
The Enigmatic Emerging Role of the C-Maf Inducing Protein in Cancer



Abstract
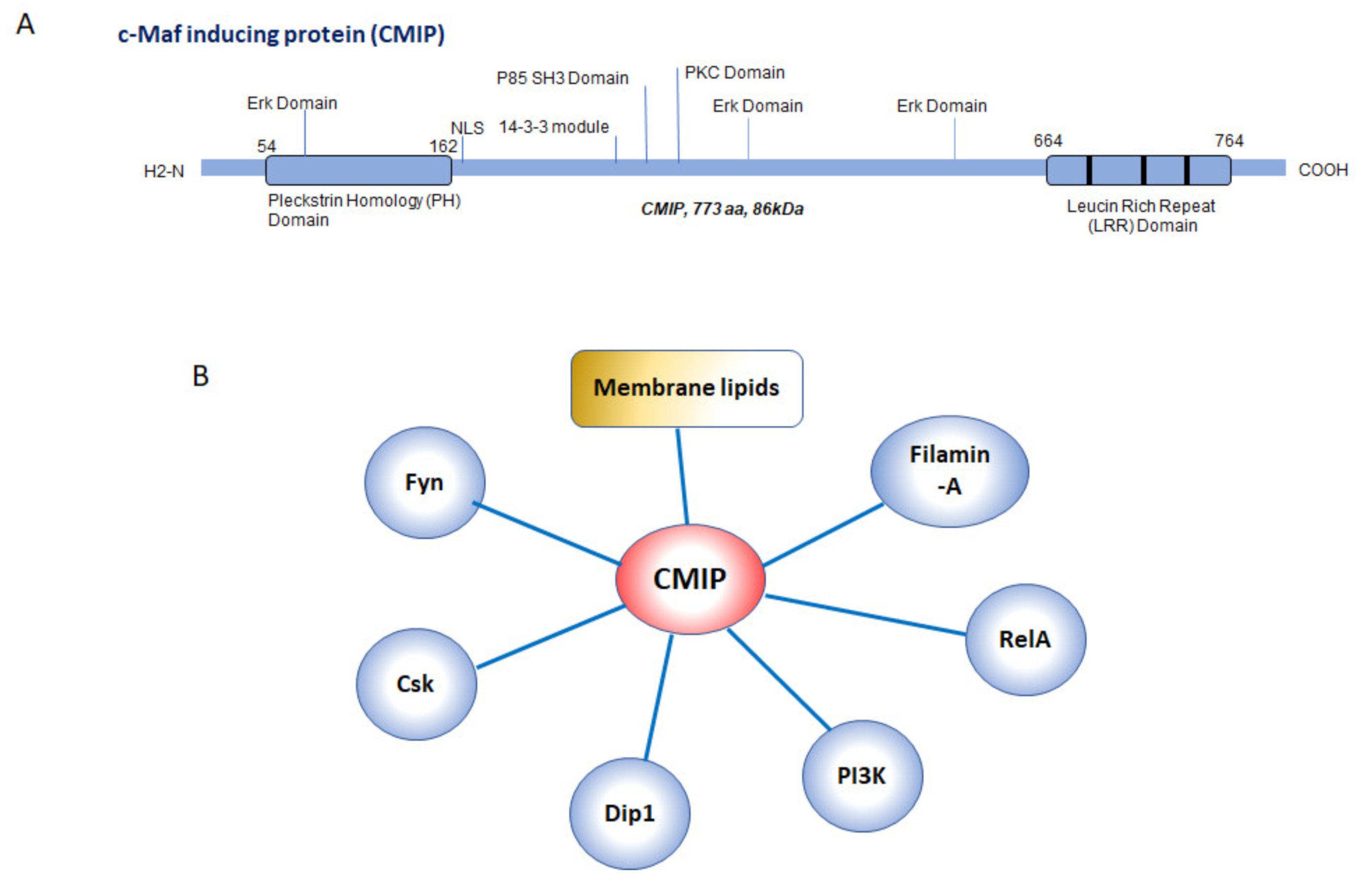
1. C-Maf Inducing Protein: Structural and Functional Aspects
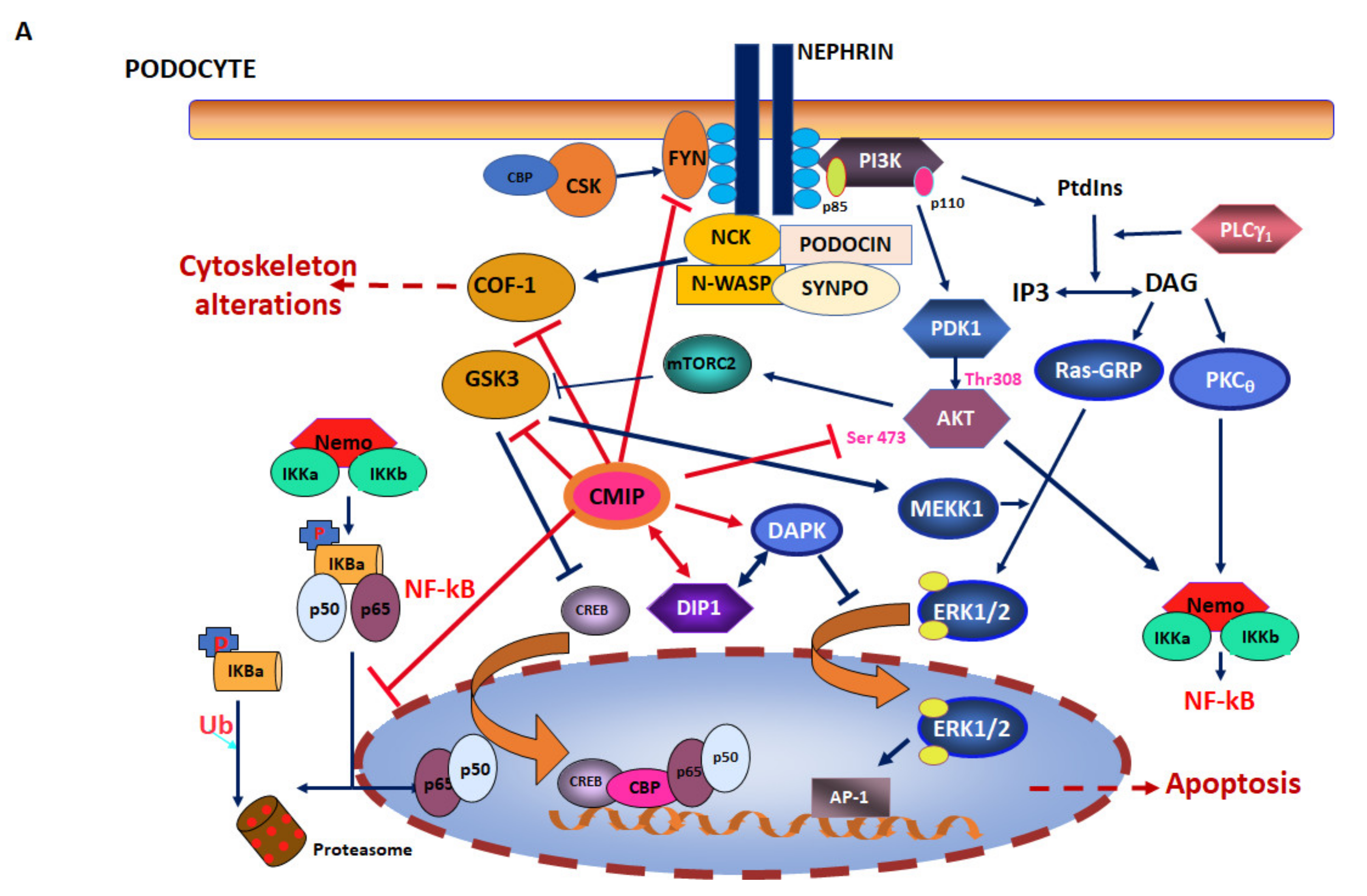
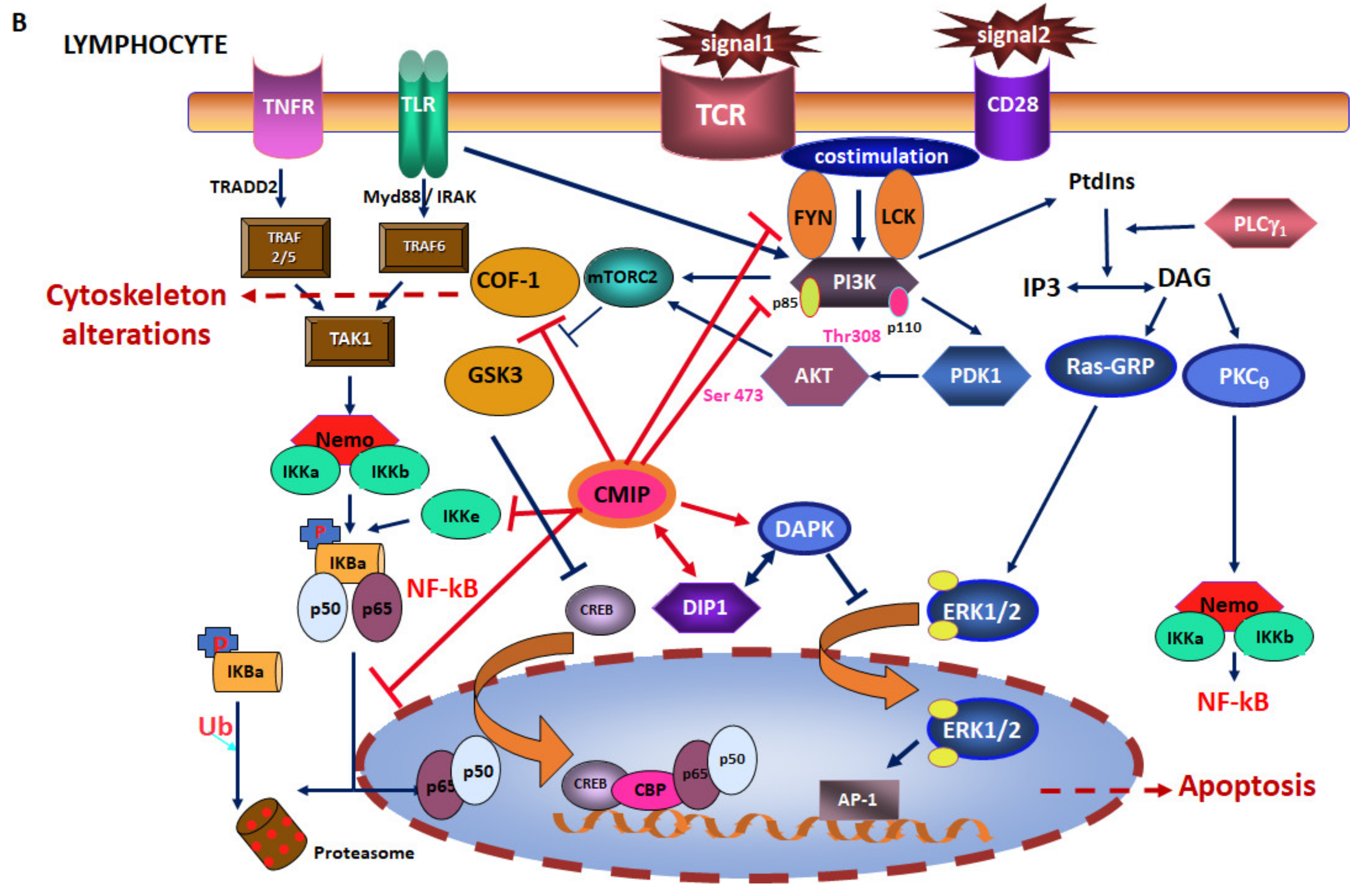
2. CMIP in Cancer Cells
2.1. CMIP in Hodgkin Lymphoma
2.2. CMIP in Solid Tumors
3. CMIP in Antiangiogenic Therapy
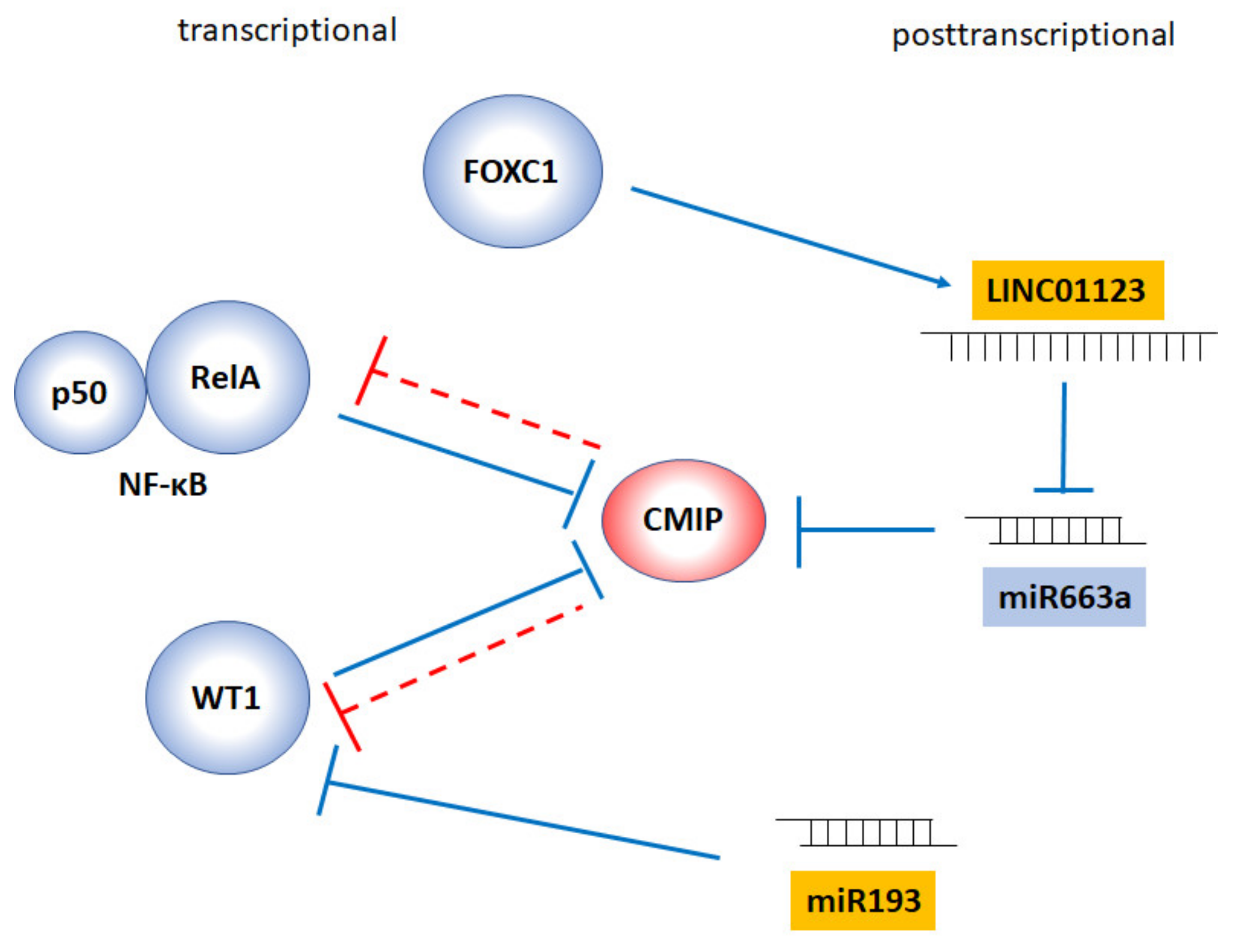
4. Regulation of CMIP Expression: The Dangerous Dance with WT1 and NF-κB
5. CMIP as a Matter of Life and Death
6. CMIP in Cancer Cells: Friend or Foe?
7. Conclusions and Open Questions
Author Contributions
Funding
Conflicts of Interest
Abbreviations
References
- Sahali, D.; Pawlak, A.; Valanciute, A.; Grimbert, P.; Lang, P.; Remy, P.; Bensman, A.; Guellaen, G. A novel approach to investigation of the pathogenesis of active minimal-change nephrotic syndrome using subtracted cDNA library screening. J. Am. Soc. Nephrol. 2002, 13, 1238–1247. [Google Scholar] [PubMed]
- Grimbert, P.; Valanciute, A.; Audard, V.; Pawlak, A.; Le gouvelo, S.; Lang, P.; Niaudet, P.; Bensman, A.; Guellaen, G.; Sahali, D. Truncation of C-mip (Tc-mip), a new proximal signaling protein, induces c-maf Th2 transcription factor and cytoskeleton reorganization. J. Exp. Med. 2003, 198, 797–807. [Google Scholar] [CrossRef] [PubMed]
- Nagase, T.; Kikuno, R.; Hattori, A.; Kondo, Y.; Okumura, K.; Ohara, O. Prediction of the coding sequences of unidentified human genes. XIX. The complete sequences of 100 new cDNA clones from brain which code for large proteins in vitro. DNA Res. 2000, 7, 347–355. [Google Scholar] [CrossRef]
- Newbury, D.F.; Winchester, L.; Addis, L.; Paracchini, S.; Buckingham, L.L.; Clark, A.; Cohen, W.; Cowie, H.; Dworzynski, K.; Everitt, A.; et al. CMIP and ATP2C2 modulate phonological short-term memory in language impairment. Am. J. Hum. Genet. 2009, 85, 264–272. [Google Scholar] [CrossRef] [PubMed]
- Moktefi, A.; Zhang, S.Y.; Vachin, P.; Ory, V.; Henique, C.; Audard, V.; Rucker-Martin, C.; Gouadon, E.; Eccles, M.; Schedl, A.; et al. Repression of CMIP transcription by WT1 is relevant to podocyte health. Kidney Int. 2016, 90, 1298–1311. [Google Scholar] [CrossRef]
- Boumediene, A.; Vachin, P.; Sendeyo, K.; Oniszczuk, J.; Zhang, S.Y.; Henique, C.; Pawlak, A.; Audard, V.; Ollero, M.; Guigonis, V.; et al. NEPHRUTIX: A randomized, double-blind, placebo vs Rituximab-controlled trial assessing T-cell subset changes in Minimal Change Nephrotic Syndrome. J. Autoimmun. 2018, 88, 91–102. [Google Scholar] [CrossRef]
- Zhang, S.Y.; Kamal, M.; Dahan, K.; Pawlak, A.; Ory, V.; Desvaux, D.; Audard, V.; Candelier, M.; BenMohamed, F.; Matignon, M.; et al. c-mip impairs podocyte proximal signaling and induces heavy proteinuria. Sci. Signal. 2010, 3, ra39. [Google Scholar] [CrossRef]
- Sendeyo, K.; Audard, V.; Zhang, S.Y.; Fan, Q.; Bouachi, K.; Ollero, M.; Rucker-Martin, C.; Gouadon, E.; Desvaux, D.; Bridoux, F.; et al. Upregulation of c-mip is closely related to podocyte dysfunction in membranous nephropathy. Kidney Int. 2013, 83, 414–425. [Google Scholar] [CrossRef]
- Bouachi, K.; Moktefi, A.; Zhang, S.Y.; Oniszczuk, J.; Sendeyo, K.; Remy, P.; Audard, V.; Pawlak, A.; Ollero, M.; Sahali, D. Expression of CMIP in podocytes is restricted to specific classes of lupus nephritis. PLoS ONE 2018, 13, e0207066. [Google Scholar] [CrossRef]
- Liu, Y.; Su, L.; Lin, Q.; Han, Y.; You, P.; Fan, Q. Induction of C-Mip by IL-17 Plays an Important Role in Adriamycin-Induced Podocyte Damage. Cell. Physiol. Biochem. 2015, 36, 1274–1290. [Google Scholar] [CrossRef]
- Kamal, M.; Valanciute, A.; Dahan, K.; Ory, V.; Pawlak, A.; Lang, P.; Guellaen, G.; Sahali, D. C-mip interacts physically with RelA and inhibits nuclear factor kappa B activity. Mol. Immunol. 2009, 46, 991–998. [Google Scholar] [CrossRef]
- Grimbert, P.; Valanciute, A.; Audard, V.; Lang, P.; Guellaen, G.; Sahali, D. The Filamin-A is a partner of Tc-mip, a new adapter protein involved in c-maf-dependent Th2 signaling pathway. Mol. Immunol. 2004, 40, 1257–1261. [Google Scholar] [CrossRef]
- Kamal, M.; Pawlak, A.; BenMohamed, F.; Valanciute, A.; Dahan, K.; Candelier, M.; Lang, P.; Guellaen, G.; Sahali, D. C-mip interacts with the p85 subunit of PI3 kinase and exerts a dual effect on ERK signaling via the recruitment of Dip1 and DAP kinase. FEBS Lett. 2010, 584, 500–506. [Google Scholar] [CrossRef]
- Audard, V.; Zhang, S.Y.; Copie-Bergman, C.; Rucker-Martin, C.; Ory, V.; Candelier, M.; Baia, M.; Lang, P.; Pawlak, A.; Sahali, D. Occurrence of minimal change nephrotic syndrome in classical Hodgkin lymphoma is closely related to the induction of c-mip in Hodgkin-Reed Sternberg cells and podocytes. Blood 2010, 115, 3756–3762. [Google Scholar] [CrossRef] [PubMed]
- Yu, L.; Lin, Q.; Feng, J.; Dong, X.; Chen, W.; Liu, Q.; Ye, J. Inhibition of nephrin activation by c-mip through Csk-Cbp-Fyn axis plays a critical role in Angiotensin II-induced podocyte damage. Cell. Signal. 2013, 25, 581–588. [Google Scholar] [CrossRef] [PubMed]
- Oniszczuk, J.; Sendeyo, K.; Chhuon, C.; Savas, B.; Cogne, E.; Vachin, P.; Henique, C.; Guerrera, I.C.; Astarita, G.; Frontera, V.; et al. CMIP is a negative regulator of T cell signaling. Cell. Mol. Immunol. 2020. [Google Scholar] [CrossRef]
- Yu, L.; Ye, J.; Liu, Q.; Feng, J.; Gu, X.; Sun, Q.; Lu, G. cMaf inducing protein inhibits cofilin1 activity and alters podocyte cytoskeleton organization. Mol. Med. Rep. 2017, 16, 4955–4963. [Google Scholar] [CrossRef] [PubMed]
- Dahlman, I.; Ryden, M.; Brodin, D.; Grallert, H.; Strawbridge, R.J.; Arner, P. Numerous Genes in Loci Associated with Body Fat Distribution Are Linked to Adipose Function. Diabetes 2016, 65, 433–437. [Google Scholar] [CrossRef]
- Sayols-Baixeras, S.; Hernaez, A.; Subirana, I.; Lluis-Ganella, C.; Munoz, D.; Fito, M.; Marrugat, J.; Elosua, R. DNA Methylation and High-Density Lipoprotein Functionality-Brief Report: The REGICOR Study (Registre Gironi del Cor). Arterioscler. Thromb. Vasc. Biol. 2017, 37, 567–569. [Google Scholar] [CrossRef]
- Li, C.X.; Gao, Y.; Gao, W.J.; Yu, C.Q.; Lyu, J.; Lyu, R.R.; Duan, J.L.; Sun, Y.; Guo, X.H.; Wang, S.F.; et al. Association between obesity and DNA methylation among the 7-16 year-old twins. Zhonghua Liu Xing Bing Xue Za Zhi 2018, 39, 443–448. [Google Scholar] [CrossRef]
- Fugmann, T.; Sofron, A.; Ritz, D.; Bootz, F.; Neri, D. The MHC Class II Immunopeptidome of Lymph Nodes in Health and in Chemically Induced Colitis. J. Immunol. 2017, 198, 1357–1364. [Google Scholar] [CrossRef] [PubMed]
- Keaton, J.M.; Gao, C.; Guan, M.; Hellwege, J.N.; Palmer, N.D.; Pankow, J.S.; Fornage, M.; Wilson, J.G.; Correa, A.; Rasmussen-Torvik, L.J.; et al. Genome-wide interaction with the insulin secretion locus MTNR1B reveals CMIP as a novel type 2 diabetes susceptibility gene in African Americans. Genet. Epidemiol. 2018, 42, 559–570. [Google Scholar] [CrossRef]
- Yan, Y.X.; Li, J.J.; Xiao, H.B.; Wang, S.; He, Y.; Wu, L.J. Association analysis of copy number variations in type 2 diabetes-related susceptible genes in a Chinese population. Acta Diabetol. 2018, 55, 909–916. [Google Scholar] [CrossRef]
- Cho, Y.S.; Chen, C.H.; Hu, C.; Long, J.; Ong, R.T.; Sim, X.; Takeuchi, F.; Wu, Y.; Go, M.J.; Yamauchi, T.; et al. Meta-analysis of genome-wide association studies identifies eight new loci for type 2 diabetes in east Asians. Nat. Genet. 2012, 44, 67–72. [Google Scholar] [CrossRef] [PubMed]
- Zhang, S.; Jamaspishvili, E.; Tong, H.; Chen, Y.; Zhou, Z.; Sun, L.; Kazakova, E.; Hong, Q. East Asian Genome-wide association study derived loci in relation to type 2 diabetes in the Han Chinese population. Acta Biochim. Pol. 2019, 66, 159–165. [Google Scholar] [CrossRef] [PubMed]
- Sakai, K.; Imamura, M.; Tanaka, Y.; Iwata, M.; Hirose, H.; Kaku, K.; Maegawa, H.; Watada, H.; Tobe, K.; Kashiwagi, A.; et al. Replication study for the association of 9 East Asian GWAS-derived loci with susceptibility to type 2 diabetes in a Japanese population. PLoS ONE 2013, 8, 6317. [Google Scholar] [CrossRef]
- Cao, Y.; Wang, T.; Wu, Y.; Juan, J.; Qin, X.; Tang, X.; Wu, T.; Hu, Y. Opposite Genetic Effects of CMIP Polymorphisms on the Risk of Type 2 Diabetes and Obesity: A Family-Based Study in China. Int. J. Mol. Sci. 2018, 19, 1011. [Google Scholar] [CrossRef] [PubMed]
- Wu, Y.; Gao, H.; Li, H.; Tabara, Y.; Nakatochi, M.; Chiu, Y.F.; Park, E.J.; Wen, W.; Adair, L.S.; Borja, J.B.; et al. A meta-analysis of genome-wide association studies for adiponectin levels in East Asians identifies a novel locus near WDR11-FGFR2. Hum. Mol. Genet. 2014, 23, 1108–1119. [Google Scholar] [CrossRef]
- Li, G.; Feng, D.; Wang, Y.; Fu, J.; Han, L.; Li, L.; Grant, S.F.A.; Li, M.; Gao, S. Loss of Cardio-Protective Effects at the CDH13 Locus Due to Gene-Sleep Interaction: The BCAMS Study. EBioMedicine 2018, 32, 164–171. [Google Scholar] [CrossRef]
- Benitez-Burraco, A.; Fernandez-Urquiza, M.; Jimenez-Romero, S. Language impairment with a microduplication in 1q42.3q43. Clin. Linguist. Phon. 2016, 1–26. [Google Scholar] [CrossRef]
- Kato, M.; Okanoya, K.; Koike, T.; Sasaki, E.; Okano, H.; Watanabe, S.; Iriki, A. Human speech- and reading-related genes display partially overlapping expression patterns in the marmoset brain. Brain Lang. 2014, 133, 26–38. [Google Scholar] [CrossRef] [PubMed]
- Newbury, D.F.; Gibson, J.L.; Conti-Ramsden, G.; Pickles, A.; Durkin, K.; Toseeb, U. Using Polygenic Profiles to Predict Variation in Language and Psychosocial Outcomes in Early and Middle Childhood. J. Speech Lang. Hear. Res. 2019, 62, 3381–3396. [Google Scholar] [CrossRef]
- Eicher, J.D.; Gruen, J.R. Language impairment and dyslexia genes influence language skills in children with autism spectrum disorders. Autism Res. 2015, 8, 229–234. [Google Scholar] [CrossRef]
- Zhenlong, C.; Jiangye, H.; Yanfen, Z.; Zhe, Z.; Zhihua, N.; Yuanxiang, H.; Weili, L.; Yongchao, L.; Daniel, R.S.; Runlin, Z.M.; et al. Differentially expressed genes identified in the main olfactory epithelium of mice with deficiency of adenylate cyclase 3 by using suppression subtractive hybridization approach. Yi Chuan 2014, 36, 574–583. [Google Scholar] [CrossRef] [PubMed]
- Pan, L.; Liao, Y.H.; Mo, M.Q.; Zhang, Q.H.; Yin, R.X. CMIP SNPs and their haplotypes are associated with dyslipidaemia and clinicopathologic features of IgA nephropathy. Biosci. Rep. 2020, 40. [Google Scholar] [CrossRef] [PubMed]
- Mo, M.Q.; Pan, L.; Lu, Q.M.; Li, Q.L.; Liao, Y.H. The association of the CMIP rs16955379 polymorphism with dyslipidemia and the clinicopathological features of IgA nephropathy. Int. J. Clin. Exp. Pathol. 2018, 11, 5008–5023. [Google Scholar]
- Bryant, E.K.; Dressen, A.S.; Bunker, C.H.; Hokanson, J.E.; Hamman, R.F.; Kamboh, M.I.; Demirci, F.Y. A multiethnic replication study of plasma lipoprotein levels-associated SNPs identified in recent GWAS. PLoS ONE 2013, 8, e63469. [Google Scholar] [CrossRef]
- Strawbridge, R.J.; Laumen, H.; Hamsten, A.; Breier, M.; Grallert, H.; Hauner, H.; Arner, P.; Dahlman, I. Effects of Genetic Loci Associated with Central Obesity on Adipocyte Lipolysis. PLoS ONE 2016, 11, e0153990. [Google Scholar] [CrossRef]
- Luo, M.; Fan, J.; Wenger, T.L.; Harr, M.H.; Racobaldo, M.; Mulchandani, S.; Dubbs, H.; Zackai, E.H.; Spinner, N.B.; Conlin, L.K. CMIP haploinsufficiency in two patients with autism spectrum disorder and co-occurring gastrointestinal issues. Am. J. Med. Genet. A 2017, 173, 2101–2107. [Google Scholar] [CrossRef]
- Audard, V.; Larousserie, F.; Grimbert, P.; Abtahi, M.; Sotto, J.J.; Delmer, A.; Boue, F.; Nochy, D.; Brousse, N.; Delarue, R.; et al. Minimal change nephrotic syndrome and classical Hodgkin’s lymphoma: Report of 21 cases and review of the literature. Kidney Int. 2006, 69, 2251–2260. [Google Scholar] [CrossRef]
- Bouatou, Y.; Koessler, T.; Oniszczuk, J.; Zhang, S.Y.; Moll, S.; Audard, V.; de Seigneux, S.; Sahali, D. Nephrotic Syndrome in Small Cell Lung Cancer and Induction of C-Mip in Podocytes. Am. J. Kidney Dis. 2017, 69, 477–480. [Google Scholar] [CrossRef]
- Zhang, J.; Huang, J.; Wang, X.; Chen, W.; Tang, Q.; Fang, M.; Qian, Y. CMIP is oncogenic in human gastric cancer cells. Mol. Med. Rep. 2017, 16, 7277–7286. [Google Scholar] [CrossRef]
- Xiang, R.; Han, X.; Ding, K.; Wu, Z. CMIP promotes Herceptin resistance of HER2 positive gastric cancer cells. Pathol. Res. Pract. 2020, 216, 152776. [Google Scholar] [CrossRef]
- Wang, B.; Wu, Z.S.; Wu, Q. CMIP Promotes Proliferation and Metastasis in Human Glioma. Biomed. Res. Int. 2017, 2017, 5340160. [Google Scholar] [CrossRef] [PubMed]
- Gerami, P.; Alsobrook, J.P., 2nd; Palmer, T.J.; Robin, H.S. Development of a novel noninvasive adhesive patch test for the evaluation of pigmented lesions of the skin. J. Am. Acad. Dermatol. 2014, 71, 237–244. [Google Scholar] [CrossRef] [PubMed]
- Li, Y.; Cui, S.; Shi, W.; Yang, B.; Yuan, Y.; Yan, S.; Xu, Y.; Zhang, Z.; Linlin, Z. Differential placental methylation in preeclampsia, preterm and term pregnancies. Placenta 2020, 93, 56–63. [Google Scholar] [CrossRef] [PubMed]
- Mouradov, D.; Sloggett, C.; Jorissen, R.N.; Love, C.G.; Li, S.; Burgess, A.W.; Arango, D.; Strausberg, R.L.; Buchanan, D.; Wormald, S.; et al. Colorectal cancer cell lines are representative models of the main molecular subtypes of primary cancer. Cancer Res. 2014, 74, 3238–3247. [Google Scholar] [CrossRef] [PubMed]
- Wang, C.C.N.; Li, C.Y.; Cai, J.H.; Sheu, P.C.; Tsai, J.J.P.; Wu, M.Y.; Li, C.J.; Hou, M.F. Identification of Prognostic Candidate Genes in Breast Cancer by Integrated Bioinformatic Analysis. J. Clin. Med. 2019, 8, 1160. [Google Scholar] [CrossRef]
- Izzedine, H.; Mangier, M.; Ory, V.; Zhang, S.Y.; Sendeyo, K.; Bouachi, K.; Audard, V.; Pechoux, C.; Soria, J.C.; Massard, C.; et al. Expression patterns of RelA and c-mip are associated with different glomerular diseases following anti-VEGF therapy. Kidney Int. 2014, 85, 457–470. [Google Scholar] [CrossRef]
- Ollero, M.; Sahali, D. Inhibition of the VEGF signalling pathway and glomerular disorders. Nephrol. Dial. Transplant. 2015, 30, 1449–1455. [Google Scholar] [CrossRef]
- Zhang, P.; Long, Q.; Zeng, S.; Wen, M.; Lu, Q. FOXC1-induced LINC01123 acts as a mediator in triple negative breast cancer. Cancer Cell Int. 2020, 20, 199. [Google Scholar] [CrossRef]
- Morrison, A.A.; Viney, R.L.; Saleem, M.A.; Ladomery, M.R. New insights into the function of the Wilms tumor suppressor gene WT1 in podocytes. Am. J. Physiol. Renal Physiol. 2008, 295, F12–F17. [Google Scholar] [CrossRef][Green Version]
- Ariyaratana, S.; Loeb, D.M. The role of the Wilms tumour gene (WT1) in normal and malignant haematopoiesis. Expert Rev. Mol. Med. 2007, 9, 1–17. [Google Scholar] [CrossRef]
- Li, T.S.; Chen, L.; Wang, S.C.; Yang, Y.Z.; Xu, H.J.; Gu, H.M.; Zhao, X.J.; Dong, P.; Pan, Y.; Shang, Z.Q.; et al. Magnesium isoglycyrrhizinate ameliorates fructose-induced podocyte apoptosis through downregulation of miR-193a to increase WT1. Biochem. Pharmacol. 2019, 166, 139–152. [Google Scholar] [CrossRef]
- Ory, V.; Fan, Q.; Hamdaoui, N.; Zhang, S.Y.; Desvaux, D.; Audard, V.; Candelier, M.; Noel, L.H.; Lang, P.; Guellaen, G.; et al. c-mip down-regulates NF-kappaB activity and promotes apoptosis in podocytes. Am. J. Pathol. 2012, 180, 2284–2292. [Google Scholar] [CrossRef]
- Zhang, T.; Saghatelian, A. Emerging roles of lipids in BCL-2 family-regulated apoptosis. Biochim. Biophys. Acta 2013, 1831, 1542–1554. [Google Scholar] [CrossRef]
- Li, G.; Kidd, J.; Kaspar, C.; Dempsey, S.; Bhat, O.M.; Camus, S.; Ritter, J.K.; Gehr, T.W.B.; Gulbins, E.; Li, P.L. Podocytopathy and Nephrotic Syndrome in Mice with Podocyte-Specific Deletion of the Asah1 Gene: Role of Ceramide Accumulation in Glomeruli. Am. J. Pathol. 2020. [Google Scholar] [CrossRef] [PubMed]
- Cancer Genome Atlas Network. Comprehensive molecular characterization of human colon and rectal cancer. Nature 2012, 487, 330–337. [Google Scholar] [CrossRef] [PubMed]
- Schulz, R.; McCole, R.B.; Woodfine, K.; Wood, A.J.; Chahal, M.; Monk, D.; Moore, G.E.; Oakey, R.J. Transcript- and tissue-specific imprinting of a tumour suppressor gene. Hum. Mol. Genet. 2009, 18, 118–127. [Google Scholar] [CrossRef]
- Yao, J.; Duan, L.; Fan, M.; Yuan, J.; Wu, X. Overexpression of BLCAP induces S phase arrest and apoptosis independent of p53 and NF-kappaB in human tongue carcinoma: BLCAP overexpression induces S phase arrest and apoptosis. Mol. Cell. Biochem. 2007, 297, 81–92. [Google Scholar] [CrossRef] [PubMed]





{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Type of Cancer | Type of Sample | Paraneoplastic Nephropathy Reported | Reference |
|---|---|---|---|
| Hodgkin Lymphoma | HRS Cells | Yes | [14] |
| Small Cell Lung Carcinoma | Cancer Tissue | Yes | [41] |
| Gastric Cancer | Cancer Tissue and Cell Lines (MKN45-HR and NCI-N87-HR) | No | [42,43] |
| Glioma | Cancer Cell Lines (A172, U251) | No | [44] |
| Melanoma | Surface Melanocytic Lesions | No | [45] |
| Choriocarcinoma | Trophoblastic Cell Lines | No | [46] |
| Colorectal cancer | Cell Lines | No | [47] |
| Breast Cancer | Cell Line Panel | No | [48] |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Ollero, M.; Sahali, D. The Enigmatic Emerging Role of the C-Maf Inducing Protein in Cancer. Diagnostics 2021, 11, 666. https://doi.org/10.3390/diagnostics11040666
Ollero M, Sahali D. The Enigmatic Emerging Role of the C-Maf Inducing Protein in Cancer. Diagnostics. 2021; 11(4):666. https://doi.org/10.3390/diagnostics11040666
Chicago/Turabian StyleOllero, Mario, and Dil Sahali. 2021. "The Enigmatic Emerging Role of the C-Maf Inducing Protein in Cancer" Diagnostics 11, no. 4: 666. https://doi.org/10.3390/diagnostics11040666
APA StyleOllero, M., & Sahali, D. (2021). The Enigmatic Emerging Role of the C-Maf Inducing Protein in Cancer. Diagnostics, 11(4), 666. https://doi.org/10.3390/diagnostics11040666