3%) |
| IV | 16 (21.4%) |
| Gene | HGVSg | Protein Change | Type of Variant | dbSNP/ClinVar ID | Clinical Significance | OncoKB-db Targeted-Drug Prediction | Number of Carriers |
|---|---|---|---|---|---|---|---|
| BRCA1 | 17:g.43091349delinsTTTAAAGTGCAGCTTTTC | p.I1395Kfs | Frameshift | - | Likely-Pathogenic * | - | 1 |
| 17:g.43093581_43093582delinsT | p.I650Kfs | Frameshift | - | Likely-Pathogenic * | Olaparib, Talazoparib | 1 | |
| 17:g.43093821_43093822delinsT | p.P570Qfs | Frameshift | - | Pathogenic * | Olaparib, Talazoparib | 1 | |
| BRCA2 | 13:g.32316515_32316516delinsT | p.C19Sfs | Frameshift | - | Pathogenic * | Olaparib, Talazoparib | 1 |
| 13:g.32332277delinsGCATACAT | p.G267Afs | Frameshift | - | Likely-Pathogenic * | Olaparib, Talazoparib | 1 | |
| 13:g.32333103delinsTA | p.H543Tfs | Frameshift | - | Likely-Pathogenic * | Olaparib, Talazoparib | 1 | |
| 13:g.32338277delinsGACTTTGACAGAAA | p.E1308Dfs | Frameshift | - | Likely-Pathogenic * | Olaparib, Talazoparib | 1 | |
| 13:g.32340935_32340951delinsA | p.G2195Ffs | Frameshift | - | Pathogenic * | Olaparib, Talazoparib | 1 | |
| 13:g.32340959delinsGATGA | p.V2203Efs | Frameshift | - | Likely-Pathogenic * | Olaparib, Talazoparib | 1 | |
| 13:g.32379885delinsCA | p.T3033Nfs | Frameshift | rs397507419 | Pathogenic | Olaparib, Talazoparib | 21 | |
| 13:g.32398375_32398376delinsA | p.T3288Nfs | Frameshift | - | Pathogenic * | Olaparib, Talazoparib | 1 | |
| TP53 | 17:g.7674917delinsTC | p.P72R | Nonsense | - | Pathogenic * | - | 2 |
| STK11 | 19:g.1219400_1219456delinsT | p.X153_splice | Splice-site | - | Pathogenic * | - | 10 |
| PTEN | 10:g.87965294delinsTCTTATCA | p.Y346Lfs | Frameshift | - | Pathogenic * | GSK2636771, AZD8186 | 1 |
| 10:g.87965293_87965297delinsC | p.L345Pfs | Frameshift | - | Pathogenic * | GSK2636771, AZD8186 | 1 | |
| BRIP1 | 17:g.61683605_61683606delinsA | p.N1147Mfs | Frameshift | - | Pathogenic * | Olaparib | 2 |
| ATM | 11:g.108227881_108227882delinsG | p.F61Lfs | Frameshift | - | Pathogenic * | Olaparib | 1 |
| 11:g.108245025_108245026delinsA | p.G301fs | Frameshift | - | Pathogenic * | Olaparib | 1 | |
| 11:g.108282707delinsCATACAACACTAAAAAATG | p.X1193_splice | Splice-site | - | Pathogenic * | Olaparib | 1 | |
| 11:g.108302968_108302969delinsC | p.L1814Wfs | Frameshift | - | Pathogenic * | Olaparib | 1 | |
| 11:g.108326058delinsCCTTCTTCCAACAGAAACGATTGT | p.L2270Pfs | Frameshift | - | Pathogenic * | Olaparib | 1 | |
| 11:g.108329023delinsACTACAGGTTTTTTTGTTGTT | p.V2365Lfs | Frameshift | - | Pathogenic * | Olaparib | 1 | |
| 11:g.108329022delinsCCCAGGGTGTCATTCACCCT | p.V2365Qfs | Frameshift | - | Pathogenic * | Olaparib | 1 | |
| 11:g.108345760delinsTCAGTAGCTCAAGGG | p.F2813Qfs | Frameshift | - | Pathogenic * | Olaparib | 1 | |
| PALB2 | 16:g.23635659_23635660delinsA | p.M296 * | Nonsense | 143979 | Pathogenic | Olaparib | 4 |
| 16:g.23629919_23629925delinsT | p.Y743 * | Nonsense | - | Likely-Pathogenic * | Olaparib | 1 | |
| MSH6 | 2:g.47803500delinsAC | p.F1088Sfs | Frameshift | rs267608078 | Pathogenic | - | 4 |
| 2:g.47803657_47803658delinsT | p.G1139Afs | Frameshift | rs587781544 | Pathogenic | - | 1 | |
| 2:g.47806453delinsCTTAGAT | p.C1269 * | Nonsense | - | Pathogenic * | - | 2 | |
| PMS2 | 7:g.5987583_5987584delinsC | p.K394Sfs | Frameshift | rs1554298067 | Pathogenic | - | 1 |
| 7:g.5987525delinsCT | p.D414Rfs | Frameshift | rs267608159 | Pathogenic | - | 5 | |
| 7:g.5987525_5987526delinsC | p.D414Tfs | Frameshift | - | Pathogenic * | - | 6 | |
| CDKN2A | 9:g.21974732_21974737delinsC | p.L31Gfs | Frameshift | - | Pathogenic * | Palbociciclib, Ribociclib, Abernaciclib | 1 |
| RAD51C | 17:g.58734130delinsAATCCAGGAAATGCAGAAGAG | p.R347Nfs | Frameshift | - | Pathogenic * | Olaparib | 1 |
| RAD50 | 5:g.132595759_132595760delinsT | p.K722Rfs | Frameshift | rs397507178 | Pathogenic | - | 6 |
| Non-P/LP-Vs Group N = 25 | P/LP-Vs Group | |||
|---|---|---|---|---|
| N = 50 | p-Value * | |||
| Mean of age of onset (SD) | 34.8 (4.7) | 33.7 (4.4) | 0.4882 | |
| N (%) | N (%) | |||
| Family history | Yes | 7 (28) | 11 (22) | 0.3272 |
| No | 18 (72) | 39 (78) | ||
| Metastatic status | Yes | 7 (28) | 9 (18) | 0.0929 |
| No | 18 (72) | 41 (82) | ||
| Triple negative | Yes | 7 (28) | 7 (14) | 0.0151 |
| No | 18 (72) | 43 (86) | ||
| HER2 overexpression | Yes | 4 (16) | 6 (12) | 0.4149 |
| No | 21 (84) | 44 (88) | ||
| Luminal B | Yes | 8 (32) | 23 (46) | 0.0424 |
| No | 17 (68) | 27 (54) | ||
| Luminal A | Yes | 6 (24) | 14 (28) | 0.5190 |
| No | 19 (76) | 36 (72) | ||
| Study | Number of Patients | Age of Onset | P/LP-Vs Variant Rate |
|---|---|---|---|
| Morocco | |||
| Jouali et al. [32] | 15 | <50 years old | 26.7% |
| Bakkach et al. [33] | 82 | <40 years old | 16.7% |
| Palestine | |||
| Hamameh et al. [34] | 79 | <40 years old | 16.5% |
| Taiwan | |||
| Wang et al. [5] | 228 | <40 years old | 8.8% |
| Turkey | |||
| Akcay et al. [35] | 110 | <45 years old | 25.5% |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Panigoro, S.S.; Paramita, R.I.; Siswiandari, K.M.; Fadilah, F. Targeted Sequencing of Germline Breast Cancer Susceptibility Genes for Discovering Pathogenic/Likely Pathogenic Variants in the Jakarta Population. Diagnostics 2022, 12, 2241. https://doi.org/10.3390/diagnostics12092241
Panigoro SS, Paramita RI, Siswiandari KM, Fadilah F. Targeted Sequencing of Germline Breast Cancer Susceptibility Genes for Discovering Pathogenic/Likely Pathogenic Variants in the Jakarta Population. Diagnostics. 2022; 12(9):2241. https://doi.org/10.3390/diagnostics12092241
Chicago/Turabian StylePanigoro, Sonar Soni, Rafika Indah Paramita, Kristina Maria Siswiandari, and Fadilah Fadilah. 2022. "Targeted Sequencing of Germline Breast Cancer Susceptibility Genes for Discovering Pathogenic/Likely Pathogenic Variants in the Jakarta Population" Diagnostics 12, no. 9: 2241. https://doi.org/10.3390/diagnostics12092241
APA StylePanigoro, S. S., Paramita, R. I., Siswiandari, K. M., & Fadilah, F. (2022). Targeted Sequencing of Germline Breast Cancer Susceptibility Genes for Discovering Pathogenic/Likely Pathogenic Variants in the Jakarta Population. Diagnostics, 12(9), 2241. https://doi.org/10.3390/diagnostics12092241