microRNA Expression Profile in Single Hormone Receptor-Positive Breast Cancers Is Mainly Dependent on HER2 Status—A Pilot Study

 , , , , , , , , and
, , , , , , , , and
Abstract
1. Introduction
2. Materials and Methods
2.1. Study Group
2.2. NanoString nCounter Assay for miRNA Profiling
2.3. The Cancer Genome Atlas (TCGA) miRNA Data Processing
2.4. miRNA Targets Prediction and Functional Annotation
2.5. The Cancer Genome Atlas (TCGA) mRNA Data Processing
3. Statistical Analysis
4. Results
4.1. Comparison of Study Groups
4.2. miRNA Expression Profile Associated with HER2 Status
4.3. miRNAs Associated with Steroid Hormone Receptor Expression
5. Discussion
6. Conclusions
Supplementary Materials
Author Contributions
Funding
Conflicts of Interest
References
- Kunc, M.; Biernat, W.; Senkus-Konefka, E. Estrogen receptor-negative progesterone receptor-positive breast cancer—“Nobody’s land“ or just an artifact? Cancer Treat. Rev. 2018, 67. [Google Scholar] [CrossRef] [PubMed]
- Yu, K.-D.; Jiang, Y.-Z.; Hao, S.; Shao, Z.-M. Molecular essence and endocrine responsiveness of estrogen receptor-negative, progesterone receptor-positive, and HER2-negative breast cancer. BMC Med. 2015, 13, 254. [Google Scholar] [CrossRef] [PubMed]
- He, Z.-Y.; Sun, J.-Y.; Li, F.-Y.; Lin, H.-X.; Wu, S.-G. Progesterone receptor loss identifies hormone receptor-positive and HER2-negative breast cancer subgroups at higher risk of relapse: A retrospective cohort study. Onco Targets Ther. 2016, 9, 1707. [Google Scholar] [CrossRef] [PubMed]
- Cui, X.; Schiff, R.; Arpino, G.; Osborne, C.K.; Lee, A.V. Biology of progesterone receptor loss in breast cancer and its implications for endocrine therapy. J. Clin. Oncol. 2005, 23, 7721–7735. [Google Scholar] [CrossRef] [PubMed]
- Senkus-Konefka, E.; Kunc, M.; Pęksa, R.; Łacko, A.; Radecka, B.; Braun, M.; Pikiel, J.; Litwiniuk, M.; Pogoda, K.; Cserni, G.; et al. ER-/PgR+ breast cancer is a separate entity characterized by distinct phenotype: Comprehensive reevaluation of cases from Polish and Hungarian centers. J. Clin. Oncol. 2020, 38, e12554. [Google Scholar] [CrossRef]
- Peng, Y.; Croce, C.M. The role of MicroRNAs in human cancer. Signal Transduct. Target. Ther. 2016, 1, 15004. [Google Scholar] [CrossRef]
- Esquela-Kerscher, A.; Slack, F.J. Oncomirs—MicroRNAs with a role in cancer. Nat. Rev. Cancer 2006, 6, 259–269. [Google Scholar] [CrossRef]
- Eichmüller, S.B.; Osen, W.; Mandelboim, O.; Seliger, B. Immune Modulatory microRNAs Involved in Tumor Attack and Tumor Immune Escape. JNCI J. Natl. Cancer Inst. 2017, 109. [Google Scholar] [CrossRef]
- Fu, S.W.; Chen, L.; Man, Y. miRNA Biomarkers in Breast Cancer Detection and Management. J. Cancer 2011, 2, 116–122. [Google Scholar] [CrossRef]
- Klinge, C.M. miRNAs and estrogen action. Trends Endocrinol. Metab. 2012, 23, 223–233. [Google Scholar] [CrossRef]
- Søkilde, R.; Persson, H.; Ehinger, A.; Pirona, A.C.; Fernö, M.; Hegardt, C.; Larsson, C.; Loman, N.; Malmberg, M.; Rydén, L.; et al. Refinement of breast cancer molecular classification by miRNA expression profiles. BMC Genom. 2019, 20. [Google Scholar] [CrossRef]
- Sekhon, J.S. Multivariate and Propensity Score Matching Software with Automated Balance Optimization: The Matching Package for R. J. Stat. Softw. 2011, 42. [Google Scholar] [CrossRef]
- Zbucka-Kretowska, M.; Niemira, M.; Paczkowska-Abdulsalam, M.; Bielska, A.; Szalkowska, A.; Parfieniuk, E.; Ciborowski, M.; Wolczynski, S.; Kretowski, A. Prenatal circulating microRNA signatures of foetal Down syndrome. Sci. Rep. 2019, 9, 2394. [Google Scholar] [CrossRef] [PubMed]
- Popeda, M.; Stokowy, T.; Bednarz-Knoll, N.; Jurek, A.; Niemira, M.; Bielska, A.; Kretowski, A.; Kalinowski, L.; Szade, J.; Markiewicz, A.; et al. NF-kappa B Signaling-Related Signatures Are Connected with the Mesenchymal Phenotype of Circulating Tumor Cells in Non-Metastatic Breast Cancer. Cancers 2019, 11, 1961. [Google Scholar] [CrossRef] [PubMed]
- The Cancer Genome Atlas Network Comprehensive molecular portraits of human breast tumours. Nature 2012, 490, 61–70. [CrossRef]
- Ciriello, G.; Gatza, M.L.; Beck, A.H.; Wilkerson, M.D.; Rhie, S.K.; Pastore, A.; Zhang, H.; McLellan, M.; Yau, C.; Kandoth, C.; et al. Comprehensive Molecular Portraits of Invasive Lobular Breast Cancer. Cell 2015, 163, 506–519. [Google Scholar] [CrossRef]
- Xu, T.; Su, N.; Liu, L.; Zhang, J.; Wang, H.; Zhang, W.; Gui, J.; Yu, K.; Li, J.; Le, T.D. miRBaseConverter: An R/Bioconductor package for converting and retrieving miRNA name, accession, sequence and family information in different versions of miRBase. BMC Bioinform. 2018, 19, 514. [Google Scholar] [CrossRef]
- Fan, Y.; Siklenka, K.; Arora, S.K.; Ribeiro, P.; Kimmins, S.; Xia, J. miRNet-dissecting miRNA-target interactions and functional associations through network-based visual analysis. Nucleic Acids Res. 2016. [Google Scholar] [CrossRef]
- Huang, D.W.; Sherman, B.T.; Lempicki, R.A. Bioinformatics enrichment tools: Paths toward the comprehensive functional analysis of large gene lists. Nucleic Acids Res. 2009, 37, 1–13. [Google Scholar] [CrossRef]
- Huang, D.W.; Sherman, B.T.; Lempicki, R.A. Systematic and integrative analysis of large gene lists using DAVID bioinformatics resources. Nat. Protoc. 2009, 4, 44–57. [Google Scholar] [CrossRef]
- Dusa, A. venn: Draw Venn Diagrams. R Packag. version 1.9. 2020. Available online: https//CRAN.R-project.org/package=venn (accessed on 28 July 2020).
- Mattie, M.D.; Benz, C.C.; Bowers, J.; Sensinger, K.; Wong, L.; Scott, G.K.; Fedele, V.; Ginzinger, D.; Getts, R.; Haqq, C. Optimized high-throughput microRNA expression profiling provides novel biomarker assessment of clinical prostate and breast cancer biopsies. Mol. Cancer 2006. [Google Scholar] [CrossRef] [PubMed]
- Lee, H.; Park, C.; Deftereos, G.; Morihara, J.; Stern, J.E.; Hawes, S.E.; Swisher, E.; Kiviat, N.B.; Feng, Q. MicroRNA expression in ovarian carcinoma and its correlation with clinicopathological features. World J. Surg. Oncol. 2012, 10, 174. [Google Scholar] [CrossRef] [PubMed]
- Wang, S.; Lin, R.-J. MicroRNA and HER2-overexpressing Cancer. MicroRNA 2013, 2, 137–147. [Google Scholar] [CrossRef] [PubMed]
- Galli de Amorim, M.; Branco, G.; Valieris, R.; Tarcitano, E.; Tojal da Silva, I.; Ferreira de Araújo, L.; Noronha Nunes, D.; Dias-Neto, E. The impact of HER2 overexpression on the miRNA and circRNA transcriptomes in two breast cell lines and their vesicles. Pharmacogenomics 2019, 20, 493–502. [Google Scholar] [CrossRef] [PubMed]
- Citron, F.; Segatto, I.; Vinciguerra, G.L.R.; Musco, L.; Russo, F.; Mungo, G.; D’Andrea, S.; Mattevi, M.C.; Perin, T.; Schiappacassi, M.; et al. Downregulation of miR-223 Expression Is an Early Event during Mammary Transformation and Confers Resistance to CDK4/6 Inhibitors in Luminal Breast Cancer. Cancer Res. 2020, 80, 1064–1077. [Google Scholar] [CrossRef]
- Ichikawa, T.; Sato, F.; Terasawa, K.; Tsuchiya, S.; Toi, M.; Tsujimoto, G.; Shimizu, K. Trastuzumab Produces Therapeutic Actions by Upregulating miR-26a and miR-30b in Breast Cancer Cells. PLoS ONE 2012, 7, e31422. [Google Scholar] [CrossRef]
- Luo, Q.; Wei, C.; Li, X.; Li, J.; Chen, L.; Huang, Y.; Song, H.; Li, D.; Fang, L. MicroRNA-195-5p is a potential diagnostic and therapeutic target for breast cancer. Oncol. Rep. 2014, 31, 1096–1102. [Google Scholar] [CrossRef]
- Yang, S.-J.; Yang, S.-Y.; Wang, D.-D.; Chen, X.; Shen, H.-Y.; Zhang, X.-H.; Zhong, S.-L.; Tang, J.-H.; Zhao, J.-H. The miR-30 family: Versatile players in breast cancer. Tumor Biol. 2017, 39, 101042831769220. [Google Scholar] [CrossRef]
- Tashkandi, H.; Shah, N.; Patel, Y.; Chen, H. Identification of new miRNA biomarkers associated with HER2-positive breast cancers. Oncoscience 2015, 2, 924–929. [Google Scholar] [CrossRef]
- Al-Nakhle, H.; Burns, P.A.; Cummings, M.; Hanby, A.M.; Hughes, T.A.; Satheesha, S.; Shaaban, A.M.; Smith, L.; Speirs, V. Estrogen Receptor β1 Expression Is Regulated by miR-92 in Breast Cancer. Cancer Res. 2010, 70, 4778–4784. [Google Scholar] [CrossRef]
- Moi, L.; Braaten, T.; Al-Shibli, K.; Lund, E.; Busund, L.-T.R. Differential expression of the miR-17-92 cluster and miR-17 family in breast cancer according to tumor type; results from the Norwegian Women and Cancer (NOWAC) study. J. Transl. Med. 2019, 17, 334. [Google Scholar] [CrossRef]
- Maillot, G.; Lacroix-Triki, M.; Pierredon, S.; Gratadou, L.; Schmidt, S.; Benes, V.; Roche, H.; Dalenc, F.; Auboeuf, D.; Millevoi, S.; et al. Widespread Estrogen-Dependent Repression of microRNAs Involved in Breast Tumor Cell Growth. Cancer Res. 2009, 69, 8332–8340. [Google Scholar] [CrossRef] [PubMed]
- Gilam, A.; Shai, A.; Ashkenazi, I.; Sarid, L.A.; Drobot, A.; Bickel, A.; Shomron, N. MicroRNA regulation of progesterone receptor in breast cancer. Oncotarget 2017, 8. [Google Scholar] [CrossRef]
- Le, X.-F.; Ling, H.; Mao, M.; Zhang, X.; Zhang, S.; Calin, G.A.; Wu, Y.; Bast, R.C. Abstract 3055: miR-495 functions as a novel regulator of the estrogen and progesterone receptorsin human breast cancers. In Proceedings of the AACR 104th Annual Meeting, Washington, DC, USA, 6–10 April 2013; p. 3055. [Google Scholar]
- Fiannaca, A.; La Rosa, M.; La Paglia, L.; Rizzo, R.; Urso, A. Analysis of miRNA expression profiles in breast cancer using biclustering. BMC Bioinform. 2015, 16 (Suppl. 4), S7. [Google Scholar] [CrossRef] [PubMed]
- Finlay-Schultz, J.; Cittelly, D.M.; Hendricks, P.; Patel, P.; Kabos, P.; Jacobsen, B.M.; Richer, J.K.; Sartorius, C.A. Progesterone downregulation of miR-141 contributes to expansion of stem-like breast cancer cells through maintenance of progesterone receptor and Stat5a. Oncogene 2015, 34, 3676–3687. [Google Scholar] [CrossRef] [PubMed]
- Lowery, A.J.; Miller, N.; Devaney, A.; McNeill, R.E.; Davoren, P.A.; Lemetre, C.; Benes, V.; Schmidt, S.; Blake, J.; Ball, G.; et al. MicroRNA signatures predict oestrogen receptor, progesterone receptor and HER2/neu receptor status in breast cancer. Breast Cancer Res. 2009, 11, R27. [Google Scholar] [CrossRef] [PubMed]
- Gorbatenko, A.; Søkilde, R.; Sorensen, E.E.; Newie, I.; Persson, H.; Morancho, B.; Arribas, J.; Litman, T.; Rovira, C.; Pedersen, S.F. HER2 and p95HER2 differentially regulate miRNA expression in MCF-7 breast cancer cells and downregulate MYB proteins through miR-221/222 and miR-503. Sci. Rep. 2019, 9, 3352. [Google Scholar] [CrossRef]
- Leivonen, S.K.; Mäkelä, R.; Östling, P.; Kohonen, P.; Haapa-Paananen, S.; Kleivi, K.; Enerly, E.; Aakula, A.; Hellström, K.; Sahlberg, N.; et al. Protein lysate microarray analysis to identify microRNAs regulating estrogen receptor signaling in breast cancer cell lines. Oncogene 2009. [Google Scholar] [CrossRef]





{kind=link}
{kind=link}
{kind=link}
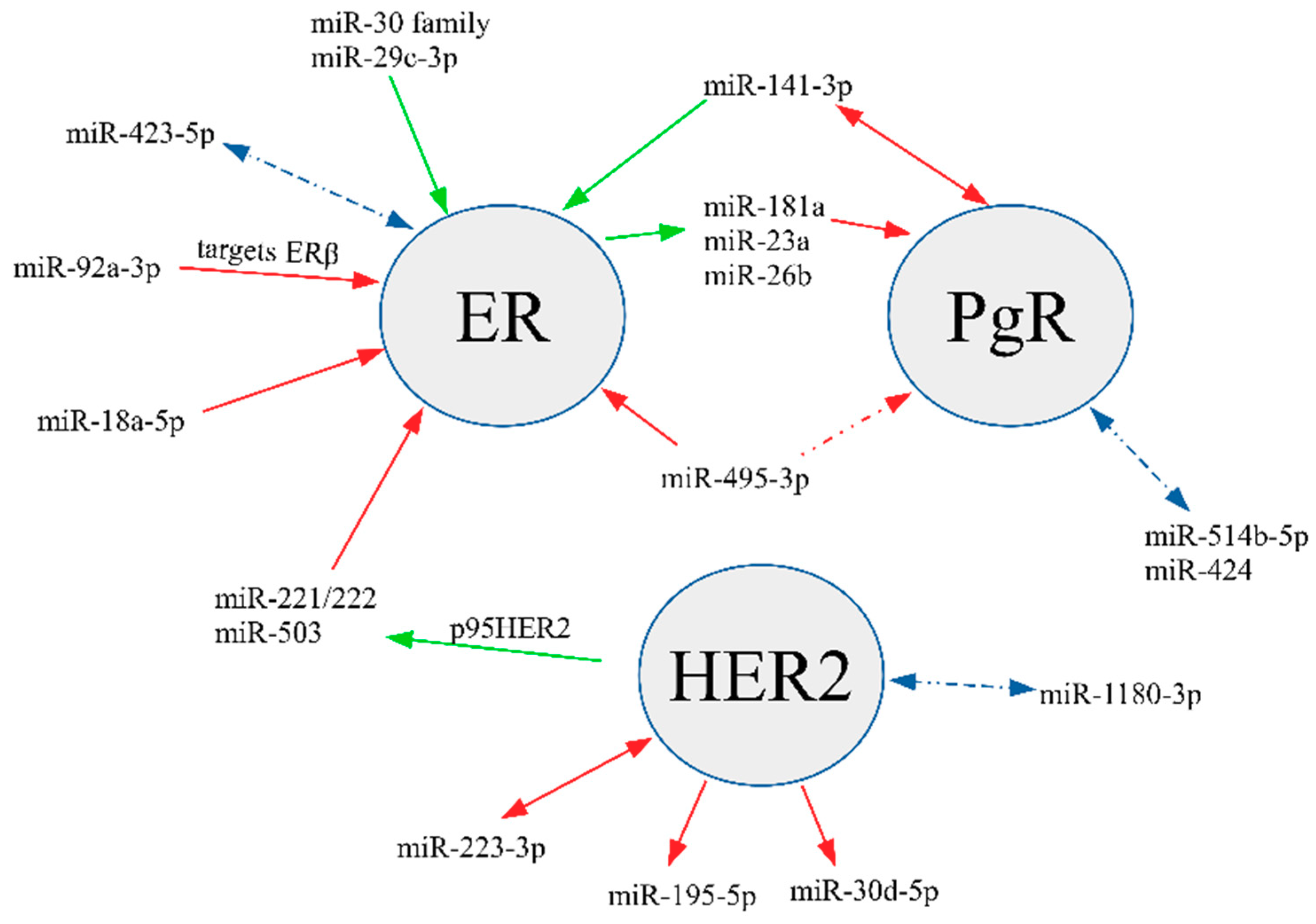{kind=link}
| Parameter | All (n = 32) | ER(+)PgR(−) (n = 14) | ER(−)PgR(+) (n = 18) | p-Value | ||||
|---|---|---|---|---|---|---|---|---|
| Age | median (range) | 62 | 29–78 | 66 | 36–76 | 53.5 | 29–78 | 0.141 |
| Grade | 1 | 0 | 0% | 0 | 0% | 0 | 0% | 0.003 |
| 2 | 6 | 19% | 6 | 19% | 0 | 0% | ||
| 3 | 26 | 81% | 8 | 25% | 18 | 56% | ||
| ER status | negative | 18 | 56% | 0 | 0% | 18 | 56% | <0.001 |
| positive | 14 | 44% | 14 | 44% | 0 | 0% | ||
| PgR status | negative | 14 | 44% | 14 | 44% | 0 | 0% | <0.001 |
| positive | 18 | 56% | 0 | 0% | 18 | 56% | ||
| HER2 status | negative | 15 | 47% | 5 | 16% | 10 | 31% | 0.308 |
| positive | 17 | 53% | 9 | 28% | 8 | 25% | ||
| Ki67 | median (range) | 47.5 | 9–90 | 30 | 9–70 | 60 | 30–90 | <0.001 |
| Tumor size [mm] | median (range) | 21 | 8–47 | 21.5 | 12–30 | 21 | 8–47 | 0.216 |
| T | 1 | 14 | 44% | 6 | 19% | 8 | 25% | 0.963 |
| 2 | 13 | 41% | 6 | 19% | 7 | 22% | ||
| 3 | 1 | 3% | 1 | 3% | 0 | 0% | ||
| 4 | 3 | 9% | 1 | 3% | 2 | 6% | ||
| NA | 1 | 3% | 0 | 0% | 1 | 3% | ||
| N | 0 | 17 | 53% | 8 | 25% | 9 | 28% | 0.351 |
| 1 | 11 | 34% | 6 | 19% | 5 | 16% | ||
| 2 | 3 | 9% | 0 | 0% | 3 | 9% | ||
| NA | 1 | 3% | 0 | 0% | 1 | 3% | ||
| M | 0 | 29 | 91% | 13 | 41% | 16 | 50% | 1.000 |
| 1 | 2 | 6% | 1 | 3% | 1 | 3% | ||
| NA | 1 | 3% | 0 | 0% | 1 | 3% | ||
| Parameter | All (n = 67) | ER(+)PgR(−) (n = 57) | ER(−)PgR(+) (n = 10) | p-Value | ||||
|---|---|---|---|---|---|---|---|---|
| Age | median (range) | 60 | 30–90 | 61 | 30–90 | 55.5 | 46–90 | 0.486 |
| ER status | negative | 10 | 15% | 0 | 0% | 10 | 15% | <0.001 |
| positive | 57 | 85% | 57 | 85% | 0 | 0% | ||
| PgR status | negative | 57 | 85% | 57 | 85% | 0 | 0% | <0.001 |
| positive | 10 | 15% | 0 | 0% | 10 | 15% | ||
| HER2 status | negative | 53 | 79% | 46 | 69% | 7 | 10% | 0.425 |
| positive | 14 | 21% | 11 | 16% | 3 | 4% | ||
| T | 1 | 13 | 19% | 12 | 18% | 1 | 1% | 0.437 |
| 2 | 44 | 66% | 35 | 52% | 9 | 13% | ||
| 3 | 9 | 13% | 9 | 13% | 0 | 0% | ||
| 4 | 1 | 1% | 1 | 1% | 0 | 0% | ||
| NA | 0 | 0% | 0 | 0% | 0 | 0% | ||
| N | 0 | 35 | 52% | 25 | 37% | 10 | 15% | 0.028 |
| 1 | 22 | 33% | 22 | 33% | 0 | 0% | ||
| 2 | 3 | 4% | 3 | 4% | 0 | 0% | ||
| 3 | 5 | 7% | 5 | 7% | 0 | 0% | ||
| NA | 2 | 3% | 2 | 3% | 0 | 0% | ||
| M | 0 | 48 | 72% | 39 | 58% | 9 | 13% | 0.260 |
| 1 | 0 | 0% | 0 | 0% | 0 | 0% | ||
| NA | 19 | 28% | 18 | 27% | 1 | 1% | ||
| miRNA | HER2 log2FC | Direction | HER2 p-Value | HER2 FDR | ER/PgR log2FC | ER/PgR p-Value | ER/PgR FDR |
|---|---|---|---|---|---|---|---|
| hsa-miR-887-5p | 2.85 | ↑ | 0.003 | 0.130 | 0.42 | 0.990 | 0.998 |
| hsa-miR-208a-3p | 2.50 | ↑ | 0.012 | 0.156 | 0.79 | 0.541 | 0.847 |
| hsa-miR-891a-5p | 2.29 | ↑ | 0.020 | 0.160 | 0.85 | 0.473 | 0.826 |
| hsa-miR-301a-5p | 2.17 | ↑ | 0.026 | 0.160 | 1.26 | 0.178 | 0.811 |
| hsa-miR-33b-5p | 2.09 | ↑ | 0.024 | 0.160 | 0.69 | 0.598 | 0.864 |
| hsa-miR-1296-3p | 1.91 | ↑ | 0.007 | 0.130 | 0.53 | 0.675 | 0.898 |
| hsa-miR-378d | 1.91 | ↑ | 0.002 | 0.130 | 0.14 | 0.702 | 0.908 |
| hsa-miR-548v | 1.82 | ↑ | 0.027 | 0.160 | 0.82 | 0.400 | 0.811 |
| hsa-miR-1295a | 1.73 | ↑ | 0.005 | 0.130 | 0.73 | 0.338 | 0.811 |
| hsa-miR-337-3p | 1.54 | ↑ | 0.014 | 0.156 | −0.11 | 0.480 | 0.827 |
| hsa-miR-514b-5p | 0.83 | ↑ | 0.023 | 0.160 | −0.67 | 0.046 | 0.809 |
| hsa-miR-185-5p | −0.71 | ↓ | 0.032 | 0.186 | 0.13 | 0.410 | 0.811 |
| hsa-miR-340-5p | −0.72 | ↓ | 0.025 | 0.160 | −0.27 | 0.666 | 0.898 |
| hsa-miR-424-5p | −0.74 | ↓ | 0.020 | 0.160 | −0.69 | 0.048 | 0.809 |
| hsa-miR-106a-5p+hsa-miR-17-5p | −0.82 | ↓ | 0.007 | 0.130 | −0.40 | 0.379 | 0.811 |
| hsa-miR-151a-3p | −0.84 | ↓ | 0.021 | 0.160 | 0.29 | 0.221 | 0.811 |
| hsa-miR-374a-5p | −0.91 | ↓ | 0.013 | 0.156 | 0.10 | 0.435 | 0.811 |
| hsa-miR-141-3p | −0.92 | ↓ | 0.034 | 0.193 | 1.10 | 0.018 | 0.809 |
| hsa-miR-26b-5p | −0.96 | ↓ | 0.025 | 0.160 | 0.54 | 0.110 | 0.809 |
| hsa-miR-126-3p | −0.97 | ↓ | 0.006 | 0.130 | 0.20 | 0.255 | 0.811 |
| hsa-miR-32-5p | −1.04 | ↓ | 0.017 | 0.160 | 0.19 | 0.349 | 0.811 |
| hsa-miR-15b-5p | −1.08 | ↓ | 0.023 | 0.160 | −0.55 | 0.425 | 0.811 |
| hsa-miR-30d-5p | −1.12 | ↓ | 0.006 | 0.130 | 0.53 | 0.078 | 0.809 |
| hsa-miR-1180-3p | −1.18 | ↓ | 0.008 | 0.142 | −0.16 | 0.867 | 0.953 |
| hsa-miR-30b-5p | −1.25 | ↓ | 0.002 | 0.130 | 0.13 | 0.310 | 0.811 |
| hsa-miR-195-5p | −1.33 | ↓ | 0.024 | 0.160 | 0.49 | 0.209 | 0.811 |
| hsa-miR-429 | −1.39 | ↓ | 0.023 | 0.160 | −0.27 | 0.997 | 0.998 |
| hsa-miR-503-5p | −1.46 | ↓ | 0.011 | 0.156 | −1.19 | 0.079 | 0.809 |
| hsa-miR-223-3p | −1.47 | ↓ | 0.025 | 0.160 | −1.30 | 0.087 | 0.809 |
| hsa-miR-450a-5p | −1.60 | ↓ | 0.006 | 0.130 | −0.74 | 0.397 | 0.811 |
| hsa-miR-29b-3p | −1.63 | ↓ | 0.001 | 0.130 | 0.36 | 0.135 | 0.811 |
| hsa-miR-135b-5p | −2.17 | ↓ | 0.016 | 0.160 | −1.46 | 0.184 | 0.811 |
| hsa-miR-660-5p | −2.30 | ↓ | 0.010 | 0.147 | −1.49 | 0.167 | 0.811 |
| miRNA | ER/PgR log2FC | Direction | ER/PgR p-Value | ER/PgR FDR | HER2 log2FC | HER2 p-Value | HER2 FDR |
|---|---|---|---|---|---|---|---|
| hsa-miR-30a-5p | 1.91 | ↑ | 0.031 | 0.809 | −1.69 | 0.046 | 0.221 |
| hsa-miR-29c-3p | 1.40 | ↑ | 0.030 | 0.809 | −1.23 | 0.047 | 0.221 |
| hsa-miR-141-3p | 1.10 | ↑ | 0.018 | 0.809 | −0.92 | 0.034 | 0.193 |
| hsa-miR-423-5p | 0.73 | ↑ | 0.045 | 0.809 | −0.49 | 0.119 | 0.338 |
| hsa-miR-514b-5p | −0.67 | ↓ | 0.046 | 0.809 | 0.83 | 0.023 | 0.160 |
| hsa-miR-424-5p | −0.69 | ↓ | 0.048 | 0.809 | −0.74 | 0.020 | 0.160 |
| hsa-miR-495-3p | −2.05 | ↓ | 0.027 | 0.809 | −0.51 | 0.773 | 0.851 |
| hsa-miR-92a-3p | −2.32 | ↓ | 0.033 | 0.809 | −1.27 | 0.206 | 0.419 |
© 2020 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Kunc, M.; Popęda, M.; Szałkowska, A.; Niemira, M.; Bieńkowski, M.; Pęksa, R.; Łacko, A.; Radecka, B.S.; Braun, M.; Pikiel, J.; et al. microRNA Expression Profile in Single Hormone Receptor-Positive Breast Cancers Is Mainly Dependent on HER2 Status—A Pilot Study. Diagnostics 2020, 10, 617. https://doi.org/10.3390/diagnostics10090617
Kunc M, Popęda M, Szałkowska A, Niemira M, Bieńkowski M, Pęksa R, Łacko A, Radecka BS, Braun M, Pikiel J, et al. microRNA Expression Profile in Single Hormone Receptor-Positive Breast Cancers Is Mainly Dependent on HER2 Status—A Pilot Study. Diagnostics. 2020; 10(9):617. https://doi.org/10.3390/diagnostics10090617
Chicago/Turabian StyleKunc, Michał, Marta Popęda, Anna Szałkowska, Magdalena Niemira, Michał Bieńkowski, Rafał Pęksa, Aleksandra Łacko, Barbara S. Radecka, Marcin Braun, Joanna Pikiel, and et al. 2020. "microRNA Expression Profile in Single Hormone Receptor-Positive Breast Cancers Is Mainly Dependent on HER2 Status—A Pilot Study" Diagnostics 10, no. 9: 617. https://doi.org/10.3390/diagnostics10090617
APA StyleKunc, M., Popęda, M., Szałkowska, A., Niemira, M., Bieńkowski, M., Pęksa, R., Łacko, A., Radecka, B. S., Braun, M., Pikiel, J., Litwiniuk, M., Pogoda, K., Iżycka-Świeszewska, E., Krętowski, A., Żaczek, A. J., Biernat, W., & Senkus-Konefka, E. (2020). microRNA Expression Profile in Single Hormone Receptor-Positive Breast Cancers Is Mainly Dependent on HER2 Status—A Pilot Study. Diagnostics, 10(9), 617. https://doi.org/10.3390/diagnostics10090617