Mitigating Harmful Cyanobacterial Blooms in a Human- and Climatically-Impacted World


Abstract
:1. Introduction


{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Toxin | Detection Method(s) * | CyanoHAB Genera |
|---|---|---|
| Aeruginosin | HPLC, MS | Microcystis, Planktothrix |
| Anatoxin-a /Homoanatoxin-a | ELISA, HPLC, MS | Anabaena, Aphanizomenon, Cylindrospermopsis, Lyngbya, Oscillatoria, Phormidium, Planktothrix, Raphidiopsis, Woronichinia |
| Anatoxin-a(S) | AEIA, MS | Anabaena |
| Aplysiatoxins | MS | Lyngbya, Oscillatoria, Schizothrix |
| beta-Methylamino-L-alanine (BMAA) | ELISA, HPLC, MS | Anabaena, Aphanizomenon, Calothrix, Cylindrospermopsis, Lyngbya, Microcystis, Nostoc, Nodularia, Planktothrix, Phormidium, Prochlorococcus, Scytonema, Synechococcus, Trichodesmium |
| Cyanopeptolin | HPLC, MS | Anabaena, Microcystis, Planktothrix |
| Cylindrospermopsin | ELISA, HPLC, MS | Anabaena, Aphanizomenon, Cylindrospermopsis, Oscillatoria, Raphidiopsis, Umezakia |
| Jamaicamides | MS | Lyngbya |
| Lyngbyatoxin | HPLC, MS | Lyngbya |
| Microcystin | ELISA, HPLC, MS, PPIA | Anabaena, Anabaenopsis, Aphanizomenon, Aphanocapsa, Cylindrospermopsis, Gloeotrichia, Hapalosiphon, Microcystis, Nostoc, Oscillatoria, Phormidium, Planktothrix, Pseudoanabaena, Synechococcus, Woronochinia |
| Nodularin | ELISA, HPLC, MS, PPIA | Nodularia |
| Saxitoxin | ELISA, HPLC, MS | Anabaena, Aphanizomenon, Cylindrospermopsis, Lyngbya, Oscillatoria, Planktothrix |
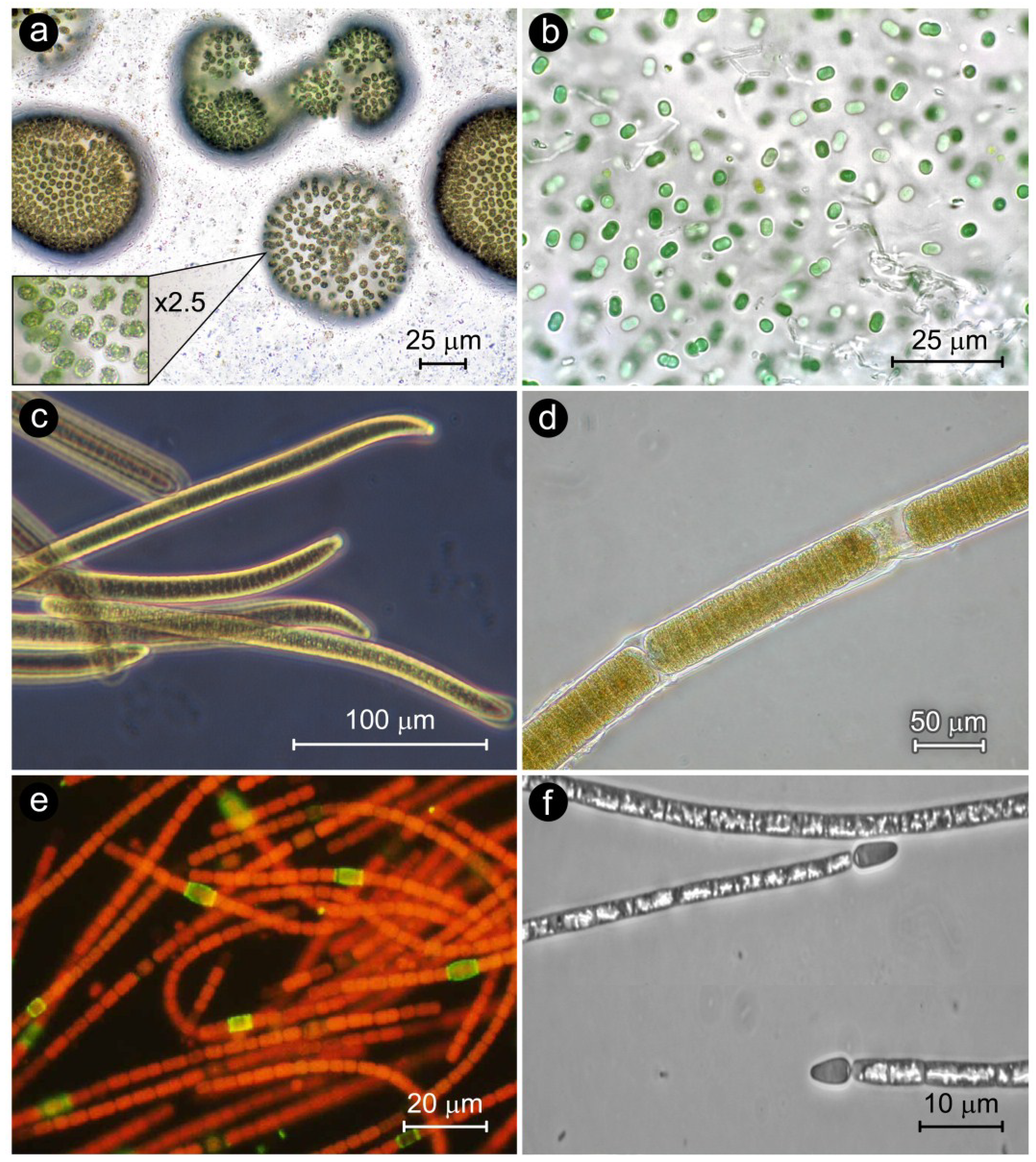
2. Cyanobacterial Bloom Taxa
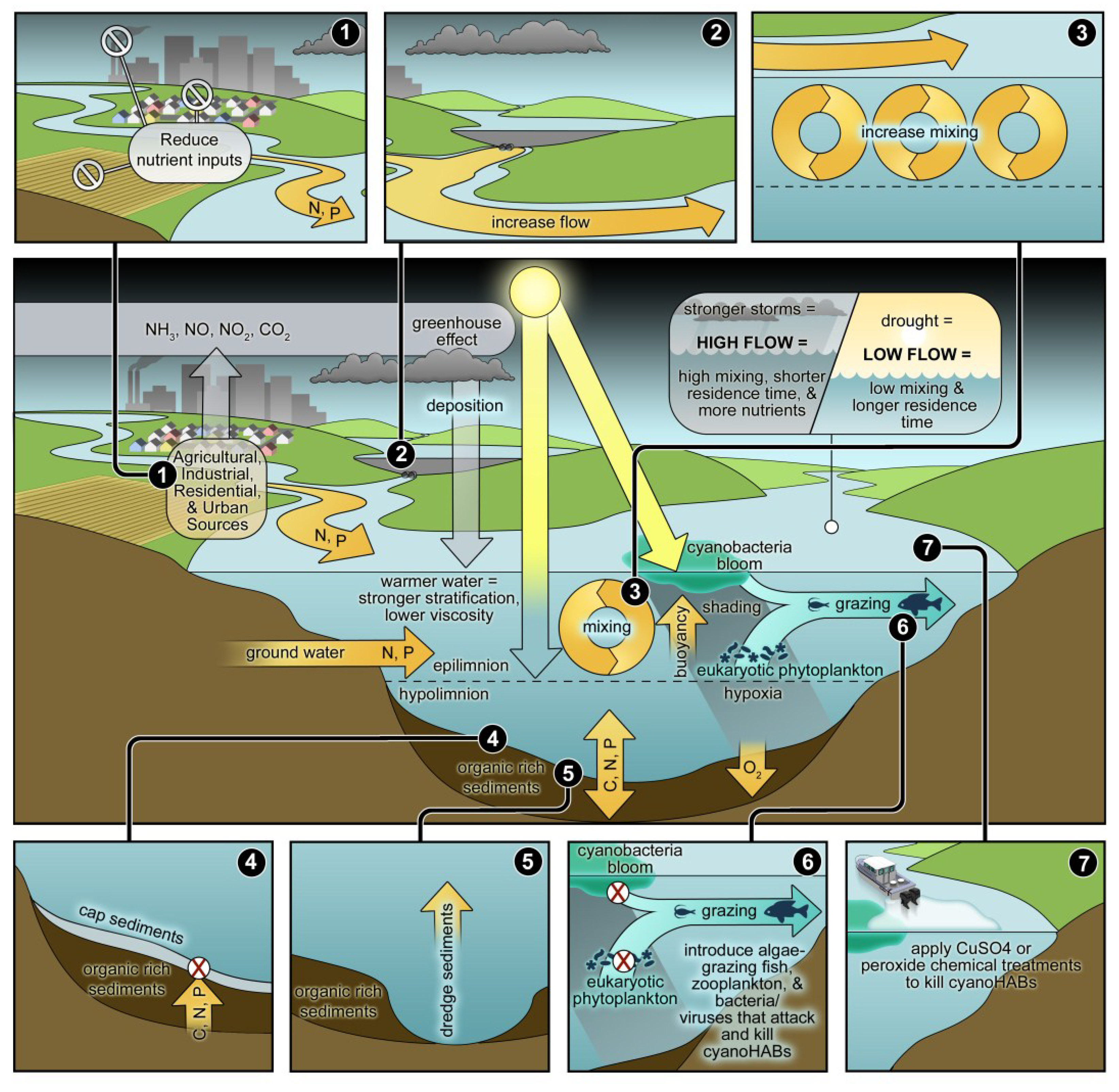
3. Managing Cyanobacterial Blooms in a Human and Climatically-Altered World
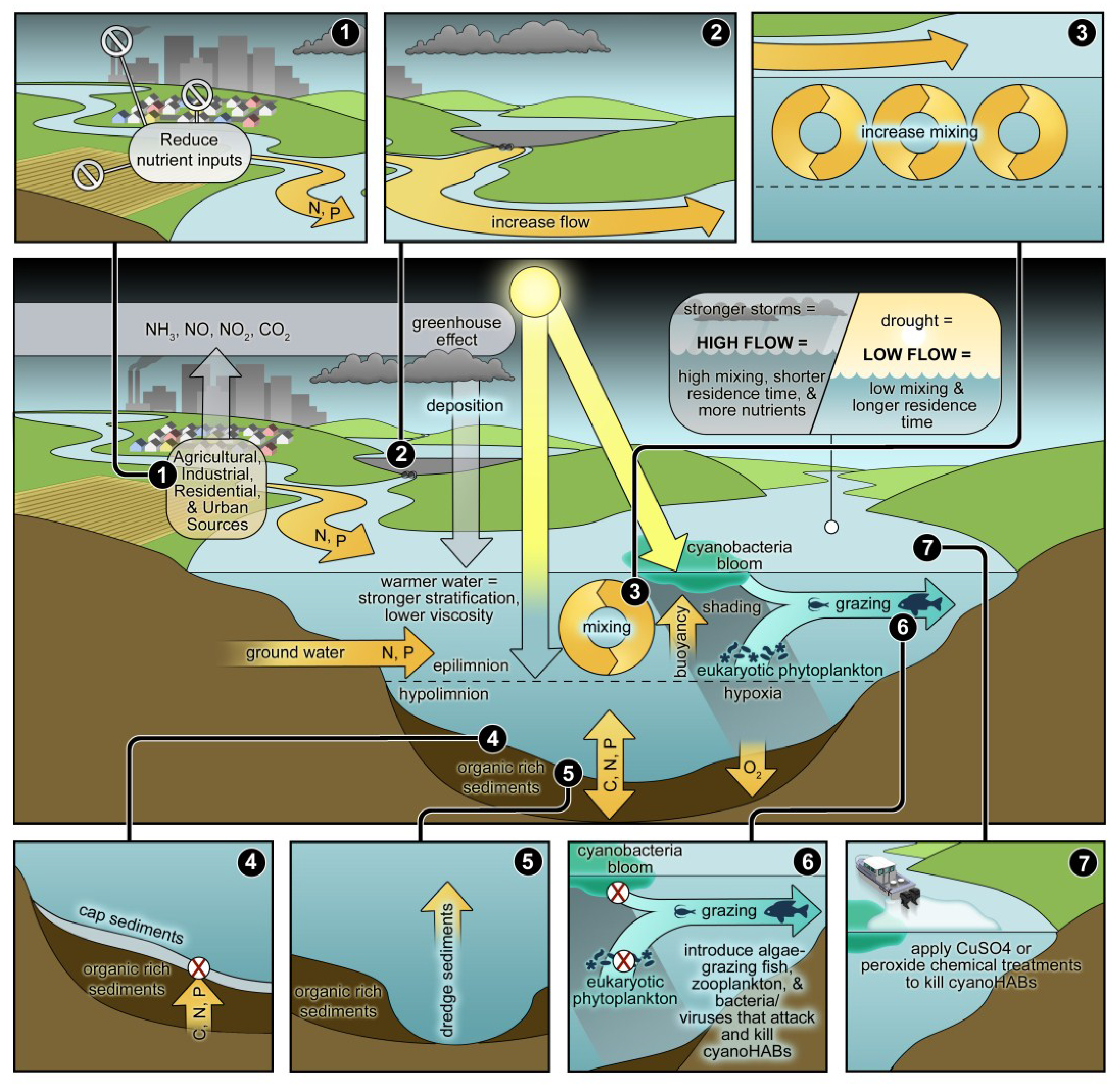
3.1. Nutrient Input Controls
3.2. Phosphorus Management
3.3. Nitrogen Management
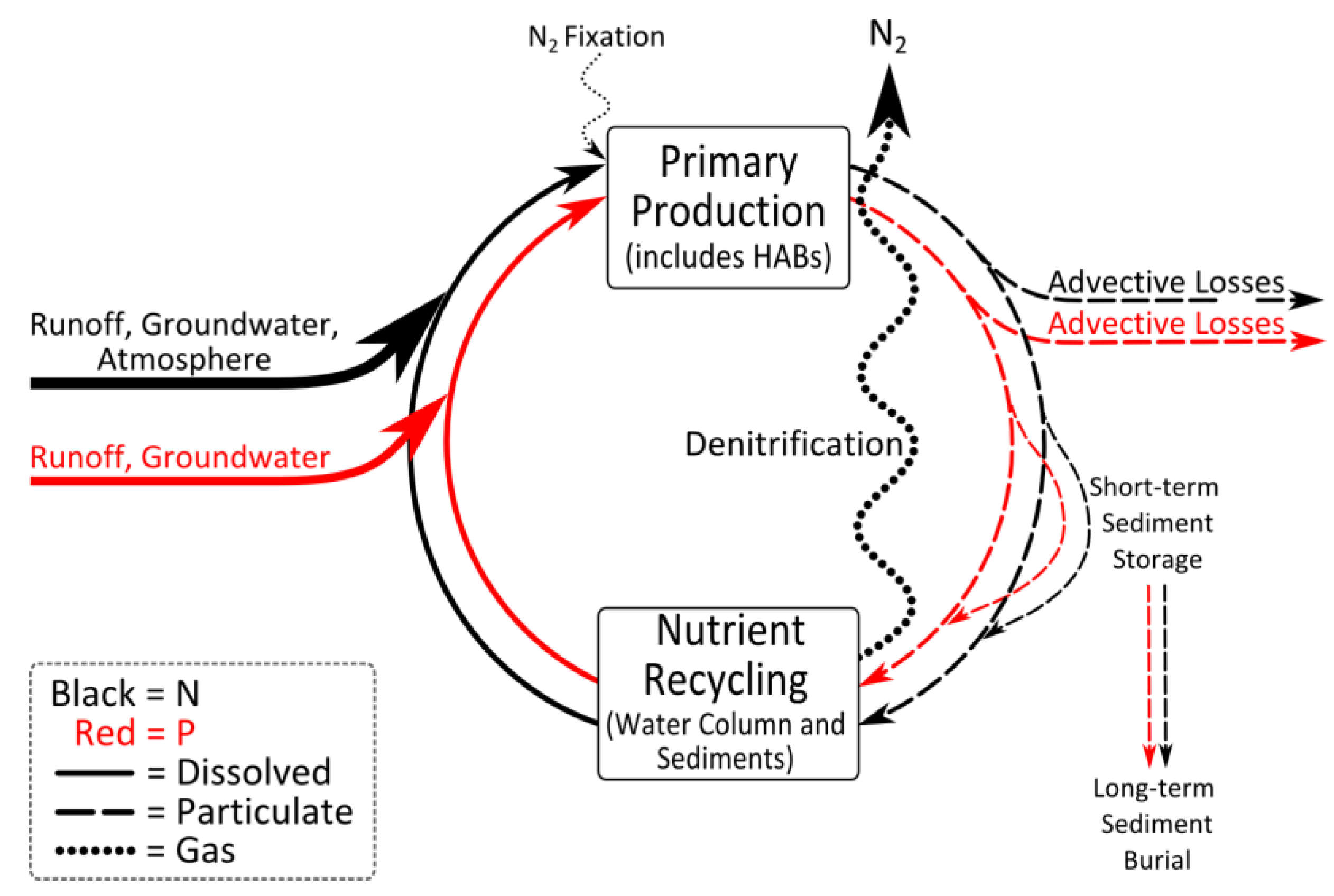
3.4. Rationale for Dual Nutrient (N and P) Controls
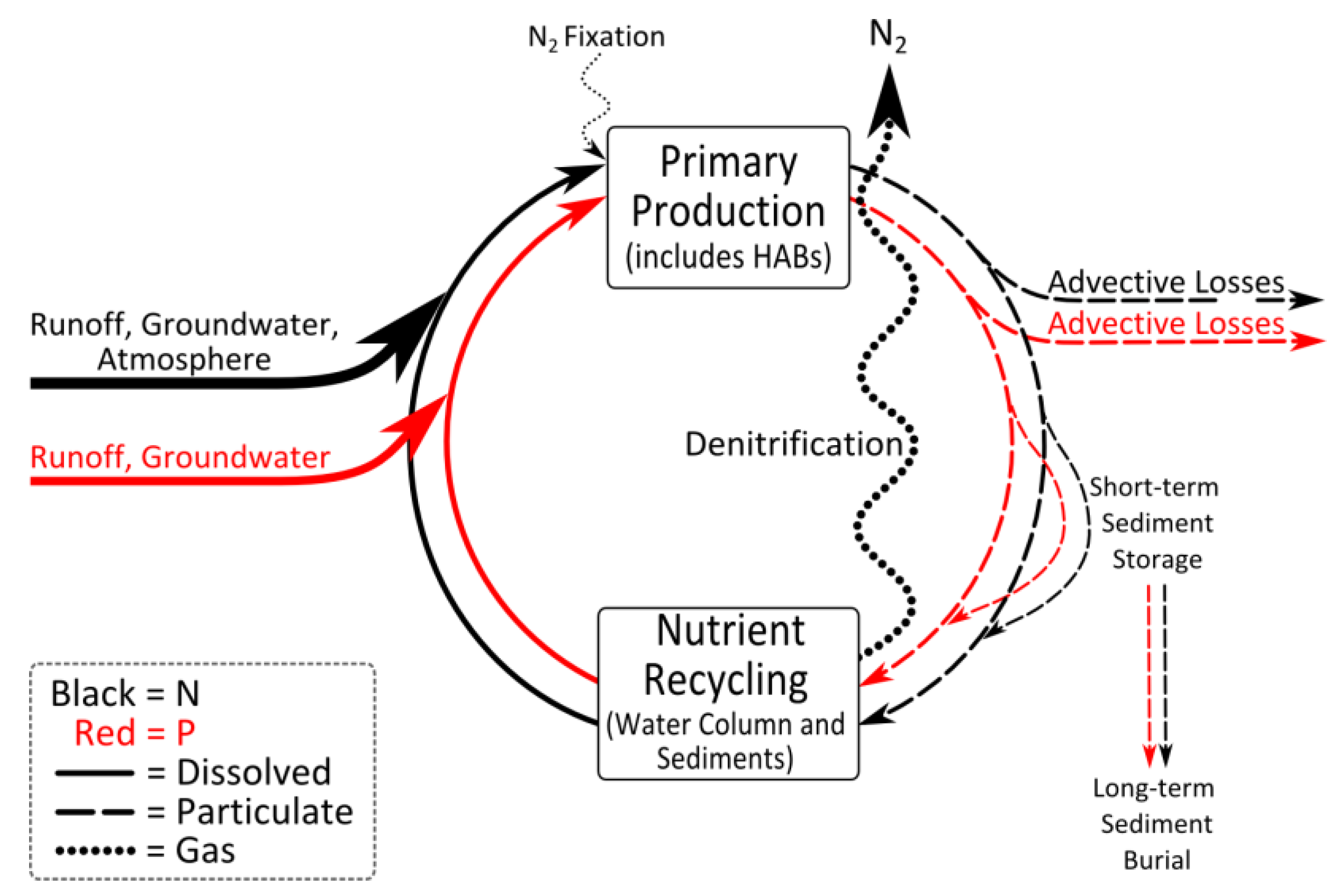
4. The Interactions of Physical, Biological and Nutrient Controls of Cyanobacteria in a Climatically-Changing World
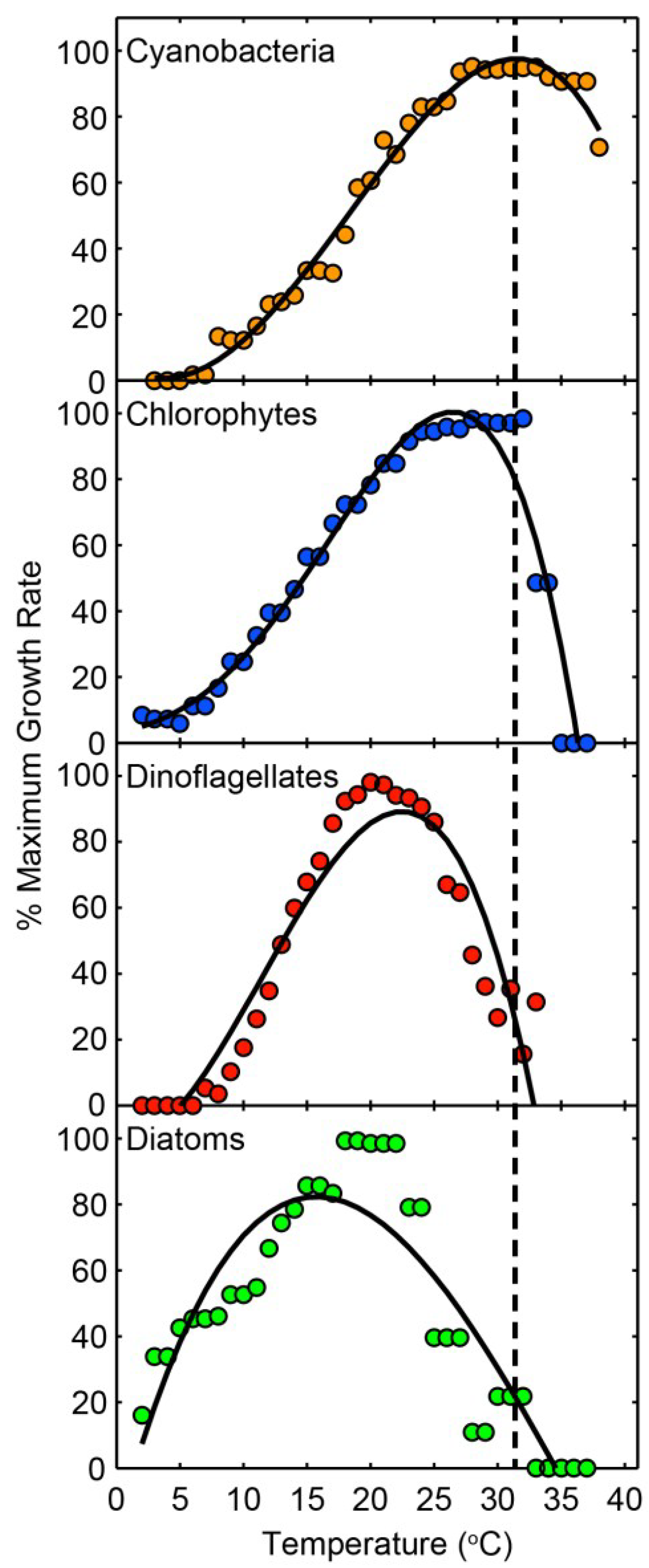
4.1. Climate Change: Its Role in Modulating CyanoHABs
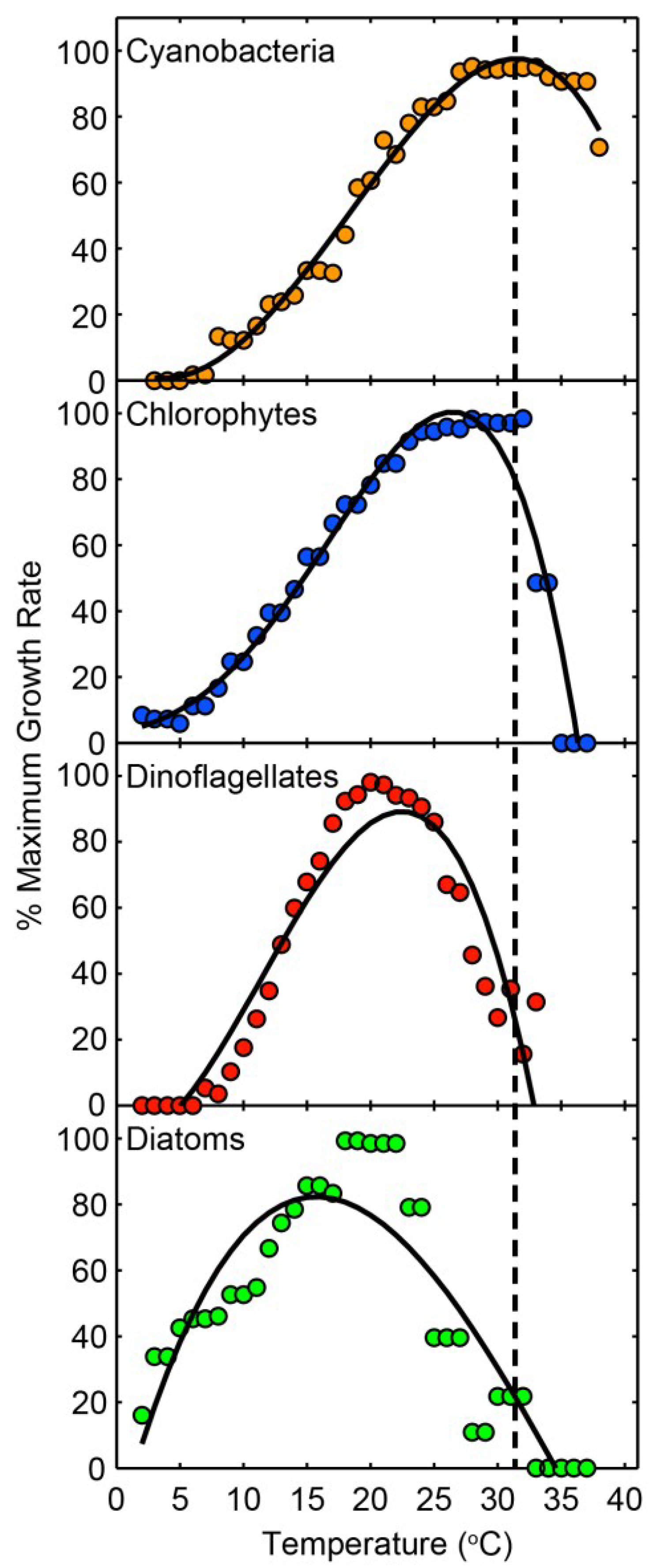
4.2. The Role of Physical Factors

4.3. Non-Nutrient Chemical Controls
4.4. Manipulating Sediments
4.5. Biological Controls
5. Concluding Remarks and Recommendations
Acknowledgments
Conflicts of Interest
References
- Schopf, J.W. The fossil record: Tracing the roots of the cyanobacterial lineage. In The Ecology of Cyanobacteria; Whitton, B.A., Potts, M., Eds.; Kluwer Academic Publishers: Dordrecht, The Netherlands, 2000; pp. 13–35. [Google Scholar]
- Potts, M.; Whitton, B.A. The Biology and Ecology of Cyanobacteria; Blackwell Scientific Publications: Oxford, UK, 2000. [Google Scholar]
- Whitton, B.A. Ecology of Cyanobacteria II: Their Diversity in Space and Time; Springer: Dordrecht, The Netherlands, 2012. [Google Scholar]
- Carmichael, W.W. Health effects of toxin producing cyanobacteria: The cyanoHABs. Hum. Ecol. Risk Assess. 2001, 7, 1393–1407. [Google Scholar] [CrossRef]
- Paerl, H.W.; Otten, T.G. Harmful Cyanobacterial Blooms: Causes, Consequences and Controls. Microb. Ecol. 2013, 65, 995–1010. [Google Scholar] [CrossRef] [PubMed]
- Reynolds, C.S. Ecology of Phytoplankton (Ecology, Biodiversity and Conservation); Cambridge University Press: Cambridge, UK, 2006. [Google Scholar]
- Gallon, J.R. Tansley Review No. 44/Reconciling the incompatible: N2 fixation and O2. New Phytol. 1992, 122, 571–609. [Google Scholar] [CrossRef]
- Wilhelm, S.W.; Trick, C.G. Iron-limited growth of cyanobacteria: Multiple siderophore production is a common response. Limnol. Oceanogr. 1994, 39, 1979–1984. [Google Scholar] [CrossRef]
- Healy, F.P. Phosphate. In The Biology of Cyanobacteria; Carr, N.G., Whitton, B.A., Eds.; Blackwell Scientific Publications: Oxford, UK, 1982; pp. 105–124. [Google Scholar]
- Paerl, H.W.; Otten, T.G. Blooms bite the hand that feeds them. Science 2013, 342, 433–434. [Google Scholar] [CrossRef] [PubMed]
- Paerl, H.W.; Millie, D.F. Physiological ecology of toxic cyanobacteria. Phycologia 1996, 35, 160–167. [Google Scholar] [CrossRef]
- Huisman, J.; Sharples, J.; Stroom, J.M.; Visser, P.M.; Kardinaal, W.E.A.; Verspagen, J.M.H.; Sommeijer, B. Changes in turbulent mixing shift competition for light between phytoplankton species. Ecology 2004, 85, 2960–2970. [Google Scholar] [CrossRef]
- Paerl, H.W. Interactions with bacteria. In The Biology of Cyanobacteria; Carr, N.G., Whitton, B.A., Eds.; Blackwell Scientific Publications Ltd.: Oxford, UK, 1982; Chapter 17; pp. 441–461. [Google Scholar]
- Paerl, H.W. Growth and reproductive strategies of freshwater blue-green algae (cyanobacteria). In Growth and Reproductive Strategies of Freshwater Phytoplankton; Sandgren, C.D., Ed.; Cambridge University Press: Cambridge, UK, 1986. [Google Scholar]
- Kuosa, H. Picoplanktonic algae in the northern Baltic Sea: Seasonal dynamics and flagellate grazing. Mar. Ecol. Prog. Ser. 1991, 73, 269–276. [Google Scholar] [CrossRef]
- Sánchez-Baracaldo, P.; Handley, B.A.; Hayes, P.K. Picocyanobacterial community structure of freshwater lakes and the Baltic Sea revealed by phylogenetic analyses and clade-specific quantitative PCR. Microbiology 2008, 11, 3347–3357. [Google Scholar] [CrossRef]
- Gaulke, A.K.; Wetz, M.S.; Paerl, H.W. Picoplankton: A major contributor to planktonic biomass and promary production in a eutrophic, river-dominated estuary. Estuar. Coast. Shelf Sci. 2010, 90, 45–54. [Google Scholar] [CrossRef]
- Wolk, C.P. Heterocyst Formation. Annu. Rev. Genet. 1996, 30, 59–78. [Google Scholar] [CrossRef] [PubMed]
- Carmichael, W.W. The cyanotoxins. Adv. Bot. Res. 1997, 27, 211–256. [Google Scholar]
- Carmichael, W.W. Microcystin concentrations in human livers, estimation of human lethal dose-lessons from Caruaru, Brazil. In Proceedings of the 4th International Conference on Toxic Cyanobacteria, Beaufort, NC, USA, 27 September–1 October 1999.
- Paerl, H.W. Nuisance phytoplankton blooms in coastal, estuarine, and inland waters. Limnol. Oceanogr. 1988, 33, 823–847. [Google Scholar] [CrossRef]
- Smith, V.H.; Schindler, D.W. Eutrophication science: Where do we go from here? Trends Ecol. Evol. 2009, 24, 201–207. [Google Scholar] [CrossRef] [PubMed]
- Paerl, H.W.; Scott, J.T. Throwing fuel on the fire: Synergistic effects of excessive nitrogen inputs and global warming on harmful algal blooms. Environ. Sci. Technol. 2010, 44, 7756–7758. [Google Scholar] [CrossRef] [PubMed]
- Paerl, H.W. Combating the global proliferation of harmful cyanobacterial blooms by integrating conceptual and technological advances in an accessible water management toolbox. Environ. Microbiol. Rep. 2013, 5, 12–14. [Google Scholar]
- Grantz, E.M.; Haggard, B.E.; Scott, J.T. Stoichiometric imbalance in rates of nitrogen and phosphorus retention, storage, and recycling can perpetuate nitrogen deficiency in highly-productive reservoirs. Limnol. Oceanogr. 2014, 59, 2203–2216. [Google Scholar] [CrossRef]
- Scott, J.T.; McCarthy, M.J.; Otten, T.G.; Steffen, M.M.; Baker, B.C.; Grantz, E.M.; Wilhelm, S.W.; Paerl, H.W. Comment: An alternative interpretation of the relationship between TN:TP and microcystins in Canadian lakes. Can. J. Fish. Aquat. Sci. 2013, 70, 1265–1268. [Google Scholar] [CrossRef]
- Smith, V.H. Low nitrogen to phosphorus ratios favor dominance by blue-green algae in lake phytoplankton. Science 1983, 221, 669–671. [Google Scholar] [CrossRef] [PubMed]
- U.S. Environmental Protection Agency. Reactive Nitrogen in the United States: An Analysis of Inputs, Flows, Consequences, and Management Options. A Report of the EPA Science Advisory Board (EPA-SAB-11-013); Scientific Advisory Board: Washington, DC, USA, 2011. [Google Scholar]
- Sharpley, A.N.; Daniel, T.; Sims, T.; Lemunyon, T.J.; Stevens, R.; Parry, R. Agricultural Phosphorus and Eutrophication, 2nd ed.; USDA-ARS, Pasture Systems & Watershed Management Research Unit: University Park, PA, USA, 2010; p. 44. [Google Scholar]
- Wetzel, R.G. Limnology: Lake and River Ecosystems, 3rd ed.; Academic Press: San Diego, CA, USA, 2001. [Google Scholar]
- Lean, D.R.S. Movement of phosphorus between its biologically important forms in lake water. J. Fish. Res. Board Can. 1973, 30, 1525–1536. [Google Scholar] [CrossRef]
- Galloway, J.N.; Dentener, F.J.; Capone, D.G.; Boyer, E.W.; Howarth, R.W.; Seitzinger, S.P.; Asner, G.P.; Cleveland, C.C.; Green, P.A.; Holland, E.A.; et al. Nitrogen cycles: Past, present, and future. Biogeochemistry 2004, 70, 153–226. [Google Scholar] [CrossRef]
- Paerl, H.W.; Piehler, M.F. Nitrogen and Marine Eutrophication. In Nitrogen in the Marine Environment; Capone, D.G., Mulholland, M., Carpenter, E., Eds.; Academic Press: Orlando, FL, USA, 2008; Volume 2, pp. 529–567. [Google Scholar]
- Blomqvist, P.; Pettersson, A.; Hyenstrand, P. Ammonium-nitrogen: A key regulatory factor causing dominance of non-nitrogen-fixing cyanobacteria in aquatic systems. Arch. Hydrobiol. 1994, 132, 141–164. [Google Scholar]
- Hyenstrand, P.; Nyvall, P.; Pettersson, A.; Blomqvist, P. Regulation of non-nitrogen-fixing cyanobacteria by inorganic nitrogen sources—Experiments from Lake Erken. Arch. Hydrobiol. 1998, 51, 29–40. [Google Scholar]
- Galloway, J.N.; Cowling, E.B.; Seitzinger, S.P.; Sokolow, R.H. Reactive nitrogen: Too much of a good thing. Ambio 2002, 31, 60–66. [Google Scholar] [PubMed]
- Paerl, H.W.; Dennis, R.L.; Whitall, D.R. Atmospheric deposition of nitrogen: Implications for nutrient over-enrichment of coastal waters. Estuaries 2002, 25, 677–693. [Google Scholar] [CrossRef]
- Elser, J.J.; Bracken, M.E.S.; Cleland, E.E.; Gruner, D.S.; Harpole, W.S.; Hillebrand, H.; Bgai, J.T.; Seabloom, E.W.; Shurin, J.B.; Smith, J.E. Global analysis of nitrogen and phosphorus limitation of primary producers in freshwater, marine and terrestrial ecosystems. Ecol. Lett. 2007, 10, 1124–1134. [Google Scholar] [CrossRef] [PubMed]
- Lewis, W.M., Jr.; Wurtsbaugh, W.A. Control of Lacustrine Phytoplankton by nutrients: Erosion of the Phosphorus Paradigm. Int. Rev. Gesamt. Hydrobiol. 2008, 93, 446–465. [Google Scholar] [CrossRef]
- Conley, D.J.; Paerl, H.W.; Howarth, R.W.; Boesch, D.F.; Seitzinger, S.P.; Havens, K.E.; Lancelot, C.; Likens, G.E. Controlling eutrophication: nitrogen and phosphorus. Science 2009, 323, 1014–1015. [Google Scholar] [CrossRef] [PubMed]
- Paerl, H.W.; Hall, N.S.; Calandrino, E.S. Controlling harmful cyanobacterial blooms in a world experiencing anthropogenic and climatic-induced change. Sci. Total Environ. 2011, 409, 1739–1745. [Google Scholar] [CrossRef] [PubMed]
- Scavia, D.; Allan, J.D.; Arend, K.K.; Bartell, S.; Beletsky, D.; Bosch, N.S.; Brandt, S.B.; Briland, R.D.; Daloğlu, I.; DePinto, J.V.; et al. Assessing and addressing the re-eutrophication of Lake Erie: Central basin hypoxia. J. Great Lakes Res. 2014, 40, 226–246. [Google Scholar] [CrossRef]
- Lewis, W.M.; Wurtsbaugh, W.A.; Paerl, H.W. Rationale for control of anthropogenic nitrogen and phosphorus in inland waters. Environ. Sci. Technol. 2011, 45, 10030–10035. [Google Scholar] [CrossRef]
- Schindler, D.W.; Hecky, R.E.; Findlay, D.L.; Stainton, M.P.; Parker, B.R.; Paterson, M.; Beaty, K.G.; Lyng, M.; Kasian, S.E.M. Eutrophication of lakes cannot be controlled by reducing nitrogen input: Results of a 37 year whole ecosystem experiment. Proc. Natl. Acad. Sci. USA 2008, 105, 11254–11258. [Google Scholar] [CrossRef] [PubMed]
- Scott, J.T.; McCarthy, M.J. Nitrogen fixation may not balance the nitrogen pool in lakes over timescales relevant to eutrophication management. Limnol. Oceangr. 2010, 55, 1265–1270. [Google Scholar] [CrossRef]
- Paerl, H.W. Physiological ecology and regulation of N2 fixation in natural waters. Adv. Microb. Ecol. 1990, 11, 305–344. [Google Scholar]
- Xu, H.; Paerl, H.W.; Qin, B.; Zhu, G.; Gao, G. Nitrogen and phosphorus inputs control phytoplankton growth in eutrophic Lake Taihu, China. Limnol. Oceanogr. 2010, 55, 420–432. [Google Scholar] [CrossRef]
- Paerl, H.W.; Xu, H.; McCarthy, M.J.; Zhu, G.; Qin, B.; Li, Y.; Gardner, W.S. Controlling harmful cyanobacterial blooms in a hyper-eutrophic lake (Lake Taihu, China): The need for a dual nutrient (N & P) management strategy. Water Res. 2011, 45, 1973–1983. [Google Scholar] [CrossRef] [PubMed]
- Smolders, A.J.P.; Lucassen, E.C.H.E.T.; Bobbink, R.; Roelofs, J.G.M.; Lamers, L.P.M. How nitrate leaching from agricultural lands provokes phosphate eutrophication in groundwater fed wetlands: the sulphur bridge. Biogeochemistry 2010, 98, 1–7. [Google Scholar] [CrossRef]
- Vincent, W.F. Dominance of bloom forming cyanobacteria (Blue-green algae). New. Zeal. J. Mar. Freshw. Res. 1987, 21, 361–542. [Google Scholar] [CrossRef]
- Paerl, H.W.; Fulton, R.S., III. Ecology of harmful cyanobacteria. In Ecology of Harmful Algae; Graneli, E., Turner, J., Eds.; Springer: Berlin, Germany, 2006; pp. 95–109. [Google Scholar]
- Jöhnk, K.D.; Huisman, J.; Sharples, J.; Sommeijer, B.; Visser, P.M.; Stroom, J.M. Summer heatwaves promote blooms of harmful cyanobacteria. Glob. Chang. Biol. 2008, 14, 495–512. [Google Scholar] [CrossRef]
- Paerl, H.W.; Huisman, J. Blooms like it hot. Science 2008, 320, 57–58. [Google Scholar] [CrossRef] [PubMed]
- Paerl, H.W.; Huisman, J. Climate change: A catalyst for global expansion of harmful cyanobacterial blooms. Environ. Microbiol. Rep. 2009, 1, 27–37. [Google Scholar] [CrossRef] [PubMed]
- Paul, V.J. Global warming and cyanobacterial harmful algal blooms. In Cyanobacterial Harmful Algal Blooms: State of the Science and Research Needs; Advances in Experimental Medicine and Biology; Volume 619, Hudnell, H.K., Ed.; Springer: New York, NY, USA, 2008; pp. 239–257. [Google Scholar]
- Kosten, S.; Huszar, V.L.M.; Bécares, E.; Costa, L.S.; van Donk, E.; Hansson, L.A.; Jeppesen, E.; Kruk, C.; Lacerot, G.; Mazzeo, N.; et al. Warmer climates boost cyanobacterial dominance in shallow lakes. Glob. Chang. Biol. 2012, 18, 118–126. [Google Scholar] [CrossRef]
- Foy, R.H.; Gibson, C.E.; Smith, R.V. The influence of daylength, light intensity and temperature on the growth rates of planktonic blue-green algae. Eur. J. Phycol. 1976, 11, 151–163. [Google Scholar] [CrossRef]
- Robarts, R.D.; Zohary, T. Temperature effects on photosynthetic capacity, respiration, and growth rates of bloom-forming cyanobacteria. New. Zeal. J. Mar. Freshw. Res. 1987, 21, 391–399. [Google Scholar] [CrossRef]
- Butterwick, C.; Heaney, S.I.; Talling, J.F. Diversity in the influence of temperature on the growth rates of freshwater algae, and its ecological relevance. J. Freshw. Biol. 2005, 50, 291–300. [Google Scholar] [CrossRef]
- Weyhenmeyer, G.A. Warmer winters: Are planktonic algal populations in Sweden’s largest lakes affected? Ambio 2001, 30, 565–571. [Google Scholar] [PubMed]
- Elliott, J.A. The seasonal sensitivity of cyanobacteria and other phytoplankton to changes in flushing rate and water temperature. Glob. Chang. Biol. 2010, 16, 864–876. [Google Scholar] [CrossRef]
- Stüken, A.; Rücker, J.; Endrulat, T.; Preussel, K.; Hemm, M.; Nixdorf, B.; Karsten, U.; Wiedner, C. Distribution of three alien cyanobacterial species (Nostocales) in northeast Germany: Cylindrospermopsis raciborskii, Anabaena bergii and Aphanizomenon aphanizomenoides. Phycologia 2006, 45, 696–703. [Google Scholar] [CrossRef]
- Peeters, F.; Straile, D.; Lorke, A.; Livingstone, D.M. Earlier onset of the spring phytoplankton bloom in lakes of the temperate zone in a warmer climate. Glob. Chang. Biol. 2007, 13, 1898–1909. [Google Scholar] [CrossRef]
- Suikkanen, S.; Laamanen, M.; Huttunen, M. Long-term changes in summer phytoplankton communities of the open northern Baltic Sea. Estuar. Coast. Shelf Sci. 2007, 71, 580–592. [Google Scholar] [CrossRef]
- Wiedner, C.; Rücker, J.; Brüggemann, R.; Nixdorf, B. Climate change affects timing and size of populations of an invasive cyanobacterium in temperate regions. Oecologia 2007, 152, 473–484. [Google Scholar] [CrossRef] [PubMed]
- Wagner, C.; Adrian, R. Cyanobacteria dominance: Quantifying the effects of climate change. Limnol. Oceanogr. 2009, 54, 2460–2468. [Google Scholar] [CrossRef]
- Vincent, W.F. Microbial Ecosystems of Antarctica; Cambridge University Press: Cambridge, UK, 1988. [Google Scholar]
- Priscu, J.C. Ecosystem Dynamics in a Polar Desert: The McMurdo Dry Valleys, Antarctica; American Geophysical Union Publications: Washington, DC, USA, 1998; Volume 72. [Google Scholar]
- Priscu, J.C.; Fritsen, C.H.; Adams, E.E.; Giovannoni, S.J.; Paerl, H.W.; McKay, C.P.; Doran, T.; Lanoil, B.D.; Pinckney, J.L. Perennial Antarctic lake ice: An Oasis for life in a polar desert. Science 1998, 280, 2095–2098. [Google Scholar] [CrossRef] [PubMed]
- Vincent, W.F.; Quesada, A. Cyanobacteria in high latitude lakes, rivers and seas. In Ecology of Cyanobacteria II: Their Diversity in Space and Time; Whitton, B.A., Ed.; Springer: New York, NY, USA, 2012; pp. 371–385. [Google Scholar]
- Vincent, W.F. Cyanobacterial dominance in the polar regions. In The Ecology of Cyanobacteria; Whitton, B.A., Potts, M., Eds.; Kluwers Academic Press: Dordrecht, The Netherlands, 2000; pp. 321–340. [Google Scholar]
- Paerl, H.W.; Ustach, J. Blue-green algal scums: An explanation for their occurrence during freshwater blooms. Limnol. Oceanogr. 1982, 27, 212–217. [Google Scholar] [CrossRef]
- Verspagen, J.M.H.; van de Waal, D.B.; Finke, J.F.; Visser, P.M.; van Donk, E.; Huisman, J. Rising CO2 levels will intensify phytoplankton blooms in eutrophic and hypertrophic lakes. PLOS One 2014. [Google Scholar] [CrossRef]
- Webster, P.J.; Holland, G.J.; Curry, J.A.; Chang, H.R. Changes in tropical cyclone number, duration, and intensity in a warming environment. Science 2005, 309, 1844–1846. [Google Scholar] [CrossRef] [PubMed]
- Intergovernmental Panel on Climate Change (IPCC). Climate Change 2007. Synthesis Report, Contribution of Working Groups I, II and III to the Fourth Assessment Report of the Intergovernmental Panel on Climate Change; Pachauri, R.K., Reisinger, A., Eds.; IPCC: Geneva, Switzerland, 2007; p. 104. [Google Scholar]
- Intergovernmental Panel on Climate Change (IPCC). Managing the risks of extreme events and disasters to advance climate change adaptation. In A Special Report of Working Groups I and II of the Intergovernmental Panel on Climate Change; Field, C., Barros, V., Stocker, T.F., Qin, D., Dokken, D.J., Ebi, K.L., Mastrandrea, M.D., Mach, K.J., Plattner, G.K., Allen, S.K., et al., Eds.; Cambridge University Press: Cambridge, UK; New York, NY, USA, 2012; p. 582. [Google Scholar]
- Holland, G.J.; Webster, P.J. Heightened tropical cyclone activity in the North Atlantic: Natural variability of climate trend? Philos. Trans. Roy. Soc. A 2007. [Google Scholar] [CrossRef]
- Allan, R.P.; Soden, B.J. Atmospheric warming and the amplification of precipitation extremes. Science 2008, 321, 1481–1484. [Google Scholar] [CrossRef] [PubMed]
- Bender, M.A.; Knutson, T.R.; Tuleya, R.E.; Sirutis, J.J.; Vecchi, G.A.; Garner, S.T.; Held, I.M. Modeled impact of anthropogenic warming on the frequency of intense Atlantic hurricanes. Science 2010, 327, 454–458. [Google Scholar] [CrossRef] [PubMed]
- Trenberth, K.E. The impact of climate change and variability on heavy precipitation, floods, and droughts. In Encyclopedia of Hydrological Sciences; Anderson, M.G., Ed.; Wiley: Hoboken, NJ, USA, 2005. [Google Scholar]
- Paerl, H.W.; Paul, V. Climate change: Links to global expansion of harmful cyanobacteria. Water Res. 2011, 46, 1349–1363. [Google Scholar] [CrossRef] [PubMed]
- Walsby, A.E.; Hayes, P.K.; Boje, R.; Stal, L.J. The selective advantage of buoyancy provided by gas vesicles for planktonic cyanobacteria in the Baltic Sea. New Phytol. 1997, 136, 407–417. [Google Scholar] [CrossRef]
- Paerl, H.W.; Tucker, J.; Bland, P.T. Carotenoid enhancement and its role in maintaining blue-green algal (Microcystis aeruginosa) surface blooms. Limnol. Oceanogr. 1983, 28, 847–857. [Google Scholar] [CrossRef]
- Tonk, L.; Bosch, K.; Visser, P.M.; Huisman, J. Salt tolerance of the harmful cyanobacterium Microcystis aeruginosa. Aquat. Microb. Ecol. 2007, 46, 117–123. [Google Scholar] [CrossRef]
- Montangnolli, W.; Zamboni, A.; Luvizotto-Santos, R.; Yunes, J.S. Acute effects of Microcystis aeruginosa from the Patos Lagoon estuary, southern Brazil, on the microcrustacean Kalliapseudes schubartii (Crustacea: Tanaidacea). Arch. Environ. Contam. Toxicol. 2004, 46, 463–469. [Google Scholar] [PubMed]
- Mazur-Marzec, H.; Żeglińska, L.; Pliński, M. The effect of salinity on the growth, toxin production, and morphology of Nodularia spumigena isolated from the Gulf of Gdansk, southern Baltic Sea. J. Appl. Phycol. 2005, 17, 171–175. [Google Scholar] [CrossRef]
- Moisander, P.H.; McClinton, E., III; Paerl, H.W. Salinity effects on growth, photosynthetic parameters, and nitrogenase activity in estuarine planktonic cyanobacteria. Microb. Ecol. 2002, 43, 432–442. [Google Scholar] [CrossRef] [PubMed]
- Padisak, J. Cylindrospermopsis raciborskii (Woloszynska) Seenayya et Subba Raju, an expanding, highly adaptive cyanobacterium: Worldwide distribution and review of its ecology. Arch. Hydrobiol. 1997, 107, 563–593. [Google Scholar]
- Hamilton, P.; Ley, M.; Dean, S.; Pick, F. The occurrence of the cyanobacterium Cylindrospermopsis raciborskii in Constance Lake: An exotic cyanoprokaryote new to Canada. Phycologia 2005, 44, 17–25. [Google Scholar] [CrossRef]
- Chapman, A.D.; Schelske, C.L. Recent appearance of Cylindrospermopsis (Cyanobacteria) in five hypereutrophic Florida lakes. J. Phycol. 1997, 33, 191–195. [Google Scholar] [CrossRef]
- Calandrino, E.S.; Paerl, H.W. Determining the potential for the proliferation of the harmful cyanobacterium Cylindrospermopsis raciborskii in Currituck Sound, North Carolina. Harmful Algae 2011, 11, 1–9. [Google Scholar] [CrossRef]
- Cirés, S.; Wörmer, L.; Wiedner, C.; Quesada, A. Temperature-dependent dispersal strategies of Aphanizomenon ovalisporum (Nostocales, Cyanobacteria): Implications for the annual life cycle. Microb. Ecol. 2012. [Google Scholar] [CrossRef]
- Watkinson, A.J.; O’Neil, J.M.; Dennison, W.C. Ecophysiology of the marine cyanobacterium Lyngbya majuscula (Oscillatoriaceae) in Moreton Bay, Australia. Harmful Algae 2005, 4, 697–715. [Google Scholar] [CrossRef]
- Osborne, N.J.; Shaw, G.R.; Webb, P.M. Health effects of recreational exposure to Moreton Bay, Australia waters during a Lyngbya majuscula bloom. Environ. Int. 2007, 33, 309–314. [Google Scholar] [CrossRef] [PubMed]
- Paerl, H.W. Marine Plankton. In Ecology of Cyanobacteria II: Their Diversity in Space and Time; Whitton, B.A., Ed.; Springer: Dordrecht, The Netherlands, 2012; pp. 127–153. [Google Scholar]
- Visser, P.M.; Ibelings, B.W.; van der Veer, B.; Koedood, J.; Mur, L.R. Artificial mixing prevents nuisance blooms of the cyanobacterium Microcystis in Lake Nieuwe Meer, The Netherlands. Freshw. Biol. 1996, 36, 435–450. [Google Scholar] [CrossRef]
- Maier, H.R.; Kingston, G.B.; Clark, T.; Frazer, A.; Sanderson, A. Risk-based approach for assessing the effectiveness of flow management in controlling cyanobacterial blooms in rivers. River Res. Appl. 2004, 20, 459–471. [Google Scholar] [CrossRef]
- Qin, B.; Zhu, G.; Gao, G.; Zhang, Y.; Li, W.; Paerl, H.W.; Carmichael, W.W. A Drinking Water Crisis in Lake Taihu, China: Linkage to Climatic Variability and Lake Management. Environ. Manag. 2010, 45, 105–112. [Google Scholar] [CrossRef]
- Matthijs, H.C.P.; Visser, P.M.; Reeze, B.; Meeuse, J.; Slot, P.C.; Wjin, G.; Talens, R.; Huisman, J. Selective suppression of harmful cyanobacteria in an entire lake with hydrogen peroxide. Water Res. 2012, 46, 1460–1472. [Google Scholar] [CrossRef] [PubMed]
- Robb, M.; Greenop, B.; Goss, Z.; Douglas, G.; Adeney, J. Application of Phoslock, an innovative phosphorous binding clay, to two Western Australian waterways: Preliminary findings. Hydrobiologia 2003, 494, 237–243. [Google Scholar] [CrossRef]
- Petersen, S.A. Lake restoration by sediment removal. J. Am. Water Resour. Assoc. 1982, 18, 423–435. [Google Scholar] [CrossRef]
- Cronberg, G. Changes in the phytoplankton of Lake Trummen induced by restoration. Hydrobiologia 1982, 86, 185–193. [Google Scholar] [CrossRef]
- McQueen, D.J. Manipulating lake community structure: Where do we go from here? Freshw. Biol. 1990, 23, 613–620. [Google Scholar] [CrossRef]
- Ghadouani, A.; Pinel-Alloul, B.; Prepas, E.E. Effects of experimentally induced cyanobacterial blooms on crustacean zooplankton communities. Freshw. Biol. 2003, 48, 363–38. [Google Scholar] [CrossRef]
- Moss, B.; Madgwick, J.; Phillips, J.G. A Guide to the Restoration of Nutrient-enriched Shallow Lakes; W.W. Hawes: London, UK, 1996. [Google Scholar]
- Scheffer, M. Ecology of Shallow Lakes; Chapman and Hall: London, UK, 1998. [Google Scholar]
- Elser, J.J. The pathway to noxious cyanobacteria blooms in lakes: The food web as the final turn. Freshw. Biol. 1999, 42, 537–543. [Google Scholar] [CrossRef]
- Jeppesen, E.; Meerhoff, M.; Jacobsen, B.A.; Hansen, R.S.; Søndergaard, M.; Jensen, J.P.; Lauridsen, T.L.; Mazzeo, N.; Branco, C.W.C. Restoration of shallow lakes by nutrient control and biomanipulation: The successful strategy varies with lake size and climate. Hydrobiologia 2007, 581, 269–285. [Google Scholar] [CrossRef]
- Jeppesen, E.; Søndergaard, M.; Meerhoff, M.; Lauridsen, T.L.; Jensen, J.P. Shallow lake restoration by nutrient loading reduction: Some recent findings and challenges ahead. Hydrobiologia 2007, 584, 239–252. [Google Scholar] [CrossRef]
© 2014 by the authors; licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Paerl, H.W. Mitigating Harmful Cyanobacterial Blooms in a Human- and Climatically-Impacted World. Life 2014, 4, 988-1012. https://doi.org/10.3390/life4040988
Paerl HW. Mitigating Harmful Cyanobacterial Blooms in a Human- and Climatically-Impacted World. Life. 2014; 4(4):988-1012. https://doi.org/10.3390/life4040988
Chicago/Turabian StylePaerl, Hans W. 2014. "Mitigating Harmful Cyanobacterial Blooms in a Human- and Climatically-Impacted World" Life 4, no. 4: 988-1012. https://doi.org/10.3390/life4040988
APA StylePaerl, H. W. (2014). Mitigating Harmful Cyanobacterial Blooms in a Human- and Climatically-Impacted World. Life, 4(4), 988-1012. https://doi.org/10.3390/life4040988