
Lithium: A Promising Anticancer Agent
 ,
,  ,
,  , ,
, ,  , , and
, , and 
Abstract
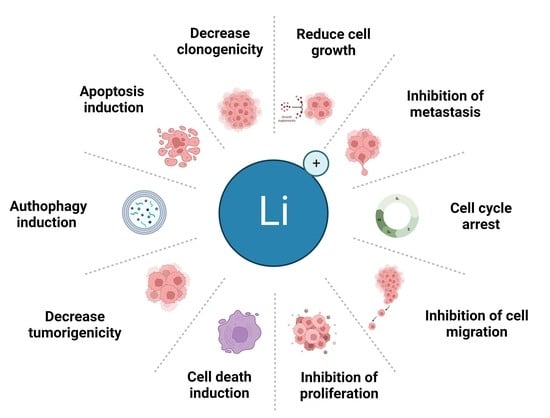
1. Introduction
2. The Effect of Lithium on Cancer Progression: Preclinical Approach
3. Lithium Transport
Lithium Transporters in Cancer
4. Lithium as a Specific Inhibitor of GSK3β
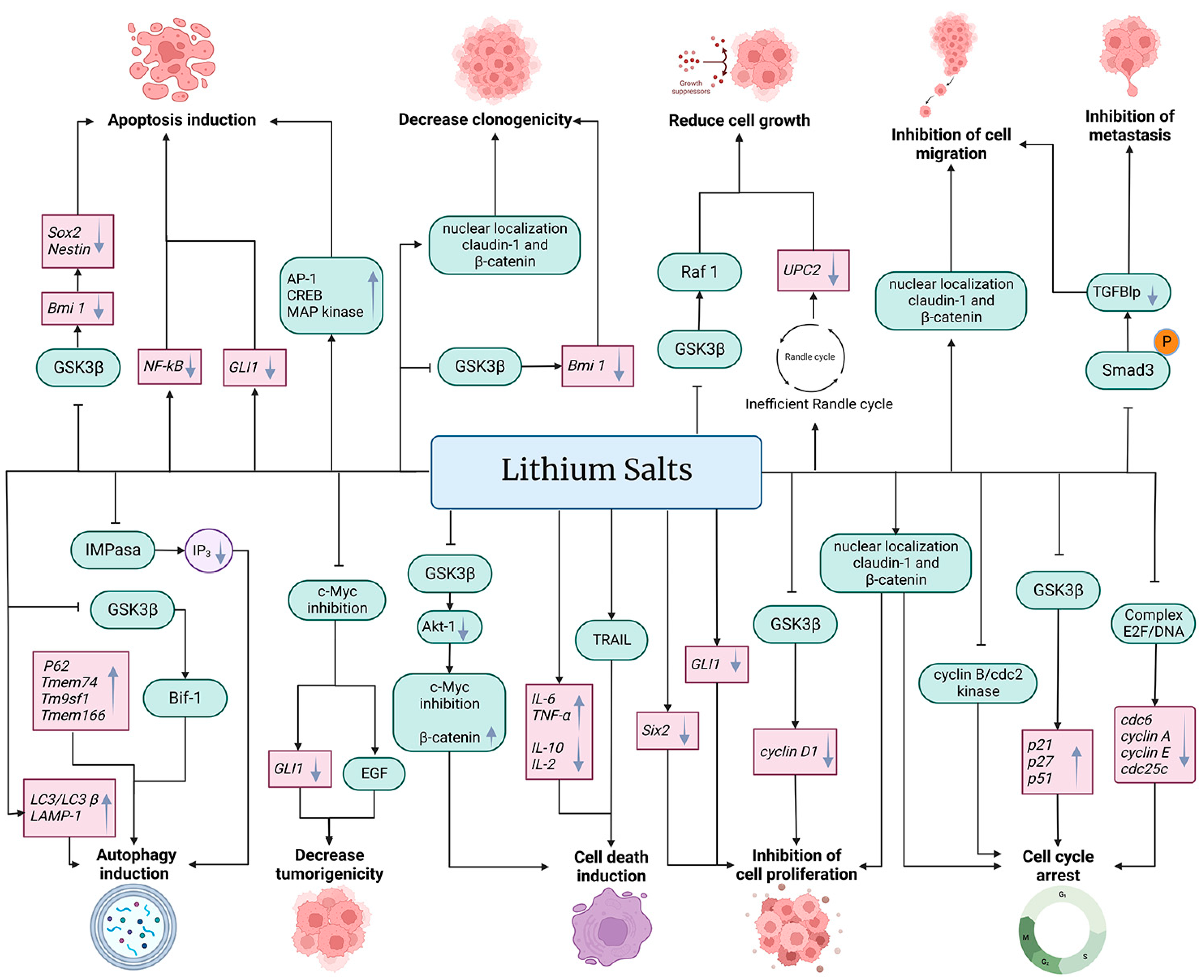
5. Effect of Lithium on Certain Cancer Hallmarks
5.1. Effect of Lithium on Apoptosis
5.2. Effect of Lithium on Autophagy
5.3. Effect of Lithium on Tumor Growth, Tumor Proliferation, Tumor Invasion and Metastasis, and Cell Cycle Arrest
6. Anti-Inflammatory Activity of Lithium
7. Synergism of Lithium with Standard Cancer Therapies
8. Recent Clinical Studies
9. Trends in the Administration of Lithium
10. Contradictory Effects of Lithium
11. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Hanahan, D. Hallmarks of cancer: New dimensions. Cancer Discov. 2022, 12, 31–46. [Google Scholar] [CrossRef]
- Warburg, O. The Metabolism of Tumors; Richard R. Smith. Inc.: New York, NY, USA, 1931. [Google Scholar]
- Poff, A.; Koutnik, A.P.; Egan, K.M.; Sahebjam, S.; D’Agostino, D.; Kumar, N.B. Targeting the Warburg effect for cancer treatment: Ketogenic diets for management of glioma. In Seminars in Cancer Biology; Elsevier: Amsterdam, The Netherlands, 2019. [Google Scholar]
- Won, E.; Kim, Y.-K. An oldie but goodie: Lithium in the treatment of bipolar disorder through neuroprotective and neurotrophic mechanisms. Int. J. Mol. Sci. 2017, 18, 2679. [Google Scholar] [CrossRef] [PubMed]
- Elbe, D. Stahl’s Essential Psychopharmacology: Neuroscientific Basis and Practical Applications. J. Can. Acad. Child Adolesc. Psychiatry 2010, 19, 230. [Google Scholar]
- de Freitas, D.M.; Leverson, B.D.; Goossens, J.L. Lithium in medicine: Mechanisms of action. In The Alkali Metal Ions: Their Role for Life; Springer: Berlin/Heidelberg, Germany, 2016; pp. 557–584. [Google Scholar]
- Edition, F. Diagnostic and statistical manual of mental disorders. Am. Psychiatr. Assoc. 2013, 21, 591–643. [Google Scholar]
- Walker, E.R.; McGee, R.E.; Druss, B.G. Mortality in mental disorders and global disease burden implications: A systematic review and meta-analysis. JAMA Psychiatry 2015, 72, 334–341. [Google Scholar] [CrossRef] [PubMed]
- Wong, M.M. Management of bipolar II disorder. Indian J. Psychol. Med. 2011, 33, 18–28. [Google Scholar] [CrossRef] [PubMed]
- Kessing, L.V.; Søndergård, L.; Forman, J.L.; Andersen, P.K. Lithium treatment and risk of dementia. Arch. Gen. Psychiatry 2008, 65, 1331–1335. [Google Scholar] [CrossRef]
- Nitrini, R.; Caramelli, P.; Herrera, E.; Bahia, V.S.; Caixeta, L.F.; Radanovic, M.; Anghinah, R.; Charchat-Fichman, H.; Porto, C.S.; Carthery, M.T.; et al. Incidence of dementia in a community-dwelling Brazilian population. Alzheimer Dis. Assoc. Disord. 2004, 18, 241–246. [Google Scholar]
- Köhrle, J. Local activation and inactivation of thyroid hormones: The deiodinase family. Mol. Cell. Endocrinol. 1999, 151, 103–119. [Google Scholar] [CrossRef]
- Berens, S.C.; Bernstein, R.S.; Robbins, J.; Wolff, J. Antithyroid effects of lithium. J. Clin. Investig. 1970, 49, 1357–1367. [Google Scholar] [CrossRef]
- Kurita, M.; Mashiko, H.; Rai, M.; Kumasaka, T.; Kouno, S.-I.; Niwa, S.-I.; Nakahata, N. Lithium chloride at a therapeutic concentration reduces Ca2+ response in protein kinase C down-regulated human astrocytoma cells. Eur. J. Pharmacol. 2002, 442, 17–22. [Google Scholar] [CrossRef]
- Briggs, K.T.; Giulian, G.G.; Li, G.; Kao, J.P.; Marino, J.P. A A Molecular Model for Lithium’s Bioactive Form. Biophys. J. 2016, 111, 294–300. [Google Scholar] [CrossRef] [PubMed]
- Volkmann, C.; Bschor, T.; Köhler, S. Lithium treatment over the lifespan in bipolar disorders. Front. Psychiatry 2020, 11, 377. [Google Scholar] [CrossRef]
- Ochoa, E.L.M. Lithium as a neuroprotective agent for bipolar disorder: An overview. Cell. Mol. Neurobiol. 2022, 42, 85–97. [Google Scholar] [CrossRef]
- Vidali, S.; Aminzadeh-Gohari, S.; Vatrinet, R.; Iommarini, L.; Porcelli, A.M.; Kofler, B.; Feichtinger, R.G. Lithium and not acetoacetate influences the growth of cells treated with lithium acetoacetate. Int. J. Mol. Sci. 2019, 20, 3104. [Google Scholar] [CrossRef]
- Hossein, G.; Janzamin, E.; Azimian-Zavareh, V. Effect of lithium chloride and antineoplastic drugs on survival and cell cycle of androgen-dependent prostate cancer LNCap cells. Indian J. Pharmacol. 2012, 44, 714. [Google Scholar] [CrossRef]
- Konenkov, V.I.; Borodin, Y.I.; Makarova, O.P.; Bgatova, N.P.; Rachkovskaya, L.N. Effects of Nanosized Lithium Carbonate Particles on the Functional Activity of Macrophages during Development of Hepatocarcinoma 29. Bull. Exp. Biol. Med. 2015, 159, 490–493. [Google Scholar] [CrossRef] [PubMed]
- O’Donovan, T.R.; Rajendran, S.; O’Reilly, S.; O’Sullivan, G.C.; McKenna, S.L. Lithium modulates autophagy in esophageal and colorectal cancer cells and enhances the efficacy of therapeutic agents in vitro and in vivo. PLoS ONE 2015, 10, e0134676. [Google Scholar] [CrossRef] [PubMed]
- Bgatova, N.P.; Gavrilova, Y.S.; Lykov, A.P.; Solovieva, A.O.; Makarova, V.; Borodin, Y.I.; Konenkov, V.I. Apoptosis and autophagy in hepatocarcinoma cells induced by different forms of lithium salts. Cell Tissue Biol. 2017, 11, 261–267. [Google Scholar] [CrossRef]
- Han, S.; Meng, L.; Jiang, Y.; Cheng, W.; Tie, X.; Xia, J.; Wu, A. Lithium enhances the antitumour effect of temozolomide against TP53 wild-type glioblastoma cells via NFAT1/FasL signalling. Br. J. Cancer 2017, 116, 1302–1311. [Google Scholar] [CrossRef]
- Yao, R.; Sun, X.; Xie, Y.; Liu, L.; Han, D.; Yao, Y.; Li, H.; Li, Z.; Xu, K. Lithium chloride inhibits cell survival, overcomes drug resistance, and triggers apoptosis in multiple myeloma via activation of the Wnt/β-catenin pathway. Am. J. Transl. Res. 2018, 10, 2610. [Google Scholar]
- Razmi, M.; Rabbani-Chadegani, A.; Hashemi-Niasari, F.; Ghadam, P. Lithium chloride attenuates mitomycin C induced necrotic cell death in MDA-MB-231 breast cancer cells via HMGB1 and Bax signaling. J. Trace Elem. Med. Biol. 2018, 48, 87–96. [Google Scholar] [CrossRef]
- Vijay, G.V.; Na Zhao, N.; Hollander, P.D.; Toneff, M.J.; Joseph, R.; Pietila, M.; Taube, J.H.; Sarkar, T.R.; Ramirez-Pena, E.; Werden, S.J.; et al. GSK3β regulates epithelial-mesenchymal transition and cancer stem cell properties in triple-negative breast cancer. Breast Cancer Res. 2019, 21, 37. [Google Scholar] [CrossRef]
- Ramshini, S.; Omidi, M. The Psychiatric Drug Lithium Increases DNA Damage and Decreases Cell Survival in MCF-7 and MDA-MB-231 Breast Cancer Cell Lines Expos ed to Ionizing Radiation. Curr. Mol. Pharmacol. 2019, 12, 301–310. [Google Scholar]
- Xue, D.; Yang, P.; Wei, Q.; Li, X.; Lin, L.; Lin, T. IL-21/IL-21R inhibit tumor growth and invasion in non-small cell lung cancer cells via suppressing Wnt/β-catenin signaling and PD-L1 expression. Int. J. Mol. Med. 2019, 44, 1697–1706. [Google Scholar] [CrossRef] [PubMed]
- Chen, C.; Shan, H. Keratin 6A gene silencing suppresses cell invasion and metastasis of nasopharyngeal carcinoma via the β-catenin cascade. Mol. Med. Rep. 2019, 19, 3477–3484. [Google Scholar] [CrossRef]
- ohen-Harazi, R.; Hofmann, S.; Kogan, V.; Fulman-Levy, H.; Abaev, K.; Shovman, O.; Brider, T.; Koman, I. Cytotoxicity of Exogenous Acetoacetate in Lithium Salt Form Is Mediated by Lithium and Not Acetoacetate. Anticancer Res. 2020, 40, 3831–3837. [Google Scholar] [CrossRef] [PubMed]
- Cammarota, F.; Conte, A.; Aversano, A.; Muto, P.; Ametrano, G.; Riccio, P.; Turano, M.; Valente, V.; DelRio, P.; Izzo, P.; et al. Lithium chloride increases sensitivity to photon irradiation treatment in primary mesenchymal colon cancer cells. Mol. Med. Rep. 2020, 21, 1501–1508. [Google Scholar] [CrossRef] [PubMed]
- Zubčić, V.; Rinčić, N.; Kurtović, M.; Trnski, D.; Musani, V.; Ozretić, P.; Levanat, S.; Leović, D.; Sabol, M. GANT61 and lithium chloride inhibit the growth of head and neck cancer cell lines through the regulation of GLI3 processing by GSK3β. Int. J. Mol. Sci. 2020, 21, 6410. [Google Scholar] [CrossRef] [PubMed]
- Taskaeva, Y.S.; Bgatova, N.P.; Dossymbekova, R.S.; Solovieva, A.O.; Miroshnichenko, S.M.; Sharipov, K.O.; Tungushbaeva, Z.B. In vitro effects of lithium carbonate on cell cycle, apoptosis, and autophagy in hepatocellular carcinoma-29 cells. Bull. Exp. Biol. Med. 2020, 170, 246–250. [Google Scholar] [CrossRef]
- Yang, B.; Wang, S.; Xie, H.; Wang, C.; Gao, X.; Rong, Y.; Liu, Z.; Lu, Y. KIF18B promotes hepatocellular carcinoma progression through activating Wnt/β-catenin-signaling pathway. J. Cell. Physiol. 2020, 235, 6507–6514. [Google Scholar] [CrossRef] [PubMed]
- Chen, H.; Zhao, Y.; Zhang, J.; Xie, Y.; Jin, M. Promoting effects of MiR-135b on human multiple myeloma cells via regulation of the Wnt/β-catenin/Versican signaling pathway. Cytokine 2021, 142, 155495. [Google Scholar] [CrossRef]
- Kim, H.; Li, S.; Lee, D.; Park, J.H.; Muramatsu, T.; Harada, H.; Jung, Y.; Jung, H. Activation of Wnt signalling reduces the population of cancer stem cells in ameloblastoma. Cell Prolif. 2021, 54, e13073. [Google Scholar] [CrossRef] [PubMed]
- Taskaeva, I.; Gogaeva, I.; Shatruk, A.; Bgatova, N. Lithium Enhances Autophagy and Cell Death in Skin Melanoma: An Ultrastructural and Immunohistochemical Study. Microsc. Microanal. 2022, 28, 1703–1711. [Google Scholar] [CrossRef] [PubMed]
- Jakobsson, E.; Argüello-Miranda, O.; Chiu, S.-W.; Fazal, Z.; Kruczek, J.; Nunez-Corrales, S.; Pandit, S.; Pritchet, L. Towards a unified understanding of lithium action in basic biology and its significance for applied biology. J. Membr. Biol. 2017, 250, 587–604. [Google Scholar] [CrossRef] [PubMed]
- de Roos, N.M.; de Vries, J.H.; Katan, M.B. Serum lithium as a compliance marker for food and supplement intake. Am. J. Clin. Nutr. 2001, 73, 75–79. [Google Scholar] [CrossRef]
- Pérez-Escuredo, J.; Van Hée, V.F.; Sboarina, M.; Falces, J.; Payen, V.L.; Pellerin, L.; Sonveaux, P. Monocarboxylate transporters in the brain and in cancer. Biochim. Biophys. Acta Mol. Cell Res. 2016, 1863, 2481–2497. [Google Scholar] [CrossRef]
- Bröer, S.; Bröer, A.; Schneider, H.-P.; Stegen, C.; Halestrap, A.P.; Deitmer, J.W. Characterization of the high-affinity monocarboxylate transporter MCT2 in Xenopus laevis oocytes. Biochem. J. 1999, 341, 529–535. [Google Scholar] [CrossRef]
- Bröer, S.; Schneider, H.-P.; Bröer, A.; Rahman, B.; Hamprecht, B.; Deitmer, J.W. Characterization of the monocarboxylate transporter 1 expressed in Xenopus laevis oocytes by changes in cytosolic pH. Biochem. J. 1998, 333, 167–174. [Google Scholar] [CrossRef]
- Halestrap, A.P. The monocarboxylate transporter family—Structure and functional characterization. IUBMB Life 2012, 64, 1–9. [Google Scholar] [CrossRef]
- Garcia, C.K.; Brown, M.S.; Pathak, R.K.; Goldstein, J.L. cDNA Cloning of MCT2, a Second Monocarboxylate Transporter Expressed in Different Cells than MCT1. J. Biol. Chem. 1995, 270, 1843–1849. [Google Scholar] [CrossRef] [PubMed]
- Halestrap, A.P.; Meredith, D. The SLC16 gene family—From monocarboxylate transporters (MCTs) to aromatic amino acid transporters and beyond. Pflügers Arch. 2004, 447, 619–628. [Google Scholar] [CrossRef] [PubMed]
- Garcia, C.K.; Goldstein, J.L.; Pathak, R.K.; Anderson, R.G.; Brown, M.S. Molecular characterization of a membrane transporter for lactate, pyruvate, and other monocarboxylates: Implications for the Cori cycle. Cell 1994, 76, 865–873. [Google Scholar] [CrossRef] [PubMed]
- Jóhannsson, E.; Nagelhus, E.A.; Mccullagh, K.J.; Sejersted, O.M.; Blackstad, T.W.; Bonen, A.; Ottersen, O.P. Cellular and subcellular expression of the monocarboxylate transporter MCT1 in rat heart: A high-resolution immunogold analysis. Circ. Res. 1997, 80, 400–407. [Google Scholar] [CrossRef]
- Kirat, D.; Inoue, H.; Iwano, H.; Yokota, H.; Taniyama, H.; Kato, S. Monocarboxylate transporter 1 (MCT1) in the liver of pre-ruminant and adult bovines. Vet. J. 2007, 173, 124–130. [Google Scholar] [CrossRef]
- Tamai, I.; Takanaga, H.; Ogihara, T.; Higashida, H.; Maeda, H.; Sai, Y.; Tsuji, A. Participation of a proton-cotransporter, MCT1, in the intestinal transport of monocarboxylic acids. Biochem. Biophys. Res. Commun. 1995, 214, 482–489. [Google Scholar] [CrossRef]
- Ritzhaupt, A.; Wood, I.S.; Ellis, A.; Hosie, K.B.; Shirazi-Beechey, S.P. Identification and characterization of a monocarboxylate transporter (MCT1) in pig and human colon: Its potential to transport l-lactate as well as butyrate. J. Physiol. 1998, 513, 719–732. [Google Scholar] [CrossRef] [PubMed]
- Orsenigo, M.N.; Tosco, M.; Bazzini, C.; Laforenza, U.; Faelli, A. A monocarboxylate transporter MCT1 is located at the basolateral pole of rat jejunum. Exp. Physiol. 1999, 84, 1033–1042. [Google Scholar] [CrossRef]
- Kirat, D.; Inoue, H.; Iwano, H.; Hirayama, K.; Yokota, H.; Taniyama, H.; Kato, S. Monocarboxylate transporter 1 gene expression in the ovine gastrointestinal tract. Vet. J. 2006, 171, 462–467. [Google Scholar] [CrossRef]
- Iwanaga, T.; Takebe, K.; Kato, I.; Karaki, S.-I.; Kuwahara, A. Cellular expression of monocarboxylate transporters (MCT) in the digestive tract of the mouse, rat, and humans, with special reference to slc5a8. Biomed. Res. 2006, 27, 243–254. [Google Scholar] [CrossRef]
- Shimoyama, Y.; Kirat, D.; Akihara, Y.; Kawasako, K.; Komine, M.; Hirayama, K.; Matsuda, K.; Okamoto, M.; Iwano, H.; Kato, S.; et al. Expression of monocarboxylate transporter 1 (MCT1) in the dog intestine. J. Vet. Med. Sci. 2007, 69, 599–604. [Google Scholar] [CrossRef] [PubMed]
- Welter, H.; Claus, R. Expression of the monocarboxylate transporter 1 (MCT1) in cells of the porcine intestine. Cell Biol. Int. 2008, 32, 638–645. [Google Scholar] [CrossRef] [PubMed]
- Poole, R.C.; Halestrap, A.P. N-terminal protein sequence analysis of the rabbit erythrocyte lactate transporter suggests identity with the cloned monocarboxylate transport protein MCT1. Biochem. J. 1994, 303, 755–759. [Google Scholar] [CrossRef]
- de Heredia, F.P.; Wood, I.S.; Trayhurn, P. Hypoxia stimulates lactate release and modulates monocarboxylate transporter (MCT1, MCT2, and MCT4) expression in human adipocytes. Pflügers Arch.-Eur. J. Physiol. 2010, 459, 509–518. [Google Scholar] [CrossRef] [PubMed]
- Hajduch, E.; Heyes, R.R.; Watt, P.W.; Hundal, H.S. Lactate transport in rat adipocytes: Identification of monocarboxylate transporter 1 (MCT1) and its modulation during streptozotocin-induced diabetes. FEBS Lett. 2000, 479, 89–92. [Google Scholar] [CrossRef] [PubMed]
- Takebe, K.; Nio-Kobayashi, J.; Takahashi-Iwanaga, H.; Yajima, T.; Iwanaga, T. Cellular expression of a monocarboxylate transporter (MCT1) in the mammary gland and sebaceous gland of mice. Histochem. Cell Biol. 2009, 131, 401–409. [Google Scholar] [CrossRef] [PubMed]
- Mac, M.; Nałęcz, K.A. Expression of monocarboxylic acid transporters (MCT) in brain cells: Implication for branched chain α-ketoacids transport in neurons. Neurochem. Int. 2003, 43, 305–309. [Google Scholar] [CrossRef]
- Hanu, R.; McKenna, M.; O’Neill, A.; Resneck, W.G.; Bloch, R.J. Monocarboxylic acid transporters, MCT1 and MCT2, in cortical astrocytes in vitro and in vivo. Am. J. Physiol. Physiol. 2000, 278, C921–C930. [Google Scholar] [CrossRef]
- Lee, Y.; Morrison, B.M.; Li, Y.; Lengacher, S.; Farah, M.H.; Hoffman, P.N.; Liu, Y.; Tsingalia, A.; Jin, L.; Zhang, P.-W.; et al. Oligodendroglia metabolically support axons and contribute to neurodegeneration. Nature 2012, 487, 443–448. [Google Scholar] [CrossRef]
- Moreira, T.J.; Pierre, K.; Maekawa, F.; Repond, C.; Cebere, A.; Liljequist, S.; Pellerin, L. Enhanced cerebral expression of MCT1 and MCT2 in a rat ischemia model occurs in activated microglial cells. J. Cereb. Blood Flow Metab. 2009, 29, 1273–1283. [Google Scholar] [CrossRef] [PubMed]
- Ainscow, E.K.; Mirshamsi, S.; Tang, T.; Ashford, M.L.; Rutter, G.A. Dynamic imaging of free cytosolic ATP concentration during fuel sensing by rat hypothalamic neurones: Evidence for ATP-independent control of ATP-sensitive K+ channels. J. Physiol. 2002, 544, 429–445. [Google Scholar] [CrossRef] [PubMed]
- Domènech-Estévez, E.; Baloui, H.; Repond, C.; Rosafio, K.; Médard, J.-J.; Tricaud, N.; Pellerin, L.; Chrast, R. Distribution of monocarboxylate transporters in the peripheral nervous system suggests putative roles in lactate shuttling and myelination. J. Neurosci. 2015, 35, 4151–4156. [Google Scholar] [CrossRef] [PubMed]
- Morrison, B.M.; Tsingalia, A.; Vidensky, S.; Lee, Y.; Jin, L.; Farah, M.H.; Lengacher, S.; Magistretti, P.J.; Pellerin, L.; Rothstein, J.D. Deficiency in monocarboxylate transporter 1 (MCT1) in mice delays regeneration of peripheral nerves following sciatic nerve crush. Exp. Neurol. 2015, 263, 325–338. [Google Scholar] [CrossRef]
- Boussouar, F.; Mauduit, C.; Tabone, E.; Pellerin, L.; Magistretti, P.J.; Benahmed, M. Developmental and hormonal regulation of the monocarboxylate transporter 2 (MCT2) expression in the mouse germ cells. Biol. Reprod. 2003, 69, 1069–1078. [Google Scholar] [CrossRef] [PubMed]
- Pierre, K.; Pellerin, L.; Debernardi, R.; Riederer, B.; Magistretti, P.J. Cell-specific localization of monocarboxylate transporters, MCT1 and MCT2, in the adult mouse brain revealed by double immunohistochemical labeling and confocal microscopy. Neuroscience 2000, 100, 617–627. [Google Scholar] [CrossRef]
- Pellerin, L.; Halestrap, A.P.; Pierre, K. Cellular and subcellular distribution of monocarboxylate transporters in cultured brain cells and in the adult brain. J. Neurosci. Res. 2005, 79, 55–64. [Google Scholar] [CrossRef] [PubMed]
- Pierre, K.; Magistretti, P.J.; Pellerin, L. MCT2 is a major neuronal monocarboxylate transporter in the adult mouse brain. J. Cereb. Blood Flow Metab. 2002, 22, 586–595. [Google Scholar] [CrossRef]
- Reiser, G.; Duhm, J. Transport pathways for lithium ions in neuroblastoma× glioma hybrid cells at ‘therapeutic’concentrations of Li+. Brain Res. 1982, 252, 247–258. [Google Scholar] [CrossRef]
- Sonveaux, P.; Vegran, F.; Schroeder, T.; Wergin, M.C.; Verrax, J.; Rabbani, Z.N.; De Saedeleer, C.J.; Kennedy, K.M.; Diepart, C.; Jordan, B.F.; et al. Targeting lactate-fueled respiration selectively kills hypoxic tumor cells in mice. J. Clin. Investig. 2008, 118, 3930–3942. [Google Scholar] [CrossRef] [PubMed]
- Baltazar, F.; Pinheiro, C.; Morais-Santos, F.; Azevedo-Silva, J.; Queiros, O.; Preto, A.; Casal, M. Monocarboxylate transporters as targets and mediators in cancer therapy response. Histol. Histopathol. 2014, 29, 1511–1524. [Google Scholar]
- Pinheiro, C.; Reis, R.M.; Ricardo, S.; Longatto-Filho, A.; Schmitt, F.; Baltazar, F. Expression of monocarboxylate transporters 1, 2, and 4 in human tumours and their association with CD147 and CD44. J. Biomed. Biotechnol. 2010, 2010, 427694. [Google Scholar] [CrossRef]
- Kennedy, K.M.; Dewhirst, M.W. Tumor metabolism of lactate: The influence and therapeutic potential for MCT and CD147 regulation. Future Oncol. 2010, 6, 127–148. [Google Scholar] [CrossRef]
- Pinheiro, C.; Longatto-Filho, A.; Azevedo-Silva, J.; Casal, M.; Schmitt, F.C.; Baltazar, F. Role of monocarboxylate transporters in human cancers: State of the art. J. Bioenerg. Biomembr. 2012, 44, 127–139. [Google Scholar] [CrossRef]
- Miranda-Gonçalves, V.; Honavar, M.; Pinheiro, C.; Martinho, O.; Pires, M.M.; Pinheiro, C.; Cordeiro, M.; Bebiano, G.; Costa, P.; Palmeirim, I.; et al. Monocarboxylate transporters (MCTs) in gliomas: Expression and exploitation as therapeutic targets. Neuro-Oncol. 2013, 15, 172–188. [Google Scholar] [CrossRef] [PubMed]
- Afonso, J.; Santos, L.L.; Miranda-Gonçalves, V.; Morais, A.; Amaro, T.; Longatto-Filho, A.; Baltazar, F. CD147 and MCT1-potential partners in bladder cancer aggressiveness and cisplatin resistance. Mol. Carcinog. 2015, 54, 1451–1466. [Google Scholar] [CrossRef] [PubMed]
- Pértega-Gomes, N.; Baltazar, F. Lactate transporters in the context of prostate cancer metabolism: What do we know? Int. J. Mol. Sci. 2014, 15, 18333–18348. [Google Scholar] [CrossRef]
- Eilertsen, M.; Andersen, S.; Al-Saad, S.; Kiselev, Y.; Donnem, T.; Stenvold, H.; Pettersen, I.; Al-Shibli, K.; Richardsen, E.; Busund, L.-T.; et al. Monocarboxylate transporters 1–4 in NSCLC: MCT1 is an independent prognostic marker for survival. PLoS ONE 2014, 9, e105038. [Google Scholar] [CrossRef] [PubMed]
- Izumi, H.; Takahashi, M.; Uramoto, H.; Nakayama, Y.; Oyama, T.; Wang, K.-Y.; Sasaguri, Y.; Nishizawa, S.; Kohno, K. Monocarboxylate transporters 1 and 4 are involved in the invasion activity of human lung cancer cells. Cancer Sci. 2011, 102, 1007–1013. [Google Scholar] [CrossRef]
- Dhup, S.; Dadhich, R.K.; Porporato, P.E.; Sonveaux, P. Multiple biological activities of lactic acid in cancer: Influences on tumor growth, angiogenesis and metastasis. Curr. Pharm. Des. 2012, 18, 1319–1330. [Google Scholar] [CrossRef]
- Morris, A.A.M. Cerebral ketone body metabolism. J. Inherit. Metab. Dis. 2004, 28, 109–121. [Google Scholar] [CrossRef]
- Bollag, G.; Tsai, J.; Zhang, J.; Zhang, C.; Ibrahim, P.; Nolop, K.; Hirth, P. Vemurafenib: The first drug approved for BRAF-mutant cancer. Nat. Rev. Drug Discov. 2012, 11, 873–886. [Google Scholar] [CrossRef] [PubMed]
- Benlloch, S.; Payá, A.; Alenda, C.; Bessa, X.; Andreu, M.; Jover, R.; Castells, A.; Llor, X.; Aranda, F.I.; Massutí, B. Detection of BRAF V600E mutation in colorectal cancer: Comparison of automatic sequencing and real-time chemistry methodology. J. Mol. Diagn. 2006, 8, 540–543. [Google Scholar] [CrossRef] [PubMed]
- Arcaini, L.; Zibellini, S.; Boveri, E.; Riboni, R.; Rattotti, S.; Varettoni, M.; Guerrera, M.L.; Lucioni, M.; Tenore, A.; Merli, M.; et al. The BRAF V600E mutation in hairy cell leukemia and other mature B-cell neoplasms. Blood 2012, 119, 188–191. [Google Scholar] [CrossRef] [PubMed]
- Chapman, M.A.; Lawrence, M.S.; Keats, J.J.; Cibulskis, K.; Sougnez, C.; Schinzel, A.C.; Harview, C.L.; Brunet, J.-P.; Ahmann, G.J.; Adli, M.; et al. Initial genome sequencing and analysis of multiple myeloma. Nature 2011, 471, 467–472. [Google Scholar] [CrossRef]
- Kang, H.-B.; Fan, J.; Lin, R.; Elf, S.; Ji, Q.; Zhao, L.; Jin, L.; Seo, J.H.; Shan, C.; Arbiser, J.L.; et al. Metabolic rewiring by oncogenic BRAF V600E links ketogenesis pathway to BRAF-MEK1 signaling. Mol. Cell 2015, 59, 345–358. [Google Scholar] [CrossRef] [PubMed]
- Ussar, S.; Voss, T. MEK1 and MEK2, different regulators of the G1/S transition. J. Biol. Chem. 2004, 279, 43861–43869. [Google Scholar] [CrossRef]
- De Saedeleer, C.J.; Copetti, T.; Porporato, P.E.; Verrax, J.; Feron, O.; Sonveaux, P. Lactate activates HIF-1 in oxidative but not in Warburg-phenotype human tumor cells. PLoS ONE 2012, 7, e46571. [Google Scholar] [CrossRef]
- Lu, H.; Dalgard, C.L.; Mohyeldin, A.; McFate, T.; Tait, A.S.; Verma, A. Reversible inactivation of HIF-1 prolyl hydroxylases allows cell metabolism to control basal HIF-1. J. Biol. Chem. 2005, 280, 41928–41939. [Google Scholar] [CrossRef]
- Lu, H.; Forbes, R.A.; Verma, A. Hypoxia-inducible factor 1 activation by aerobic glycolysis implicates the Warburg effect in carcinogenesis. J. Biol. Chem. 2002, 277, 23111–23115. [Google Scholar] [CrossRef]
- Semenza, G.L. Defining the role of hypoxia-inducible factor 1 in cancer biology and therapeutics. Oncogene 2010, 29, 625–634. [Google Scholar] [CrossRef]
- Wang, Z.; Smith, K.S.; Murphy, M.; Piloto, O.; Somervaille, T.C.P.; Cleary, M.L. Glycogen synthase kinase 3 in MLL leukaemia maintenance and targeted therapy. Nature 2008, 455, 1205–1209. [Google Scholar] [PubMed]
- Woodgett, J.R. Regulation and functions of the glycogen synthase kinase-3 subfamily. Semin. Cancer Biol. 1994, 5, 269–275. [Google Scholar] [PubMed]
- Ougolkov, A.V.; Fernandez-Zapico, M.E.; Savoy, D.N.; Urrutia, R.A.; Billadeau, D.D. Glycogen synthase kinase-3β participates in nuclear factor κB–mediated gene transcription and cell survival in pancreatic cancer cells. Cancer Res. 2005, 65, 2076–2081. [Google Scholar]
- Kotliarova, S.; Pastorino, S.; Kovell, L.C.; Kotliarov, Y.; Song, H.; Zhang, W.; Bailey, R.; Maric, D.; Zenklusen, J.C.; Lee, J.; et al. Glycogen synthase kinase-3 inhibition induces glioma cell death through c-MYC, nuclear factor-κB, and glucose regulation. Cancer Res. 2008, 68, 6643–6651. [Google Scholar] [CrossRef]
- Marchand, B.; Tremblay, I.; Cagnol, S.; Boucher, M.-J. Inhibition of glycogen synthase kinase-3 activity triggers an apoptotic response in pancreatic cancer cells through JNK-dependent mechanisms. Carcinogenesis 2012, 33, 529–537. [Google Scholar] [PubMed]
- Shakoori, A.; Ougolkov, A.; Yu, Z.W.; Zhang, B.; Modarressi, M.H.; Billadeau, D.D.; Mai, M.; Takahashi, Y.; Minamoto, T. Deregulated GSK3β activity in colorectal cancer: Its association with tumor cell survival and proliferation. Biochem. Biophys. Res. Commun. 2005, 334, 1365–1373. [Google Scholar] [CrossRef]
- Mazor, M.; Kawano, Y.; Zhu, H.; Waxman, J.; Kypta, R.M. Inhibition of glycogen synthase kinase-3 represses androgen receptor activity and prostate cancer cell growth. Oncogene 2004, 23, 7882–7892. [Google Scholar] [CrossRef]
- Ryves, W.; Harwood, A.J. Lithium inhibits glycogen synthase kinase-3 by competition for magnesium. Biochem. Biophys. Res. Commun. 2001, 280, 720–725. [Google Scholar] [CrossRef]
- Klein, P.S.; Melton, D.A. A molecular mechanism for the effect of lithium on development. Proc. Natl. Acad. Sci. USA 1996, 93, 8455–8459. [Google Scholar] [CrossRef] [PubMed]
- Li, X.; Friedman, A.B.; Zhu, W.; Wang, L.; Boswell, S.; May, R.S.; Davis, L.L.; Jope, R.S. Lithium regulates glycogen synthase kinase-3β in human peripheral blood mononuclear cells: Implication in the treatment of bipolar disorder. Biol. Psychiatry 2007, 61, 216–222. [Google Scholar] [CrossRef]
- Qu, L.; Huang, S.; Baltzis, D.; Rivas-Estilla, A.-M.; Pluquet, O.; Hatzoglou, M.; Koumenis, C.; Taya, Y.; Yoshimura, A.; Koromilas, A.E. Endoplasmic reticulum stress induces p53 cytoplasmic localization and prevents p53-dependent apoptosis by a pathway involving glycogen synthase kinase-3β. Genes Dev. 2004, 18, 261–277. [Google Scholar] [CrossRef] [PubMed]
- Watcharasit, P.; Bijur, G.N.; Song, L.; Zhu, J.; Chen, X.; Jope, R.S. Glycogen synthase kinase-3β (GSK3β) binds to and promotes the actions of p53. J. Biol. Chem. 2003, 278, 48872–48879. [Google Scholar] [CrossRef] [PubMed]
- Watcharasit, P.; Bijur, G.N.; Zmijewski, J.W.; Song, L.; Zmijewska, A.; Chen, X.; Johnson, G.V.W.; Jope, R.S. Direct, activating interaction between glycogen synthase kinase-3β and p53 after DNA damage. Proc. Natl. Acad. Sci. USA 2002, 99, 7951–7955. [Google Scholar] [CrossRef]
- Beurel, E.; Kornprobst, M.; Eggelpoël, M.-J.B.-V.; Ruiz-Ruiz, C.; Cadoret, A.; Capeau, J.; Desbois-Mouthon, C. GSK-3β inhibition by lithium confers resistance to chemotherapy-induced apoptosis through the repression of CD95 (Fas/APO-1) expression. Exp. Cell Res. 2004, 300, 354–364. [Google Scholar] [CrossRef]
- Cao, Q.; Lu, X.; Feng, Y.-J. Glycogen synthase kinase-3β positively regulates the proliferation of human ovarian cancer cells. Cell Res. 2006, 16, 671–677. [Google Scholar] [CrossRef] [PubMed]
- Kunnimalaiyaan, M.; Vaccaro, A.M.; Ndiaye, M.A.; Chen, H. Inactivation of glycogen synthase kinase-3β, a downstream target of the raf-1 pathway, is associated with growth suppression in medullary thyroid cancer cells. Mol. Cancer Ther. 2007, 6, 1151–1158. [Google Scholar] [CrossRef]
- Wang, J.S.; Wang, C.L.; Wen, J.F.; Wang, Y.J.; Hu, Y.B.; Ren, H.Z. Lithium inhibits proliferation of human esophageal cancer cell line Eca-109 by inducing a G2/M cell cycle arrest. J. Gastroenterol. 2008, 14, 3982. [Google Scholar] [CrossRef]
- Korur, S.; Huber, R.M.; Sivasankaran, B.; Petrich, M.; Morin, P., Jr.; Hemmings, B.A.; Merlo, A.; Lino, M.M. GSK3β regulates differentiation and growth arrest in glioblastoma. PLoS ONE 2009, 4, e7443. [Google Scholar] [CrossRef]
- Maurer, G.D.; Brucker, D.P.; Bähr, O.; Harter, P.N.; Hattingen, E.; Walenta, S.; Mueller-Klieser, W.; Steinbach, J.P.; Rieger, J. Differential utilization of ketone bodies by neurons and glioma cell lines: A rationale for ketogenic diet as experimental glioma therapy. BMC Cancer 2011, 11, 315. [Google Scholar] [CrossRef]
- Bijur, G.N.; De Sarno, P.; Jope, R.S. Glycogen synthase kinase-3β facilitates staurosporine-and heat shock-induced apoptosis: Protection by lithium. J. Biol. Chem. 2000, 275, 7583–7590. [Google Scholar] [CrossRef]
- Chen, R.-W.; Chuang, D.-M. Long term lithium treatment suppresses p53 and Bax expression but increases Bcl-2 expression: A prominent role in neuroprotection against excitotoxicity. J. Biol. Chem. 1999, 274, 6039–6042. [Google Scholar] [CrossRef]
- Cross, D.A.; Alessi, D.R.; Cohen, P.; Andjelkovich, M.; Hemmings, B.A. Inhibition of glycogen synthase kinase-3 by insulin mediated by protein kinase B. Nature 1995, 378, 785–789. [Google Scholar]
- King, T.D.; Bijur, G.N.; Jope, R.S. Caspase-3 activation induced by inhibition of mitochondrial complex I is facilitated by glycogen synthase kinase-3β and attenuated by lithium. Brain Res. 2001, 919, 106–114. [Google Scholar] [CrossRef] [PubMed]
- Gustin, J.P.; Karakas, B.; Weiss, M.B.; Abukhdeir, A.M.; Lauring, J.; Garay, J.P.; Cosgrove, D.; Tamaki, A.; Konishi, H.; Konishi, Y.; et al. Knockin of mutant PIK3CA activates multiple oncogenic pathways. Proc. Natl. Acad. Sci. USA 2009, 106, 2835–2840. [Google Scholar] [CrossRef] [PubMed]
- Karlovic, D.; Jakopec, S.; Dubravcic, K.; Batinic, D.; Buljan, D.; Osmak, M. Lithium increases expression of p21WAF/Cip1 and survivin in human glioblastoma cells. Cell Biol. Toxicol. 2007, 23, 83–90. [Google Scholar] [CrossRef]
- Mora, A.; Sabio, G.; Alonso, J.C.; Soler, G.; Centeno, F. Different dependence of lithium and valproate on PI3K/PKB pathway. Bipolar Disord. 2002, 4, 195–200. [Google Scholar] [CrossRef] [PubMed]
- Adler, J.T.; Hottinger, D.G.; Kunnimalaiyaan, M.; Chen, H. Inhibition of growth in medullary thyroid cancer cells with histone deacetylase inhibitors and lithium chloride. J. Surg. Res. 2010, 159, 640–644. [Google Scholar] [CrossRef]
- Liao, X.; Zhang, L.; Thrasher, J.B.; Du, J.; Li, B. Glycogen synthase kinase-3β suppression eliminates tumor necrosis factor-related apoptosis-inducing ligand resistance in prostate cancer. Mol. Cancer Ther. 2003, 2, 1215–1222. [Google Scholar] [PubMed]
- Ozaki, N.; Chuang, D.M. Lithium increases transcription factor binding to AP-1 and cyclic AMP-responsive element in cultured neurons and rat brain. J. Neurochem. 1997, 69, 2336–2344. [Google Scholar] [CrossRef]
- Chen, R.W.; Qin, Z.H.; Ren, M.; Kanai, H.; Chalecka-Franaszek, E.; Leeds, P.; Chuang, D.M. Regulation of c-Jun N-terminal kinase, p38 kinase and AP-1 DNA binding in cultured brain neurons: Roles in glutamate excitotoxicity and lithium neuroprotection. J. Neurochem. 2003, 84, 566–575. [Google Scholar] [CrossRef]
- Wang, M.J.; Huang, H.Y.; Chen, W.F.; Chang, H.F.; Kuo, J.S. Glycogen synthase kinase-3β inactivation inhibits tumor necrosis factor-α production in microglia by modulating nuclear factor κB and MLK3/JNK signaling cascades. J. Neuroinflamm. 2010, 7, 99. [Google Scholar] [CrossRef] [PubMed]
- Peng, Z.; Ji, Z.; Mei, F.; Lu, M.; Ou, Y.; Cheng, X. Lithium inhibits tumorigenic potential of PDA cells through targeting hedgehog-GLI signaling pathway. PLoS ONE 2013, 8, e61457. [Google Scholar] [CrossRef]
- Li, H.; Huang, K.; Liu, X.; Liu, J.; Lu, X.; Tao, K.; Wang, G.; Wang, J. Lithium chloride suppresses colorectal cancer cell survival and proliferation through ROS/GSK-3β/NF-κB signaling pathway. Oxidative Med. Cell. Longev. 2014, 2014, 241864. [Google Scholar] [CrossRef]
- Sarkar, S.; Rubinsztein, D.C. Inositol and IP3 levels regulate autophagy—Biology and therapeutic speculations. Autophagy 2006, 2, 132–134. [Google Scholar] [CrossRef]
- Sarkar, S.; Floto, R.A.; Berger, Z.; Imarisio, S.; Cordenier, A.; Pasco, M.; Cook, L.J.; Rubinsztein, D.C. Lithium induces autophagy by inhibiting inositol monophosphatase. J. Cell Biol. 2005, 170, 1101–1111. [Google Scholar] [CrossRef]
- Chiu, C.-T.; Chuang, D.-M. Molecular actions and therapeutic potential of lithium in preclinical and clinical studies of CNS disorders. Pharmacol. Ther. 2010, 128, 281–304. [Google Scholar] [CrossRef] [PubMed]
- Sarkar, S.; Perlstein, E.O.; Imarisio, S.; Pineau, S.; Cordenier, A.; Maglathlin, R.L.; Webster, J.A.; Lewis, T.A.; O’Kane, C.J.; Schreiber, S.L.; et al. Small molecules enhance autophagy and reduce toxicity in Huntington’s disease models. Nat. Chem. Biol. 2007, 3, 331–338. [Google Scholar] [CrossRef]
- Heiseke, A.; Aguib, Y.; Riemer, C.; Baier, M.; Schätzl, H.M. Lithium induces clearance of protease resistant prion protein in prion-infected cells by induction of autophagy. J. Neurochem. 2009, 109, 25–34. [Google Scholar] [CrossRef]
- Bgatova, N.; Taskaeva, I.; Makarova, V. Influence of distant tumor growth and lithium treatment on ultrastructural organization of kidney proximal tubules. Ultrastruct. Pathol. 2021, 45, 212–223. [Google Scholar] [CrossRef] [PubMed]
- Taskaeva, I.; Bgatova, N. Ultrastructural and immunofluorescent analysis of lithium effects on autophagy in hepatocellular carcinoma cells. Asian Pac. J. Cancer Biol. 2018, 3, 83–87. [Google Scholar] [CrossRef]
- Taskaeva, I.S.; Bgatova, N.P.; Solovieva, A.O. Autophagy in Hepatocellular Carcinoma-29 after Single or Combined Administration of Lithium Carbonate and Rapamycin. Cell Tissue Biol. 2019, 13, 353–359. [Google Scholar] [CrossRef]
- Kim, E.C.; Meng, H.; Jun, A.S. Lithium treatment increases endothelial cell survival and autophagy in a mouse model of Fuchs endothelial corneal dystrophy. Br. J. Ophthalmol. 2013, 97, 1068–1073. [Google Scholar] [CrossRef]
- Yang, J.; Takahashi, Y.; Cheng, E.; Liu, J.; Terranova, P.F.; Zhao, B.; Thrasher, J.B.; Wang, H.-G.; Li, B. GSK-3β promotes cell survival by modulating Bif-1-dependent autophagy and cell death. J. Cell Sci. 2010, 123, 861–870. [Google Scholar] [CrossRef] [PubMed]
- Li, L.; Song, H.; Zhong, L.; Yang, R.; Yang, X.-Q.; Jiang, K.-L.; Liu, B.-Z. Lithium chloride promotes apoptosis in human leukemia NB4 cells by inhibiting glycogen synthase kinase-3 beta. Int. J. Med Sci. 2015, 12, 805. [Google Scholar] [CrossRef]
- Fine, E.J.; Miller, A.; Quadros, E.V.; Sequeira, J.M.; Feinman, R.D. Acetoacetate reduces growth and ATP concentration in cancer cell lines which over-express uncoupling protein 2. Cancer Cell Int. 2009, 9, 14. [Google Scholar] [PubMed]
- Franklin, S.; Young, J.; Nonnecke, B. Effects of ketones, acetate, butyrate, and glucose on bovine lymphocyte proliferation. J. Dairy Sci. 1991, 74, 2507–2514. [Google Scholar] [CrossRef] [PubMed]
- Bartmann, C.; Raman, S.R.J.; Flöter, J.; Schulze, A.; Bahlke, K.; Willingstorfer, J.; Strunz, M.; Wöckel, A.; Klement, R.J.; Kapp, M.; et al. Beta-hydroxybutyrate (3-OHB) can influence the energetic phenotype of breast cancer cells, but does not impact their proliferation and the response to chemotherapy or radiation. Cancer Metab. 2018, 6, 8. [Google Scholar] [CrossRef]
- Liu, J.; Ju, P.; Zhou, Y.; Zhao, Y.; Xie, Y.; Long, Y.; Gu, Y.; Ni, D.; Lyv, Z.; Mao, Z.; et al. Six2 is a coordinator of LiCl-induced cell proliferation and apoptosis. Int. J. Mol. Sci. 2016, 17, 1504. [Google Scholar]
- Zanni, G.; Di Martino, E.; Omelyanenko, A.; Andäng, M.; Delle, U.; Elmroth, K.; Blomgren, K. Lithium increases proliferation of hippocampal neural stem/progenitor cells and rescues irradiation-induced cell cycle arrest in vitro. Oncotarget 2015, 6, 37083. [Google Scholar] [CrossRef] [PubMed]
- Smits, V.A.; Essers, M.A.; Loomans, D.S.; Klompmaker, R.; Rijksen, G.; Medema, R.H. Inhibition of cell proliferation by lithium is associated with interference in cdc2 activation. FEBS Lett. 1999, 457, 23–27. [Google Scholar] [CrossRef]
- Fu, Y.; Zheng, Y.; Chan, K.G.; Liang, A.; Hu, F. Lithium chloride decreases proliferation and migration of C6 glioma cells harboring isocitrate dehydrogenase 2 mutant via GSK-3β. Mol. Biol. Rep. 2014, 41, 3907–3913. [Google Scholar] [CrossRef]
- Grimes, C.A.; Jope, R.S. The multifaceted roles of glycogen synthase kinase 3β in cellular signaling. Prog. Neurobiol. 2001, 65, 391–426. [Google Scholar] [CrossRef]
- Ding, Q.; Xia, W.; Liu, J.-C.; Yang, J.-Y.; Lee, D.-F.; Xia, J.; Bartholomeusz, G.; Li, Y.; Pan, Y.; Li, Z.; et al. associates with and primes GSK-3β for its inactivation resulting in upregulation of β-catenin. Mol. Cell 2005, 19, 159–170. [Google Scholar] [CrossRef] [PubMed]
- Díaz, M.; Vidal, F.; Cruz, A.L.; De Araujo, W.M.; Tanaka, M.N.; Viola, J.P.; Morgado-Diaz, J.A. Lithium reduces tumorigenic potential in response to EGF signaling in human colorectal cancer cells. Int. J. Oncol. 2011, 38, 1365–1373. [Google Scholar]
- Farina, A.K.; Bong, Y.-S.; Feltes, C.M.; Byers, S.W. Post-transcriptional regulation of cadherin-11 expression by GSK-3 and β-catenin in prostate and breast cancer cells. PLoS ONE 2009, 4, e4797. [Google Scholar] [CrossRef]
- Ohira, T.; Gemmill, R.M.; Ferguson, K.; Kusy, S.; Roche, J.; Brambilla, E.; Zeng, C.; Baron, A.; Bemis, L.; Erickson, P.; et al. WNT7a induces E-cadherin in lung cancer cells. Proc. Natl. Acad. Sci. USA 2003, 100, 10429–10434. [Google Scholar] [CrossRef] [PubMed]
- Maeng, Y.-S.; Lee, R.; Lee, B.; Choi, S.-I.; Kim, E.K. Lithium inhibits tumor lymphangiogenesis and metastasis through the inhibition of TGFBIp expression in cancer cells. Sci. Rep. 2016, 6, 20739. [Google Scholar] [CrossRef]
- Nowicki, M.O.; Dmitrieva, N.; Stein, A.M.; Cutter, J.L.; Godlewski, J.; Saeki, Y.; Nita, M.; Berens, M.E.; Sander, L.M.; Newton, H.B.; et al. Lithium inhibits invasion of glioma cells; possible involvement of glycogen synthase kinase-3. Neuro-Oncology 2008, 10, 690–699. [Google Scholar] [CrossRef]
- Mao, C.D.; Hoang, P.; DiCorleto, P.E. Lithium inhibits cell cycle progression and induces stabilization of p53 in bovine aortic endothelial cells. J. Biol. Chem. 2001, 276, 26180–26188. [Google Scholar] [CrossRef]
- Morin, P.J.; Sparks, A.B.; Korinek, V.; Barker, N.; Clevers, H.; Vogelstein, B.; Kinzler, K.W. Activation of β-catenin-Tcf signaling in colon cancer by mutations in β-catenin or APC. Science 1997, 275, 1787–1790. [Google Scholar] [CrossRef]
- Gradl, D.; Kühl, M.; Wedlich, D. The Wnt/Wg signal transducer β-catenin controls fibronectin expression. Mol. Cell. Biol. 1999, 19, 5576–5587. [Google Scholar] [CrossRef]
- Tetsu, O.; McCormick, F.J.N. β-Catenin regulates expression of cyclin D1 in colon carcinoma cells. Mol. Cell. Biol. 1999, 398, 422–426. [Google Scholar]
- He, T.-C.; Sparks, A.B.; Rago, C.; Hermeking, H.; Zawel, L.; da Costa, L.T.; Morin, P.J.; Vogelstein, B.; Kinzler, K.W. Identification of c-MYC as a target of the APC pathway. Science 1998, 281, 1509–1512. [Google Scholar] [CrossRef] [PubMed]
- Sun, A.; Shanmugam, I.; Song, J.; Terranova, P.F.; Thrasher, J.B.; Li, B. Lithium suppresses cell proliferation by interrupting E2F–DNA interaction and subsequently reducing S–phase gene expression in prostate cancer. Prostate 2007, 67, 976–988. [Google Scholar] [CrossRef]
- Matsebatlela, T.; Gallicchio, V.; Becker, R. Lithium modulates cancer cell growth, apoptosis, gene expression and cytokine production in HL-60 promyelocytic leukaemia cells and their drug-resistant sub-clones. Biol. Trace Element Res. 2012, 149, 323–330. [Google Scholar] [CrossRef]
- Mantovani, A.; Allavena, P.; Sica, A.; Balkwill, F. Cancer-related inflammation. Nature 2008, 454, 436–444. [Google Scholar] [CrossRef]
- Singh, N.; Baby, D.; Rajguru, J.P.; Patil, P.B.; Thakkannavar, S.S.; Pujari, V.B. Inflammation and cancer. Ann. Afr. Med. 2019, 18, 121. [Google Scholar] [CrossRef] [PubMed]
- Lee, J.-H.; Kim, S.-W.; Kim, J.-H.; Kim, H.-J.; Um, J.; Jung, D.-W.; Williams, D. Lithium chloride protects against sepsis-induced skeletal muscle atrophy and cancer cachexia. Cells 2021, 10, 1017. [Google Scholar] [CrossRef]
- Huang, W.-C.; Lin, Y.-S.; Wang, C.-Y.; Tsai, C.-C.; Tseng, H.-C.; Chen, C.-L.; Lu, P.-J.; Chen, P.-S.; Qian, L.; Hong, J.-S.; et al. Glycogen synthase kinase-3 negatively regulates anti-inflammatory interleukin-10 for lipopolysaccharide-induced iNOS/NO biosynthesis and RANTES production in microglial cells. Immunology 2009, 128, e275–e286. [Google Scholar] [CrossRef]
- Nahman, S.; Belmaker, R.; Azab, A.N. Effects of lithium on lipopolysaccharide-induced inflammation in rat primary glia cells. Endotoxin Res. 2012, 18, 447–458. [Google Scholar] [CrossRef]
- Nassar, A.; Azab, A.N. Effects of lithium on inflammation. ACS Chem. Neurosci. 2014, 5, 451–458. [Google Scholar] [CrossRef]
- Scholtz, W. IL-6 in disease: Cause or cure. Immunopathology 1996, 3, 131–150. [Google Scholar]
- Bgatova, N.; Taskaeva, I. Ultrastructure of the kidney filtration barrier in conditions of distant tumor growth and lithium treatment. Ultrastruct. Pathol. 2020, 44, 412–421. [Google Scholar] [CrossRef] [PubMed]
- Novetsky, A.P.; Thompson, D.M.; Zighelboim, I.; Thaker, P.H.; Powell, M.A.; Mutch, D.G.; Goodfellow, P.J. Lithium chloride and inhibition of glycogen synthase kinase 3β as a potential therapy for serous ovarian cancer. Int. J. Gynecol. Cancer 2013, 23, 361–366. [Google Scholar] [CrossRef] [PubMed]
- Lubner, S.J.; Kunnimalaiyaan, M.; Holen, K.D.; Ning, L.; Ndiaye, M.; LoConte, N.K.; Mulkerin, D.L.; Schelman, W.R.; Chen, H. A preclinical and clinical study of lithium in low-grade neuroendocrine tumors. Oncology 2011, 16, 452–457. [Google Scholar] [CrossRef]
- Khasraw, M.; Ashley, D.; Wheeler, G.; Berk, M. Using lithium as a neuroprotective agent in patients with cancer. BMC Med. 2012, 10, 131. [Google Scholar] [CrossRef]
- Rouhani, M.; Goliaei, B.; Khodagholi, F.; Nikoofar, A. Lithium increases radiosensitivity by abrogating DNA repair in breast cancer spheroid culture. Arch. Iran. Med. 2014, 17, 352–360. [Google Scholar]
- Bilir, A.; Aynacioğlu, A.; Tuna, M.Y. The possible interactions and therapeutic roles of lithium chloride and midkine on cancer treatment. Crit. Rev. Oncog. 2019, 24, 35–45. [Google Scholar] [CrossRef]
- Huo, K.; Sun, Y.; Li, H.; Du, X.; Wang, X.; Karlsson, N.; Zhu, C.; Blomgren, K. Lithium reduced neural progenitor apoptosis in the hippocampus and ameliorated functional deficits after irradiation to the immature mouse brain. Mol. Cell. Neurosci. 2012, 51, 32–42. [Google Scholar] [CrossRef]
- Malaterre, J.; McPherson, C.S.; Denoyer, D.; Lai, E.; Hagekyriakou, J.; Lightowler, S.; Shudo, K.; Ernst, M.; Ashley, D.M.; Short, J.L.; et al. Enhanced lithium-induced brain recovery following cranial irradiation is not impeded by inflammation. Stem Cells Transl. Med. 2012, 1, 469–479. [Google Scholar] [CrossRef]
- Zhukova, N.; Ramaswamy, V.; Remke, M.; Martin, D.C.; Castelo-Branco, P.; Zhang, C.H.; Fraser, M.; Tse, K.; Poon, R.; Shih, D.J.; et al. WNT activation by lithium abrogates TP53 mutation associated radiation resistance in medulloblastoma. Acta Neuropathol. Commun. 2014, 2, 174. [Google Scholar] [CrossRef]
- Yang, C.; Wang, W.; Zhu, K.; Liu, W.; Luo, Y.; Yuan, X.; Wang, J.; Cheng, T.; Zhang, X. Lithium chloride with immunomodulatory function for regulating titanium nanoparticle-stimulated inflammatory response and accelerating osteogenesis through suppression of MAPK signaling pathway. Int. J. Nanomed. 2019, 14, 7475. [Google Scholar] [CrossRef]
- Yavari, S.; Geramifar, P.; Fallahpoor, M.; Changizi, V.; Gholami, M.; Meysamie, A.; Farzanehfar, S.; Abbasi, M. The effect of lithium on radioiodine thyroid tissue ablation. Intern. J. Radiat. Res. 2021, 19, 1045–1048. [Google Scholar] [CrossRef]
- Boehmerle, W.; Zhang, K.; Sivula, M.; Heidrich, F.M.; Lee, Y.; Jordt, S.-E.; Ehrlich, B.E. Chronic exposure to paclitaxel diminishes phosphoinositide signaling by calpain-mediated neuronal calcium sensor-1 degradation. Proc. Natl. Acad. Sci. USA 2007, 104, 11103–11108. [Google Scholar] [CrossRef] [PubMed]
- Ibrahim, E.Y.; Ehrlich, B.E. Prevention of chemotherapy-induced peripheral neuropathy: A review of recent findings. Crit. Rev. Oncol./Hematol. 2020, 145, 102831. [Google Scholar] [CrossRef]
- Sánchez, J.C.; Muñoz, L.V.; Ehrlich, B.E. Modulating TRPV4 channels with paclitaxel and lithium. Cell Calcium 2020, 91, 102266. [Google Scholar] [CrossRef] [PubMed]
- Ge, W.; Jakobsson, E. Systems biology understanding of the effects of lithium on cancer. Front. Oncol. 2019, 9, 296. [Google Scholar] [CrossRef]
- Huang, R.-Y.; Hsieh, K.-P.; Huang, W.-W.; Yang, Y.-H. of lithium and cancer risk in patients with bipolar disorder: Population-based cohort study. Br. J. Psychiatry 2016, 209, 393–399. [Google Scholar] [CrossRef] [PubMed]
- Najafi, S.; Heidarali, Z.; Rajabi, M.; Omidi, Z.; Zayeri, F.; Salehi, M.; Haghighat, S. Lithium and preventing chemotherapy-induced peripheral neuropathy in breast cancer patients: A placebo-controlled randomized clinical trial. Trials 2021, 22, 835. [Google Scholar] [CrossRef]
- Ueda, M.; Stefan, T.; Stetson, L.; Ignatz-Hoover, J.J.; Tomlinson, B.; Creger, R.J.; Cooper, B.; Lazarus, H.M.; de Lima, M.; Wald, D.N.; et al. Phase I Trial of Lithium and Tretinoin for Treatment of Relapsed and Refractory Non-promyelocytic Acute Myeloid Leukemia. Front Oncol. 2020, 10, 327. [Google Scholar] [CrossRef]
- Sherman, S.I. Thyroid carcinoma. Lancet 2003, 361, 501–511. [Google Scholar] [CrossRef] [PubMed]
- Wen, P.Y.; Macdonald, D.R.; Reardon, D.A.; Cloughesy, T.F.; Sorensen, A.G.; Galanis, E.; DeGroot, J.; Wick, W.; Gilbert, M.R.; Lassman, A.B.; et al. Updated response assessment criteria for high-grade gliomas: Response assessment in neuro-oncology working group. J. Clin. Oncol. 2010, 28, 1963–1972. [Google Scholar] [CrossRef]
- Steinbach, G.; Hockenbery, D.M.; Huls, G.; Furlong, T.; Myerson, D.; Loeb, K.R.; Fann, J.R.; Castilla-Llorente, C.; McDonald, G.B.; Martin, P.J. Pilot study of lithium to restore intestinal barrier function in severe graft-versus-host disease. PLoS ONE 2017, 12, e0183284. [Google Scholar] [CrossRef] [PubMed]
- Linssen, J.D.G.; van Neerven, S.M.; Aelvoet, A.S.; Elbers, C.C.; Vermeulen, L.; Dekker, E. The CHAMP-study: The CHemopreventive effect of lithium in familial AdenoMatous Polyposis; study protocol of a phase II trial. BMC Gastroenterol 2022, 22, 383. [Google Scholar] [CrossRef] [PubMed]
- Hattinger, C.M.; Patrizio, M.P.; Magagnoli, F.; Luppi, S.; Serra, M. An update on emerging drugs in osteosarcoma: Towards tailored therapies? Expert Opin. Emerg. Drugs 2019, 24, 153–171. [Google Scholar] [CrossRef]
- Bgatova, N.P.; Borodin, Y.I.; Makarova, V.; Pozhidaeva, A.A.; Rachkovskaya, L.N.; Konenkov, V.I. Effects of Nanosized Lithium Carbonate Particles on Intact Muscle Tissue and Tumor Growth. Bull. Exp. Biol. Med. 2014, 157, 89–94. [Google Scholar] [CrossRef]
- Bgatova, N.P.; Makarova, O.P.; Pozhidayeva, A.A.; Borodin, Y.I.; Rachkovskaya, L.N.; Konenkov, V.I. Effects of Lithium Nano-Scaled Particles on Local and Systemic Structural and Functional Organism Transformations Under Tumour Growth. Achiev. Life Sci. 2014, 8, 101–111. [Google Scholar] [CrossRef]
- Lykov, A.P.; Poveshchenko, O.V.; Bondarenko, N.A.; Bogatova, N.P.; Makarova, O.P.; Konenkov, V.I. Antiproliferative potential of officinal forms and nanoparticles of lithium salts. Bull. Exp. Biol. Med. 2016, 160, 827. [Google Scholar] [CrossRef]
- Gavrilova, Y.S.; Bgatova, N.P.; Solov’Eva, A.O.; Trifonova, K.E.; Lykov, A.; Borodin, Y.I.; Konenkov, V.I. Target cells for lithium in different forms within a heterogeneous hepatocarcinoma-29 population. Cell Tissue Biol. 2016, 10, 284–289. [Google Scholar] [CrossRef]
- Hosseini, Y.; Alavi, S.E.; Akbarzadeh, A.; Heidarinasab, A. Improving lithium carbonate therapeutics by pegylated liposomal technology: An in vivo study. Comp. Clin. Pathol. 2016, 25, 211–218. [Google Scholar] [CrossRef]
- Iqbal, H.; Razzaq, A.; Uzair, B.; Ain, N.U.; Sajjad, S.; Althobaiti, N.A.; Albalawi, A.E.; Menaa, B.; Haroon, M.; Khan, M.; et al. Breast cancer inhibition by biosynthesized titanium dioxide nanoparticles is comparable to free doxorubicin but appeared safer in BALB/c mice. Materials 2021, 14, 3155. [Google Scholar] [CrossRef] [PubMed]
- Özer, S.; Şenel, B.; Yazan, Y. Preparation and in vitro evaluation of in situ gelling system containing lithium carbonate for parenteral administration. Polym. Bull. 2020, 77, 599–622. [Google Scholar] [CrossRef]
- Patel, V.; Chudasama, A.; Nivsarkar, M.; Vasu, K.; Shishoo, C. Push-pull osmotic pump for zero order delivery of lithium carbonate: Development and in vitro characterization. Pharm. Dev. Technol. 2012, 17, 375–382. [Google Scholar] [CrossRef] [PubMed]
- Sasaki, M.; Hashimoto, S.; Sawa, T.; Amaya, F. Tumor necrosis factor-alpha induces expression of C/EBP-beta in primary afferent neurons following nerve injury. Neuroscience 2014, 279, 1–9. [Google Scholar] [CrossRef] [PubMed]
- Gambelli, F.; Di, Y.P.; Niu, X.; Friedman, M.; Hammond, T.; Riches, D.W.; Ortiz, L. Phosphorylation of tumor necrosis factor receptor 1 (p55) protects macrophages from silica-induced apoptosis. J. Biol. Chem. 2004, 279, 2020–2029. [Google Scholar] [CrossRef]
- Saleh, K.A.; Aldulmani, S.A.A.; Awwad, N.S.; Ibrahium, H.A.; Asiri, T.H.; Hamdy, M.S. Utilization of lithium incorporated mesoporous silica for preventing necrosis and increase apoptosis in different cancer cells. BMC Chem. 2019, 13, 8. [Google Scholar] [CrossRef] [PubMed]
- Belahmar, A.; Mikou, M.; Hoehr, C.; El Ghalmi, M. Cumulative dose experiments on Lithium formate monohydrate as an EPR-dosimeter for use in different radiation therapy scenarios. Nucl. Instrum. Methods Phys. Res. Sect. B Beam Interact. Mater. At. 2022, 532, 1–6. [Google Scholar] [CrossRef]
- Zhu, Z.; Xing, H.; Tang, R.; Qian, S.; He, S.; Hu, Q.; Zhang, N. The preconditioning of lithium promotes mesenchymal stem cell-based therapy for the degenerated intervertebral disc via upregulating cellular ROS. Stem Cell Res. Ther. 2021, 12, 239. [Google Scholar] [CrossRef]
- Farmani, A.R.; Salmeh, M.A.; Golkar, Z.; Moeinzadeh, A.; Ghiasi, F.F.; Amirabad, S.Z.; Shoormeij, M.H.; Mahdavinezhad, F.; Momeni, S.; Moradbeygi, F.; et al. Li-Doped Bioactive Ceramics: Promising Biomaterials for Tissue Engineering and Regenerative Medicine. J. Funct. Biomater. 2022, 13, 162. [Google Scholar] [CrossRef]
- Yazdanpanah, A.; Moztarzadeh, F.; Arabyazdi, S. A heat-generating lithium-ferrite doped bioactive glass for cancer hyperthermia. Phys. B Condens. Matter 2020, 593, 412298. [Google Scholar] [CrossRef]
- Zhou, Z.; Wang, X.; Zhang, H.; Huang, H.; Sun, L.; Ma, L.; Du, Y.; Pei, C.; Zhang, Q.; Li, H.; et al. Activating layered metal oxide nanomaterials via structural engineering as biodegradable nanoagents for photothermal cancer therapy. Small 2021, 17, 2007486. [Google Scholar] [CrossRef]
- Zhou, Z.; Li, X.; Hu, T.; Xue, B.; Chen, H.; Ma, L.; Liang, R.; Tan, C. Molybdenum-Based Nanomaterials for Photothermal Cancer Therapy. Adv. NanoBiomed Res. 2022, 2, 2200065. [Google Scholar] [CrossRef]
- Yu, X.; Zhang, Y.-C.; Yang, X.; Huang, Z.; Zhang, T.; Yang, L.; Meng, W.; Liu, X.; Gong, P.; Forni, A.; et al. Bonsai-inspired AIE nanohybrid photosensitizer based on vermiculite nanosheets for ferroptosis-assisted oxygen self-sufficient photodynamic cancer therapy. Nano Today 2022, 44, 101477. [Google Scholar] [CrossRef]
- Suganthi, M.; Sangeetha, G.; Gayathri, G.; Sankar, B.R. Biphasic dose-dependent effect of lithium chloride on survival of human hormone-dependent breast cancer cells (MCF-7). Biol. Trace Elem. Res. 2012, 150, 477–486. [Google Scholar] [CrossRef]
- Walker, R.J.; Weggery, S.; Bedford, J.J.; Mcdonald, F.J.; Ellis, G.; Leader, J.P. Lithium-induced reduction in urinary concentrating ability and urinary aquaporin 2 (AQP2) excretion in healthy volunteers. Kidney Int. 2005, 67, 291–294. [Google Scholar] [CrossRef]
- Pinna, M.; Manchia, M.; Puddu, S.; Minnai, G.; Tondo, L.; Salis, P. Cutaneous adverse reaction during lithium treatment: A case report and updated systematic review with meta-analysis. Int. J. Bipolar Disord. 2017, 5, 20. [Google Scholar] [CrossRef] [PubMed]
- Nair, C.G.; Menon, R.; Jacob, P.; Babu, M. Lithium-induced parathyroid dysfunction: A new case. Indian J. Endocrinol. Metab. 2013, 17, 930–932. [Google Scholar] [CrossRef]
- Meehan, A.D.; Udumyan, R.; Kardell, M.; Landén, M.; Järhult, J.; Wallin, G. Lithium-associated hypercalcemia: Pathophysiology, prevalence, management. World J. Surg. 2018, 42, 415–424. [Google Scholar] [CrossRef] [PubMed]
- Poels, E.M.P.; Bijma, H.H.; Galbally, M.; Bergink, V. Lithium during pregnancy and after delivery: A review. Int. J. Bipolar Disord. 2018, 6, 26. [Google Scholar] [CrossRef]
- Matsebatlela, T.M.; Mogodiri, R.K.; Hart, D.A.; Gallicchio, V.S.; Becker, R.W. Resistance to lithium-induced apoptosis in a lithium tolerant clone of HL-60 promyelocytes. J. Trace Microprobe Tech. 2000, 18, 163–170. [Google Scholar]
- Lucas, K.C.; Hart, D.A.; Becker, R.W. Porcine proximal tubular cells (LLC-PK1) are able to tolerate high levels of lithium chloride in vitro: Assessment of the influence of 1–20 mM LiCl on cell death and alterations in cell biology and biochemistry. Cell Biol. Int. 2010, 34, 225–233. [Google Scholar] [CrossRef] [PubMed]



{kind=link}
{kind=link}
{kind=link}
| Type of Salt | Type of Cancer | Experimental Model | Dose of Lithium | Outcomes of the Study | Ref. |
|---|---|---|---|---|---|
| LiCl | Lymph Node Carcinoma of the Prostate cells | Human prostate carcinoma LNCaP cells | 2.5, 10, 25, and 45 mM | LiCl showed a cytotoxic effect in a dose- and time-dependent manner (p < 0.001). LiCl increased apoptosis of Lymph Node Carcinoma of the Prostate cells in the presence of etoposide, which is S- and G2-phase-specific drug. | [19] |
| Li2CO3 particles | Hepatocarcinoma 29 | Male CBA mice | Li2CO3 particles 10–20 nm in diameter (0.058 mg) | Nanosized lithium carbonate particles were inessential for the function of peritoneal macrophages. | [20] |
| LiCl | Gastrointestinal tract cancers (colorectal carcinoma) | Female balb/c mice | Lithium chloride (200 mg/kg) | The efficacy of chemotherapeutic agents was improved with lithium, by enhancing non-apoptotic cell death. | [21] |
| Li3C6H5O7 and Li2CO3 particles | Hepatocellular carcinoma | Cell line HCC-29 (hepatocarcinoma) | 5 mM lithium salts | Nanosized lithium citrate mainly induced apoptosis, whereas lithium carbonate induced apoptosis and autophagic cell death. | [22] |
| LiCl | Glioblastoma | Human TP53wt U87 and TP53mut U251glioblastoma cells with methylated MGMT promoters | 1.2 mM | Low doses of lithium in combination with temozolomide induced glioma cell death via NFAT1/FasL signaling. | [23] |
| LiCl | Multiple myeloma | Human multiple myeloma cell lines (RPMI-8226 and U266) | 40 mM | Lithium inhibited cell proliferation and induced cell cycle arrest. | [24] |
| LiCl | Breast cancer | MDA-MB-231 breast cancer cells | ND | Induced autophagy in breast cancer cells treated with LiCl and mitomycin C. | [25] |
| LiAcAc LiCl | Neuroblastoma, renal cell carcinoma | Human embryonic kidney cell line HEK293, human normal dermal fibroblasts (HDFn) and RCC 786-O cells | 2.5, 5, and 10 mM LiAcAc and with equimolar concentrations of lithium chloride (LiCl) | LiAcAc and LiCl affected the growth of all cell lines, either negatively or positively. | [18] |
| LiCl | Breast cancer | SUM159 MCF7 MDA-MB-231 and HEK293T | 20 mM | In triple-negative breast cancers, GSK3β was upregulated and correlated with worse survival in patients. Inhibition of GSK3β via LiCl decreased the expression of markers of the mesenchymal phenotype, indicating its ability to impede the process of epithelial-mesenchymal transition (EMT). | [26] |
| LiCl | Breast cancer | MCF7 MDA-MB-231 | 30 and 20 mM | Treatment with LiCl increased the active GSK3β protein, and DNA damage, and decreased survival, independently of estrogen receptor status in breast cancer cells exposed to ionizing radiation. | [27] |
| LiCl | Lung cancer | A549 | 50 ng/ml | The protein expression levels of PD-L1 were upregulated after treatment with LiCl. PD-L1 is the major ligand of PD-1 and is expressed in a variety of tumors, including in NSCLC. Overexpression of PD-L1 was implicated in tumor immunity, and inhibition of PD-L1 enhanced antitumor immunity by preventing tumor cells from escaping host immune responses. | [28] |
| LiCl | Nasopharyngeal carcinoma | C666-1,5-8F and SUNE-1 | 20 mM | LiCl was used to activate Wnt/β-catenin, which is correlated with the EMT process in cancer. | [29] |
| LiAcAc | Breast cancer | Breast cancer cell lines (MCF7, MDA-MB-231, Hs578T) | 3 mM, 10 mM, or 30 mM | Cytotoxic effects of LiAcAc treatment were significantly similar to those caused by LiCl. | [30] |
| LiCl | Colorectal cancer | T88 primary colon cancer cells from a patient with sporadic colon cancer. | 30 mM | LiCl could be used to sensitize colon cancer cells to radiation therapy. | [31] |
| LiCl | Human head and neck squamous cell carcinoma (HNSCC) | HNSCC SCC9, SCC25, A253, DETROIT562 & Fadu cell lines | 11.45 mM, 15.24 mM, 14.56 mM, 11.4 mM, 5.53 mM | LiCl reduced the proliferation and colony formation capabilities of HNSCC cell lines and influenced wound closure. LiCl increased the inhibitory Ser9 phosphorylation of GSK3β, leading to an increment of the GLI3 repressor form; inhibiting HH-GLI pathway activity. | [32] |
| Li2CO3 | Hepatocellular carcinoma | HCC-29 cell line | 5 mM | Li2CO3 arrested the cell cycle in the G2/M-phase and induced apoptosis and autophagy in HCC-29 cells. The upregulation of autophagy markers LC3B and LAMP1 was assessed using the presence of Li2CO3. | [33] |
| LiCl | Hepatocellular carcinoma | HepG2 cell line | 20 mM | Antitumor effects of KIF18B-siRNA, which targets KIF18B, an important in pairing and separation of chromosomes, could be reversed by LiCl treatment, through the Wnt/β-catenin pathway | [34] |
| LiCl | Myeloma | Human MM cell line RPMI8226 | 20 mmol/L | MiR-135b is involved in the development and progression of various cancers. In this study, LiCl was used to activate the Wnt/β-catenin pathway, reversing the effects of downregulating miR-135b on the proliferation, migration, invasion, and apoptosis of MM cells. | [35] |
| LiCl | Ameloblastoma, an odontogenic epithelial tumor | AM-1 cell line and human ameloblastoma cells | 1 and 4 mM | Lithium chloride (LiCl) increased the size and decreased the proliferation of cells, and expression of Sox2. | [36] |
| Li2CO3 | Skin melanoma | B16 melanoma cells | ND | Li2CO3 inhibited cell proliferation and stimulated cell death in melanoma cells through induction of autophagy and apoptosis. | [37] |
| NCT Number | Title | Status | Conditions | Interventions | Characteristics | Population | Results | Ref. |
|---|---|---|---|---|---|---|---|---|
| NCT03153280 | Dose Escalation Study of Lithium with Oxaliplatin and Capecitabine in Advanced Oesophago-Gastric or Colorectal Cancer (Lithium) | Recruiting |
|
|
|
| No data were analyzed, reported, or published on this study. | ND |
| NCT00582712 | An Initial Study of Lithium in Patients with Medullary Thyroid Cancer | Terminated |
|
|
|
| No data were analyzed, reported, or published on this study. | ND |
| NCT00501540 | Lithium for Low-Grade Neuroendocrine Tumors | Completed |
|
|
|
|
| [168] |
| NCT02198859 | Evaluation of lithium and its effect on clinically localized prostate cancer | Completed |
|
|
|
|
| ND |
| NCT00251316 | Effect of Lithium Carbonate on Low-Dose Radioiodine Therapy in Early Thyroid Cancer | Completed |
|
|
|
|
| [184] |
| NCT01486459 | A Feasibility Trial Using Lithium as A Neuroprotective Agent in Patients Undergoing Prophylactic Cranial Irradiation for Small Cell Lung Cancer (TULIP) | Terminated |
|
|
|
|
| ND |
| NCT05221593 | Efficacy of Lithium Against Chemotherapy Induced Neutropenia in Breast Cancer Patients | Completed |
|
|
|
|
| ND |
| NCT00469937 | Ph I Study of Lithium During Whole Brain Radiotherapy for Patients with Brain Metastases | Terminated |
|
|
|
|
| ND |
| NCT01553916 | Neuroprotective Effects of Lithium in Patients with Small Cell Lung Cancer Undergoing Radiation Therapy to the Brain | Completed |
|
|
Phase 1 Phase 2 |
|
| ND |
| NCT01105702 | Temodar (Temozolomide), Bevacizumab, Lithium and Radiation for High Grade Glioma | Terminated |
|
|
|
|
| [185] |
| NCT00408681 | Lithium Carbonate in Treating Patients with Acute Intestinal Graft-Versus-Host-Disease (GVHD) After Donor Stem Cell Transplant | Completed |
|
|
|
|
| [186] |
| NCT05402891 | The CHAMP-study: The Chemo preventive Effect of Lithium in Familial Adenomatous Polyposis | Enrolling by invitation |
|
|
|
|
| [187] |
| NCT01669369 | Clinical Trial of Lithium Carbonate Combined with Neoadjuvant Chemotherapy to Treat Osteosarcoma | Recruiting |
|
|
|
|
| [188] |
| NCT01820624 | Lithium Carbonate and Tretinoin in Treating Patients with Relapsed or Refractory Acute Myeloid Leukemia | Completed |
|
|
|
|
| [183] |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Villegas-Vázquez, E.Y.; Quintas-Granados, L.I.; Cortés, H.; González-Del Carmen, M.; Leyva-Gómez, G.; Rodríguez-Morales, M.; Bustamante-Montes, L.P.; Silva-Adaya, D.; Pérez-Plasencia, C.; Jacobo-Herrera, N.; et al. Lithium: A Promising Anticancer Agent. Life 2023, 13, 537. https://doi.org/10.3390/life13020537
Villegas-Vázquez EY, Quintas-Granados LI, Cortés H, González-Del Carmen M, Leyva-Gómez G, Rodríguez-Morales M, Bustamante-Montes LP, Silva-Adaya D, Pérez-Plasencia C, Jacobo-Herrera N, et al. Lithium: A Promising Anticancer Agent. Life. 2023; 13(2):537. https://doi.org/10.3390/life13020537
Chicago/Turabian StyleVillegas-Vázquez, Edgar Yebrán, Laura Itzel Quintas-Granados, Hernán Cortés, Manuel González-Del Carmen, Gerardo Leyva-Gómez, Miguel Rodríguez-Morales, Lilia Patricia Bustamante-Montes, Daniela Silva-Adaya, Carlos Pérez-Plasencia, Nadia Jacobo-Herrera, and et al. 2023. "Lithium: A Promising Anticancer Agent" Life 13, no. 2: 537. https://doi.org/10.3390/life13020537
APA StyleVillegas-Vázquez, E. Y., Quintas-Granados, L. I., Cortés, H., González-Del Carmen, M., Leyva-Gómez, G., Rodríguez-Morales, M., Bustamante-Montes, L. P., Silva-Adaya, D., Pérez-Plasencia, C., Jacobo-Herrera, N., Reyes-Hernández, O. D., & Figueroa-González, G. (2023). Lithium: A Promising Anticancer Agent. Life, 13(2), 537. https://doi.org/10.3390/life13020537