
Investigation of PACAP38 and PAC1 Receptor Expression in Human Retinoblastoma and the Effect of PACAP38 Administration on Human Y-79 Retinoblastoma Cells

 , , ,
, , ,
Abstract
1. Introduction
2. Materials and Methods
2.1. Human Eyes
Immunohistochemistry
2.2. Cell Culture
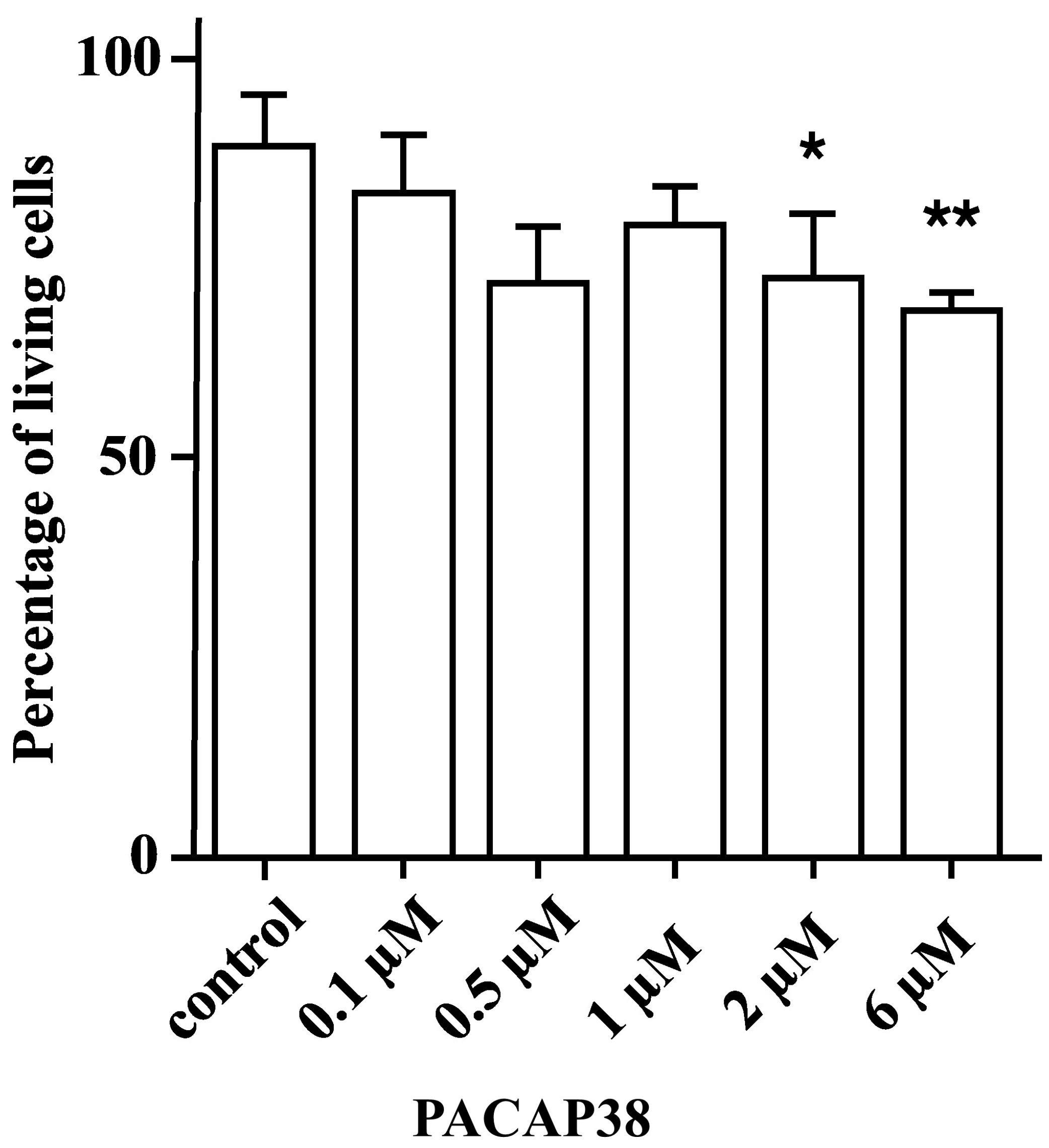
2.2.1. Cell Viability Assay
2.2.2. Statistical Analysis
3. Results
3.1. Human Eyes
3.1.1. Clinical Data
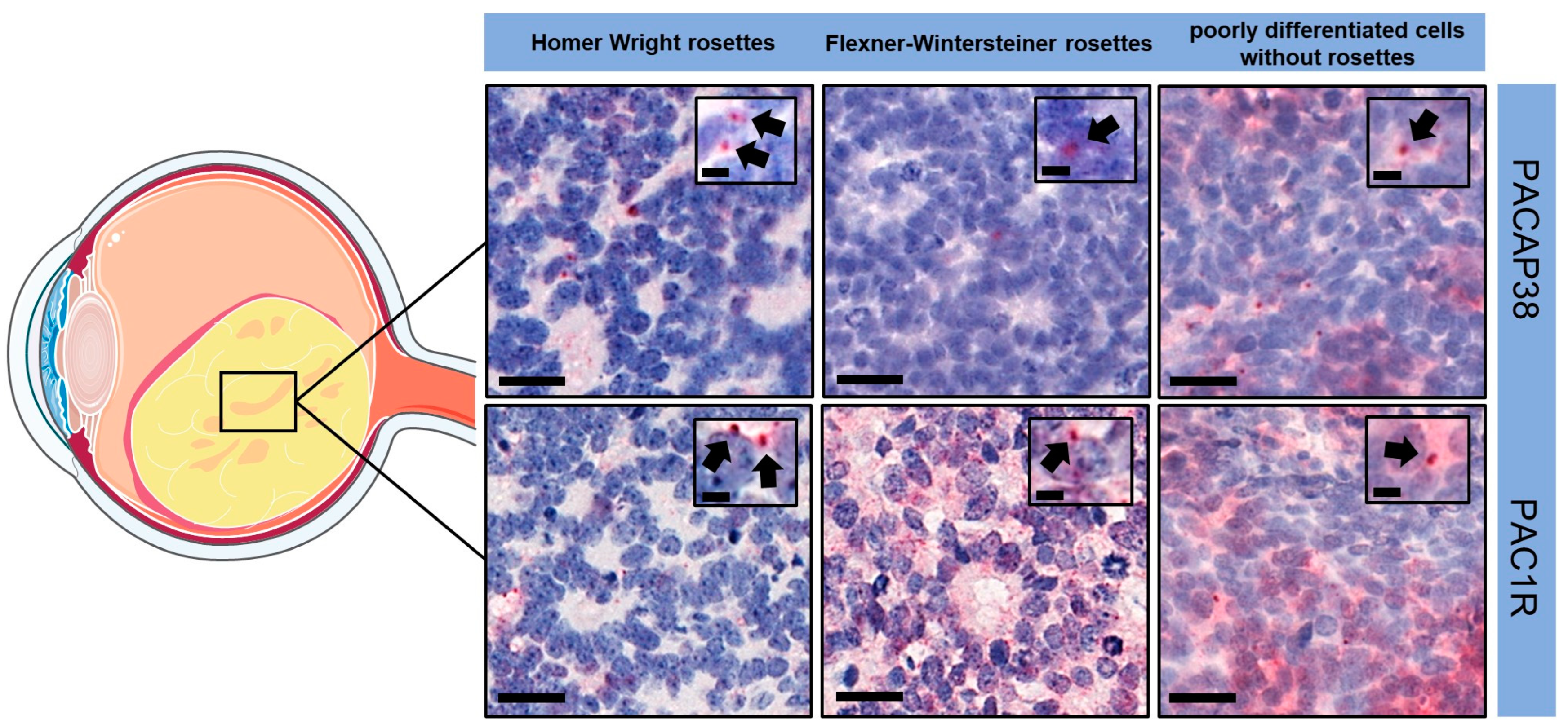
3.1.2. Immunohistochemistry
3.2. Cell Culture
4. Discussion
5. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Yanoff, M.; Sassani, J.W. Retinoblastoma and simulating lesions. In Ocular Pathology, 8th ed.; Yanoff, M., Sassani, J.W., Eds.; Elsevier: London, UK, 2020; pp. 726–759. [Google Scholar]
- Dimaras, H.; Corson, T.W.; Cobrinik, D.; White, A.; Zhao, J.; Munier, F.L.; Abramson, D.H.; Shields, C.L.; Chantada, G.L.; Njuguna, F.; et al. Retinoblastoma. Nat. Rev. Dis. Primers 2015, 1, 15021. [Google Scholar] [CrossRef]
- Orjuela-Grimm, M.; Singh, N.; Bhatt-Carreño, S.; Singh, A.D. Retinoblastoma: Incidence and etiologic factors. In Clinical Ophthalmic Oncology; Berry, J.L., Kim, J.W., Damato, B.E., Singh, A.D., Eds.; Springer International Publishing: Cham, Switzerland, 2019; pp. 39–56. [Google Scholar]
- Dimaras, H.; Kimani, K.; Dimba, E.A.; Gronsdahl, P.; White, A.; Chan, H.S.; Gallie, B.L. Retinoblastoma. Lancet 2012, 379, 1436–1446. [Google Scholar] [CrossRef] [PubMed]
- Gupta, A.K.; Meena, J.P. A narrative review of retinoblastoma and recent advances in its management. Pediatr. Med. 2020, 3, 20. [Google Scholar] [CrossRef]
- Knudson, A.G. Mutation and cancer: Statistical study of retinoblastoma. Proc. Natl. Acad. Sci. USA 1971, 68, 820–823. [Google Scholar] [CrossRef] [PubMed]
- Wong, J.R.; Tucker, M.A.; Kleinerman, R.A.; Devesa, S.S. Retinoblastoma incidence patterns in the US surveillance, epidemiology, and end results program. JAMA Ophthalmol. 2014, 132, 478. [Google Scholar] [CrossRef] [PubMed]
- Rushlow, D.E.; Mol, B.M.; Kennett, J.Y.; Yee, S.; Pajovic, S.; Thériault, B.L.; Prigoda-Lee, N.L.; Spencer, C.; Dimaras, H.; Corson, T.W.; et al. Characterisation of retinoblastomas without RB1 mutations: Genomic, gene expression, and clinical studies. Lancet Oncol. 2013, 14, 327–334. [Google Scholar] [CrossRef] [PubMed]
- Bremner, R.; Sage, J. The origin of human retinoblastoma. Nature 2014, 514, 313. [Google Scholar] [CrossRef]
- Bouchoucha, Y.; Matet, A.; Berger, A.; Carcaboso, A.M.; Gerrish, A.; Moll, A.; Jenkinson, H.; Ketteler, P.; Dorsman, J.C.; Chantada, G.; et al. Retinoblastoma: From genes to patient care. Eur. J. Med. Genet. 2023, 66, 104674. [Google Scholar] [CrossRef]
- Rootman, D.B.; Gonzalez, E.; Mallipatna, A.; VandenHoven, C.; Hampton, L.; Dimaras, H.; Chan, H.S.L.; Gallie, B.L.; Heon, E. Hand-held high-resolution spectral domain optical coherence tomography in retinoblastoma: Clinical and morphologic considerations. Br. J. Ophthalmol. 2013, 97, 59–65. [Google Scholar] [CrossRef]
- Xu, X.L.; Fang, Y.; Lee, T.C.; Forrest, D.; Gregory-Evans, C.; Almeida, D.; Liu, A.; Jhanwar, S.C.; Abramson, D.H.; Cobrinik, D. Retinoblastoma has properties of a cone precursor tumor and depends upon cone-specific MDM2 signaling. Cell 2009, 137, 1018–1031. [Google Scholar] [CrossRef]
- Xu, X.L.; Singh, H.P.; Wang, L.; Qi, D.-L.; Poulos, B.K.; Abramson, D.H.; Jhanwar, S.C.; Cobrinik, D. Rb suppresses human cone-precursor-derived retinoblastoma tumours. Nature 2014, 514, 385–388. [Google Scholar] [CrossRef]
- Kooi, I.E.; Mol, B.M.; Moll, A.C.; Van Der Valk, P.; De Jong, M.C.; De Graaf, P.; Van Mil, S.E.; Schouten-van Meeteren, A.Y.N.; Meijers-Heijboer, H.; Kaspers, G.L.; et al. Loss of photoreceptorness and gain of genomic alterations in retinoblastoma reveal tumor progression. EBioMedicine 2015, 2, 660–670. [Google Scholar] [CrossRef]
- Singh, H.P.; Wang, S.; Stachelek, K.; Lee, S.; Reid, M.W.; Thornton, M.E.; Craft, C.M.; Grubbs, B.H.; Cobrinik, D. Developmental stage-specific proliferation and retinoblastoma genesis in RB-deficient human but not mouse cone precursors. Proc. Natl. Acad. Sci. USA 2018, 115, E9391–E9400. [Google Scholar] [CrossRef] [PubMed]
- Liu, H.; Zhang, Y.; Zhang, Y.-Y.; Li, Y.-P.; Hua, Z.-Q.; Zhang, C.-J.; Wu, K.-C.; Yu, F.; Zhang, Y.; Su, J.; et al. Human embryonic stem cell-derived organoid retinoblastoma reveals a cancerous origin. Proc. Natl. Acad. Sci. USA 2020, 117, 33628–33638. [Google Scholar] [CrossRef] [PubMed]
- Liu, J.; Ottaviani, D.; Sefta, M.; Desbrousses, C.; Chapeaublanc, E.; Aschero, R.; Sirab, N.; Lubieniecki, F.; Lamas, G.; Tonon, L.; et al. A high-risk retinoblastoma subtype with stemness features, dedifferentiated cone states and neuronal/ganglion cell gene expression. Nat. Commun. 2021, 12, 5578. [Google Scholar] [CrossRef]
- Singh, H.P.; Shayler, D.W.H.; Fernandez, G.E.; Thornton, M.E.; Craft, C.M.; Grubbs, B.H.; Cobrinik, D. An immature, dedifferentiated, and lineage-deconstrained cone precursor origin of N-Myc–initiated retinoblastoma. Proc. Natl. Acad. Sci. USA 2022, 119, e2200721119. [Google Scholar] [CrossRef] [PubMed]
- Singh, L.; Kashyap, S. Update on pathology of retinoblastoma. Int. J. Ophthalmol. 2018, 11, 2011–2016. [Google Scholar] [CrossRef]
- Alsharif, H.; Helmi, H.; Maktabi, A. Histopathological characteristics and classification for prognostic indicators. In Retinoblastoma—Past, Present and Future; Manaa Alkatan, H., Ed.; IntechOpen: London, UK, 2019. [Google Scholar] [CrossRef]
- Eagle, R.C. The pathology of ocular cancer. Eye 2013, 27, 128–136. [Google Scholar] [CrossRef]
- Mallipatna, A.C.; Gallie, B.L.; Chévez-Barrios, P.; Rouic, L.L.-L.; Chantada, G.L.; Doz, F.; Brisse, H.J.; Munier, F.L.; Albert, D.M.; Català-Mora, J.; et al. Retinoblastoma. In AJCC Cancer Staging Manual, 8th ed.; Amin, M.B., Edge, S.B., Greene, F.L., Byrd, D.R., Brookland, R.K., Washington, M.K., Gershenwald, J.E., Compton, C.C., Hess, K.R., Sullivan, D.C., et al., Eds.; Springer International Publishing: Cham, Switzerland, 2017; pp. 827–839. [Google Scholar] [CrossRef]
- Martínez-Sánchez, M.; Hernandez-Monge, J.; Rangel, M.; Olivares-Illana, V. Retinoblastoma: From discovery to clinical management. FEBS J. 2022, 289, 4371–4382. [Google Scholar] [CrossRef]
- Manecka, D.-L.; Boukhzar, L.; Falluel-Morel, A.; Lihrmann, I.; Anouar, Y. PACAP signaling in neuroprotection. In Pituitary Adenylate Cyclase Activating Polypeptide—PACAP; Current Topics in Neurotoxicity; Reglodi, D., Tamas, A., Eds.; Springer International Publishing: Cham, Switzerland, 2016; Volume 11, pp. 549–561. [Google Scholar]
- Toth, D.; Szabo, E.; Tamas, A.; Juhasz, T.; Horvath, G.; Fabian, E.; Opper, B.; Szabo, D.; Maugeri, G.; D’Amico, A.G.; et al. Protective effects of PACAP in peripheral organs. Front. Endocrinol. 2020, 11, 377. [Google Scholar] [CrossRef]
- Miyata, A.; Arimura, A.; Dahl, R.R.; Minamino, N.; Uehara, A.; Jiang, L.; Culler, M.D.; Coy, D.H. Isolation of a novel 38 residue-hypothalamic polypeptide which stimulates adenylate cyclase in pituitary cells. Biochem. Biophys. Res. Commun. 1989, 164, 567–574. [Google Scholar] [CrossRef]
- Miyata, A.; Jiang, L.; Dahl, R.D.; Kitada, C.; Kubo, K.; Fujino, M.; Minamino, N.; Arimura, A. Isolation of a neuropeptide corresponding to the n-terminal 27 residues of the pituitary adenylate cyclase activating polypeptide with 38 residues (PACAP38). Biochem. Biophys. Res. Commun. 1990, 170, 643–648. [Google Scholar] [CrossRef]
- Vaudry, D.; Falluel-Morel, A.; Bourgault, S.; Basille, M.; Burel, D.; Wurtz, O.; Fournier, A.; Chow, B.K.C.; Hashimoto, H.; Galas, L.; et al. Pituitary adenylate cyclase-activating polypeptide and its receptors: 20 years after the discovery. Pharmacol. Rev. 2009, 61, 283–357. [Google Scholar] [CrossRef]
- Olianas, M.C.; Ingianni, A.; Sogos, V.; Onali, P. Expression of pituitary adenylate cyclase-activating polypeptide (PACAP) receptors and PACAP in human fetal retina. J. Neurochem. 2002, 69, 1213–1218. [Google Scholar] [CrossRef]
- Patko, E.; Szabo, E.; Toth, D.; Tornoczky, T.; Bosnyak, I.; Vaczy, A.; Atlasz, T.; Reglodi, D. Distribution of PACAP and PAC1 Receptor in the Human Eye. J. Mol. Neurosci. 2022, 72, 2176–2187. [Google Scholar] [CrossRef]
- Reglodi, D.; Tamas, A.; Koppan, M.; Szogyi, D.; Welke, L. Role of PACAP in female fertility and reproduction at gonadal level—Recent advances. Front. Endocrinol. 2012, 3, 155. [Google Scholar] [CrossRef] [PubMed]
- Watanabe, J.; Seki, T.; Shioda, S. PACAP and neural development. In Pituitary Adenylate Cyclase Activating Polypeptide—PACAP; Current Topics in Neurotoxicity; Reglodi, D., Tamas, A., Eds.; Springer International Publishing: Cham, Switzerland, 2016; Volume 11, pp. 65–82. [Google Scholar]
- Reglodi, D.; Atlasz, T.; Szabo, E.; Jungling, A.; Tamas, A.; Juhasz, T.; Fulop, B.D.; Bardosi, A. PACAP deficiency as a model of aging. GeroScience 2018, 40, 437–452. [Google Scholar] [CrossRef]
- Reglodi, D.; Helyes, Z.; Nemeth, J.; Vass, R.A.; Tamas, A. PACAP as a potential biomarker: Alterations of PACAP levels in human physiological and pathological conditions. In Pituitary Adenylate Cyclase Activating Polypeptide—PACAP; Current Topics in Neurotoxicity; Reglodi, D., Tamas, A., Eds.; Springer International Publishing: Cham, Switzerland, 2016; Volume 11, pp. 815–832. [Google Scholar]
- Toth, D.; Reglodi, D.; Schwieters, L.; Tamas, A. Role of endocrine PACAP in age-related diseases. Front. Endocrinol. 2023, 14, 1118927. [Google Scholar] [CrossRef] [PubMed]
- Fabian, E.; Reglodi, D.; Mester, L.; Szabo, A.; Szabadfi, K.; Tamas, A.; Toth, G.; Kovacs, K. Effects of PACAP on intracellular signaling pathways in human retinal pigment epithelial cells exposed to oxidative stress. J. Mol. Neurosci. 2012, 48, 493–500. [Google Scholar] [CrossRef] [PubMed]
- Maugeri, G.; D’Amico, A.G.; Saccone, S.; Federico, C.; Cavallaro, S.; D’Agata, V. PACAP and VIP inhibit HIF-1α-mediated VEGF expression in a model of diabetic macular edema. J. Cell. Physiol. 2017, 232, 1209–1215. [Google Scholar] [CrossRef] [PubMed]
- Maugeri, G.; D’Amico, A.G.; Castrogiovanni, P.; Saccone, S.; Federico, C.; Reibaldi, M.; Russo, A.; Bonfiglio, V.; Avitabile, T.; Longo, A.; et al. PACAP through EGFR transactivation preserves human corneal endothelial integrity. J. Cell. Biochem. 2019, 120, 10097–10105. [Google Scholar] [CrossRef] [PubMed]
- Fabian, E.; Reglodi, D.; Horvath, G.; Opper, B.; Toth, G.; Fazakas, C.; Vegh, A.G.; Wilhelm, I.; Krizbai, I.A. Pituitary adenylate cyclase activating polypeptide acts against neovascularization in retinal pigment epithelial cells. Ann. N. Y. Acad. Sci. 2019, 1455, 160–172. [Google Scholar] [CrossRef] [PubMed]
- Maugeri, G.; D’Amico, A.G.; Bucolo, C.; D’Agata, V. Protective effect of PACAP-38 on retinal pigmented epithelium in an in vitro and in vivo model of diabetic retinopathy through EGFR-dependent mechanism. Peptides 2019, 119, 170108. [Google Scholar] [CrossRef] [PubMed]
- Maugeri, G.; Longo, A.; D’Amico, A.G.; Rasà, D.M.; Reibaldi, M.; Russo, A.; Bonfiglio, V.; Avitabile, T.; D’Agata, V. Trophic effect of PACAP on human corneal endothelium. Peptides 2018, 99, 20–26. [Google Scholar] [CrossRef] [PubMed]
- Atlasz, T.; Szabadfi, K.; Kiss, P.; Tamas, A.; Toth, G.; Reglodi, D.; Gabriel, R. Evaluation of the protective effects of PACAP with cell-specific markers in ischemia-induced retinal degeneration. Brain Res. Bull. 2010, 81, 497–504. [Google Scholar] [CrossRef]
- Vaczy, A.; Kovari, P.; Kovacs, K.; Farkas, K.; Szabo, E.; Kvarik, T.; Kocsis, B.; Fulop, B.; Atlasz, T.; Reglodi, D. Protective role of endogenous PACAP in inflammation-induced retinal degeneration. Curr. Pharm. Des. 2018, 24, 3534–3542. [Google Scholar] [CrossRef] [PubMed]
- Kvarik, T.; Reglodi, D.; Werling, D.; Vaczy, A.; Kovari, P.; Szabo, E.; Kovacs, K.; Hashimoto, H.; Ertl, T.; Gyarmati, J.; et al. The protective effects of endogenous PACAP in oxygen-induced retinopathy. J. Mol. Neurosci. 2021, 71, 2546–2557. [Google Scholar] [CrossRef]
- Szabo, E.; Patko, E.; Vaczy, A.; Molitor, D.; Csutak, A.; Toth, G.; Reglodi, D.; Atlasz, T. Retinoprotective effects of PACAP eye drops in microbead-induced glaucoma model in rats. Int. J. Mol. Sci. 2021, 22, 8825. [Google Scholar] [CrossRef]
- Atlasz, T.; Szabadfi, K.; Reglődi, D.; Kiss, P.; Tamás, A.; Tóth, G.; Molnár, A.; Szabó, K.; Gábriel, R. Effects of pituitary adenylate cyclase activating polypeptide and its fragments on retinal degeneration induced by neonatal monosodium glutamate treatment. Ann. N. Y. Acad. Sci. 2009, 1163, 348–352. [Google Scholar] [CrossRef]
- Seki, T.; Itoh, H.; Nakamachi, T.; Shioda, S. Suppression of ganglion cell death by PACAP following optic nerve transection in the rat. J. Mol. Neurosci. 2008, 36, 57–60. [Google Scholar] [CrossRef]
- Lindholm, D.; Mäkelä, J.; Korhonen, L. PACAP and neural progenitor cells. In Pituitary Adenylate Cyclase Activating Polypeptide—PACAP; Current Topics in Neurotoxicity; Reglodi, D., Tamas, A., Eds.; Springer International Publishing: Cham, Switzerland, 2016; Volume 11, pp. 53–63. [Google Scholar]
- Horvath, G.; Reglodi, D.; Fabian, E.; Opper, B. Effects of pituitary adenylate cyclase activating polypeptide on cell death. Int. J. Mol. Sci. 2022, 23, 4953. [Google Scholar] [CrossRef]
- Denes, V.; Geck, P.; Mester, A.; Gabriel, R. Pituitary adenylate cyclase-activating polypeptide: 30 years in research spotlight and 600 million years in service. J. Clin. Med. 2019, 8, 1488. [Google Scholar] [CrossRef]
- Moody, T.W.; Jensen, R.T. PACAP and cancer. In Pituitary Adenylate Cyclase Activating Polypeptide—PACAP; Current Topics in Neurotoxicity; Reglodi, D., Tamas, A., Eds.; Springer International Publishing: Cham, Switzerland, 2016; Volume 11, pp. 795–814. [Google Scholar]
- Moody, T.W.; Nuche-Berenguer, B.; Jensen, R.T. Vasoactive intestinal peptide/pituitary adenylate cyclase activating polypeptide, and their receptors and cancer. Curr. Opin. Endocrinol. Diabetes Obes. 2016, 23, 38–47. [Google Scholar] [CrossRef] [PubMed]
- Moody, T.W.; Jensen, R.T. Pituitary adenylate cyclase-activating polypeptide/vasoactive intestinal peptide [Part 1]: Biology, pharmacology, and new insights into their cellular basis of action/signaling which are providing new therapeutic targets. Curr. Opin. Endocrinol. Diabetes Obes. 2021, 28, 198–205. [Google Scholar] [CrossRef]
- Zibara, K.; Zeidan, A.; Mallah, K.; Kassem, N.; Awad, A.; Mazurier, F.; Badran, B.; El-Zein, N. Signaling pathways activated by PACAP in MCF-7 breast cancer cells. Cell. Signal. 2018, 50, 37–47. [Google Scholar] [CrossRef] [PubMed]
- Maugeri, G.; D’Amico, A.G.; Saccone, S.; Federico, C.; Rasà, D.M.; Caltabiano, R.; Broggi, G.; Giunta, S.; Musumeci, G.; D’Agata, V. Effect of PACAP on hypoxia-induced angiogenesis and epithelial—Mesenchymal transition in glioblastoma. Biomedicines 2021, 9, 965. [Google Scholar] [CrossRef]
- Lochner, R.; Couce, M. Retinoblastoma. Available online: https://www.pathologyoutlines.com/topic/eyeretinaretinoblastoma.html (accessed on 15 November 2023).
- Bardosi, S.; Bardosi, A.; Nagy, Z.; Reglodi, D. Expression of PACAP and PAC1 receptor in normal human thyroid gland and in thyroid papillary carcinoma. J. Mol. Neurosci. 2016, 60, 171–178. [Google Scholar] [CrossRef]
- García-Fernández, M.O.; Bodega, G.; Ruíz-Villaespesa, A.; Cortés, J.; Prieto, J.C.; Carmena, M.J. PACAP expression and distribution in human breast cancer and healthy tissue. Cancer Lett. 2004, 205, 189–195. [Google Scholar] [CrossRef]
- García-Fernández, M.O.; Collado, B.; Bodega, G.; Cortés, J.; Ruíz-Villaespesa, A.; Carmena, M.J.; Prieto, J.C. Pituitary adenylate cyclase-activating peptide/vasoactive intestinal peptide receptors in human normal mammary gland and breast cancer tissue. Gynecol. Endocrinol. 2005, 20, 327–333. [Google Scholar] [CrossRef]
- Ferencz, S.; Reglodi, D.; Kaszas, B.; Bardosi, A.; Toth, D.; Vekony, Z.; Vicena, V.; Karadi, O.; Kelemen, D. PACAP and PAC1 receptor expression in pancreatic ductal carcinoma. Oncol. Lett. 2019, 18, 5725–5730. [Google Scholar] [CrossRef] [PubMed]
- Ferencz, S.; Toth, D.; Kaszas, B.; Bardosi, S.; Vicena, V.; Karadi, O.; Reglodi, D.; Kelemen, D. PACAP and PAC1 receptor expression in human insulinomas. Int. J. Pept. Res. Ther. 2021, 27, 1719–1728. [Google Scholar] [CrossRef]
- Szanto, Z.; Sarszegi, Z.; Reglodi, D.; Nemeth, J.; Szabadfi, K.; Kiss, P.; Varga, A.; Banki, E.; Csanaky, K.; Gaszner, B.; et al. PACAP immunoreactivity in human malignant tumor samples and cardiac diseases. J. Mol. Neurosci. 2012, 48, 667–673. [Google Scholar] [CrossRef]
- Tamas, A.; Javorhazy, A.; Reglodi, D.; Sarlos, D.P.; Banyai, D.; Semjen, D.; Nemeth, J.; Lelesz, B.; Fulop, D.B.; Szanto, Z. Examination of PACAP-like immunoreactivity in urogenital tumor samples. J. Mol. Neurosci. 2016, 59, 177–183. [Google Scholar] [CrossRef] [PubMed]
- García-Fernández, M.O.; Bodega, G.; Solano, R.M.; Ruíz-Villaespesa, A.; Sánchez-Chapado, M.; Carmena, M.J.; Prieto, J.C. Expression and distribution of pituitary adenylate cyclase-activating peptide in human prostate and prostate cancer tissues. Regul. Pept. 2002, 110, 9–15. [Google Scholar] [CrossRef] [PubMed]
- Olianas, M.C.; Ennas, M.G.; Lampis, G.; Onali, P. Presence of pituitary adenylate cyclase-activating polypeptide receptors in Y-79 human retinoblastoma cells. J. Neurochem. 2002, 67, 1293–1300. [Google Scholar] [CrossRef] [PubMed]
- Wojcieszak, J.; Zawilska, J.B. PACAP38 and PACAP6-38 exert cytotoxic activity against human retinoblastoma y79 cells. J. Mol. Neurosci. 2014, 54, 463–468. [Google Scholar] [CrossRef] [PubMed]
- D’Amico, A.G.; Maugeri, G.; Vanella, L.; Pittalà, V.; Reglodi, D.; D’Agata, V. Multimodal role of PACAP in glioblastoma. Brain Sci. 2021, 11, 994. [Google Scholar] [CrossRef] [PubMed]
- Brubel, R.; Boronkai, A.; Reglodi, D.; Racz, B.; Nemeth, J.; Kiss, P.; Lubics, A.; Toth, G.; Horvath, G.; Varga, T.; et al. Changes in the expression of pituitary adenylate cyclase-activating polypeptide in the human placenta during pregnancy and its effects on the survival of JAR choriocarcinoma cells. J. Mol. Neurosci. 2010, 42, 450–458. [Google Scholar] [CrossRef] [PubMed]
- Horvath, G.; Brubel, R.; Kovacs, K.; Reglodi, D.; Opper, B.; Ferencz, A.; Szakaly, P.; Laszlo, E.; Hau, L.; Kiss, P.; et al. Effects of PACAP on oxidative stress-induced cell death in rat kidney and human hepatocyte cells. J. Mol. Neurosci. 2011, 43, 67–75. [Google Scholar] [CrossRef] [PubMed]
- Farini, D.; Puglianiello, A.; Mammi, C.; Siracusa, G.; Moretti, C. Dual effect of pituitary adenylate cyclase activating polypeptide on prostate tumor LNCaP cells: Short- and long-term exposure affect proliferation and neuroendocrine differentiation. Endocrinology 2003, 144, 1631–1643. [Google Scholar] [CrossRef]
- Doan, N.-D.; Chatenet, D.; Létourneau, M.; Vaudry, H.; Vaudry, D.; Fournier, A. Receptor-independent cellular uptake of pituitary adenylate cyclase-activating polypeptide. Biochim. Biophys. Acta 2012, 1823, 940–949. [Google Scholar] [CrossRef]
- Yu, R.; Zhong, J.; Li, M.; Guo, X.; Zhang, H.; Chen, J. PACAP induces the dimerization of PAC1 on the nucleus associated with the cAMP increase in the nucleus. Neurosci. Lett. 2013, 549, 92–96. [Google Scholar] [CrossRef]
- Reubi, J.C.; Läderach, U.; Waser, B.; Gebbers, J.O.; Robberecht, P.; Laissue, J.A. Vasoactive intestinal peptide/pituitary adenylate cyclase-activating peptide receptor subtypes in human tumors and their tissues of origin. Cancer Res. 2000, 60, 3105–3112. [Google Scholar]
- Bourgault, S.; Vaudry, D.; Botia, B.; Couvineau, A.; Laburthe, M.; Vaudry, H.; Fournier, A. Novel stable PACAP analogs with potent activity towards the PAC1 receptor. Peptides 2008, 29, 919–932. [Google Scholar] [CrossRef] [PubMed]
- Apostol, C.R.; Bernard, K.; Tanguturi, P.; Molnar, G.; Bartlett, M.J.; Szabò, L.; Liu, C.; Ortiz, J.B.; Saber, M.; Giordano, K.R.; et al. Design and synthesis of brain penetrant glycopeptide analogues of PACAP with neuroprotective potential for traumatic brain injury and parkinsonism. Front. Drug Discov. 2022, 1, 818003. [Google Scholar] [CrossRef] [PubMed]
- Werling, D.; Banks, W.; Salameh, T.; Kvarik, T.; Kovacs, L.; Vaczy, A.; Szabo, E.; Mayer, F.; Varga, R.; Tamas, A.; et al. Passage through the ocular barriers and beneficial effects in retinal ischemia of topical application of PACAP1-38 in rodents. Int. J. Mol. Sci. 2017, 18, 675. [Google Scholar] [CrossRef]
- Hirabayashi, T.; Shibato, J.; Kimura, A.; Yamashita, M.; Takenoya, F.; Shioda, S. Potential therapeutic role of pituitary adenylate cyclase-activating polypeptide for dry eye disease. Int. J. Mol. Sci. 2022, 23, 664. [Google Scholar] [CrossRef] [PubMed]
- Boisvilliers, M.D.; Perrin, F.; Hebache, S.; Balandre, A.-C.; Bensalma, S.; Garnier, A.; Vaudry, D.; Fournier, A.; Festy, F.; Muller, J.-M.; et al. VIP and PACAP analogs regulate therapeutic targets in high-risk neuroblastoma cells. Peptides 2016, 78, 30–41. [Google Scholar] [CrossRef]


{kind=link}
{kind=link}
{kind=link}
| Case | Sex | Age at Enucleation (Months) | Eye Involved | Tumor Site | Number of Tumor Foci | RB1 Mutation |
|---|---|---|---|---|---|---|
| 1 | male | 9.9 | right | superotemporal | monofocal | no |
| 2 | female | 16.6 | right | superotemporal | multifocal | no |
| 3 | male | 9.3 | right | superotemporal | monofocal | no |
| 4 | male | 4.1 | right | adjacent to the optic disc | monofocal | yes |
| 5 | male | 36.3 | left | whole eye | monofocal | no |
| 6 | male | 19.9 | left | whole eye | monofocal | no |
| 7 | male | 18.3 | right | whole eye | monofocal | no |
| Case | Histomorphology | Necrosis | Calcification | Mib-1 (%) | Stage |
|---|---|---|---|---|---|
| 1 | tumor cells arranged in sheets, nests, and trabeculae (G4) | large | focal | 80 | pT2 |
| 2 | tumor with many Homer Wright rosettes (G2) | small | focal | 90 | pT2 |
| 3 | tumor with many Homer Wright rosettes (G2) | small | focal | 90 | pT2 |
| 4 | tumor with many Flexner–Wintersteiner rosettes (G2) | large | no | 50–60 | pT2 |
| 5 | tumor with occasional Homer Wight rosettes (G3) | large | focal | 80 | pT2 |
| 6 | tumor cells arranged in sheets, nests, and trabeculae (G4) | large | focal | n.d. | pT2 |
| 7 | tumor cells arranged in sheets, nests, and trabeculae (G4) | large | no | n.d. | pT2 |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2024 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Tóth, D.; Fábián, E.; Szabó, E.; Patkó, E.; Vicena, V.; Váczy, A.; Atlasz, T.; Tornóczky, T.; Reglődi, D. Investigation of PACAP38 and PAC1 Receptor Expression in Human Retinoblastoma and the Effect of PACAP38 Administration on Human Y-79 Retinoblastoma Cells. Life 2024, 14, 185. https://doi.org/10.3390/life14020185
Tóth D, Fábián E, Szabó E, Patkó E, Vicena V, Váczy A, Atlasz T, Tornóczky T, Reglődi D. Investigation of PACAP38 and PAC1 Receptor Expression in Human Retinoblastoma and the Effect of PACAP38 Administration on Human Y-79 Retinoblastoma Cells. Life. 2024; 14(2):185. https://doi.org/10.3390/life14020185
Chicago/Turabian StyleTóth, Dénes, Eszter Fábián, Edina Szabó, Evelin Patkó, Viktória Vicena, Alexandra Váczy, Tamás Atlasz, Tamás Tornóczky, and Dóra Reglődi. 2024. "Investigation of PACAP38 and PAC1 Receptor Expression in Human Retinoblastoma and the Effect of PACAP38 Administration on Human Y-79 Retinoblastoma Cells" Life 14, no. 2: 185. https://doi.org/10.3390/life14020185
APA StyleTóth, D., Fábián, E., Szabó, E., Patkó, E., Vicena, V., Váczy, A., Atlasz, T., Tornóczky, T., & Reglődi, D. (2024). Investigation of PACAP38 and PAC1 Receptor Expression in Human Retinoblastoma and the Effect of PACAP38 Administration on Human Y-79 Retinoblastoma Cells. Life, 14(2), 185. https://doi.org/10.3390/life14020185