Predator Avoidance in Extremophile Fish

Abstract
:1. Introduction
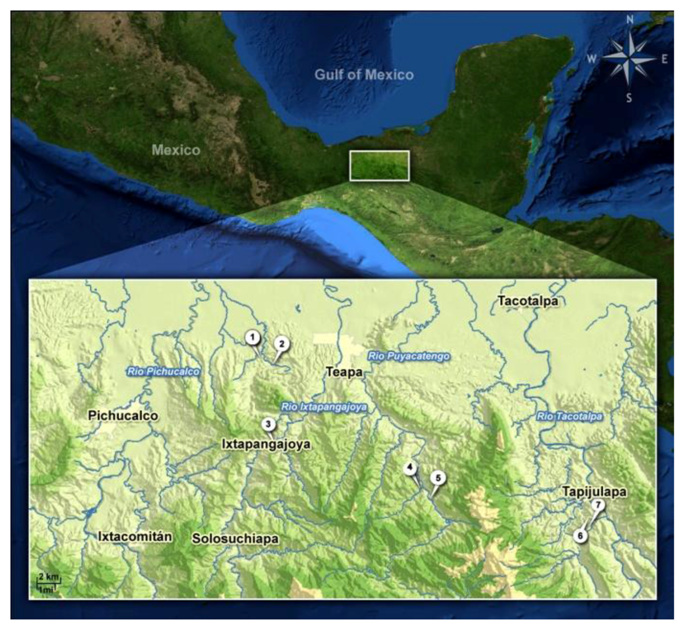
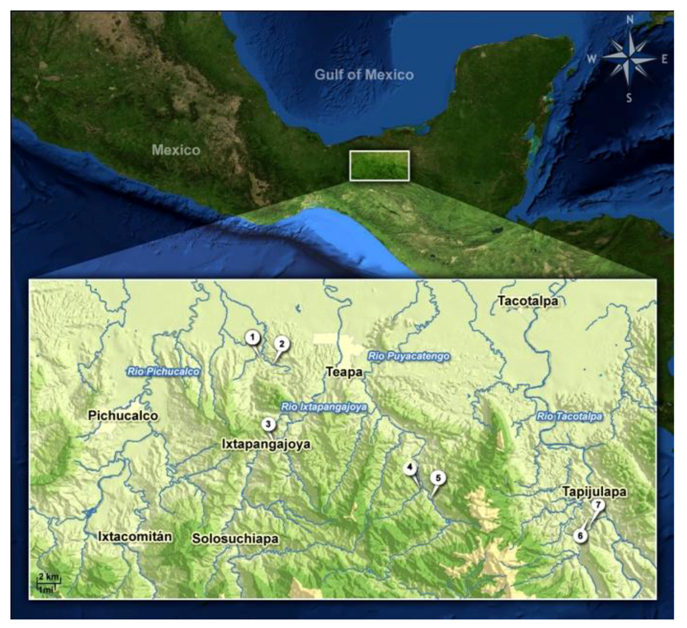
2. Experimental Section
2.1. Study Organisms, Sampling Sites and Maintenance of the Test Animals

{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Population | Species | Habitat / Treatment | SL [mm] | Sampling point | ||
|---|---|---|---|---|---|---|
| males | females | Latitude | Longitude | |||
| Río Ixtapangajoya (Río Puyacatengo drainage) | Poecilia mexicana | wc*, n, sf | 28.4 ± 0.4 (N = 28) | 28.9 ± 0.9 (N = 24) | 17.49450 | −92.99763 |
| Río Ixtapangajoya (Río Puyacatengo drainage) | P. mexicana | lab, n, sf | 39.5 ± 1.2 (N = 28) | 43.67 ± 2.0 (N = 24) | 17.49450 | −92.99763 |
| Río El Azufre (Río Pichucalco dranage) | P. mexicana | wc, n, sf | 40.5 ± 2.2 (N = 28) | 43.8 ± 1.6 (N = 24) | 17.55634 | −93.00762 |
| Baños del Azufre (Río Pichucalco drainage) | Poecilia sulphuraria | wc, s, sf | 24.6 ± 0.6 (N = 28) | 26.04 ± 1.1 (N = 24) | 17.55225 | −92.99859 |
| RíoPuyacatengo | P. mexicana | wc, n, sf | 39.2 ± 1.3 (N = 28) | 40.4 ± 2.3 (N = 25) | 17.47000 | −92.89573 |
| La Lluvia (Río Puyacatengo drainage) | P. mexicana | wc, s, sf | 28.2 ± 0.5 (N = 28) | 35.8 ± 0.9 (N = 25) | 17.46387 | −92.89541 |
| Cueva del Azufre (Río Tacotalpa drainage) | P. mexicana | wc, s, sf | 31.1 ± 0.6 (N = 28) | 39.3 ± 1.2 (N = 24) | 17.44225 | −92.77447 |
| El Azufre II (Río Tacotalpa drainage) | P. mexicana | wc, s, ca | 33.3 ± 0.9 (N = 28) | 35.8 ± 1.1 (N = 24) | 17.43843 | −92.77476 |
2.2. Experimental Design
2.3. Statistical Analyses
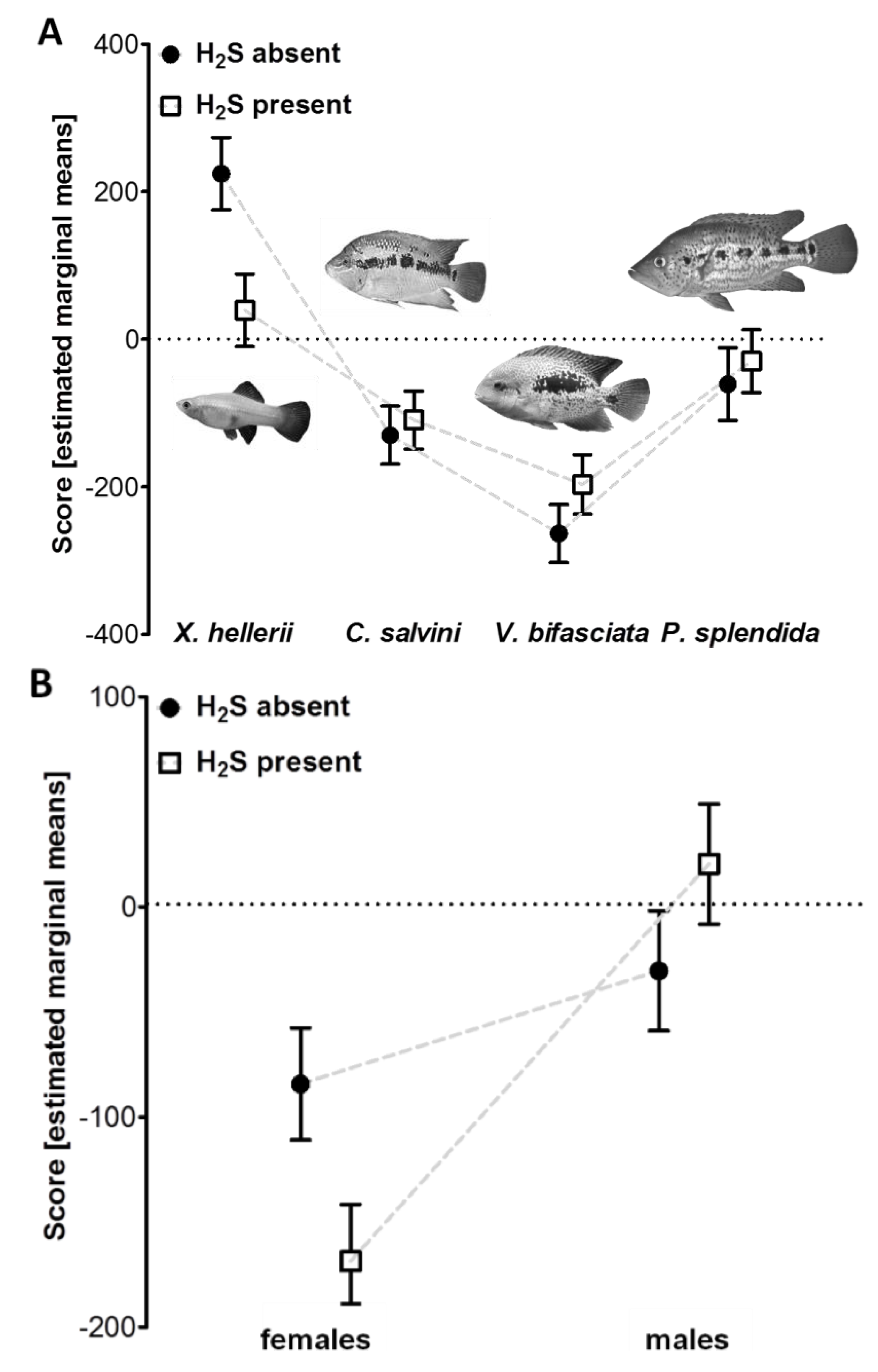
2.3.1. Experiment 1: Predator Recognition in Populations from Sulfide Springs
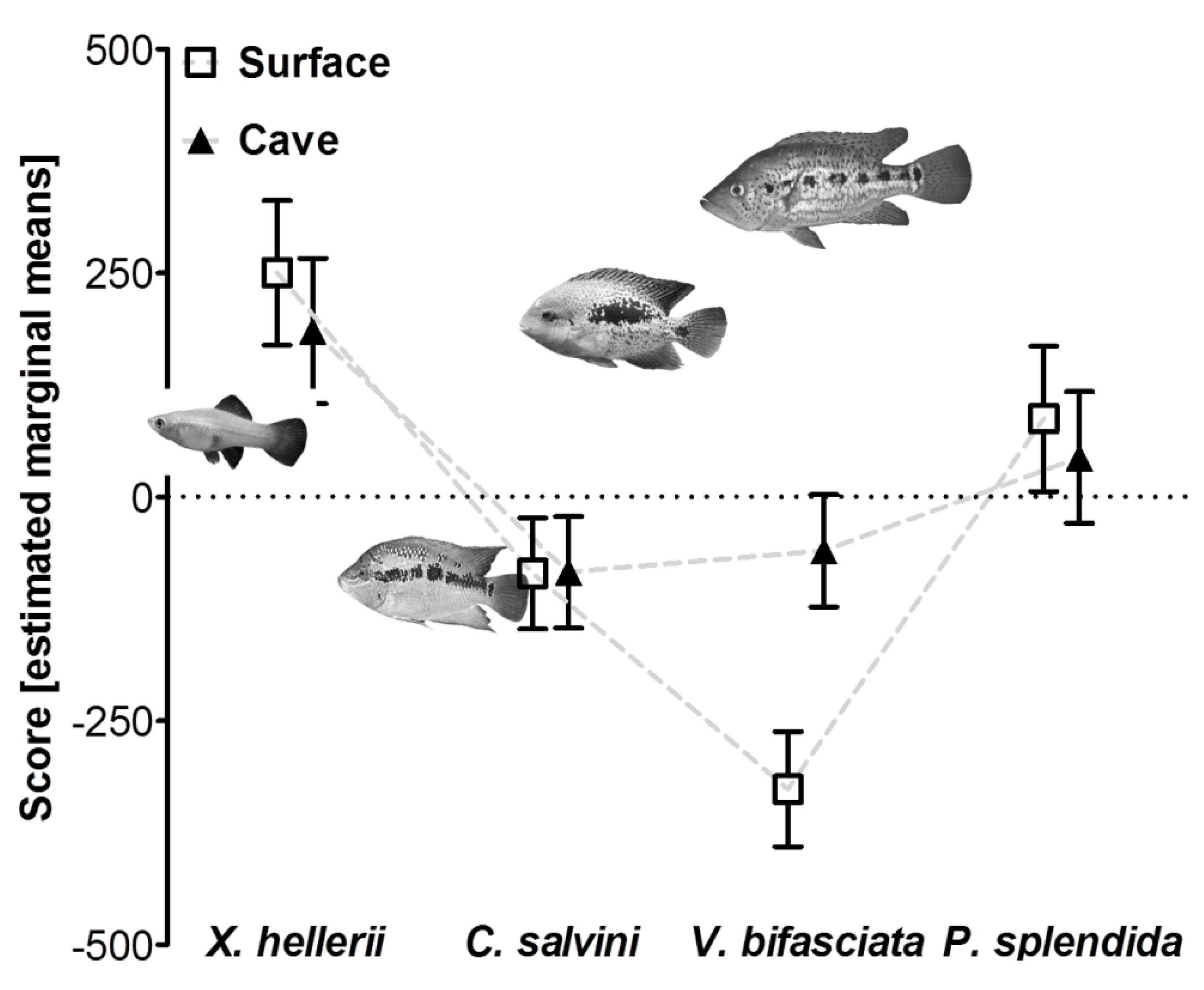
2.3.2. Experiment 2: Predator Recognition in Cave-Dwelling P. mexicana
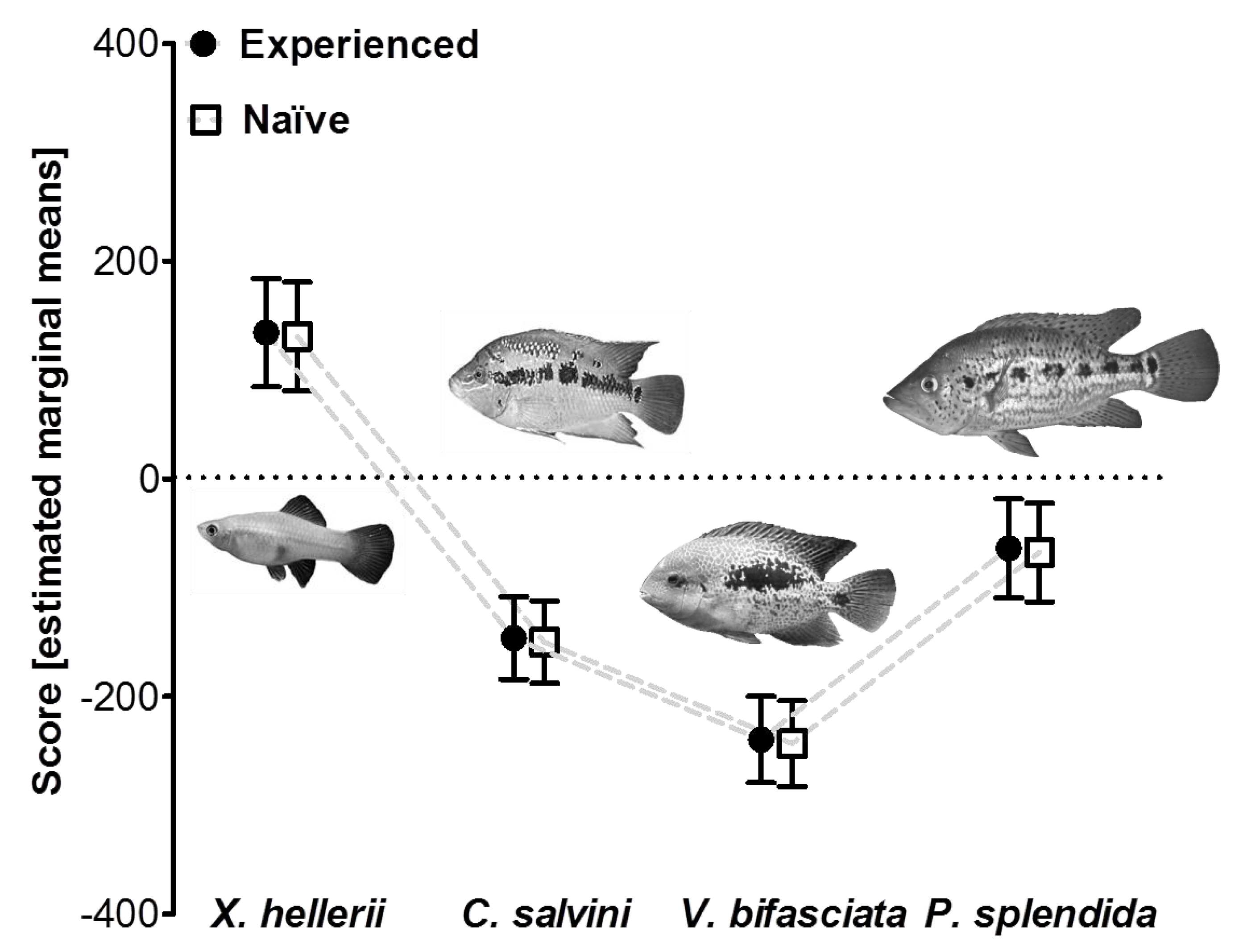
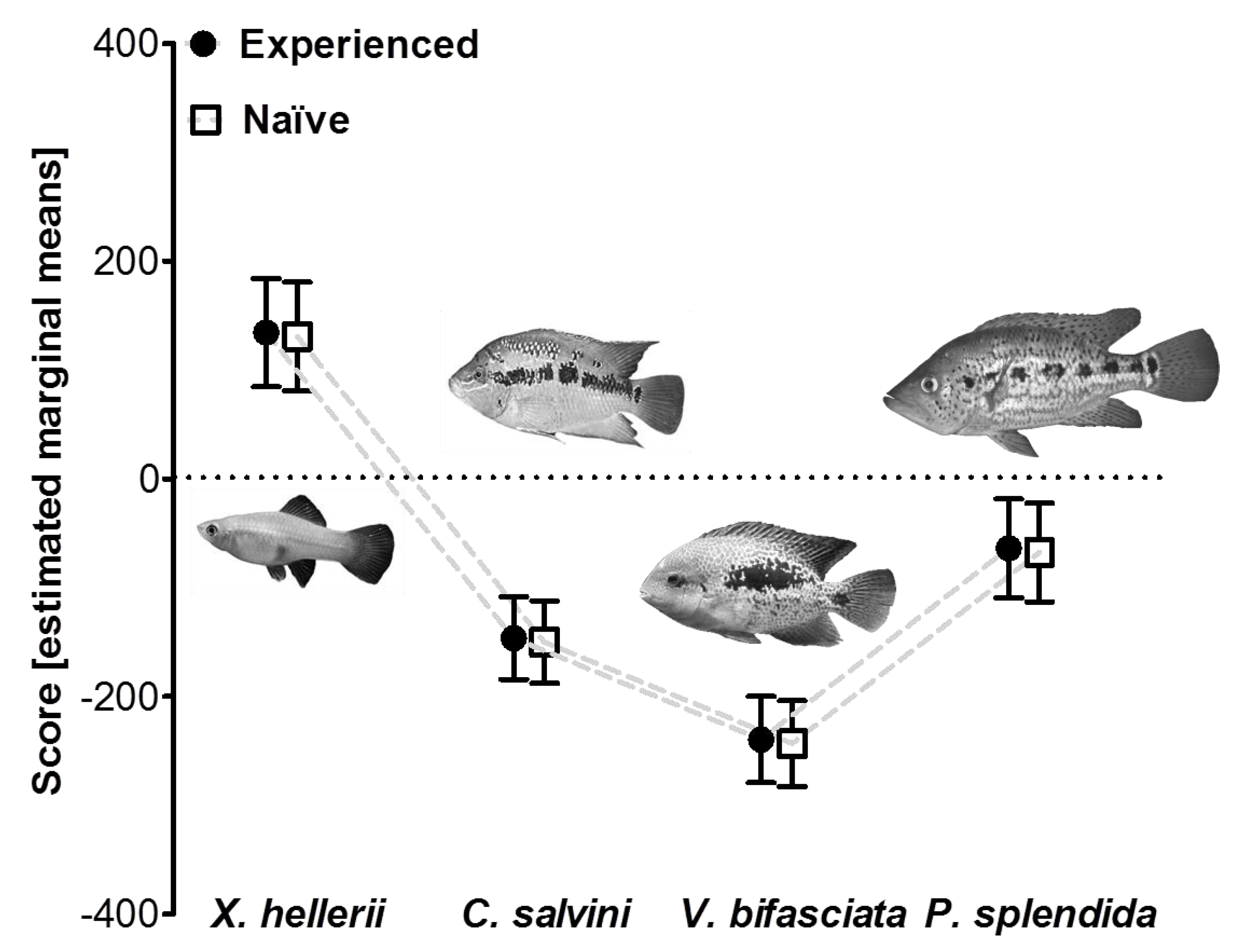
2.3.3. Experiment 3: Influence of Predator Experience
3. Results
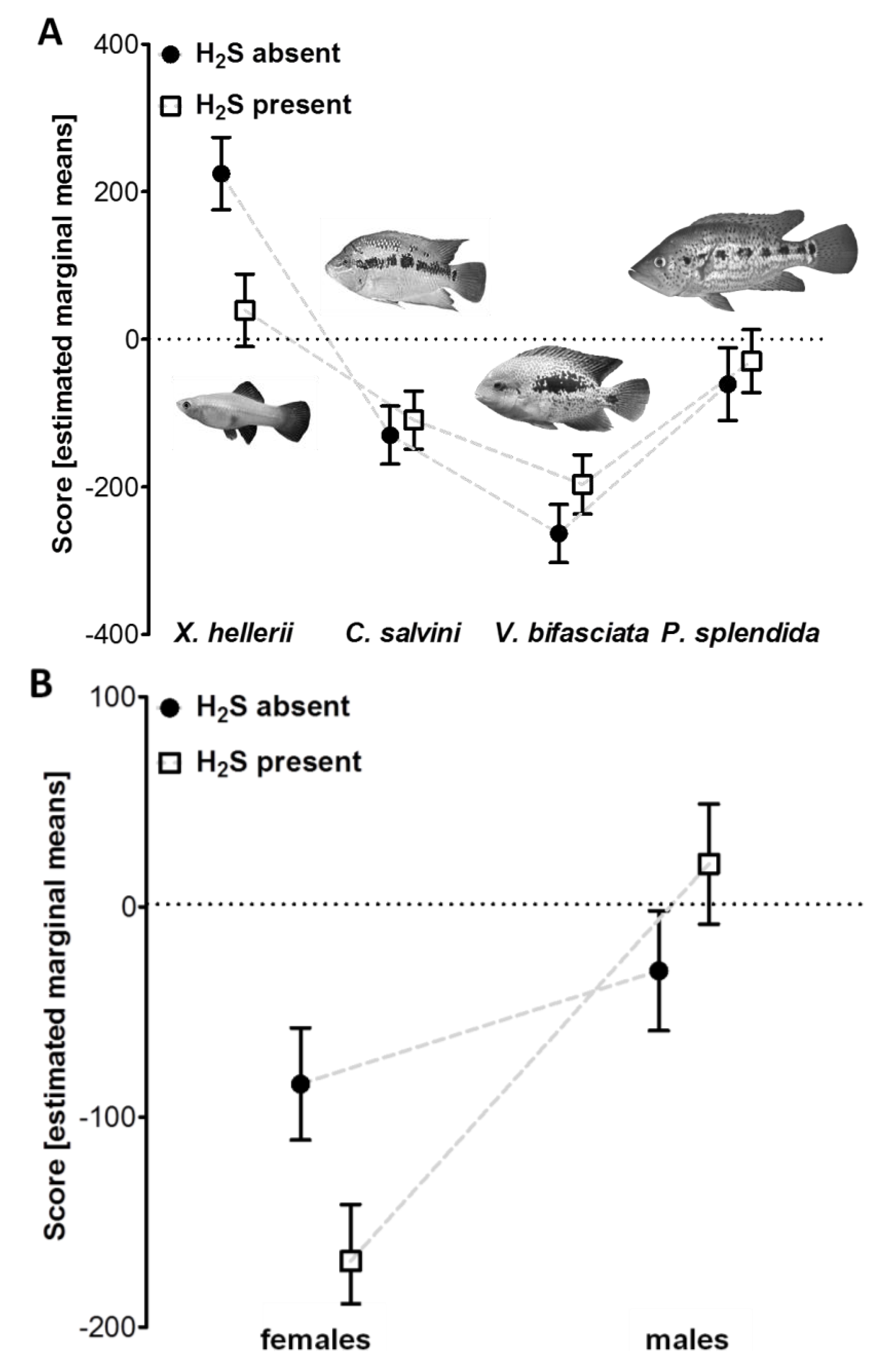
3.1. Reduced Predator Avoidance in H2S-Adapted Poecilia?
| df | F | P | partial eta2 | |
|---|---|---|---|---|
| (A) Experiment 1 | ||||
| predator type | 3 | 21.72 | <0.001 | 0.256 |
| drainage | 1 | 9.89 | 0.002 | 0.050 |
| H2S | 1 | 0.36 | 0.55 | 0.002 |
| sex | 1 | 19.28 | <0.001 | 0.093 |
| predator body size (SL) | 1 | 16.00 | <0.001 | 0.078 |
| predator type× drainage | 3 | 1.92 | 0.13 | 0.030 |
| predator type × H2S | 3 | 4.25 | 0.006 | 0.063 |
| predator type× sex | 3 | 1.30 | 0.28 | 0.020 |
| drainage× H2S | 1 | 0.61 | 0.44 | 0.003 |
| drainage× sex | 1 | 2.13 | 0.15 | 0.011 |
| H2S × sex | 1 | 5.95 | 0.016 | 0.030 |
| error | 189 | |||
| (B) Experiment 2 | ||||
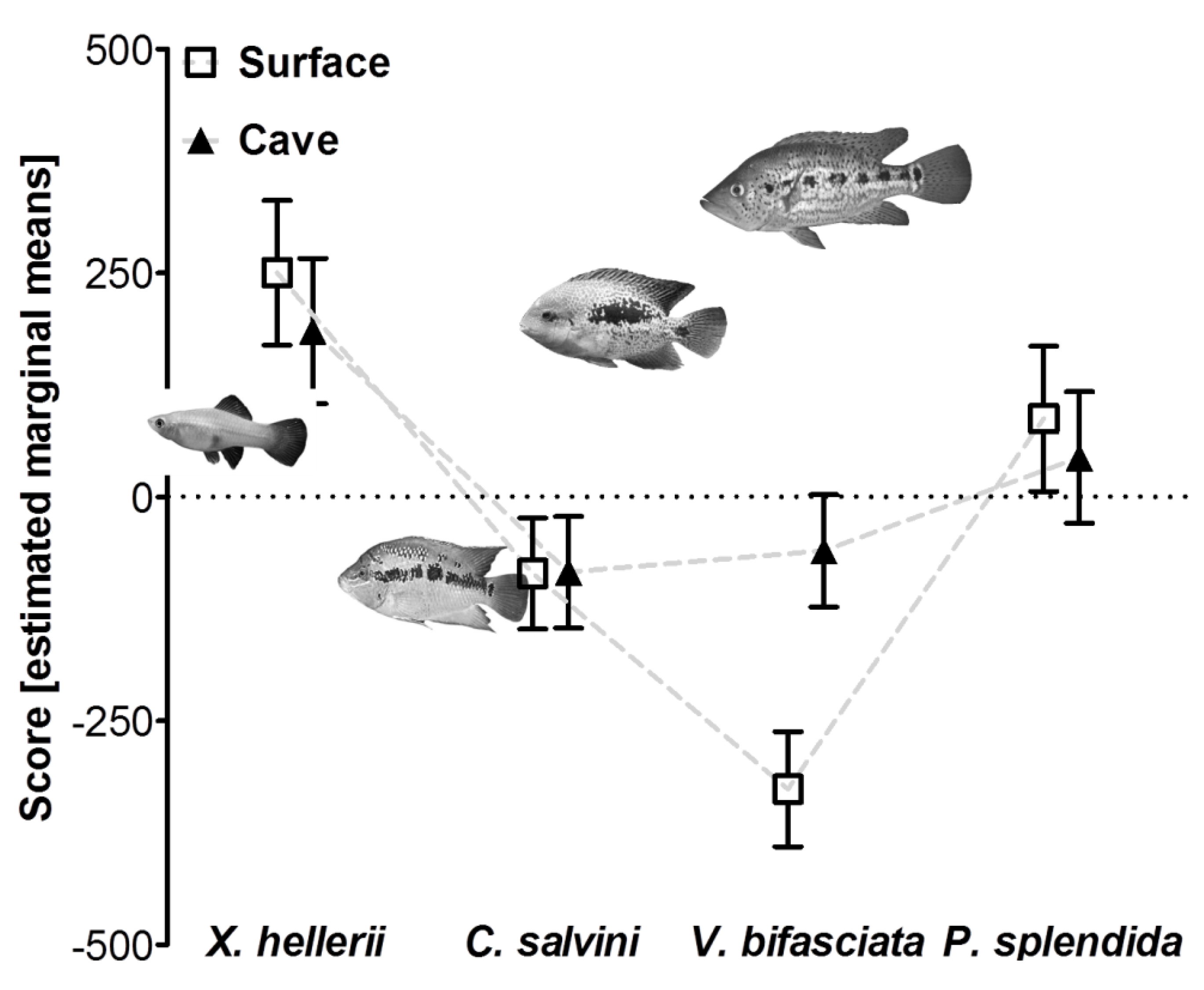
| predator type | 3 | 14.43 | <0.001 | 0.325 |
| sex | 1 | 0.61 | 0.44 | 0.007 |
| light regime | 1 | 0.84 | 0.36 | 0.009 |
| predator body size (SL) | 1 | 9.01 | 0.003 | 0.091 |
| predator type × sex | 3 | 0.90 | 0.44 | 0.029 |
| predator type × light regime | 3 | 3.20 | 0.027 | 0.096 |
| sex × light regime | 1 | 3.33 | 0.071 | 0.036 |
| error | 90 | |||
| (C) Experiment 3 | ||||
| predator type | 3 | 16.01 | <0.001 | 0.331 |
| sex | 1 | 0.05 | 0.82 | 0.001 |
| experience | 1 | 0.01 | 0.91 | <0.001 |
| predator body size (SL) | 1 | 12.93 | 0.001 | 0.118 |
| error | 97 |
3.2. Reduced Predator Avoidance in Cave Mollies?
3.3. Influence of Predator Experience on Predator Avoidance Responses in P. mexicana
4. Discussion
5. Conclusions
Acknowledgments
References and Notes
- Magnhagen, C. Predation risk as a cost of reproduction. Trends Ecol. Evol. 1991, 6, 183–186. [Google Scholar] [CrossRef]
- Kelley, J.L.; Magurran, A.E. Learned predator recognition and antipredator responses in fishes. Fish. Fish. 2003, 4, 216–226. [Google Scholar] [CrossRef]
- Utne-Palm, A. Response of naive two-spotted gobies Gobiusculus flavescens to visual and chemical stimuli of their natural predator, cod Gadus morhua. Mar. Ecol. Prog. Ser. 2001, 218, 267–274. [Google Scholar] [CrossRef]
- Kelley, J.L.; Magurran, A.E. Effects of relaxed predation pressure on visual predator recognition in the guppy. Behav. Ecol. Sociobiol. 2003, 54, 225–232. [Google Scholar] [CrossRef]
- Chivers, D.P.; Smith, R.J.F. Chemical alarm signalling in aquatic predator-prey systems: A review and prospectus. Écoscience 1998, 5, 338–352. [Google Scholar]
- Crowl, T.A.; Covich, A.P. Responses of a freshwater shrimp to chemical and tactile stimuli from a large decapod predator. J. N. Am. Benthol. Soc. 1994, 13, 291–298. [Google Scholar]
- Mahon, A.R.; Amsler, C.D.; McClintock, J.B.; Baker, B.J. Chemo-tactile predator avoidance responses of the common Antarctic limpet Nacella concinna. Polar Biol. 2002, 25, 469–473. [Google Scholar]
- Wisenden, B.; Pogatshnik, J.; Gibson, D.; Bonacci, L.; Schumacher, A.; Willett, A. Sound the alarm: learned association of predation risk with novel auditory stimuli by fathead minnows (Pimephales promelas) and glowlight tetras (Hemigrammus erythrozonus) after single simultaneous pairings with conspecific chemical alarm cues. Environ. Biol. Fish. 2008, 81, 141–147. [Google Scholar]
- Blumstein, D.T. The multipredator hypothesis and the evolutionary persistence of antipredator behavior. Ethology 2006, 112, 209–217. [Google Scholar] [CrossRef]
- Endler, J.A. Natural selection on color patterns in Poecilia reticulata. Evolution 1980, 34, 76–91. [Google Scholar] [CrossRef]
- Endler, J.A.; Houde, A.E. Geographic variation in female preferences for males traits in Poecilia reticulata. Evolution 1995, 49, 456–468. [Google Scholar] [CrossRef]
- Byers, J.A. American pronghorn: Social adaptations and the ghosts of predators past; University of Chicago Press: Chicago, IL, USA, 1997. [Google Scholar]
- Coss, R.G. Effects of relaxed natural selection on the evolution of behavior. In Geographic Variation in Behavior: Perspectives on Evolutionary Mechanisms; Foster, S.A., Endler, J.A., Eds.; Oxford Univ. Press: Oxford, UK, 1999; pp. 180–208. [Google Scholar]
- Blumstein, D.T.; Daniel, J.C. Isolation from mammalian predators differentially affects two congeners. Behav. Ecol. 2002, 13, 657–663. [Google Scholar] [CrossRef]
- Griffin, A.S.; Blumstein, D.T.; Evans, C. Training captive-bred or translocated animals to avoid predators. Conserv. Biol. 2000, 14, 1317–1326. [Google Scholar] [CrossRef]
- Magurran, A.E. The causes and consequences of geographic variation in antipredator behavior: perspectives from fish populations. In Geographic Variation in Behavior: Perspectives on Evolutionary Mechanisms; Foster, S.A., Endler, J.A., Eds.; Oxford Univ. Press: New York, USA, 1999; pp. 139–163. [Google Scholar]
- Brown, G.E.; Chivers, D.P.; Smith, R.J.F. Differential learning rates of chemical versus visual cues of a northern pike by fathead minnows in a natural habitat. Environ. Biol. Fish. 1997, 49, 89–96. [Google Scholar] [CrossRef]
- Bierbach, D.; Schulte, M.; Herrmann, N.; Tobler, M.; Stadler, S.; Jung, C.T.; Kunkel, B.; Riesch, R.; Klaus, S.; Ziege, M.; Indy, J.R.; Arias-Rodriguez, L.; Plath, M. Predator-induced changes of female mating preferences: innate and experiential effects. BMC Evol. Biol. 2011, 11, 190. [Google Scholar]
- Tobler, M.; Schlupp, I.; Heubel, K.U.; Riesch, R.; Garcia de León, F.J.; Giere, O.; Plath, M. Life on the edge: hydrogen sulfide and the fish communities of a Mexican cave and surrounding waters. Extremophiles 2006, 10, 577–585. [Google Scholar] [CrossRef]
- Riesch, R.; Duwe, V.; Herrmann, N.; Padur, L.; Ramm, A.; Scharnweber, K.; Schulte, M.; Schulz-Mirbach, T.; Ziege, M.; Plath, M. Variation along the shy–bold continuum in extremophile fishes (Poecilia mexicana, Poecilia sulphuraria). Behav. Ecol. Sociobiol. 2009, 63, 1515–1526. [Google Scholar] [CrossRef]
- Bagarinao, T. Sulfide as an environmental factor and toxicant: tolerance and adaptations in aquatic organisms. Aquat. Toxicol. 1992, 24, 21. [Google Scholar] [CrossRef]
- Grieshaber, M.K.; Völkel, S. Animal adaptations for tolerance and exploitation of poisonous sulfide. Annu. Rev. Physiol. 1998, 60, 33–53. [Google Scholar] [CrossRef]
- Plath, M.; Tobler, M.; Riesch, R.; Garcia de León, F.J.; Giere, O.; Schlupp, I. Survival in an extreme habitat: the roles of behaviour and energy limitation. Naturwissenschaften 2007, 94, 991–996. [Google Scholar] [CrossRef]
- Tobler, M.; Riesch, R.; Tobler, C.M.; Plath, M. Compensatory behavior in response to sulphide-induced hypoxia affects time budgets, feeding efficiency, and predation risk. Evol. Ecol. Res. 2009, 11, 935–948. [Google Scholar]
- Riesch, R.; Oranth, A.; Dzienko, J.; Karau, N.; Schießl, A.; Stadler, S.; Wigh, A.; Zimmer, C.; Arias-Rodriguez, L.; Schlupp, I.; Plath, M. Extreme habitats are not refuges: Poeciliids suffer from increased aerial predation risk in sulphidic southern Mexican habitats. Biol. J. Linn. Soc. 2010, 101, 417–426. [Google Scholar] [CrossRef]
- Roach, K.A.; Tobler, M.; Winemiller, K.O. Hydrogen sulfide, bacteria, and fish: a unique, subterranean food chain. Ecology 2011, 92, 2056–2062. [Google Scholar]
- Plath, M.; Heubel, K.U.; Garcia de León, F.J.; Schlupp, I. Cave molly females (Poecilia mexicana, Poeciliidae, Teleostei) like well-fed males. Behav. Ecol. Sociobiol. 2005, 58, 144–151. [Google Scholar] [CrossRef]
- Riesch, R.; Plath, M.; Schlupp, I. Toxic hydrogen sulphide and dark caves: pronounced male life-history divergence among locally adapted Poecilia mexicana (Poeciliidae). J. Evol. Biol. 2011, 24, 596–606. [Google Scholar] [CrossRef]
- Riesch, R.; Plath, M.; Schlupp, I. Toxic hydrogen sulfide and dark caves: life-history adaptations in a livebearing fish (Poecilia mexicana, Poeciliidae). Ecology 2010, 91, 1494–1505. [Google Scholar]
- Gordon, M.S.; Rosen, D.E. A cavernicolous form of the poeciliid dish Poecilia sphenops from Tabasco, México. Copeia 1962, 1962, 360–368. [Google Scholar] [CrossRef]
- Tobler, M.; Palacios, M.; Chapman, L.J.; Mitrofanov, I.; Bierbach, D.; Plath, M.; Arias-Rodriguez, L.; Garcia de León, F.J.; Mateos, M. Evolution in extreme environments: replicated phenotypic differentiation in livebearing fish inhabiting sulfidic springs. Evolution 2011, 65, 2213–2228. [Google Scholar] [CrossRef]
- Tobler, M.; Riesch, R.; Garcia de León, F.J.; Schlupp, I.; Plath, M. Two endemic and endangered fishes, Poecilia sulphuraria (Alvarez, 1948) and Gambusia eurystoma Miller, 1975 (Poeciliidae, Teleostei) as only survivors in a small sulphidic habitat. J. Fish. Biol. 2008, 72, 523–533. [Google Scholar] [CrossRef]
- Tobler, M.; Franssen, C.M.; Plath, M. Male-biased predation of a cave fish by a giant water bug. Naturwissenschaften 2008, 95, 775–779. [Google Scholar] [CrossRef]
- Tobler, M.; Schlupp, I.; Plath, M. Predation of a cave fish (Poecilia mexicana, Poeciliidae) by a giant water-bug (Belostoma, Belostomatidae) in a Mexican sulphur cave. Ecol. Entomol. 2007, 32, 492–495. [Google Scholar]
- Horstkotte, J.; Riesch, R.; Plath, M.; Jäger, P. Predation on a cavefish (Poecilia mexicana) by three species of spiders in a Mexican sulfur cave. Bull. Br. Arachnol. Soc. 2010, 15, 55–58. [Google Scholar]
- Klaus, S.; Plath, M. Predation on a cave fish by the freshwater crab Avotrichodactylus bidens (Bott, 1969) (Brachyura, Trichodactylidae) in a Mexican sulfur cave. Crustaceana 2011, 84, 411–418. [Google Scholar] [CrossRef]
- Miller, R.R. Freshwater Fishes of Mexico; University of Chicago Press: Chicago, IL, USA, 2006. [Google Scholar]
- Conkel, D. Cichlids of North. and Central America; T.F.H. Publications: Neptune City, NJ, USA, 1993. [Google Scholar]
- Plath, M.; Schlupp, I. Parallel evolution leads to reduced shoaling behavior in two cave dwelling populations of Atlantic mollies (Poecilia mexicana, Poeciliidae, Teleostei). Environ. Biol. Fish. 2008, 82, 289–297. [Google Scholar] [CrossRef]
- Parzefall, J. Zur vergleichenden Ethologie verschiedener Mollienesia-Arten einschließlich einer Höhlenform von Mollienesia sphenops. Behaviour 1969, 33, 1–38. [Google Scholar] [CrossRef]
- Parzefall, J. Rückbildung aggressiver Verhaltensweisen bei einer Höhlenform von Poecilia sphenops (Pisces, Poeciliidae). Z. Tierpsychol. 1974, 35, 66–84. [Google Scholar] [CrossRef]
- Bierbach, D.; Klein, M.; Sassmannshausen, V.; Schlupp, I.; Riesch, R.; Parzefall, J.; Plath, M. Divergent evolution of male aggressive behaviour: another reproductive isolation mechanism in extremophile poeciliid fishes. Int. J. Evol. Biol. 2012, 2012. [Google Scholar]
- Peters, N.; Peters, G.; Parzefall, J.; Wilkens, H. Über degenerative und konstruktive Merkmale bei einer phylogenetisch jungen Höhlenform von Poecilia sphenops (Pisces, Poeciliidae). Int. Rev. Gesamten Hydrobiol. Hydrogr. 1973, 58, 417–436. [Google Scholar] [CrossRef]
- Plath, M.; Hauswaldt, J.S.; Moll, K.; Tobler, M.; Garcia de León, F.J.; Schlupp, I.; Tiedemann, R. Local adaptation and pronounced genetic differentiation in an extremophile fish, Poecilia mexicana, inhabiting a Mexican cave with toxic hydrogen sulphide. Mol. Ecol. 2007, 16, 967–976. [Google Scholar]
- Fontanier, M.; Tobler, M. A morphological gradient revisited: cave mollies vary not only in eye size. Environ. Biol. Fish. 2009, 86, 285–292. [Google Scholar]
- Körner, K.E.; Schlupp, I.; Plath, M.; Loew, E.R. Spectral sensitivity of mollies: comparing surface- and cave-dwelling Atlantic mollies, Poecilia mexicana. J. Fish. Biol. 2006, 69, 54–65. [Google Scholar] [CrossRef]
- Ferrari, M.C.O.; Gonzalo, A.; Messier, F.; Chivers, D.P. Generalization of learned predator recognition: an experimental test and framework for future studies. Proc. Roy. Soc. B 2007, 274, 1853–1859. [Google Scholar]
- Greven, H. Gonads, genitals, and reproductive biology. In Ecology and Evolution of Poeciliid Fishes; Evans, J., Pilastro, A., Schlupp, I., Eds.; Chicago University Press: Chicago, IL, USA, 2011. [Google Scholar]
- Evans, J.; Pilastro, A. Postcopulatory sexual selection. In Ecology and Evolution of Poeciliid Fishes; Evans, J., Pilastro, A., Schlupp, I., Eds.; Chicago University Press: Chicago, IL, USA, 2011. [Google Scholar]
- Plath, M.; Makowicz, A.M.; Schlupp, I.; Tobler, M. Sexual harassment in live-bearing fishes (Poeciliidae): Comparing courting and noncourting species. Behav. Ecol. 2007, 18, 680–688. [Google Scholar]
- Parzefall, J. A review of morphological and behavioural changes in the cave molly, Poecilia mexicana, from Tabasco, Mexico. Environ. Biol. Fish. 2001, 62, 263–275. [Google Scholar] [CrossRef]
- Kullander, S.O. Cichlidae (Cichlids); EDIPUCRS: Porto Alegre, Brasil, 2003. [Google Scholar]
- Peckarsky, B.L.; Penton, M.A. Why do Ephemerella nymphs scorpion posture - A ghost of predation past. Oikos 1988, 53, 185–193. [Google Scholar] [CrossRef]
- Warburton, K.; Lees, N. Species discrimination in guppies: learned responses to visual cues. Anim. Behav. 1996, 52, 371–378. [Google Scholar]
- Harris, S.; Ramnarine, I.W.; Smith, H.G.; Pettersson, L.B. Picking personalities apart: estimating the influence of predation, sex and body size on boldness in the guppy Poecilia reticulata. Oikos 2010, 119, 1711–1718. [Google Scholar] [CrossRef]
- Riesch, R.; Schlupp, I.; Tobler, M.; Plath, M. Reduction of the association preference for conspecifics in cave-dwelling Atlantic mollies, Poecilia mexicana. Behav. Ecol. Sociobiol. 2006, 60, 794–802. [Google Scholar] [CrossRef]
- Krause, J.; Ruxton, G.D. Living in groups; Oxford University Press: Oxford, 2002. [Google Scholar]
- Botham, M.S.; Hayward, R.K.; Morrell, L.J.; Croft, D.P.; Ward, J.R.; Ramnarine, I.; Krause, J. Risk-sensitive antipredator behavior in the Trinidadian guppy, Poecilia reticulata. Ecology 2008, 89, 3174–3185. [Google Scholar] [CrossRef]
- Karplus, I.; Algom, D. Visual cues for predator face recognition by reef fishes. Z. Tierpsychol. 1981, 55, 343–364. [Google Scholar]
- Brown, G.E.; Chivers, D.P. Learning about danger: chemical alarm cues and local risk assessment in prey fishes. In Fish Cognition and Behaviour; Brown, C., Laland, K.N., Krause, J., Eds.; Blackwell: London, UK, 2006; pp. 49–69. [Google Scholar]
- Kelley, J.L.; Brown, C. Predation risk and decision-making in poeciliid prey. In Ecology and Evolution of Poeciliid Fishes; Evans, J.P., Pilastro, A., Schlupp, I., Eds.; University of Chicago Press: Chicago, IL, USA, 2011; pp. 174–184. [Google Scholar]
- Fisher, H.S.; Wong, B.B.M.; Rosenthal, G.G. Alteration of the chemical environment disrupts communication in a freshwater fish. Proc. Roy. Soc. B 2006, 273, 1187–1193. [Google Scholar]
© 2013 by the authors; licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution license (http://creativecommons.org/licenses/by/3.0/).
Share and Cite
Bierbach, D.; Schulte, M.; Herrmann, N.; Zimmer, C.; Arias-Rodriguez, L.; Indy, J.R.; Riesch, R.; Plath, M. Predator Avoidance in Extremophile Fish. Life 2013, 3, 161-180. https://doi.org/10.3390/life3010161
Bierbach D, Schulte M, Herrmann N, Zimmer C, Arias-Rodriguez L, Indy JR, Riesch R, Plath M. Predator Avoidance in Extremophile Fish. Life. 2013; 3(1):161-180. https://doi.org/10.3390/life3010161
Chicago/Turabian StyleBierbach, David, Matthias Schulte, Nina Herrmann, Claudia Zimmer, Lenin Arias-Rodriguez, Jeane Rimber Indy, Rüdiger Riesch, and Martin Plath. 2013. "Predator Avoidance in Extremophile Fish" Life 3, no. 1: 161-180. https://doi.org/10.3390/life3010161
APA StyleBierbach, D., Schulte, M., Herrmann, N., Zimmer, C., Arias-Rodriguez, L., Indy, J. R., Riesch, R., & Plath, M. (2013). Predator Avoidance in Extremophile Fish. Life, 3(1), 161-180. https://doi.org/10.3390/life3010161