
Rutin Linoleate Triggers Oxidative Stress-Mediated Cytoplasmic Vacuolation in Non-Small Cell Lung Cancer Cells



{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
1. Introduction
2. Materials and Methods
2.1. Specific Reagents and Instruments
2.2. Safety Profile Assessment Using the EpiAirway™ 3D Model
2.3. Two-Dimensional Cell Culture Conditions
2.4. Cell Viability Assessment—The MTT Test
2.5. Kinetic Cell Proliferation Monitoring
2.6. Lactate Dehydrogenase (LDH) Release Quantification
2.7. Cell Morphology Evaluation
2.8. Neutral Red Staining and Uptake
2.9. Immunofluorescence-Based Examination of Cellular Components
2.10. ROS Production Measurement
3. Results
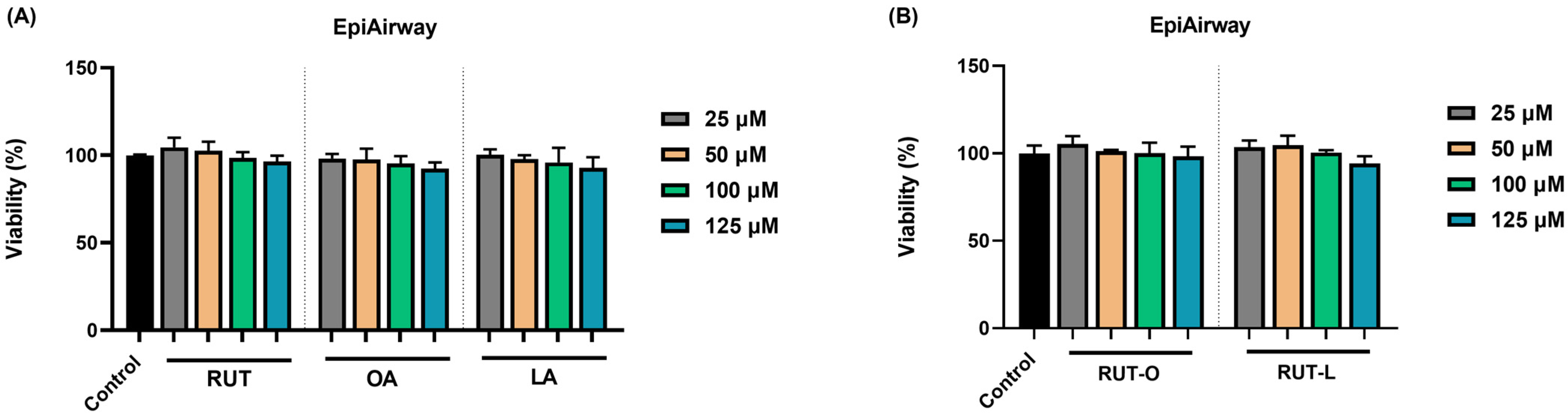
3.1. RUT-O and RUT-L Lack Cytotoxicity in EpiAirway™ 3D Tissues
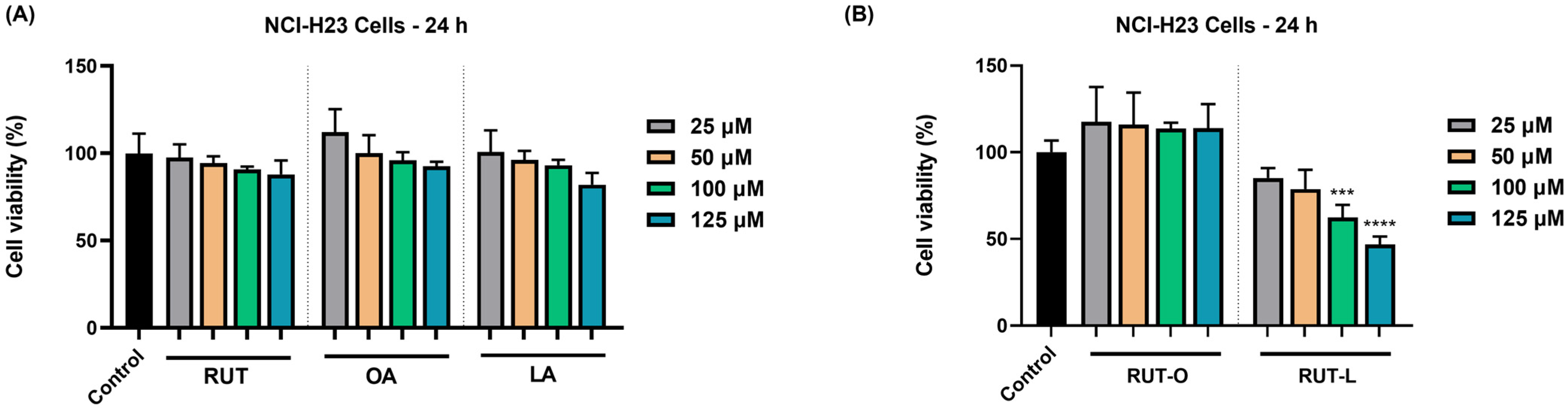
3.2. RUT-L Significantly Reduces the Viability of NCI-H23 Cells
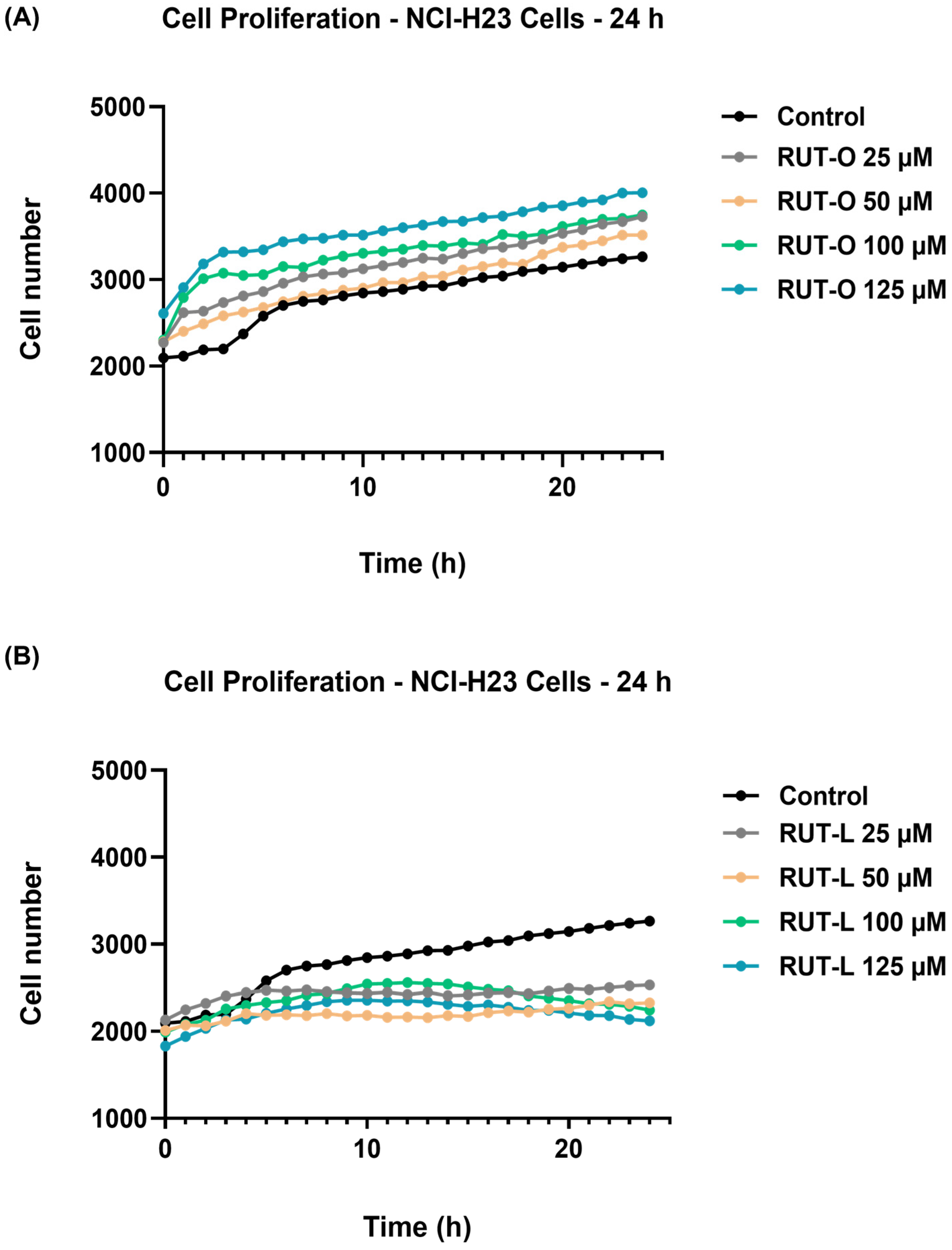
3.3. RUT-L Inhibits the Proliferation of NCI-H23 Cells
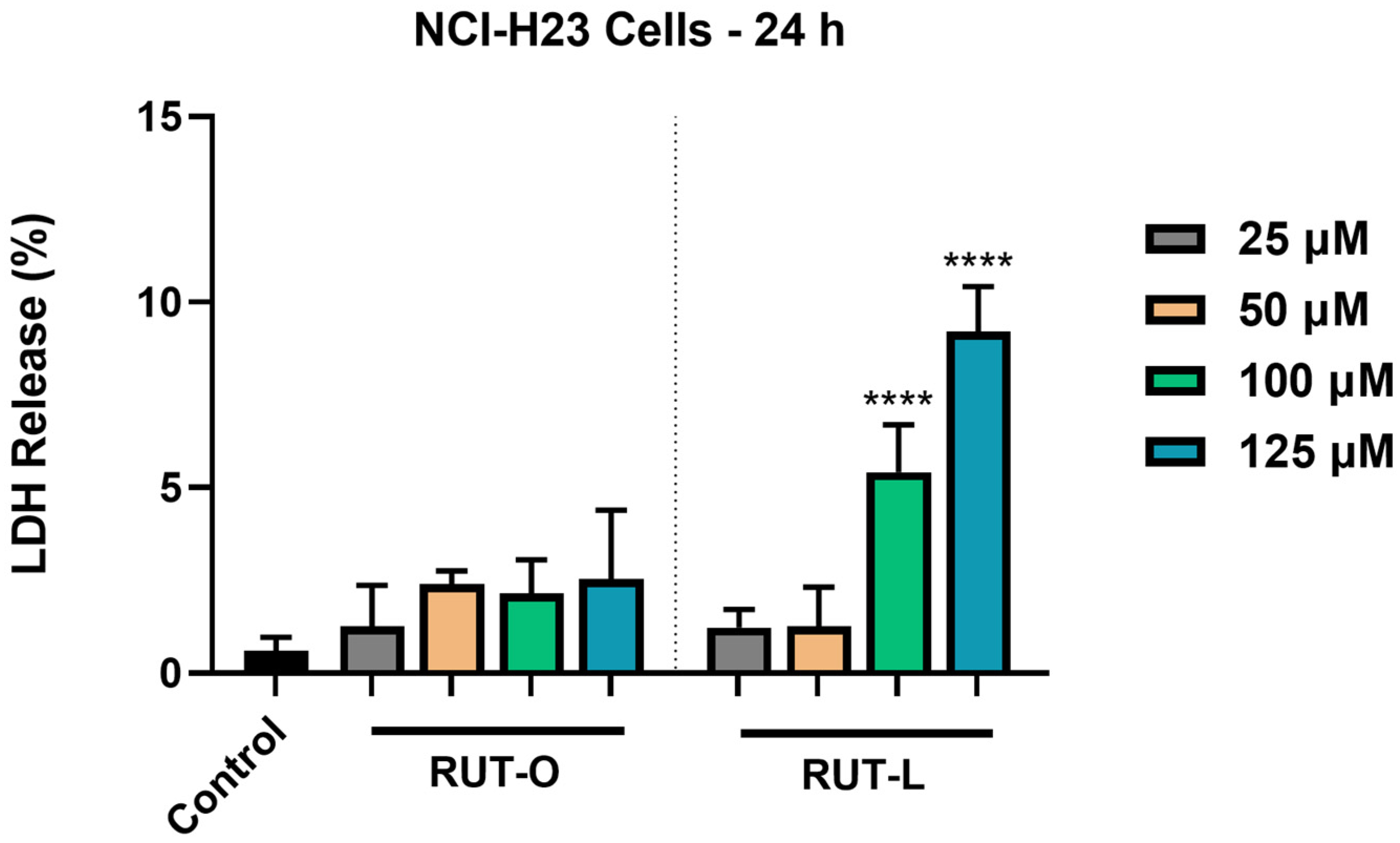
3.4. RUT-L Significantly Increases LDH Release from NCI-H23 Cells
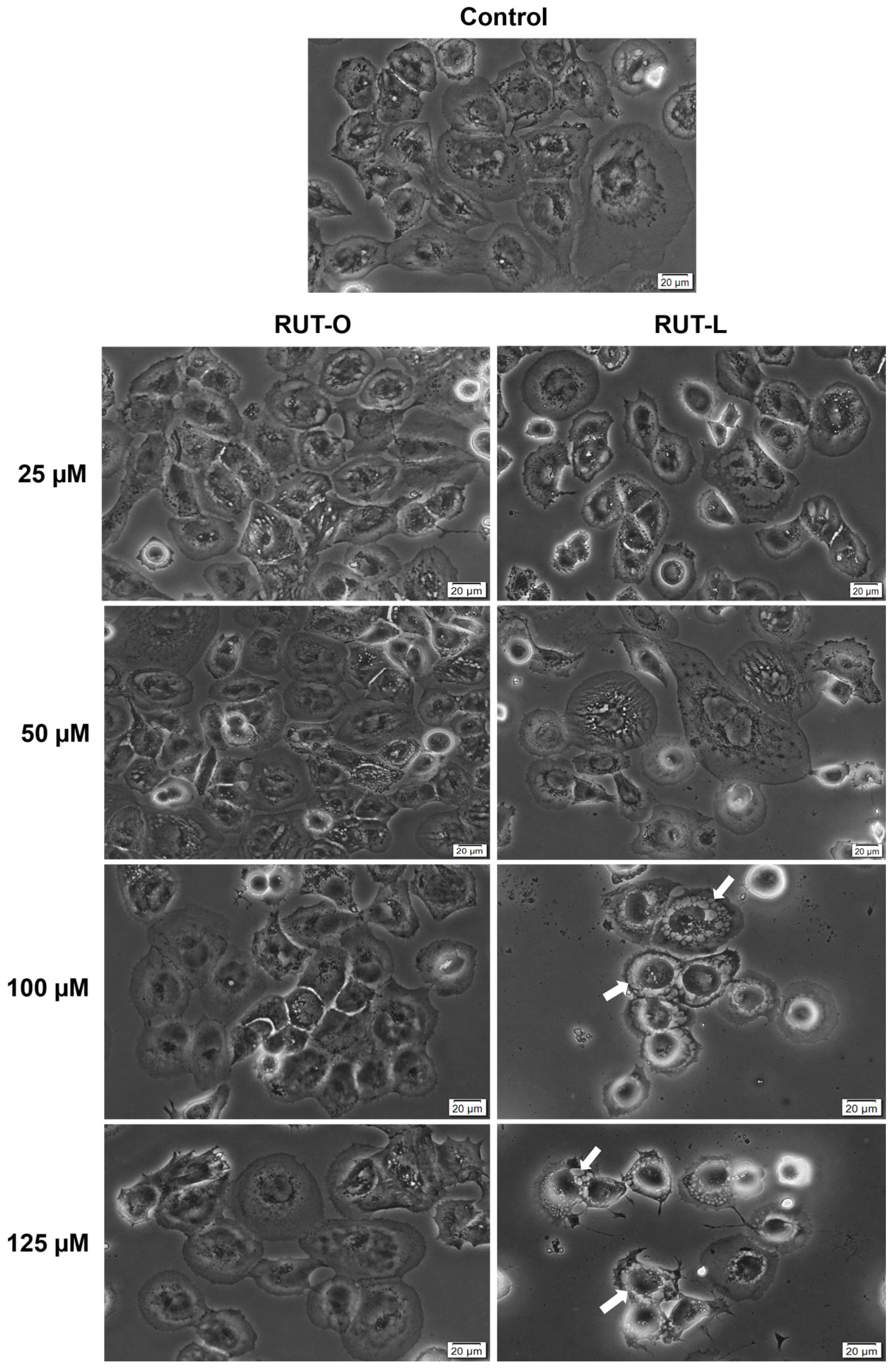
3.5. RUT-L Triggers Cytoplasmic Vacuolation in NCI-H23 Cells
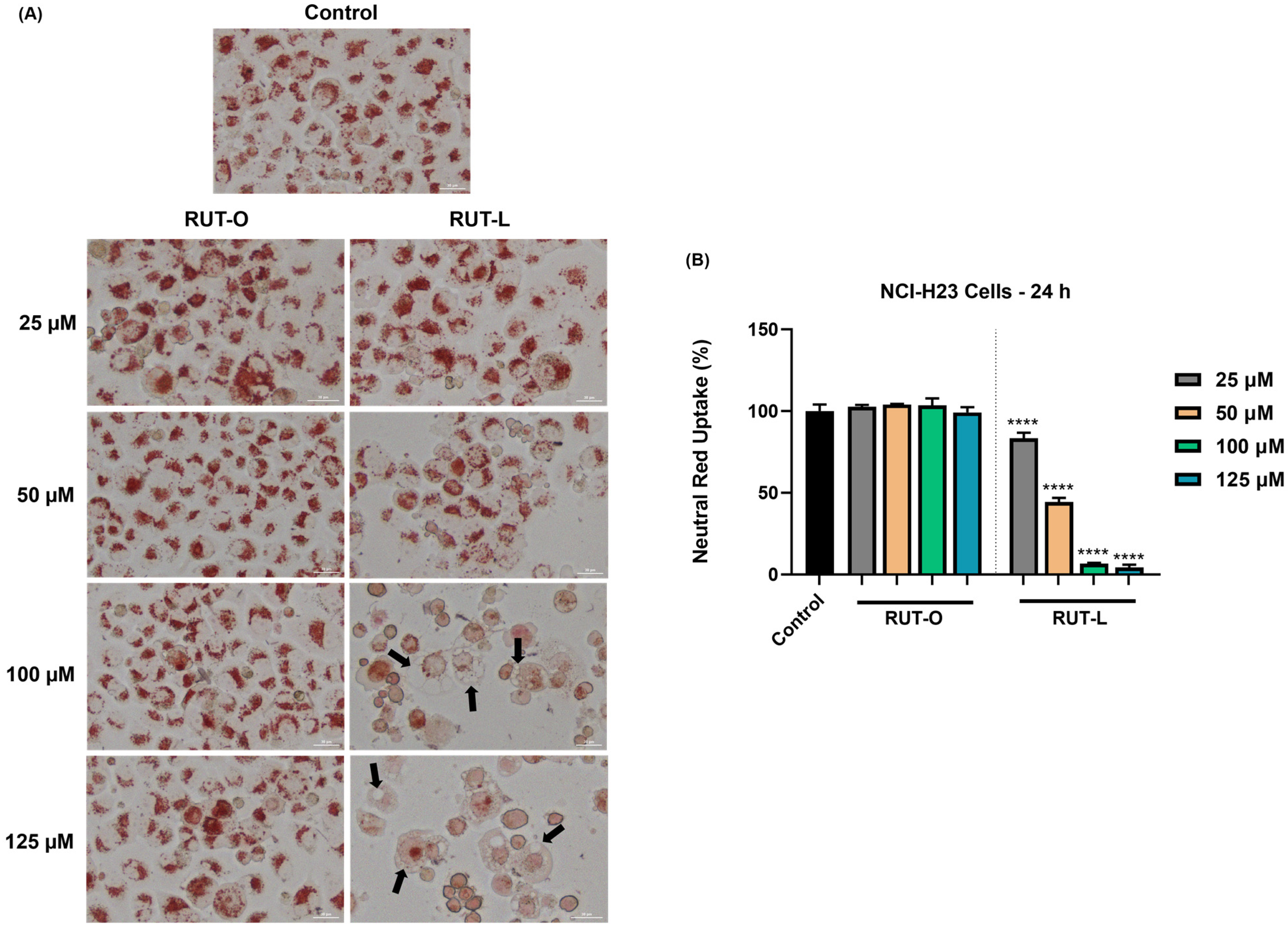
3.6. RUT-L Blocks Neutral Red Uptake in NCI-H23 Cells
3.7. RUT-L Alters Nuclear and Cytoskeletal Appearance in NCI-H23 Cells
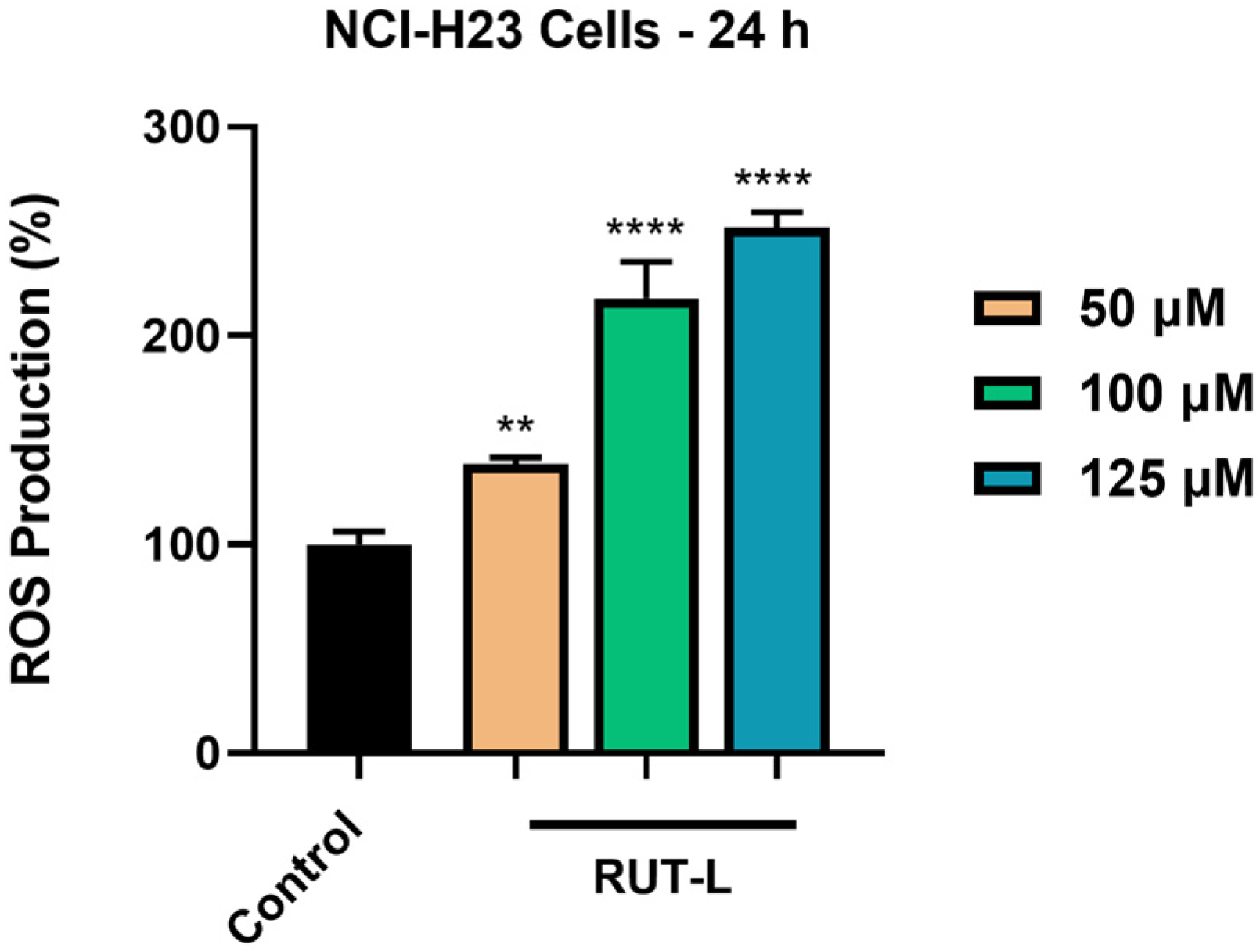
3.8. RUT-L Generates Oxidative Stress in NCI-H23 Cells
4. Discussion
5. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Thandra, K.C.; Barsouk, A.; Saginala, K.; Aluru, J.S.; Barsouk, A. Epidemiology of lung cancer. Contemp. Oncol. 2021, 25, 45–52. [Google Scholar]
- Walser, T.; Cui, X.; Yanagawa, J.; Lee, J.M.; Heinrich, E.; Lee, G.; Sharma, S.; Dubinett, S.M. Smoking and lung cancer: The role 540 of inflammation. Proc. Am. Thorac. Soc. 2008, 5, 811–815. [Google Scholar] [CrossRef]
- Lupo, P.J.; Spector, L.G. Cancer progress and priorities: Childhood cancer. Cancer Epidemiol. Biomark. Prev. 2020, 29, 1081–1094. [Google Scholar] [CrossRef]
- De Groot, P.; Munden, R.F. Lung Cancer Epidemiology, Risk Factors, and Prevention. Radiol. Clin. N. Am. 2012, 50, 863–876. [Google Scholar] [CrossRef]
- Lemjabbar-Alaoui, H.; Hassan, O.U.I.; Yang, Y.W.; Buchanan, P. Lung cancer: Biology and treatment options. Biochim. Biophys. Acta 2015, 1856, 189–210. [Google Scholar] [CrossRef]
- Nouri, Z.; Fakhri, S.; Nouri, K.; Wallace, C.E.; Farzaei, M.H.; Bishayee, A. Targeting Multiple Signaling Pathways in Cancer: The Rutin Therapeutic Approach. Cancers 2020, 12, 2276. [Google Scholar] [CrossRef]
- Naeem, A.; Hu, P.; Yang, M.; Zhang, J.; Liu, Y.; Zhu, W.; Zheng, Q. Natural Products as Anticancer Agents: Current Status and Future Perspectives. Molecules 2022, 27, 8367. [Google Scholar] [CrossRef]
- Sharma, S.; Ali, A.; Ali, J.; Sahni, J.K.; Baboota, S. Rutin: Therapeutic potential and recent advances in drug delivery. Expert. Opin. Investig. Drugs 2013, 22, 1063–1079. [Google Scholar] [CrossRef]
- Kopustinskiene, D.M.; Jakstas, V.; Savickas, A.; Bernatoniene, J. Flavonoids as anticancer agents. Nutrients 2020, 12, 457. [Google Scholar] [CrossRef]
- Dehelean, C.A.; Coricovac, D.; Pinzaru, I.; Marcovici, I.; Macasoi, I.G.; Semenescu, A.; Lazar, G.; Pinzaru, S.C.; Radulov, I.; Alexa, E.; et al. Rutin bioconjugates as potential nutraceutical prodrugs: An in vitro and in ovo toxicological screening. Front. Pharmacol. 2022, 13, 1000608. [Google Scholar] [CrossRef]
- Ganeshpurkar, A.; Saluja, A.K. The Pharmacological Potential of Rutin. Saudi Pharm. J. 2017, 25, 149–164. [Google Scholar] [CrossRef]
- Semwal, R.; Joshi, S.K.; Semwal, R.B.; Semwal, D.K. Health benefits and limitations of rutin—A natural flavonoid with high nutraceutical value. Phytochem. Lett. 2021, 46, 119–128. [Google Scholar] [CrossRef]
- Rashidinejad, A.; Jameson, G.B.; Singh, H. The Effect of pH and Sodium Caseinate on the Aqueous Solubility, Stability, and Crystallinity of Rutin towards Concentrated Colloidally Stable Particles for the Incorporation into Functional Foods. Molecules 2022, 27, 534. [Google Scholar] [CrossRef]
- Gullón, B.; Lú-Chau, T.A.; Moreira, M.T.; Lema, J.M.; Eibes, G. Rutin: A review on extraction, identification and purification methods, biological activities and approaches to enhance its bioavailability. Trends Food Sci. Technol. 2017, 67, 220–235. [Google Scholar] [CrossRef]
- Bhat, M. Opportunities and challenges of fatty acid conjugated therapeutics. Chem. Phys. Lipids 2021, 236, 105053. [Google Scholar] [CrossRef]
- Mecenas, A.S.; Malafaia, C.R.A.; Sangenito, L.S.; Simas, D.L.R.; de Barros Machado, T.; Amaral, A.C.F.; dos Santos, A.L.S.; Freire, D.M.G.; Leal, I.C.R. Rutin derivatives obtained by transesterification reactions catalyzed by Novozym 435: Antioxidant properties and absence of toxicity in mammalian cells. PLoS ONE 2018, 13, e0203159. [Google Scholar] [CrossRef]
- Viskupicova, J.; Maliar, T. Rutin fatty acid esters: From synthesis to biological health effects and application. J. Food Nutr. Res. 2017, 56, 232–243. [Google Scholar]
- Moacă, E.A.; Watz, C.; Faur, A.C.; Lazăr, D.; Socoliuc, V.; Păcurariu, C.; Ianoș, R.; Rus, C.I.; Minda, D.; Barbu-Tudoran, L.; et al. Biologic Impact of Green Synthetized Magnetic Iron Oxide Nanoparticles on Two Different Lung Tumorigenic Monolayers and a 3D Normal Bronchial Model—EpiAirwayTM Microtissue. Pharmaceutics 2023, 15, 2. [Google Scholar] [CrossRef]
- Gag, O.; Macasoi, I.; Pinzaru, I.; Dinu, S.; Popovici, R.; Cosoroaba, M.R.; Buzatu, R.; Cabuta, M.; Chiriac, S.D. In Vitro Assessment of the Impact of Ultraviolet B Radiation on Oral Healthy and Tumor Cells. Photonics 2023, 10, 464. [Google Scholar] [CrossRef]
- Repetto, G.; del Peso, A.; Zurita, J.L. Neutral red uptake assay for the estimation of cell viability/cytotoxicity. Nat. Protoc. 2008, 3, 1125–1131. [Google Scholar] [CrossRef]
- Cabezón-gutiérrez, L.; Sereno, M.; Cervera-calero, R.; Mielgo-rubio, X.; Higuera, O. High tumor burden in non-small cell lung cancer. Review of the literature. J. Clin. Transl. Res. 2022, 8, 403–413. [Google Scholar]
- Bourreau, C.; Treps, L.; Faure, S.; Fradin, D.; Clere, N. Therapeutic strategies for non-small cell lung cancer: Experimental models and emerging biomarkers to monitor drug efficacies. Pharmacol. Ther. 2023, 242, 108347. [Google Scholar] [CrossRef]
- Satari, A. Rutin: A Flavonoid as an Effective Sensitizer for Anticancer Therapy; Insights into Multifaceted Mechanisms and Applicability for Combination Therapy. EPMA J. 2021, 12, 155–176. [Google Scholar] [CrossRef]
- Jackson, G.R.; Maione, A.G.; Klausner, M.; Hayden, P.J. Prevalidation of an Acute Inhalation Toxicity Test Using the EpiAirway in Vitro Human Airway Model. Appl. Vitr. Toxicol. 2018, 4, 149–158. [Google Scholar] [CrossRef]
- Available online: https://www.mattek.com/ (accessed on 20 November 2023).
- Available online: https://www.atcc.org/ (accessed on 5 December 2023).
- Neitzel, J.J. Fatty Acid Molecules: Fundamentals and Role in Signaling. Nat. Educ. 2010, 3, 57. [Google Scholar]
- Czumaj, A.; Śledziński, T. Biological role of unsaturated fatty acid desaturases in health and disease. Nutrients 2020, 12, 356. [Google Scholar] [CrossRef]
- Zárate, R.; el Jaber-Vazdekis, N.; Tejera, N.; Pérez, J.A.; Rodríguez, C. Significance of long chain polyunsaturated fatty acids in human health. Clin. Transl. Med. 2017, 6, 25. [Google Scholar] [CrossRef]
- ISO 10993-5:2009(En); Biological Evaluation of Medical Devices—Part 5: Tests for In Vitro Cytotoxicity. ISO: Geneva, Switzerland, 2009. Available online: https://www.iso.org/obp/ui/#iso:std:iso:10993:-5:ed-3:v1:en (accessed on 6 December 2023).
- Hoai, T.T.; Yen, P.T.; Bich Dao, T.T.; Long, L.H.; Anh, D.X.; Minh, L.H.; Anh, B.Q.; Thuong, N.T. Evaluation of the cytotoxic effect of rutin prenanoemulsion in lung and colon cancer cell lines. J. Nanomater. 2020, 2020, 8867669. [Google Scholar] [CrossRef]
- ben Sghaier, M.; Pagano, A.; Mousslim, M.; Ammari, Y.; Kovacic, H.; Luis, J. Rutin inhibits proliferation, attenuates superoxide production and decreases adhesion and migration of human cancerous cells. Biomed. Pharmacother. 2016, 84, 1972–1978. [Google Scholar] [CrossRef]
- Yao, Q.; Li, H.; Fan, L.; Huang, S.; Wang, J.; Zheng, N. The combination of lactoferrin and linolenic acid inhibits colorectal tumor growth through activating AMPK/JNK-related apoptosis pathway. PeerJ 2021, 9, e11072. [Google Scholar] [CrossRef]
- Onishi, H.; Jimi, S.; Tsugu, H.; Lee, S.; Fukushima, T. Mechanisms of small globular protein-induced plasma membrane permeability and cytotoxicity in U87-MG human malignant glioblastoma cells. Anticancer Res. 2006, 26, 4033–4041. [Google Scholar]
- Han, Z.; Feng, D.; Wang, W.; Wang, Y.; Cheng, M.; Yang, H.; Liu, Y. Influence of Fatty Acid Modification on the Anticancer Activity of the Antimicrobial Peptide Figainin 1. ACS Omega 2023, 8, 41876–41884. [Google Scholar] [CrossRef]
- Viskupicova, J.; Danihelova, M.; Ondrejovic, M.; Liptaj, T.; Sturdik, E. Lipophilic rutin derivatives for antioxidant protection of oil-based foods. Food Chem. 2010, 123, 45–50. [Google Scholar] [CrossRef]
- Nair, S.V.G.; Ziaullah; Vasantha Rupasinghe, H.P. Fatty acid esters of phloridzin induce apoptosis of human liver cancer cells through altered gene expression. PLoS ONE 2014, 9, e107149. [Google Scholar] [CrossRef]
- Shubin, A.V.; Demidyuk, I.V.; Komissarov, A.A.; Rafieva, L.M.; Kostrov, S.V. C Cytoplasmic vacuolization in cell death and survival. Oncotarget 2016, 7, 55863–55889. [Google Scholar] [CrossRef]
- Fontana, F.; Raimondi, M.; Marzagalli, M.; Di Domizio, A.; Limonta, P. The emerging role of paraptosis in tumor cell biology: Perspectives for cancer prevention and therapy with natural compounds. Biochim. Biophys. Acta Rev. Cancer 2020, 1873, 188338. [Google Scholar] [CrossRef]
- Hanson, S.; Dharan, A.; Jinsha, P.V.; Pal, S.; Nair, B.G.; Kar, R.; Mishra, N. Paraptosis: A unique cell death mode for targeting cancer. Front. Pharmacol. 2023, 14, 1159409. [Google Scholar] [CrossRef]
- Maltese, W.A.; Overmeyer, J.H. Methuosis: Nonapoptotic cell death associated with vacuolization of macropinosome and endosome compartments. Am. J. Pathol. 2014, 184, 1630–1642. [Google Scholar] [CrossRef]
- Chen, X.; Chen, X.; Zhang, X.; Wang, L.; Cao, P.; Rajamanickam, V.; Wu, C.; Zhou, H.; Cai, Y.; Liang, G.; et al. Curcuminoid B63 induces ROS-mediated paraptosis-like cell death by targeting TrxR1 in gastric cells. Redox Biol. 2019, 21, 101061. [Google Scholar] [CrossRef]
- Kessel, D. Apoptosis, Paraptosis and Autophagy: Death and Survival Pathways Associated with Photodynamic Therapy. J. Pediatr. 2013, 176, 139–148. [Google Scholar] [CrossRef]
- Ates, G.; Vanhaecke, T.; Rogiers, V.; Rodrigues, R.M. Assaying Cellular Viability Using the Neutral Red Uptake Assay. Methods Mol. Biol. 2017, 1601, 19–26. [Google Scholar]
- Dubrovsky, J.G.; Guttenberger, M.; Saralegui, A.; Napsucialy-Mendivil, S.; Voigt, B.; Baluška, F.; Menzel, D. Neutral red as a probe for confocal laser scanning microscopy studies of plant roots. Ann. Bot. 2006, 97, 1127–1138. [Google Scholar] [CrossRef]
- Schwab, B.; Hülskamp, M. Neutral Red Staining for Plant Vacuoles. Cold Spring Harb. Protoc. 2010, 2010, pdb.prot4953. [Google Scholar] [CrossRef]
- Kubitschke, H.; Schnauss, J.; Nnetu, K.D.; Warmt, E.; Stange, R.; Kaes, J. Actin and microtubule networks contribute differently to cell response for small and large strains. New J. Phys. 2017, 19, 093003. [Google Scholar] [CrossRef]
- NavaneethaKrishnan, S.; Rosales, J.L.; Lee, K.Y. ROS-mediated cancer cell killing through dietary phytochemicals. Oxid. Med. Cell. Longev. 2019, 2019, 9051542. [Google Scholar] [CrossRef]
- Slika, H.; Mansour, H.; Wehbe, N.; Nasser, S.A.; Iratni, R.; Nasrallah, G.; Shaito, A.; Ghaddar, T.; Kobeissy, F.; Eid, A.H. Therapeutic potential of flavonoids in cancer: ROS-mediated mechanisms. Biomed. Pharmacother. 2022, 146, 112442. [Google Scholar] [CrossRef]
- Khan, F.; Pandey, P.; Jha, N.K.; Khalid, M.; Ojha, S. Rutin mediated apoptotic cell death in Caski cervical cancer cells via notch-621 1 and hes-1 downregulation. Life 2021, 11, 761. [Google Scholar] [CrossRef]

Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2024 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Marcovici, I.; Vlad, D.; Buzatu, R.; Popovici, R.A.; Cosoroaba, R.M.; Chioibas, R.; Geamantan, A.; Dehelean, C. Rutin Linoleate Triggers Oxidative Stress-Mediated Cytoplasmic Vacuolation in Non-Small Cell Lung Cancer Cells. Life 2024, 14, 215. https://doi.org/10.3390/life14020215
Marcovici I, Vlad D, Buzatu R, Popovici RA, Cosoroaba RM, Chioibas R, Geamantan A, Dehelean C. Rutin Linoleate Triggers Oxidative Stress-Mediated Cytoplasmic Vacuolation in Non-Small Cell Lung Cancer Cells. Life. 2024; 14(2):215. https://doi.org/10.3390/life14020215
Chicago/Turabian StyleMarcovici, Iasmina, Daliborca Vlad, Roxana Buzatu, Ramona Amina Popovici, Raluca Mioara Cosoroaba, Raul Chioibas, Andreea Geamantan, and Cristina Dehelean. 2024. "Rutin Linoleate Triggers Oxidative Stress-Mediated Cytoplasmic Vacuolation in Non-Small Cell Lung Cancer Cells" Life 14, no. 2: 215. https://doi.org/10.3390/life14020215
APA StyleMarcovici, I., Vlad, D., Buzatu, R., Popovici, R. A., Cosoroaba, R. M., Chioibas, R., Geamantan, A., & Dehelean, C. (2024). Rutin Linoleate Triggers Oxidative Stress-Mediated Cytoplasmic Vacuolation in Non-Small Cell Lung Cancer Cells. Life, 14(2), 215. https://doi.org/10.3390/life14020215