
Taming Prebiotic Chemistry: The Role of Heterogeneous and Interfacial Catalysis in the Emergence of a Prebiotic Catalytic/Information Polymer System

{kind=link}
{kind=link}
{kind=link}
Abstract
:1. Introduction
2. RNA World Hypothesis
2.1. Availability of the Ribonucleotides
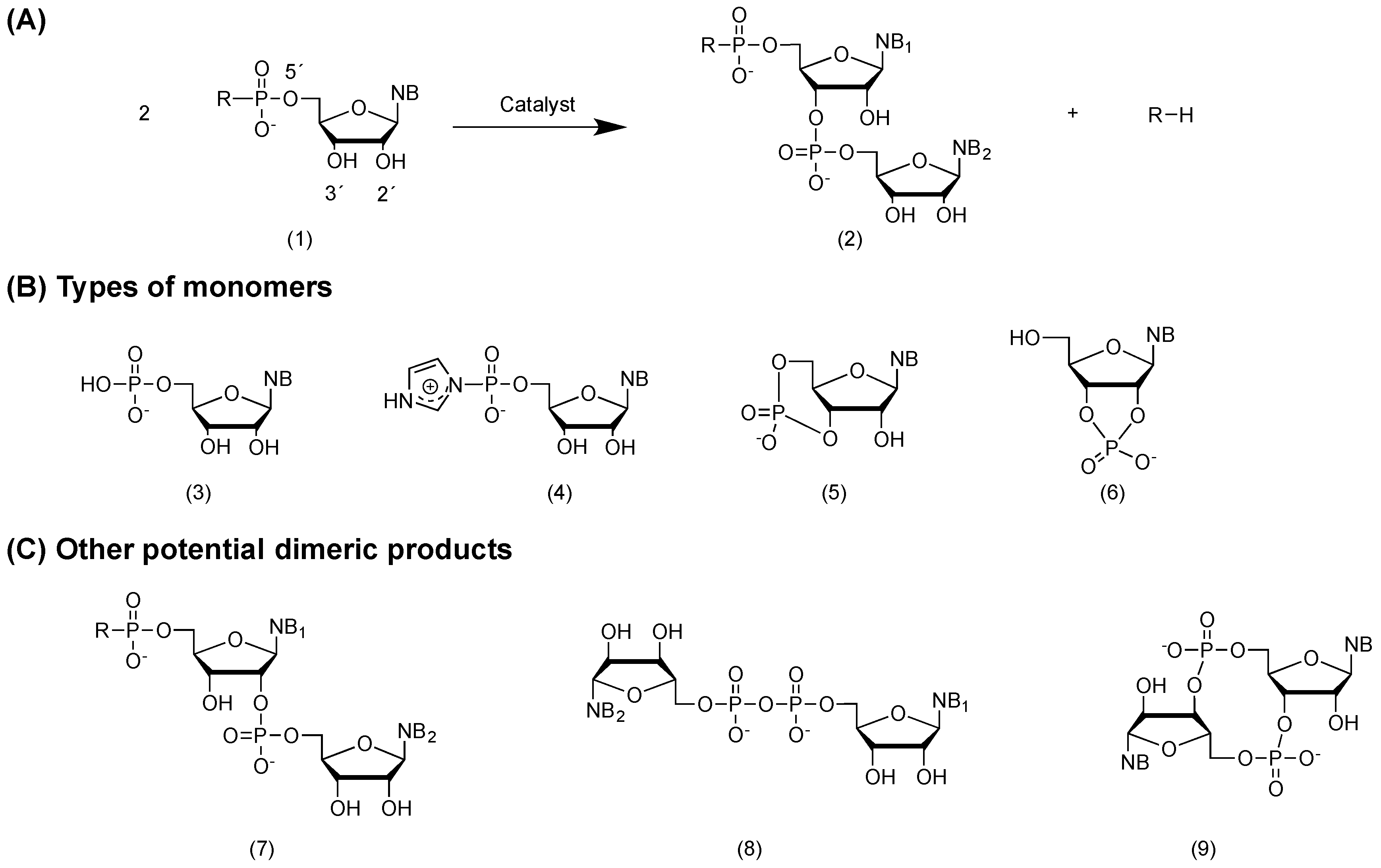
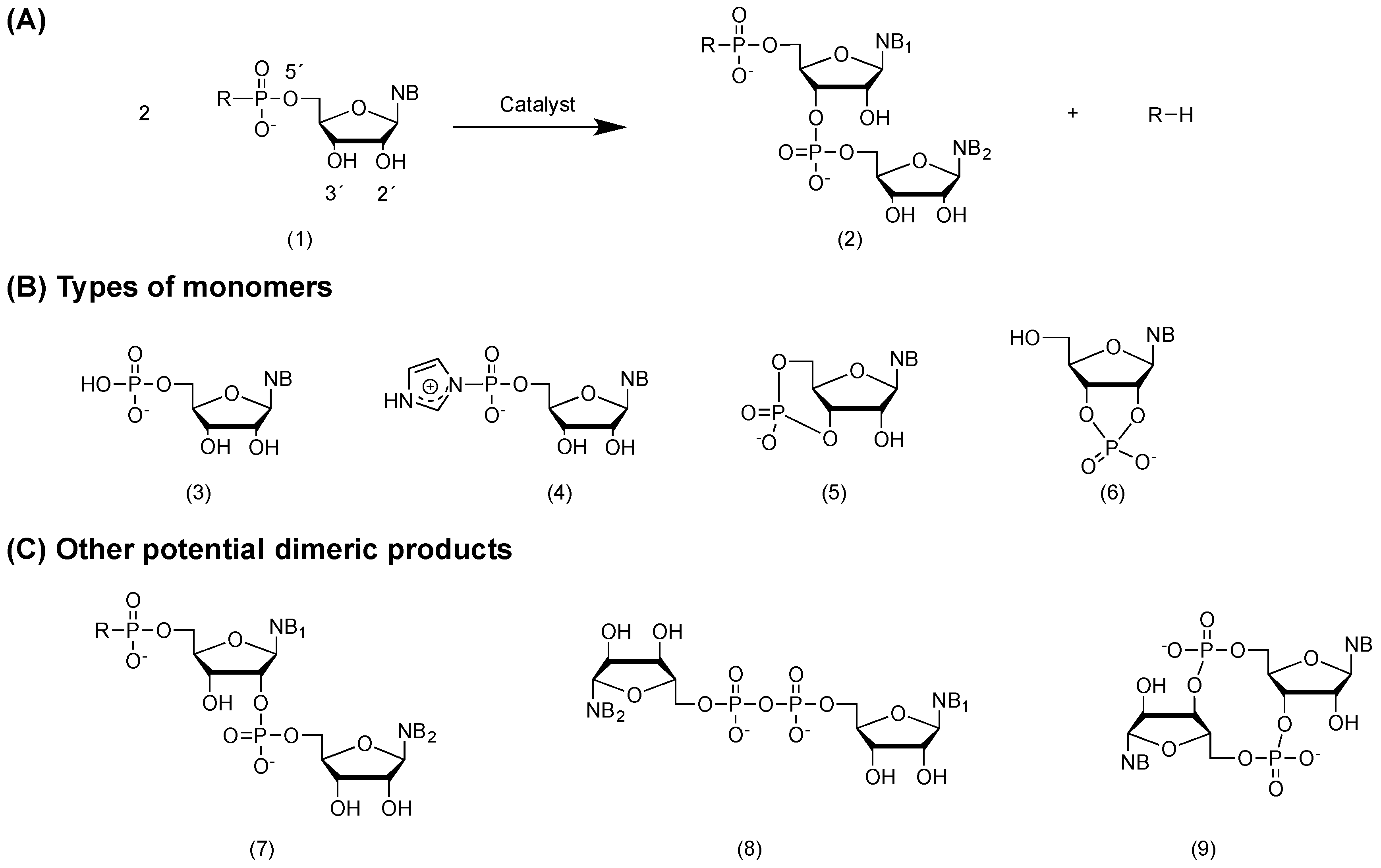
2.2. General Consideration on Non-Enzymatic RNA Polymerization from Monomers
2.3. Chemical Activation of RNA Monomers
3. RNA Polymerization in Bulk Aqueous Media
3.1. Non-Enzymatic Self-Condensation of RNA Monomers in Aqueous Media
3.2. Non-Enzymatic, Template-Directed Polymerization of RNA Monomers in Aqueous Media
4. Heterogeneous Media as Supporting Environments for RNA Polymerization
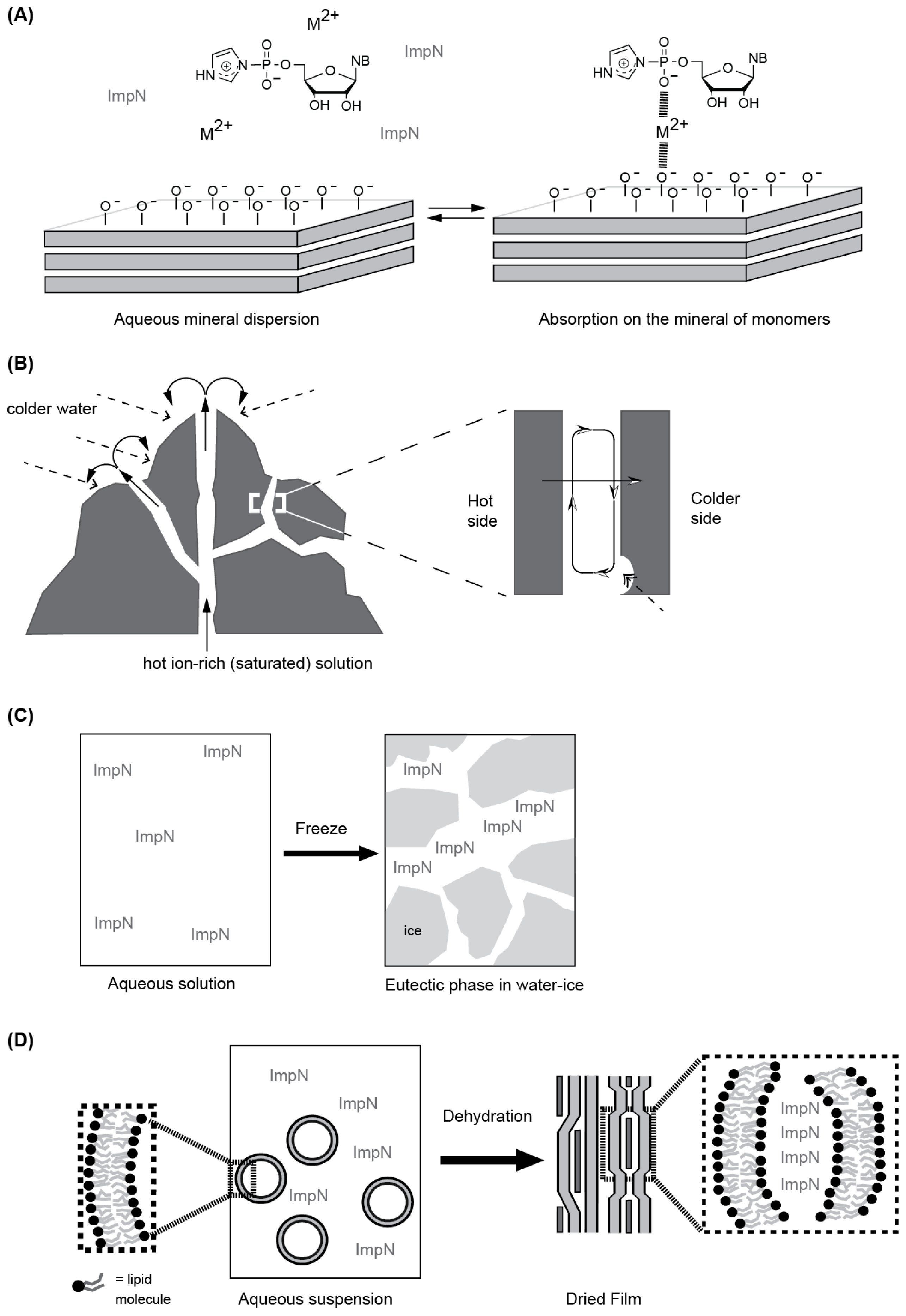
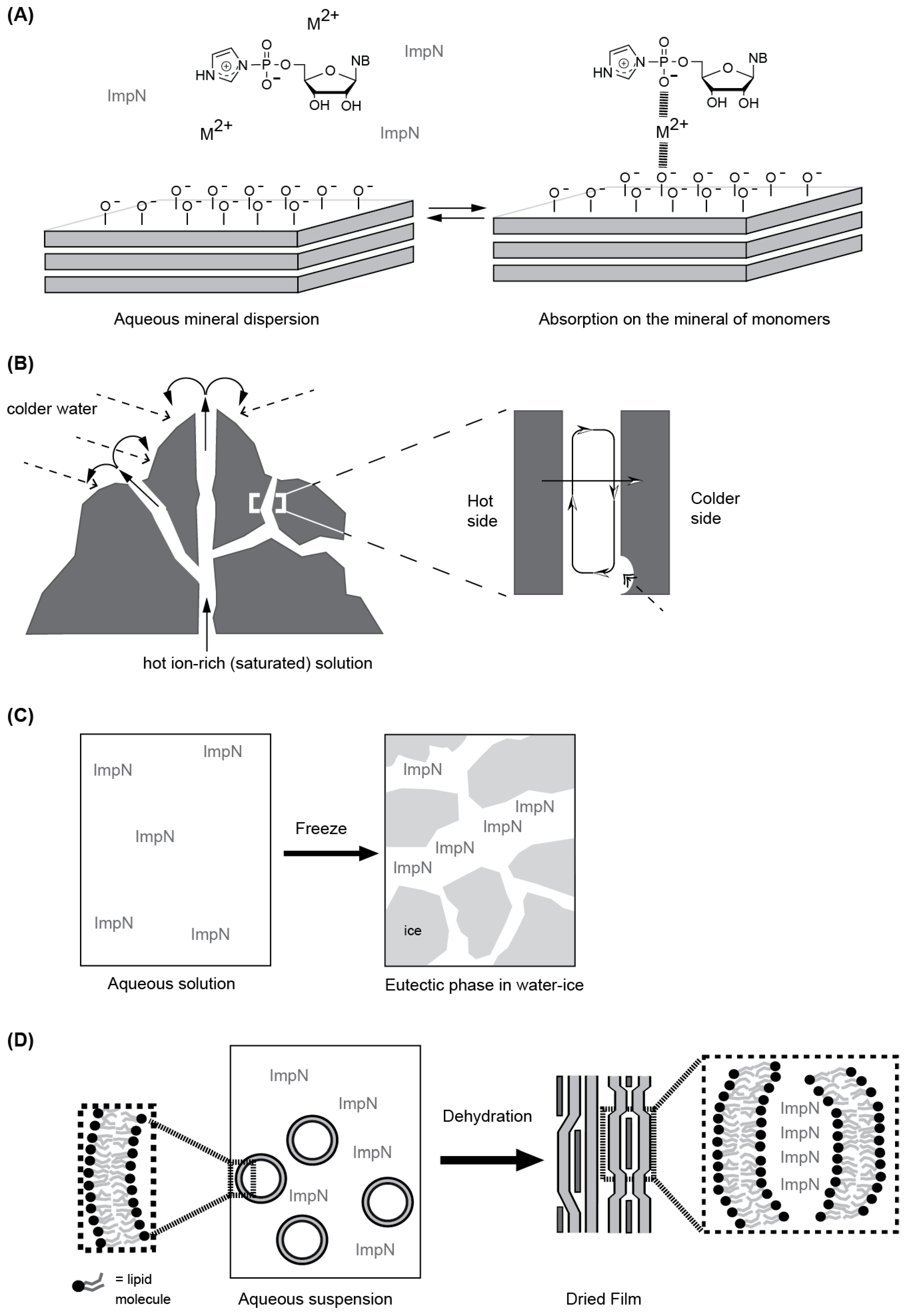
4.1. Rationales behind the Heterogeneous Media and Their Nature
4.1.1. Mineral Surfaces and Formations
4.1.2. Eutectic Phases
4.1.3. Self-Assembled Molecular Systems
5. Relevance of the Various Approaches to the Non-Enzymatic RNA Polymerization
6. Conclusions
Acknowledgments
Conflicts of Interest
References
- Orgel, L.E. Prebiotic chemistry and the origin of the RNA world. Crit. Rev. Biochem. Mol. Biol. 2004, 39, 99–123. [Google Scholar] [PubMed]
- Pressman, A.; Bianco, C.; Chen, I.A. The RNA world as a model system to study the origins of life. Curr. Biol. Rev. 2015, 25, R953–R963. [Google Scholar] [CrossRef] [PubMed]
- Martin, L.; Unrau, P.; Müller, U. RNA synthesis by in vitro selected ribozymes for recreating an RNA world. Life 2015, 5, 247–268. [Google Scholar] [CrossRef] [PubMed]
- Kruger, K.; Grabowski, P.J.; Zaug, A.J.; Sands, J.; Gottschling, D.E.; Cech, T.R. Self-splicing RNA: Autoexcision and autocyclization of the ribosomal RNA intervening sequence of tetrahymena. Cell 1982, 31, 147–157. [Google Scholar] [CrossRef]
- Guerrier-Takada, C.; Gardiner, K.; Marsh, T.; Pace, N.; Altman, S. The RNA moiety of ribonuclease p is the catalytic subunit of the enzyme. Cell 1983, 35, 849–857. [Google Scholar] [CrossRef]
- Nissen, P.; Hassen, J.; Ban, N.; Moore, P.B.; Steitz, T.A. The structural basis of ribosome activity in peptide bond synthesis. Science 2000, 289, 920–930. [Google Scholar] [CrossRef] [PubMed]
- Sprengel, G.; Follmann, H. Evidence for the reductive pathway of deoxyribonucleotide synthesis in an archaebacterium. FEBS Lett. 1981, 132, 207–209. [Google Scholar] [CrossRef]
- Menor-Salván, C.; Ruiz-Bermejo, D.M.; Guzmán, M.I.; Osuna-Esteban, S.; Veintemillas-Verdaguer, S. Synthesis of pyrimidines and triazines in ice: Implications for the prebiotic chemistry of nucleobases. Chem. A Eur. J. 2009, 15, 4411–4418. [Google Scholar] [CrossRef] [PubMed]
- Kopetzki, D.; Antonietti, M. Hydrothermal formose reaction. New J. Chem. 2011, 35, 1787–1794. [Google Scholar] [CrossRef]
- Pasek, M.A.; Dworkin, J.P.; Lauretta, D.S. A radical pathway for organic phosphorylation during schreibersite corrosion with implications for the origin of life. Geochim. Cosmochim. Acta 2007, 71, 1721–1736. [Google Scholar] [CrossRef]
- Powner, M.W.; Gerland, B.; Sutherland, J.D. Synthesis of activated pyrimidine ribonucleotides in prebiotically plausible conditions. Nature 2009, 459, 239–242. [Google Scholar] [CrossRef] [PubMed]
- Levy, M.; Miller, S.L. The stability of the RNA bases: Implications for the origin of life. Proc. Natl. Acad. Sci. USA 1998, 95, 7933–7938. [Google Scholar] [CrossRef] [PubMed]
- Larralde, R.; Robertson, M.P.; Miller, S.L. Rates of decomposition of ribose and other sugars: Implication for chemical evolution. Proc. Natl. Acad. Sci. USA 1995, 92, 8158–8160. [Google Scholar] [CrossRef] [PubMed]
- Cafferty, B.J.; Hud, N.V. Abiotic synthesis of RNA in water: A common goal of prebiotic chemistry and bottom-up synthetic biology. Curr. Opin. Chem. Biol. 2014, 22, 146–157. [Google Scholar] [CrossRef] [PubMed]
- Cafferty, B.J.; Hud, N.V. Was a pyrimidine-pyrimidine base pair the ancestor of watson-crick base pairs? Insights from a systematic approach to the origin of RNA. Isr. J. Chem. 2015, 55, 891–905. [Google Scholar] [CrossRef]
- Krishnamurthy, R. On the emergence of RNA. Isr. J. Chem. 2015, 55, 837–850. [Google Scholar] [CrossRef]
- Dörr, M.; Löffler, P.M.G.; Monnard, P.-A. Nonenzymatic, template-directed nucleic acid replication: Relevance to the origins of infor-mation transfer and to novel synthetic metholodogies. Curr. Org. Synth. 2012, 9, 735–763. [Google Scholar]
- Joyce, G.F. The antiquity of RNA-base evolution. Nature 2002, 418, 214–221. [Google Scholar] [CrossRef] [PubMed]
- Kanavarioti, A.; Rosenbach, M.T.; Hurley, T.B. Nucleotides as nucleophiles: Reactions of nucleotides with phosphoimidazolides activated guanosine. Orig. Life Evol. Biosphere 1992, 21, 199–217. [Google Scholar] [CrossRef]
- Dickson, K.S.; Burns, C.M.; Richardson, J.P. Determination of the free-energy change for repair of a DNA phosphodiester bond. J. Biol. Chem. 2000, 275, 15828–15831. [Google Scholar] [CrossRef] [PubMed]
- Von Kiedrowski, G. A self-replicating hexadeoxynucleotide. Angew. Chem. Int. Ed. Engl. 1986, 25, 932–935. [Google Scholar] [CrossRef]
- Weimann, B.J.; Lohrmann, R.; Orgel, L.E.; Schneider-Bernloehr, H.; Sulston, J.E. Template-directed synthesis with adenosine-5′-phosphorimidazolide. Science 1968, 161, 387–398. [Google Scholar] [CrossRef] [PubMed]
- Röthlingshöfer, M.; Richert, C. Chemical primer extension at submillimolar concentration of deoxynucleotides. J. Org. Chem. 2010, 75, 3945–3952. [Google Scholar] [CrossRef] [PubMed]
- Jauker, M.; Griesser, H.; Richert, C. Copying of RNA sequences without pre-activation. Angew. Chem. Int. Ed. 2015, 54, 14559–14563. [Google Scholar] [CrossRef] [PubMed]
- Stütz, J.A.R.; Kervio, E.; Deck, C.; Richert, C. Chemical primer extension: Individual steps of spontaneous replication. Chem. Biodivers. 2007, 4, 784–802. [Google Scholar] [CrossRef] [PubMed]
- Prabahar, K.J.; Ferris, J.P. Effect of phosphate activating group on oligonucleotide formation on montmorillonite: The regioselective formation of 3′,5′-linked oligoadenylates. J. Am. Chem. Soc 1994, 116, 10914–10920. [Google Scholar] [CrossRef] [PubMed]
- Lohrmann, R.; Orgel, L.E. Prebiotic activation processes. Nature 1973, 244, 418–420. [Google Scholar] [CrossRef] [PubMed]
- Ibanez, J.D.; Kimball, A.P.; Oro, J. Possible prebiotic condensation of mononucleotides by cyanamide. Science 1971, 173, 444–446. [Google Scholar] [CrossRef] [PubMed]
- DOlinnaya, N.G.; Blumenfeld, M.; Merenkova, I.N.; Oretskaya, T.S.; Krynetskaya, N.; Ivanovskaya, M.G.; Vasseur, M.; Shabarova, Z.A. Oligonucleotide circularization by template-directed chemical ligation. Nucleic Acid Res. 1993, 21, 5403–5407. [Google Scholar] [CrossRef]
- Dolinnaya, N.G.; Sokolova, N.I.; Ashirbekova, D.T.; Shabarova, Z.A. The use of BrCN for assembling modified DNA duplexes and DNA-RNA hybrids; comparison with water-soluble carbodiimide. Nucleic Acid Res. 1991, 19, 3067–3072. [Google Scholar] [CrossRef]
- Sutherland, J.D. The origin of life—Out of the blue. Angew. Chem. Int. Ed. 2016, 55, 104–121. [Google Scholar] [CrossRef] [PubMed]
- Pino, S.; Sponer, J.; Costanzo, G.; Saladino, R.; Mauro, E. From formamide to RNA, the path is tenuous but continuous. Life 2015, 5, 372–384. [Google Scholar] [CrossRef] [PubMed]
- Morasch, M.; Mast, C.B.; Langer, J.K.; Schilcher, P.; Braun, D. Dry polymerization of 3′,5′-cyclic GMP to long strands of RNA. ChemBioChem 2014, 15, 879–883. [Google Scholar] [CrossRef] [PubMed]
- Goldberg, R.N.; Tewari, Y.B. Thermodynamics of the hydrolysis reactions of adenosine 3′,5′-(cyclic)phosphate(aq) and phosphoenolpyruvate(aq); the standard molar formation properties of 3′,5′-(cyclic)phosphate(aq) and phosphoenolpyruvate(aq). J. Chem. Thermodyn. 2003, 35, 1809–1830. [Google Scholar] [CrossRef]
- Crothers, D.M.; Zimm, B.H. Theory of the melting transition of synthetic polynucleotides: Evaluation of the stacking free energy. J. Mol. Biol. 1964, 9, 1–9. [Google Scholar] [CrossRef]
- Sawai, H. Catalysis of internucleotide bond formation by divalent metal ions. J. Am. Chem. Soc. 1976, 98, 7037–7039. [Google Scholar] [CrossRef] [PubMed]
- Sigel, R.K.O.; Sigel, H. A stability concept for metal ion coordination to single-stranded nucleic acids and affinities of individual sites. Acc. Chem. Res. 2010, 43, 974–984. [Google Scholar] [CrossRef] [PubMed]
- Inoue, T.; Orgel, L.E. Oligomerization of (guanosine 5′-phosphor)-2-methylimidazolide on poly(c). J. Mol. Biol. 1982, 162, 201–218. [Google Scholar] [CrossRef]
- Kanavarioti, A.; Lee, L.F.; Gangopadhyay, S. Relative reactivity of ribosyl 2′-oh vs. 3′-oh in concentrated aqueous solutions of phosphoimidazolide activated nucleotides. Orig. Life Evol. Biosphere 1999, 29, 473–487. [Google Scholar] [CrossRef] [PubMed]
- Kanavarioti, A. Dimerization in highly concentrated solutions of phospho-imidazolide activated mononucleotides. Orig. Life Evol. Biosphere 1997, 27, 357–376. [Google Scholar] [CrossRef] [PubMed]
- Tapiero, C.M.; Nagyvary, J. Prebiotic formation of cytidine nucleotides. Nature 1971, 231, 42–43. [Google Scholar] [CrossRef] [PubMed]
- Verlander, M.S.; Lohrmann, R.; Orgel, L.E. Catalysts for the self-polymerization of adenosine cyclic 2′,3′-phosphate. J. Mol. Evol. 1973, 2, 303–316. [Google Scholar] [CrossRef] [PubMed]
- Kanavarioti, A.; Bernssconi, C.F.; Alberas, D.J.; Baird, E.E. Kinetic dissection of individual steps in the poly(c)-directed oligoguanylate synthesis from guanosine 5′-phospho-2-methylimidazolide. J. Am. Chem. Soc. 1993, 115, 8537–8546. [Google Scholar] [CrossRef] [PubMed]
- Kanavarioti, A. Template-directed chemistry and the origins of the RNA world. Orig. Life Evol. Biosphere 1994, 24, 479–494. [Google Scholar] [CrossRef]
- Schrum, J.P.; Ricardo, A.; Krishnamurthy, M.; Blain, J.C.; Szostak, J.W. Efficient and rapid template-directed nucleic acid copying using 2′-amino-2′,3′-dideoxyribonucleoside-5′-phosphorimidazolide monomers. J. Am. Chem. Soc. 2009, 131, 14560–14570. [Google Scholar] [CrossRef] [PubMed]
- Vogel, S.R.; Deck, C.; Richert, C. Accelerating chemical replication steps of RNA involving activated ribonucleotides and downstream-binding elements. Chem. Commun. 2005, 39, 4922–4924. [Google Scholar] [CrossRef] [PubMed]
- Varani, G. Exceptionally stable nucleic acid hairpins. Annu. Rev. Biophys. Biomol. Struct. 1995, 24, 379–404. [Google Scholar] [CrossRef] [PubMed]
- Sawai, H. Oligoadenylates formation on an oligouridylate template in the presence of a lead catalyst. J. Mol. Evol. 1981, 17, 48–51. [Google Scholar] [CrossRef]
- Fakhrai, H.; van Roode, J.H.G.; Orgel, L.E. Synthesis of oligoguanylates on oligocytidylate templates. J. Mol. Evol. 1981, 17, 295–302. [Google Scholar] [CrossRef] [PubMed]
- Wu, T.; Orgel, L.E. Nonenzymatic template-directed synthesis on hairpin oligonucleotides. 3. Incorporation of adenosine and uridine residues. J. Am. Chem. Soc. 1992, 114, 7963–7969. [Google Scholar] [CrossRef] [PubMed]
- Wu, T.; Orgel, L.E. Nonenzymatic template-directed synthesis on oligodeoxycytidylate sequences in hairpin oligonucleotides. J. Am. Chem. Soc. 1992, 114, 317–322. [Google Scholar] [CrossRef] [PubMed]
- Lohrmann, R.; Orgel, L.E. Efficient catalysis of polycytidylic acid directed oligoguanylate formation by Pb2+. J. Mol. Biol. 1980, 142, 555–567. [Google Scholar] [CrossRef]
- Lohrmann, R.; Bridson, P.K.; Orgel, L.E. Efficient metal-ion catalyzed template-directed oligonucleotide synthesis. Science 1980, 208, 1464–1465. [Google Scholar] [CrossRef] [PubMed]
- Inoue, T.; Orgel, L.E. Substituent control of the poly(c)-directed oligomerization of guanosine 5′-phosphorimidazolide. J. Am. Chem. Soc. 1981, 103, 7666–7667. [Google Scholar] [CrossRef]
- Sawai, H.; Totuka, S.; Yamamoto, H. Helical structure formation between complementary oligonucleotides. Minimum chain length required for the template-directed synthesis of oligonucleotides. Orig. Life Evol. Biosphere 1997, 27, 525–533. [Google Scholar] [CrossRef] [PubMed]
- Joyce, G.F.; Orgel, L.E. Non-enzymatic template-directed synthesis on RNA random copolymers. Poly(c,a) templates. J. Mol. Biol. 1988, 202, 677–681. [Google Scholar] [CrossRef]
- Chaput, J.C.; Sinha, S.; Switzer, C. 5-propynyluracil.Diaminopurine: An efficient base-pair for non-enzymatic transcription of DNA. Chem. Commun. 2002, 1568–1569. [Google Scholar] [CrossRef]
- Sawai, H.; Wada, M. Nonenzymatic template-directed condensation of short-chained oligouridylates on poly(a) template. Orig. Life Evol. Biosphere 2000, 30, 503–511. [Google Scholar] [CrossRef] [PubMed]
- Hill, A.R., Jr.; Orgel, L.E. The limits of template-directed synthesis with nucleoside-5′-phosphoro(2-methyl)imidazole. Orig. Life Evol. Biosphere 1993, 23, 285–290. [Google Scholar] [CrossRef] [PubMed]
- Hey, M.; Hartel, C.; Göbel, M.W. Nonenzymatic oligomerization of ribonucleotides: Towards in vitro selection experiments. Helv. Chim. Acta 2003, 86, 844–854. [Google Scholar] [CrossRef]
- Hud, N.V.; Cafferty, B.J.; Krishnamurthy, R.; Williams, L.D. The origin of RNA and “my grandfather’s axe”. Chem. Biol. 2013, 20, 466–474. [Google Scholar] [CrossRef] [PubMed]
- Heuberger, B.D.; Pal, A.; Del Frate, F.; Topkar, V.V.; Szostak, J.W. Replacing uridine with 2-thiouridine enhances the rate and fidelity of nonenzymatic RNA primer extension. J. Am. Chem. Soc. 2015, 137, 2769–2775. [Google Scholar] [CrossRef] [PubMed]
- Zielinski, W.S.; Orgel, L.E. Oligomerization of activated derivatives of 3′-amino-3′-deoxyguanosine on poly(c) and poly(dc) templates. Nucleic Acid Res. 1985, 13, 2469–2484. [Google Scholar] [CrossRef]
- Hagenbuch, P.; Kervio, E.; Hochgesand, A.; Plutowski, U.; Richert, C. Chemical primer extension: Efficiently determining single nucleotides in DNA. Angew. Chem. Int. Ed. Engl. 2005, 44, 6588–6592. [Google Scholar] [CrossRef] [PubMed]
- Rajamani, S.; Ichida, J.K.; Antal, T.; Treco, D.A.; Leu, K.; Nowak, M.A.; Szostak, J.W.; Chen, I.A. Effect of stalling after mismatches on the error catastrophe in nonenzymatic nucleic acid replication. J. Am. Chem. Soc. 2010, 132, 5880–5885. [Google Scholar] [PubMed]
- Leu, K.; Kervio, E.; Obermayer, B.; Turk-MacLeod, R.M.; Yuan, C.; Luevano, J.-M.; Chen, E.; Gerland, U.; Richert, C.; Chen, I.A. Cascade of reduced speed and accuracy after errors in enzyme-free copying of nucleic acid sequences. J. Am. Chem. Soc. 2013, 135, 354–366. [Google Scholar] [CrossRef] [PubMed]
- Bernal, J.D. The Physical Basis of Life; Routledge and Paul Publisher: London, UK, 1951. [Google Scholar]
- Patel, B.H.; Percivalle, C.; Ritson, D.J.; DuffyColm, D.; Sutherland, J.D. Common origins of RNA, protein and lipid precursors in a cyanosulfidic protometabolism. Nat. Chem. 2015, 7, 301–307. [Google Scholar] [CrossRef] [PubMed]
- Ferris, J.P. Montmorillonite-catalysed formation of RNA oligomers: The possible role of catalysis in the origins of life. Philos. Trans. R. Soc. Lond. B Biol. Sci. 2006, 361, 1777–1786. [Google Scholar] [CrossRef] [PubMed]
- Martin, W.; Baross, J.; Kelley, D.; Russell, M.J. Hydrothermal vents and the origin of life. Nat. Rev. Microbiol. 2008, 6, 805–814. [Google Scholar] [CrossRef] [PubMed]
- Baaske, P.; Weinert, F.M.; Duhr, S.; Lemke, K.H.; Russell, M.J.; Braun, D. Extreme accumulation of nucleotides in simulated hydrothermal pore systems. Proc. Natl. Acad. Sci. USA 2007, 104, 9346–9351. [Google Scholar] [CrossRef] [PubMed]
- Mast, C.B.; Schink, S.; Gerland, U.; Braun, D. Escalation of polymerization in a thermal gradient. Proc. Natl. Acad. Sci. USA 2013, 110, 8030–8035. [Google Scholar] [CrossRef] [PubMed]
- Kanavarioti, A.; Monnard, P.-A.; Deamer, D.W. Eutectic phases in ice facilitate non-enzymatic nucleic acid synthesis. Astrobiology 2001, 1, 271–281. [Google Scholar] [CrossRef] [PubMed]
- Attwater, J.; Wochner, A.; Pinheiro, V.B.; Coulson, A.; Holliger, P. Ice as a protocellular medium for RNA replication. Nat. Commun. 2010, 1, 76. [Google Scholar] [CrossRef] [PubMed]
- Damer, B.; Deamer, D. Coupled phases and combinatorial selection in fluctuating hydrothermal pools: A scenario to guide experimental approaches to the origin of cellular life. Life 2015, 5, 872. [Google Scholar] [CrossRef] [PubMed]
- Georgelin, T.; Jaber, M.; Fournier, F.; Laurent, G.; Costa-Torro, F.; Maurel, M.-C.; Lambert, J.-F. Stabilization of ribofuranose by a mineral surface. Carbohydr. Res. 2015, 402, 241–244. [Google Scholar] [CrossRef] [PubMed]
- Ricardo, A.; Carrigan, M.A.; Olcott, A.N.; Benner, S.A. Borate minerals stabilize ribose. Science 2004, 303, 196. [Google Scholar] [CrossRef] [PubMed]
- Ferris, J.P.; Hill, A.R., Jr.; Liu, R.; Orgel, L.E. Synthesis of long prebiotic oligomers on mineral surfaces. Nature 1996, 381, 59–61. [Google Scholar] [CrossRef] [PubMed]
- Huang, W.; Ferris, J.P. One-step, regioselective synthesis of up to 50-mer of RNA oligomers by montmorillonite catalysis. J. Am. Chem. Soc. 2006, 128, 8914–8919. [Google Scholar] [CrossRef] [PubMed]
- Miyakawa, S.; Ferris, J.P. Sequence- and regioselectivity in the montmorillonite-catalyzed synthesis of RNA. J. Am. Chem. Soc. 2003, 125, 8202–8208. [Google Scholar] [CrossRef] [PubMed]
- Franchi, M.; Ferris, J.P.; Gallori, E. Cations as mediators of the adsorption of nucleic acids on clay surfaces in prebiotic environments. Orig. Life Evol. Biosphere 2003, 33, 1–16. [Google Scholar] [CrossRef] [PubMed]
- Aldersley, M.F.; Joshi, P.C.; Price, J.D.; Ferris, J.P. The role of montmorillonite in its catalysis of RNA synthesis. Appl. Clay Sci. 2011, 54, 1–14. [Google Scholar] [CrossRef]
- Gibbs, D.; Lohrmann, R.; Orgel, L.E. Template-directed synthesis and selective absorption of oligoadenylates on hydroxyapatite. J. Mol. Evol. 1980, 15, 347–354. [Google Scholar] [CrossRef] [PubMed]
- Acevedo, O.L.; Orgel, L.E. Template-directed oligonucleotide ligation on hydroxylapatite. Nature 1986, 321, 790–792. [Google Scholar] [CrossRef] [PubMed]
- Schwartz, A.W.; Orgel, L.E. Template-directed polynucleotide synthesis on mineral surfaces. J. Mol. Evol. 1985, 21, 299–300. [Google Scholar] [CrossRef]
- Ertem, G.; Ferris, J.P. Synthesis of RNA oligomers on heterogeneous templates. Nature 1996, 379, 238–240. [Google Scholar] [CrossRef] [PubMed]
- Budin, I.; Bruckner, R.J.; Szostak, J.W. Formation of protocell-like vesicles in a thermal diffusion column. J. Am. Chem. Soc. 2009, 131, 9628–9629. [Google Scholar] [CrossRef] [PubMed]
- Keil, L.; Hartmann, M.; Lanzmich, S.; Braun, D. Probing of molecular replication and accumulation in shallow heat gradients through numerical simulations. Phys. Chem. Chem. Phys. 2016, 18, 20153–20159. [Google Scholar] [CrossRef] [PubMed]
- Mast, C.B.; Braun, D. Thermal trap for DNA replication. Phys. Rev. Lett. 2010, 104, 188102. [Google Scholar] [CrossRef] [PubMed]
- Da Silva, L.; Maurel, M.-C.; Deamer, D. Salt-promoted synthesis of RNA-like molecules in simulated hydrothermal conditions. J. Mol. Evol. 2015, 80, 86–97. [Google Scholar] [CrossRef] [PubMed]
- Bada, J.L.; Bigham, C.; Miller, S.L. Impact melting of frozen oceans on the early earth: Implications for the origin of life. Proc. Natl. Acad. Sci. USA 1994, 91, 1248–1250. [Google Scholar] [CrossRef] [PubMed]
- Arndt, N.T.; Nisbet, E.G. Processes on the young earth and the habitats of early life. Annu. Rev. Earth Planet. Sci. 2012, 40, 521–549. [Google Scholar] [CrossRef]
- Mulkidjanian, A.Y.; Bychkov, A.Y.; Dibrova, D.V.; Galperin, M.Y.; Koonin, E.V. Origin of first cells at terrestrial, anoxic geothermal fields. Proc. Natl. Acad. Sci. USA 2012, 109, E821–E830. [Google Scholar] [CrossRef] [PubMed]
- Cocks, F.H.; Brower, W.E. Phase diagram relationships in cryobiology. Cryobiology 1974, 11, 340–358. [Google Scholar] [CrossRef]
- Monnard, P.-A.; Kanavarioti, A.; Deamer, D.W. Eutectic phase polymerization of activated ribonucleotide mixtures yields quasi-equimolar incorporation of purine and pyrimidine nucleobases. J. Am. Chem. Soc. 2003, 125, 13734–13740. [Google Scholar] [CrossRef] [PubMed]
- Wieczorek, R.; Dörr, M.; Chotera, A.; Luisi, P.L.; Monnard, P.-A. A dipeptide can catalyze the formation of phosphodiester bonds: A possible origin of protein catalysis. Chembiochem 2013, 14, 217–223. [Google Scholar] [CrossRef] [PubMed]
- Monnard, P.-A.; Szostak, J.W. Metal-ion catalyzed polymerization in the eutectic phase in water-ice: A possible approach to template-directed RNA polymerization. J. Inorg. Biochem. 2008, 102, 1104–1111. [Google Scholar] [CrossRef] [PubMed]
- Löffler, P.M.G.; Gronen, J.; Dörr, M.; Monnard, P.-A. Sliding over the barriers in enzyme-free RNA copying—One-pot primer extension in ice. PLoS ONE 2013, 8, e75617. [Google Scholar] [CrossRef] [PubMed]
- Monnard, P.-A.; Apel, C.L.; Kanavarioti, A.; Deamer, D.W. Influence of ionic solutes on self-assembly and polymerization processes related to early forms of life: Implications for a prebiotic aqueous medium. Astrobiology 2002, 2, 139–152. [Google Scholar] [CrossRef] [PubMed]
- Vlassov, A.; Johnston, B.H.; Landweber, L.F.; Kazakov, S.A. Ligation activity of fragmented ribozymes in frozen solution: Implication for the RNA world. Nucleic Acids. Res. 2004, 32, 2966–2974. [Google Scholar] [CrossRef] [PubMed]
- Lie, L.; Biliya, S.; Vannberg, F.; Wartell, R.M. Ligation of RNA oligomers by the schistosoma mansoni hammerhead ribozyme in frozen solution. J. Mol. Evol. 2016, 82, 81–92. [Google Scholar] [CrossRef] [PubMed]
- Attwater, J.; Wochner, A.; Holliger, P. In-ice evolution of RNA polymerase ribozyme activity. Nat. Chem. 2013, 5, 1011–1018. [Google Scholar] [CrossRef] [PubMed]
- Mutschler, H.; Wochner, A.; Holliger, P. Freeze–Thaw cycles as drivers of complex ribozyme assembly. Nat. Chem. 2015, 7, 502–508. [Google Scholar] [CrossRef] [PubMed]
- Syren, R.M.; Sanjur, A.; Fox, S.W. Proteinoid microspheres more stable in hot than in cold water. Biosystems 1985, 17, 275–280. [Google Scholar] [CrossRef]
- Weber, A.L. Growth of organic microspherules in sugar-ammonia reactions. Orig. Life Evol. Biosphere 2005, 35, 523–536. [Google Scholar] [CrossRef] [PubMed]
- Simoneit, B.R.T.; Rushdi, A.I.; Deamer, D.W. Abiotic formation of acylglycerols under simulated hydrothermal conditions and self-assembly properties of such lipid products. Adv. Space Res. 2007, 40, 1649–1656. [Google Scholar] [CrossRef]
- Toppozini, L.; Dies, H.; Deamer, D.W.; Rheinstädter, M.C. Adenosine monophosphate forms ordered arrays in multilamellar lipid matrices: Insights into assembly of nucleic acid for primitive life. PLoS ONE 2013, 8, e62810. [Google Scholar] [CrossRef] [PubMed]
- Kawamura, K. Behaviour of RNA under hydrothermal conditions and the origins of life. Int. J. Astrobiol. 2004, 3, 301–309. [Google Scholar] [CrossRef]
- Rajamani, S.; Vlassov, A.; Coombs, A.; Olasagasti, F.; Deamer, D.W. Lipid-assisted synthesis of RNA-like polymers from mononucleotides. Orig. Life Evol. Biosphere 2008, 38, 57–74. [Google Scholar] [CrossRef] [PubMed]
- Olasagasti, F.; Kim, H.J.; Pourmand, N.; Deamer, D.W. Non-enzymatic transfer of sequence information under plausible prebiotic conditions. Biochimie 2011, 93, 556–561. [Google Scholar] [CrossRef] [PubMed]
- Mungi, C.; Rajamani, S. Characterization of RNA-like oligomers from lipid-assisted nonenzymatic synthesis: Implications for origin of informational molecules on early earth. Life 2015, 5, 65–84. [Google Scholar] [CrossRef] [PubMed]
- DeGuzman, V.; Vercoutere, W.; Shenasa, H.; Deamer, D. Generation of oligonucleotides under hydrothermal conditions by non-enzymatic polymerization. J. Mol. Evol. 2014, 78, 251–262. [Google Scholar] [CrossRef] [PubMed]
- Monnard, P.-A.; Walde, P. Current ideas about prebiological compartmentalization. Life 2015, 5, 1239–1263. [Google Scholar] [CrossRef] [PubMed]
- Black, R.A.; Blosser, M.C.; Stottrup, B.L.; Tavakley, R.; Deamer, D.W.; Keller, S.L. Nucleobases bind to and stabilize aggregates of a prebiotic amphiphile, providing a viable mechanism for the emergence of protocells. Proc. Natl. Acad. Sci. USA 2013, 110, 13272–13276. [Google Scholar] [CrossRef] [PubMed]
- Hud, N.V.; Anet, F.A.L. Intercalation-mediated synthesis and replication: A new approach to the origin of life. J. Theor. Biol. 2000, 205, 543–562. [Google Scholar] [CrossRef] [PubMed]

© 2016 by the author; licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC-BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Monnard, P.-A. Taming Prebiotic Chemistry: The Role of Heterogeneous and Interfacial Catalysis in the Emergence of a Prebiotic Catalytic/Information Polymer System. Life 2016, 6, 40. https://doi.org/10.3390/life6040040
Monnard P-A. Taming Prebiotic Chemistry: The Role of Heterogeneous and Interfacial Catalysis in the Emergence of a Prebiotic Catalytic/Information Polymer System. Life. 2016; 6(4):40. https://doi.org/10.3390/life6040040
Chicago/Turabian StyleMonnard, Pierre-Alain. 2016. "Taming Prebiotic Chemistry: The Role of Heterogeneous and Interfacial Catalysis in the Emergence of a Prebiotic Catalytic/Information Polymer System" Life 6, no. 4: 40. https://doi.org/10.3390/life6040040
APA StyleMonnard, P.-A. (2016). Taming Prebiotic Chemistry: The Role of Heterogeneous and Interfacial Catalysis in the Emergence of a Prebiotic Catalytic/Information Polymer System. Life, 6(4), 40. https://doi.org/10.3390/life6040040