

Aging and Cerebral Glucose Metabolism: 18F-FDG-PET/CT Reveals Distinct Global and Regional Metabolic Changes in Healthy Patients

 , ,
, ,  and
and 
Abstract
:1. Introduction
2. Methods
2.1. Study Population
2.2. Subjects
2.3. Brain PET/CT Acquisition Protocol
2.4. Brain PET/CT Data Analysis
3. Results
3.1. Clinical Characteristics of the Study Population
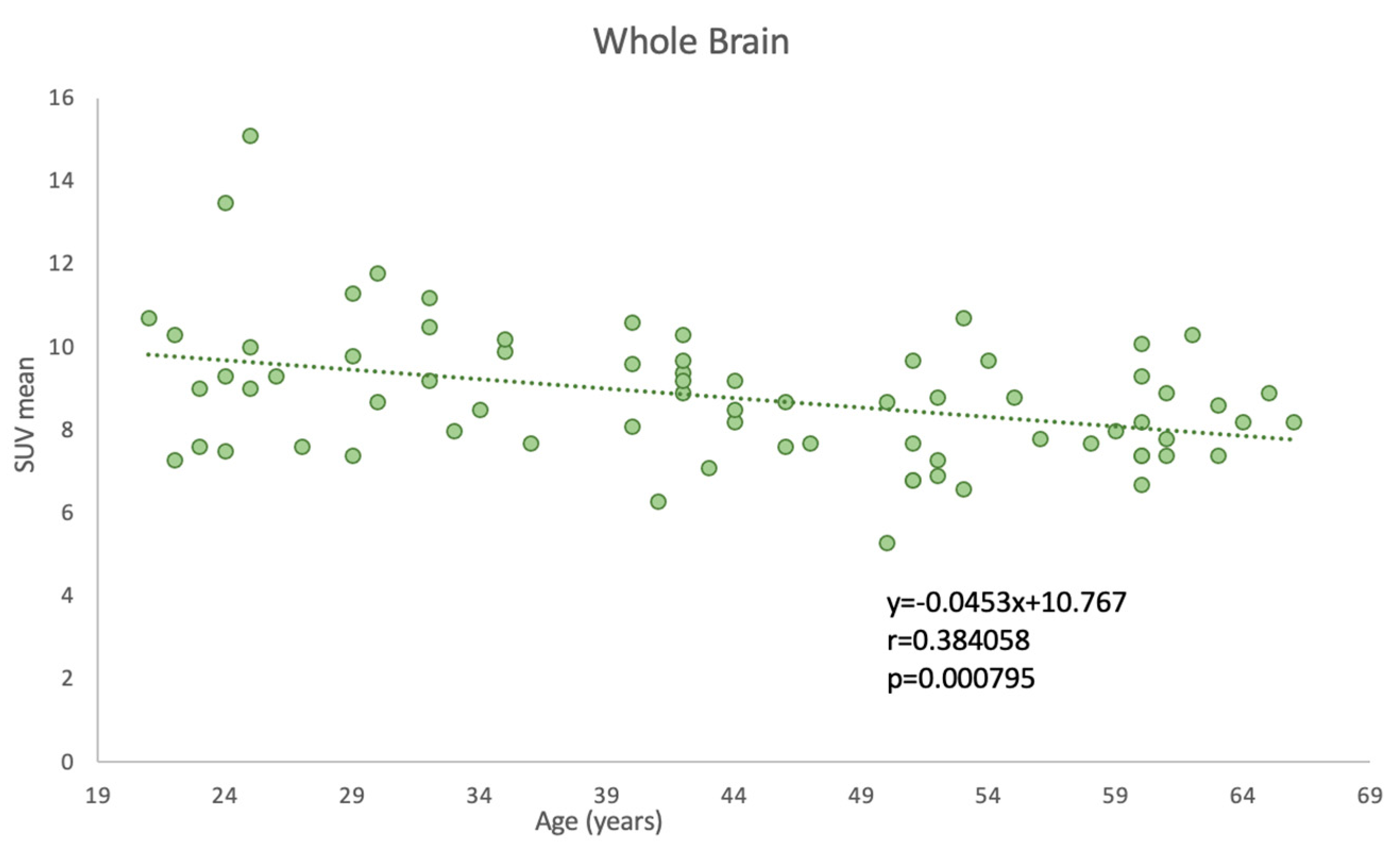
3.2. Effect of Aging on Global Cerebral Metabolic Activity
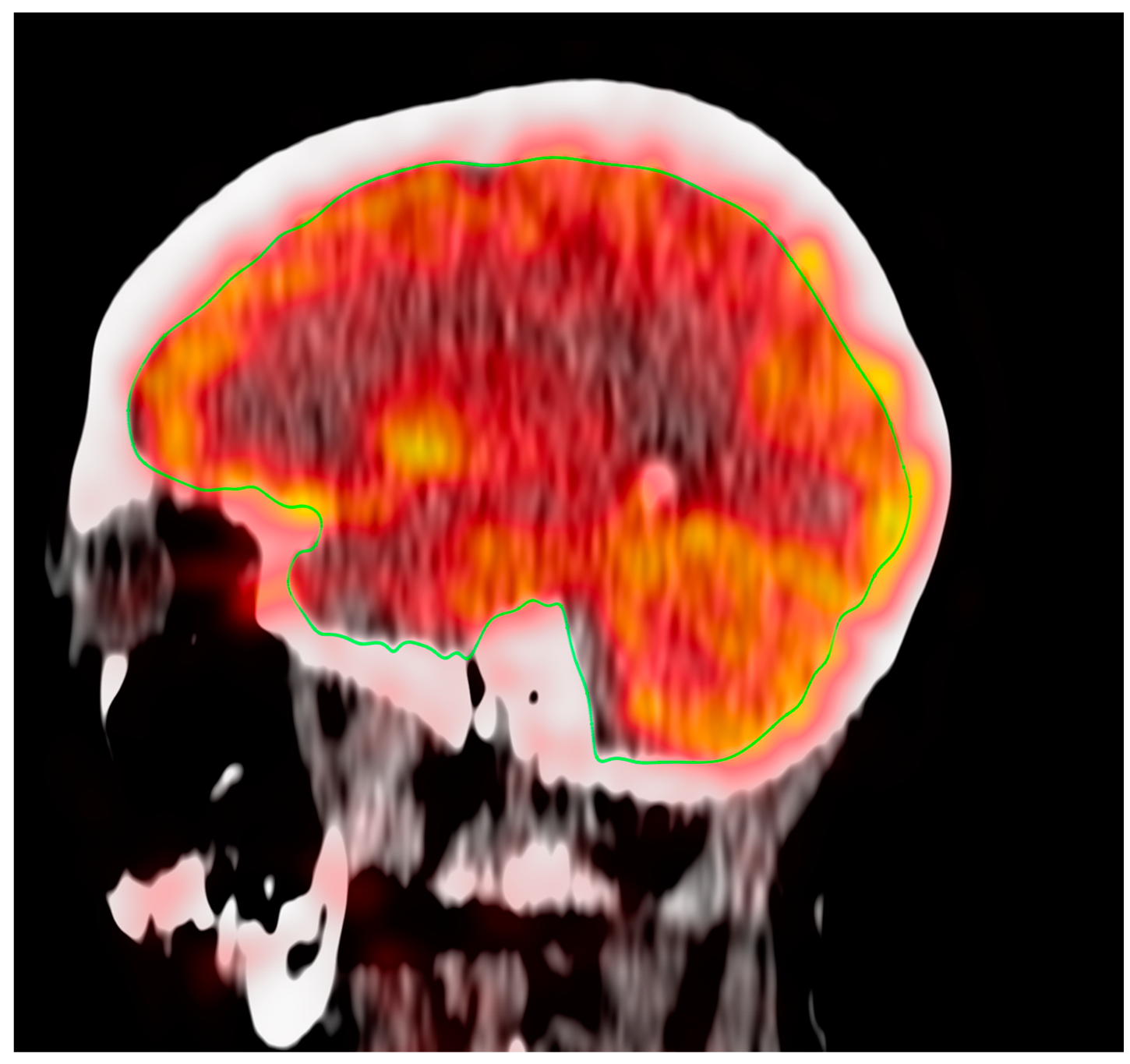
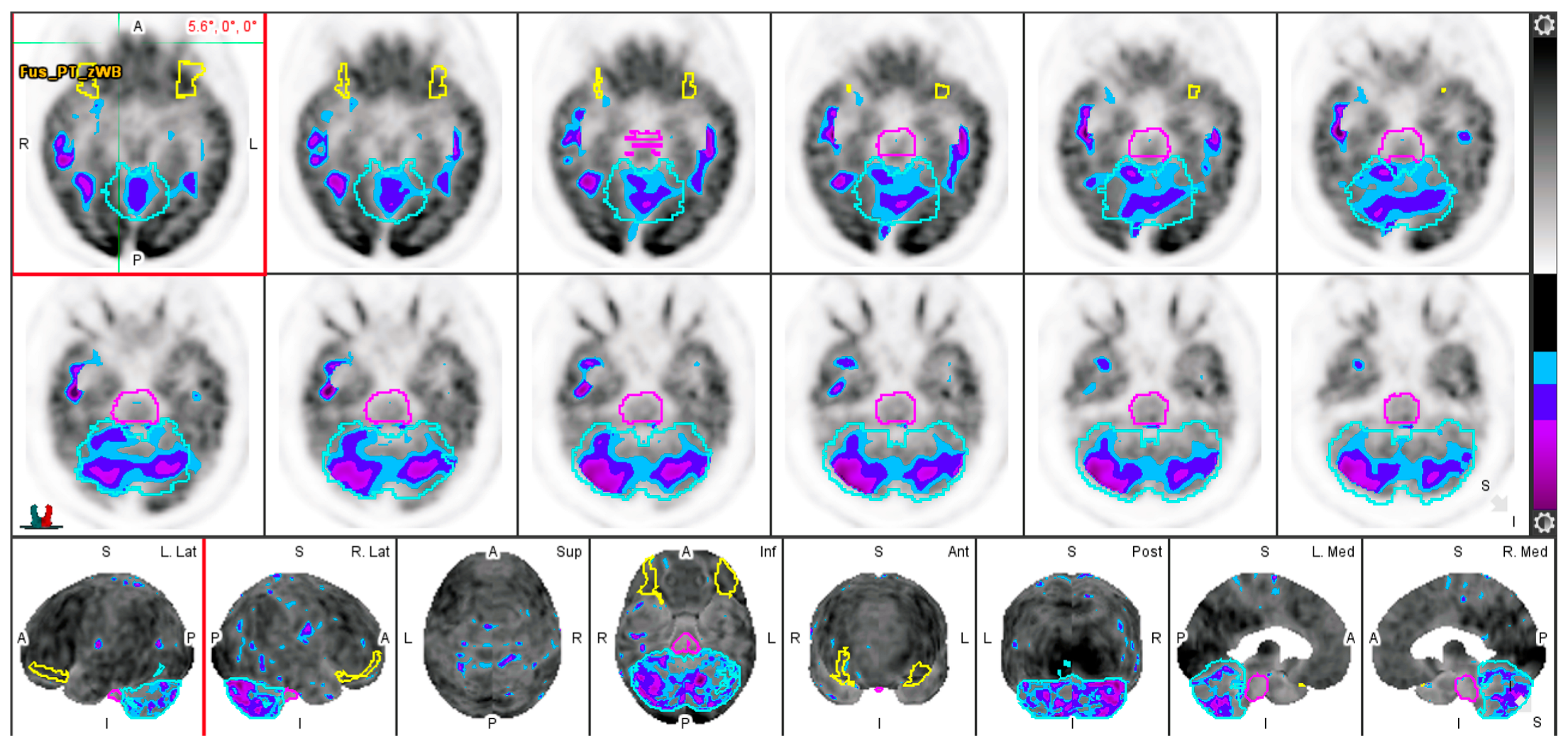
3.3. Effect of Aging on Regional Cerebral Metabolic Activity
4. Discussion
5. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Blinkouskaya, Y.; Weickenmeier, J. Brain Shape Changes Associated with Cerebral Atrophy in Healthy Aging and Alzheimer’s Disease. Front. Mech. Eng. 2021, 7, 705653. [Google Scholar] [CrossRef]
- Schott, J.M. The Neurology of Ageing: What Is Normal? Pract. Neurol. 2017, 17, 172–182. [Google Scholar] [CrossRef]
- Mattson, M.P.; Arumugam, T.V. Hallmarks of Brain Aging: Adaptive and Pathological Modification by Metabolic States. Cell Metab. 2018, 27, 1176–1199. [Google Scholar] [CrossRef]
- Lee, J.; Kim, H.-J. Normal Aging Induces Changes in the Brain and Neurodegeneration Progress: Review of the Structural, Biochemical, Metabolic, Cellular, and Molecular Changes. Front. Aging Neurosci. 2022, 14, 931536. [Google Scholar] [CrossRef] [PubMed]
- Castelli, V.; Benedetti, E.; Antonosante, A.; Catanesi, M.; Pitari, G.; Ippoliti, R.; Cimini, A.; d’Angelo, M. Neuronal Cells Rearrangement during Aging and Neurodegenerative Disease: Metabolism, Oxidative Stress and Organelles Dynamic. Front. Mol. Neurosci. 2019, 12, 132. [Google Scholar] [CrossRef] [PubMed]
- Massaad, C.A.; Klann, E. Reactive Oxygen Species in the Regulation of Synaptic Plasticity and Memory. Antioxid. Redox Signal. 2011, 14, 2013–2054. [Google Scholar] [CrossRef]
- Goyal, M.S.; Hawrylycz, M.; Miller, J.A.; Snyder, A.Z.; Raichle, M.E. Aerobic Glycolysis in the Human Brain Is Associated with Development and Neotenous Gene Expression. Cell Metab. 2014, 19, 49–57. [Google Scholar] [CrossRef]
- Goyal, M.S.; Vlassenko, A.G.; Blazey, T.M.; Su, Y.; Couture, L.E.; Durbin, T.J.; Bateman, R.J.; Benzinger, T.L.-S.; Morris, J.C.; Raichle, M.E. Loss of Brain Aerobic Glycolysis in Normal Human Aging. Cell Metab. 2017, 26, 353–360.e3. [Google Scholar] [CrossRef] [PubMed]
- Harada, N.; Nishiyama, S.; Satoh, K.; Fukumoto, D.; Kakiuchi, T.; Tsukada, H. Age-Related Changes in the Striatal Dopaminergic System in the Living Brain: A Multiparametric PET Study in Conscious Monkeys. Synapse 2002, 45, 38–45. [Google Scholar] [CrossRef]
- Kumar, J.S.D.; Mann, J.J. PET Tracers for Serotonin Receptors and Their Applications. Cent. Nerv. Syst. Agents Med. Chem. 2014, 14, 96–112. [Google Scholar] [CrossRef]
- Yuan, Y.; Gu, Z.-X.; Wei, W.-S. Fluorodeoxyglucose-Positron-Emission Tomography, Single-Photon Emission Tomography, and Structural MR Imaging for Prediction of Rapid Conversion to Alzheimer Disease in Patients with Mild Cognitive Impairment: A Meta-Analysis. AJNR Am. J. Neuroradiol. 2009, 30, 404–410. [Google Scholar] [CrossRef]
- Kato, T.; Inui, Y.; Nakamura, A.; Ito, K. Brain Fluorodeoxyglucose (FDG) PET in Dementia. Ageing Res. Rev. 2016, 30, 73–84. [Google Scholar] [CrossRef] [PubMed]
- Sipilä, H.T.; Clark, J.C.; Peltola, O.; Teräs, M. An automatic [15O]H2O production system for heart and brain studies. J. Label. Comp. Radiopharm. 2001, 44, S1066–S1068. [Google Scholar] [CrossRef]
- Friedland, R.P.; Jagust, W.J.; Huesman, R.H.; Koss, E.; Knittel, B.; Mathis, C.A.; Ober, B.A.; Mazoyer, B.M.; Budinger, T.F. Regional Cerebral Glucose Transport and Utilization in Alzheimer’s Disease. Neurology 1989, 39, 1427–1434. [Google Scholar] [CrossRef]
- Jagust, W.J.; Seab, J.P.; Huesman, R.H.; Valk, P.E.; Mathis, C.A.; Reed, B.R.; Coxson, P.G.; Budinger, T.F. Diminished Glucose Transport in Alzheimer’s Disease: Dynamic PET Studies. J. Cereb. Blood Flow. Metab. 1991, 11, 323–330. [Google Scholar] [CrossRef]
- De Leon, M.J.; Convit, A.; Wolf, O.T.; Tarshish, C.Y.; DeSanti, S.; Rusinek, H.; Tsui, W.; Kandil, E.; Scherer, A.J.; Roche, A.; et al. Prediction of Cognitive Decline in Normal Elderly Subjects with 2-[18F]Fluoro-2-Deoxy-d-Glucose/Positron-Emission Tomography (FDG/PET). Proc. Natl. Acad. Sci. USA 2001, 98, 10966–10971. [Google Scholar] [CrossRef] [PubMed]
- Blomberg, B.A.; de Jong, P.A.; Thomassen, A.; Lam, M.G.E.; Vach, W.; Olsen, M.H.; Mali, W.P.T.M.; Narula, J.; Alavi, A.; Høilund-Carlsen, P.F. Thoracic Aorta Calcification but Not Inflammation Is Associated with Increased Cardiovascular Disease Risk: Results of the CAMONA Study. Eur. J. Nucl. Med. Mol. Imaging 2017, 44, 249–258. [Google Scholar] [CrossRef] [PubMed]
- Teichner, E.M.; You, J.C.; Hriso, C.; Wintering, N.A.; Zabrecky, G.P.; Alavi, A.; Bazzan, A.J.; Monti, D.A.; Newberg, A.B. Alterations in Cerebral Glucose Metabolism as Measured by 18F-Fluorodeoxyglucose-PET in Patients with Persistent Postconcussion Syndrome. Nucl. Med. Commun. 2021, 42, 772–781. [Google Scholar] [CrossRef] [PubMed]
- Partovi, S.; Yuh, R.; Pirozzi, S.; Lu, Z.; Couturier, S.; Grosse, U.; Schluchter, M.D.; Nelson, A.; Jones, R.; O’Donnell, J.K.; et al. Diagnostic Performance of an Automated Analysis Software for the Diagnosis of Alzheimer’s Dementia with 18F FDG PET. Am. J. Nucl. Med. Mol. Imaging 2017, 7, 12–23. [Google Scholar]
- Deery, H.A.; Di Paolo, R.; Moran, C.; Egan, G.F.; Jamadar, S.D. Lower Brain Glucose Metabolism in Normal Ageing Is Predominantly Frontal and Temporal: A Systematic Review and Pooled Effect Size and Activation Likelihood Estimates Meta-analyses. Hum. Brain Mapp. 2022, 44, 1251–1277. [Google Scholar] [CrossRef]
- Hirose, S.; Osada, T.; Ogawa, A.; Tanaka, M.; Wada, H.; Yoshizawa, Y.; Imai, Y.; Machida, T.; Akahane, M.; Shirouzu, I.; et al. Lateral-Medial Dissociation in Orbitofrontal Cortex-Hypothalamus Connectivity. Front. Hum. Neurosci. 2016, 10, 244. [Google Scholar] [CrossRef]
- Nestor, P.G.; Nakamura, M.; Niznikiewicz, M.; Thompson, E.; Levitt, J.J.; Choate, V.; Shenton, M.E.; McCarley, R.W. In Search of the Functional Neuroanatomy of Sociality: MRI Subdivisions of Orbital Frontal Cortex and Social Cognition. Soc. Cogn. Affect. Neurosci. 2013, 8, 460–467. [Google Scholar] [CrossRef] [PubMed]
- Ishibashi, K.; Onishi, A.; Fujiwara, Y.; Oda, K.; Ishiwata, K.; Ishii, K. Longitudinal Effects of Aging on 18F-FDG Distribution in Cognitively Normal Elderly Individuals. Sci. Rep. 2018, 8, 11557. [Google Scholar] [CrossRef]
- Manto, M.; Bower, J.M.; Conforto, A.B.; Delgado-García, J.M.; da Guarda, S.N.F.; Gerwig, M.; Habas, C.; Hagura, N.; Ivry, R.B.; Mariën, P.; et al. Consensus Paper: Roles of the Cerebellum in Motor Control—The Diversity of Ideas on Cerebellar Involvement in Movement. Cerebellum 2012, 11, 457–487. [Google Scholar] [CrossRef]
- Schmahmann, J.D.; Ko, R.; MacMore, J. The Human Basis Pontis: Motor Syndromes and Topographic Organization. Brain 2004, 127, 1269–1291. [Google Scholar] [CrossRef]
- Koziol, L.F.; Budding, D.; Andreasen, N.; D’Arrigo, S.; Bulgheroni, S.; Imamizu, H.; Ito, M.; Manto, M.; Marvel, C.; Parker, K.; et al. Consensus Paper: The Cerebellum’s Role in Movement and Cognition. Cerebellum 2014, 13, 151–177. [Google Scholar] [CrossRef]
- Glickstein, M.; Doron, K. Cerebellum: Connections and Functions. Cerebellum 2008, 7, 589–594. [Google Scholar] [CrossRef]
- Ward, N.S. Compensatory Mechanisms in the Aging Motor System. Ageing Res. Rev. 2006, 5, 239–254. [Google Scholar] [CrossRef]
- Hou, Y.; Dan, X.; Babbar, M.; Wei, Y.; Hasselbalch, S.G.; Croteau, D.L.; Bohr, V.A. Ageing as a Risk Factor for Neurodegenerative Disease. Nat. Rev. Neurol. 2019, 15, 565–581. [Google Scholar] [CrossRef] [PubMed]
- Dickerson, B.C.; Bakkour, A.; Salat, D.H.; Feczko, E.; Pacheco, J.; Greve, D.N.; Grodstein, F.; Wright, C.I.; Blacker, D.; Rosas, H.D.; et al. The Cortical Signature of Alzheimer’s Disease: Regionally Specific Cortical Thinning Relates to Symptom Severity in Very Mild to Mild AD Dementia and Is Detectable in Asymptomatic Amyloid-Positive Individuals. Cereb. Cortex 2009, 19, 497–510. [Google Scholar] [CrossRef] [PubMed]
- Planche, V.; Manjon, J.V.; Mansencal, B.; Lanuza, E.; Tourdias, T.; Catheline, G.; Coupé, P. Structural Progression of Alzheimer’s Disease over Decades: The MRI Staging Scheme. Brain Commun. 2022, 4, fcac109. [Google Scholar] [CrossRef]
- Jin, K.; Zhang, T.; Shaw, M.; Sachdev, P.; Cherbuin, N. Relationship Between Sulcal Characteristics and Brain Aging. Front. Aging Neurosci. 2018, 10, 339. [Google Scholar] [CrossRef] [PubMed]
- Dickstein, D.L.; Kabaso, D.; Rocher, A.B.; Luebke, J.I.; Wearne, S.L.; Hof, P.R. Changes in the Structural Complexity of the Aged Brain. Aging Cell 2007, 6, 275–284. [Google Scholar] [CrossRef]
- Nedergaard, M.; Goldman, S.A. Glymphatic Failure as a Final Common Pathway to Dementia. Science 2020, 370, 50–56. [Google Scholar] [CrossRef]
- Peters, R. Ageing and the Brain. Postgrad. Med. J. 2006, 82, 84–88. [Google Scholar] [CrossRef] [PubMed]
- Sweeney, M.D.; Kisler, K.; Montagne, A.; Toga, A.W.; Zlokovic, B.V. The Role of Brain Vasculature in Neurodegenerative Disorders. Nat. Neurosci. 2018, 21, 1318–1331. [Google Scholar] [CrossRef] [PubMed]
- Hurtz, S.; Woo, E.; Kebets, V.; Green, A.E.; Zoumalan, C.; Wang, B.; Ringman, J.M.; Thompson, P.M.; Apostolova, L.G. Age Effects on Cortical Thickness in Cognitively Normal Elderly Individuals. Dement. Geriatr. Cogn. Dis. Extra 2014, 4, 221–227. [Google Scholar] [CrossRef] [PubMed]
- Nugent, S.; Castellano, C.-A.; Goffaux, P.; Whittingstall, K.; Lepage, M.; Paquet, N.; Bocti, C.; Fulop, T.; Cunnane, S.C. Glucose Hypometabolism Is Highly Localized, but Lower Cortical Thickness and Brain Atrophy Are Widespread in Cognitively Normal Older Adults. Am. J. Physiol. Endocrinol. Metab. 2014, 306, E1315–E1321. [Google Scholar] [CrossRef] [PubMed]
- Chételat, G.; Desgranges, B.; Landeau, B.; Mézenge, F.; Poline, J.B.; de la Sayette, V.; Viader, F.; Eustache, F.; Baron, J.-C. Direct Voxel-Based Comparison between Grey Matter Hypometabolism and Atrophy in Alzheimer’s Disease. Brain 2008, 131, 60–71. [Google Scholar] [CrossRef]
- Lee, J.; Burkett, B.J.; Min, H.-K.; Senjem, M.L.; Lundt, E.S.; Botha, H.; Graff-Radford, J.; Barnard, L.R.; Gunter, J.L.; Schwarz, C.G.; et al. Deep Learning-Based Brain Age Prediction in Normal Aging and Dementia. Nat. Aging 2022, 2, 412–424. [Google Scholar] [CrossRef] [PubMed]
- Mosconi, L.; Tsui, W.H.; Herholz, K.; Pupi, A.; Drzezga, A.; Lucignani, G.; Reiman, E.M.; Holthoff, V.; Kalbe, E.; Sorbi, S.; et al. Multicenter Standardized 18F-FDG PET Diagnosis of Mild Cognitive Impairment, Alzheimer’s Disease, and Other Dementias. J. Nucl. Med. 2008, 49, 390–398. [Google Scholar] [CrossRef] [PubMed]
- Mosconi, L.; Mistur, R.; Switalski, R.; Tsui, W.H.; Glodzik, L.; Li, Y.; Pirraglia, E.; De Santi, S.; Reisberg, B.; Wisniewski, T.; et al. 18F-FDG-PET/CT Changes in Brain Glucose Metabolism from Normal Cognition to Pathologically Verified Alzheimer’s Disease. Eur. J. Nucl. Med. Mol. Imaging 2009, 36, 811–822. [Google Scholar] [CrossRef] [PubMed]


{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Demographics | |
| Female, n (%) | 33 (82.5) |
| Age, years | 35.8 ± 13.1 |
| Body mass index, kg/m2 | 27.4 ± 4.5 |
| Comorbidities | |
| Smoking history, n (%) | 17 (42.5) |
| Family history of coronary artery disease, n (%) | 13 (32.5) |
| Arterial hypertension, n (%) | 10 (25.0) |
| Hypercholesterolemia, n (%) | 6 (15.0) |
| Atrial fibrillation, n (%) | 3 (7.5) |
| Heart valve disease, n (%) | 2 (5.0) |
| Peripheral artery disease, n (%) | 1 (2.5) |
| History of previous stroke/transient ischemic attack, n (%) | 0 (0.0) |
| Laboratory tests | |
| Total cholesterol, mmol/L | 5.2 ± 0.8 |
| HDL cholesterol, mmol/L | 3.3 ± 0.8 |
| LDL cholesterol, mmol/L | 1.4 ± 0.4 |
| Triglycerides, mmol/L | 1.1 ± 0.7 |
| HbA1c, mmol/L | 36.4 ± 6.2 |
| C-reactive protein, mg/L | 2.6 ± 4.1 |
| White blood cell count, 109 cells/L | 6.3 ± 2.5 |
| Fibrinogen, μmol/L | 9.6 ± 1.6 |
| Creatinine, μmol/L | 81.5 ± 13 |
| Estimated glomerular filtration rate, mL/min/1.73 m2 | 79.1 ± 13.4 |
| Medications | |
| Aspirin, n (%) | 6 (15.0) |
| Beta blockers, n (%) | 5 (12.5) |
| Angiotensin-converting enzyme inhibitors/angiotensin receptor blockers, n (%) | 5 (12.5) |
| Lipid-lowering medication, n (%) | 3 (7.5) |
| Risk profile | |
| 10-year Framingham risk, % (25–75th percentile) | 4.7 (0.7–4.2) |
| CHADS-VASc score, % (25–75th percentile) | 0.8 (0–1) |
| Level of physical activity, % (25–75th percentile) | 2.1 (1.3–3) |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Subtirelu, R.C.; Teichner, E.M.; Su, Y.; Al-Daoud, O.; Patel, M.; Patil, S.; Writer, M.; Werner, T.; Revheim, M.-E.; Høilund-Carlsen, P.F.; et al. Aging and Cerebral Glucose Metabolism: 18F-FDG-PET/CT Reveals Distinct Global and Regional Metabolic Changes in Healthy Patients. Life 2023, 13, 2044. https://doi.org/10.3390/life13102044
Subtirelu RC, Teichner EM, Su Y, Al-Daoud O, Patel M, Patil S, Writer M, Werner T, Revheim M-E, Høilund-Carlsen PF, et al. Aging and Cerebral Glucose Metabolism: 18F-FDG-PET/CT Reveals Distinct Global and Regional Metabolic Changes in Healthy Patients. Life. 2023; 13(10):2044. https://doi.org/10.3390/life13102044
Chicago/Turabian StyleSubtirelu, Robert Christopher, Eric Michael Teichner, Yvonne Su, Omar Al-Daoud, Milan Patel, Shiv Patil, Milo Writer, Thomas Werner, Mona-Elisabeth Revheim, Poul Flemming Høilund-Carlsen, and et al. 2023. "Aging and Cerebral Glucose Metabolism: 18F-FDG-PET/CT Reveals Distinct Global and Regional Metabolic Changes in Healthy Patients" Life 13, no. 10: 2044. https://doi.org/10.3390/life13102044
APA StyleSubtirelu, R. C., Teichner, E. M., Su, Y., Al-Daoud, O., Patel, M., Patil, S., Writer, M., Werner, T., Revheim, M.-E., Høilund-Carlsen, P. F., & Alavi, A. (2023). Aging and Cerebral Glucose Metabolism: 18F-FDG-PET/CT Reveals Distinct Global and Regional Metabolic Changes in Healthy Patients. Life, 13(10), 2044. https://doi.org/10.3390/life13102044