
Effects of Cannabidiol on Locomotor Activity

 ,
,  , and
, and
Abstract
:1. Introduction
2. Materials and Methods
3. Results
3.1. Effects of Cannabidiolic Acid, Precursor of Cannabidiol, on Locomotor Activity
3.2. Effects of Cannabidiol on Locomotor Activity
3.3. Effects of Cannabidiol on Locomotor Modifications Induced by Experimental Models of Diseases
3.4. Effects of Cannabidiol Combination with Δ9-Tetrahydrocannabinol on Locomotor Activity
3.5. Effects of Cannabidiol on Locomotor Changes Induced by Other Substances

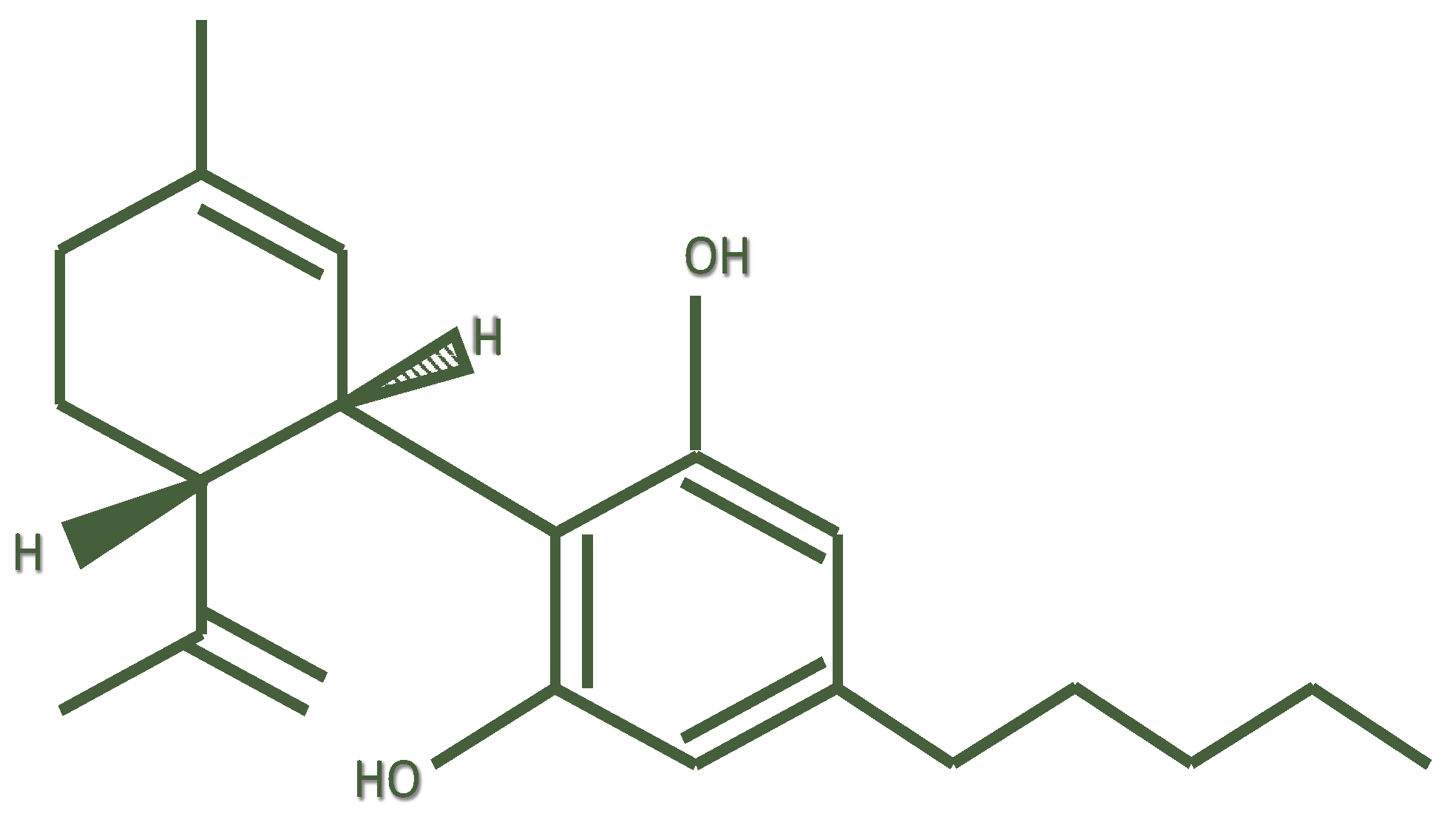{kind=link}
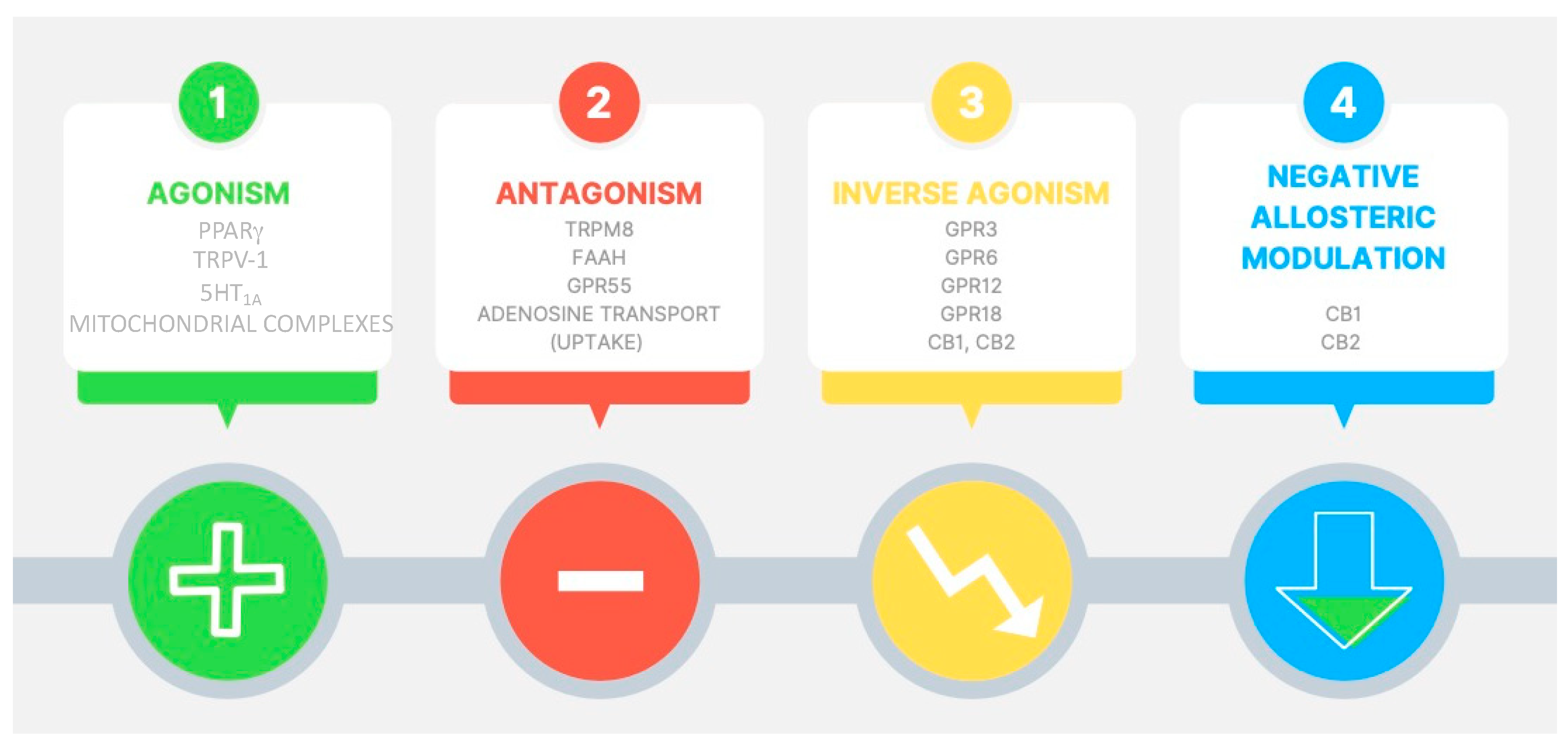{kind=link}
| Species | Substance | CBD | Locomotor Activity | Reference |
|---|---|---|---|---|
| Rat | Single i.p. rimonabant (0.1 mg/kg; i.p.) | Single i.p. CBD (20 mg/kg) | No effect of rimonabant on CBD-induced reduction of vertical activity. | [69] |
| Single i.p. WAY-100,635 (0.5 mg/kg) | Single i.p. CBD (20 mg/kg) | WAY-100,635 restored CBD-induced reduction of vertical activity. | ||
| Single i.p. 8-OH-DPAT (1 mg/kg) | Single i.p. CBD (20 mg/kg) | CBD potentiated 8-OH-DPAT reduction of vertical activity. | ||
| Rat | Single oral haloperidol (5 mg/kg) | Single oral CBD (3, 5, 10 mg/kg) | Haloperidol or CBD caused an increase in time spent in the open arms. This effect was slightly reduced by concomitant CBD/haloperidol administration. | [75] |
| Haloperidol reduced motor activity. CBD (5 mg/kg) antagonized hypolocomotion induced by haloperidol. | ||||
| Mouse | Daily progressive i.p. dose of cocaine (15 to 60 mg/kg) three times a day for 12 days | Single i.p. CBD (10, 20, and 40 mg/kg) given 270 min after the last cocaine administration | All doses of CBD blocked the increase in total distance values induced by cocaine withdrawal. | [76] |
| Rat | Single i.p. ketamine (30, 60 mg/kg) alone or 10 min after CBD | Single i.p. CBD (10 or 30 mg/kg) alone or 10 min before ketamine | CBD (30 mg/kg) alone decreased motor activity; ketamine (30, 60 mg/kg) alone increased motor activity. CBD did not reverse ketamine-induced short-lasting hyperactivity but prolonged it over time. | [78] |
| Mouse | Single i.p. S-ketamine (2.5, 3, 5, 10, 30 mg/kg) | I.p. CBD (3, 10, 30 mg/kg) 30 min before S-ketamine | CBD produced antidepressant effects without any influence on motor activity. Higher dose of S-ketamine showed antidepressant effects with increase in motor activity. | [79] |
4. Discussion
5. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Conflicts of Interest
References
- Turner, C.E.; Hadley, K.W. Constituents of Cannabis sativa L. 3. Clear and discrete separation of cannabidiol and cannabichromene. J. Pharm. Sci. 1973, 62, 1083–1086. [Google Scholar] [CrossRef] [PubMed]
- Mechoulam, R.; Shvo, Y. Hashish—I: The structure of Cannabidiol. Tetrahedron 1963, 19, 2073–2078. [Google Scholar] [CrossRef]
- Grotenhermen, F. Pharmacokinetics and pharmacodynamics of cannabinoids. Clin. Pharmacokinet. 2003, 42, 327–360. [Google Scholar] [CrossRef] [PubMed]
- Pattnaik, F.; Nanda, S.; Mohanty, S.; Dalai, A.K.; Kumar, V.; Ponnusamy, S.K.; Naik, S. Cannabis: Chemistry, extraction and therapeutic applications. Chemosphere 2022, 289, 133012. [Google Scholar] [CrossRef] [PubMed]
- Tura, M.; Mandrioli, M.; Toschi, T.G. Preliminary Study: Comparison of Antioxidant Activity of Cannabidiol (CBD) and α-Tocopherol Added to Refined Olive and Sunflower Oils. Molecules 2019, 24, 3485. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Burstein, S. Cannabidiol (CBD) and its analogs: A review of their effects on inflammation. Bioorg. Med. Chem. 2015, 23, 1377–1385. [Google Scholar] [CrossRef]
- Calapai, F.; Cardia, L.; Sorbara, E.E.; Navarra, M.; Gangemi, S.; Calapai, G.; Mannucci, C. Cannabinoids, Blood-Brain Barrier, and Brain Disposition. Pharmaceutics 2020, 12, 265. [Google Scholar] [CrossRef] [Green Version]
- Karaźniewicz-Łada, M.; Główka, A.K.; Mikulska, A.A.; Główka, F.K. Pharmacokinetic Drug-Drug Interactions among Antiepileptic Drugs, Including CBD, Drugs Used to Treat COVID-19 and Nutrients. Int. J. Mol. Sci. 2021, 22, 9582. [Google Scholar] [CrossRef]
- Jiang, R.; Yamaori, S.; Takeda, S.; Yamamoto, I.; Watanabe, K. Identification of cytochrome P450 enzymes responsible for metabolism of cannabidiol by human liver microsomes. Life Sci. 2011, 89, 165–170. [Google Scholar] [CrossRef]
- Watanabe, K.; Usami, N.; Osada, S.; Narimatsu, S.; Yamamoto, I.; Yoshimura, H. Cannabidiol metabolism revisited: Tentative identification of novel decarbonylated metabolites of cannabidiol formed by human liver microsomes and recombinant cytochrome P450 3A4. Forensic Toxicol. 2019, 37, 449–455. [Google Scholar] [CrossRef] [Green Version]
- Ujváry, I.; Hanuš, L. Human Metabolites of Cannabidiol: A Review on Their Formation, Biological Activity, and Relevance in Therapy. Cannabis Cannabinoid Res. 2016, 1, 90–101. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Petitet, F.; Jeantaud, B.; Reibaud, M.; Imperato, A.; Dubroeucq, M.C. Complex pharmacology of natural cannabinoids: Evidence for partial agonist activity of delta9-tetrahydrocannabinol and antagonist activity of cannabidiol on rat brain cannabinoid receptors. Life Sci. 1998, 63, PL1–PL6. [Google Scholar] [CrossRef]
- Mechoulam, R.; Peters, M.; Murillo-Rodriguez, E.; Hanus, L.O. Cannabidiol--recent advances. Chem. Biodivers. 2007, 4, 1678–1692. [Google Scholar] [CrossRef] [PubMed]
- McPartland, J.M.; Duncan, M.; Di Marzo, V.; Pertwee, R.G. Are cannabidiol and Δ(9)-tetrahydrocannabivarin negative modulators of the endocannabinoid system? A systematic review. Br. J. Pharmacol. 2015, 172, 737–753. [Google Scholar] [CrossRef] [Green Version]
- Russo, E.B. Cannabidiol claims and misconceptions. Trends Pharmacol. Sci. 2017, 38, 198–201. [Google Scholar] [CrossRef]
- Pertwee, R.G. The diverse CB1 and CB2 receptor pharmacology of three plant cannabinoids: Δ9-tetrahydrocannabinol, cannabidiol and Δ9-tetrahydrocannabivarin. Br. J. Pharmacol. 2008, 153, 199–215. [Google Scholar] [CrossRef] [Green Version]
- Elsohly, M.A.; Slade, D. Chemical constituents of marijuana: The complex mixture of natural cannabinoids. Life Sci. 2005, 78, 539–548. [Google Scholar] [CrossRef]
- Iannotti, F.A.; Hill, C.L.; Leo, A.; Alhusaini, A.; Soubrane, C.; Mazzarella, E.; Russo, E.; Whalley, B.J.; Di Marzo, V.; Stephens, G.J. Nonpsychotropic plant cannabinoids, cannabidivarin (CBDV) and cannabidiol (CBD), activate and desensitize transient receptor potential vanilloid 1 (TRPV1) channels in vitro: Potential for the treatment of neuronal hyperexcitability. ACS Chem. Neurosci. 2014, 5, 1131–1141. [Google Scholar] [CrossRef] [Green Version]
- Chiba, T.; Ueno, S.; Obara, Y.; Nakahata, N. A synthetic cannabinoid, CP55940, inhibits lipopolysaccharide-induced cytokine mRNA expression in a cannabinoid receptor-independent mechanism in rat cerebellar granule cells. J. Pharm. Pharmacol. 2011, 63, 636–647. [Google Scholar] [CrossRef]
- de Filippis, D.; Iuvone, T.; d’amico, A.; Esposito, G.; Steardo, L.; Herman, A.G.; Pelckmans, P.A.; de Winter, B.Y.; de Man, J.G. Effect of cannabidiol on sepsis-induced motility disturbances in mice: Involvement of CB receptors and fatty acid amide hydrolase. Neurogastroenterol. Motil. 2008, 20, 919–927. [Google Scholar] [CrossRef]
- De Gregorio, D.; McLaughlin, R.J.; Posa, L.; Ochoa-Sanchez, R.; Enns, J.; Lopez-Canul, M.; Aboud, M.; Maione, S.; Comai, S.; Gobbi, G. Cannabidiol modulates serotonergic transmission and reverses both allodynia and anxiety-like behavior in a model of neuropathic pain. Pain 2019, 160, 136–150. [Google Scholar] [CrossRef] [PubMed]
- O’Sullivan, S.E.; Kendall, D.A. Cannabinoid activation of peroxisome proliferator-activated receptors: Potential for modulation of inflammatory disease. Immunobiology 2010, 215, 611–616. [Google Scholar] [CrossRef] [PubMed]
- Pandolfo, P.; Silveirinha, V.; dos Santos-Rodrigues, A.; Venance, L.; Ledent, C.; Takahashi, R.N.; Cunha, R.A.; Köfalvi, A. Cannabinoids inhibit the synaptic uptake of adenosine and dopamine in the rat and mouse striatum. Eur. J. Pharmacol. 2011, 655, 38–45. [Google Scholar] [CrossRef] [PubMed]
- Mazeh, A.C.; Angus, J.A.; Wright, C.E. Cannabidiol selectively inhibits the contraction of rat small resistance arteries: Possible role for CGRP and voltage-gated calcium channels. Eur. J. Pharmacol. 2021, 891, 173767. [Google Scholar] [CrossRef]
- Kathmann, M.; Flau, K.; Redmer, A.; Tränkle, C.; Schlicker, E. Cannabidiol is an allosteric modulator at mu- and delta-opioid receptors. Naunyn-Schmiedebergs Arch. Exp. Pharmakol. 2006, 372, 354–361. [Google Scholar] [CrossRef]
- Song, J.H.; Shin, S.H.; Wang, W.; Ross, G.M. Involvement of oxidative stress in ascorbate-induced proapoptotic death of PC12 cells. Exp. Neurol. 2001, 169, 425–437. [Google Scholar] [CrossRef]
- Ryan, D.; Drysdale, A.J.; Lafourcade, C.; Pertwee, R.G.; Platt, B. Cannabidiol targets mitochondria to regulate intracellular Ca2+ levels. J. Neurosci. 2009, 29, 2053–2063. [Google Scholar] [CrossRef]
- Kim, J.; Choi, H.; Kang, E.K.; Ji, G.Y.; Kim, Y.; Choi, I.S. In Vitro Studies on Therapeutic Effects of Cannabidiol in Neural Cells: Neurons, Glia, and Neural Stem Cells. Molecules 2021, 26, 6077. [Google Scholar] [CrossRef]
- Watt, G.; Karl, T. In vivo Evidence for Therapeutic Properties of Cannabidiol (CBD) for Alzheimer’s Disease. Front Pharmacol. 2017, 8, 20. [Google Scholar] [CrossRef] [Green Version]
- Mannucci, C.; Navarra, M.; Calapai, F.; Spagnolo, E.V.; Busardò, F.P.; Cas, R.D.; Ippolito, F.M.; Calapai, G. Neurological Aspects of Medical Use of Cannabidiol. CNS Neurol. Disord. Drug Targets 2017, 16, 541–553. [Google Scholar] [CrossRef] [Green Version]
- McCartney, D.; Benson, M.J.; Desbrow, B.; Irwin, C.; Suraev, A.; McGregor, I.S. Cannabidiol and Sports Performance: A Narrative Review of Relevant Evidence and Recommendations for Future Research. Sports Med. Open. 2020, 6, 27. [Google Scholar] [CrossRef] [PubMed]
- Iffland, K.; Grotenhermen, F. An Update on Safety and Side Effects of Cannabidiol: A Review of Clinical Data and Relevant Animal Studies. Cannabis Cannabinoid Res. 2017, 2, 139–154. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Koturbash, I.; MacKay, D. Cannabidiol and Other Cannabinoids: From Toxicology and Pharmacology to the Development of a Regulatory Pathway. J. Diet. Suppl. 2020, 17, 487–492. [Google Scholar] [CrossRef]
- Mokrý, J. Experimental models and behavioural tests used in the study of Parkinson’s disease. Physiol. Res. 1995, 44, 143–150. [Google Scholar] [PubMed]
- Perras, C. Sativex for the management of multiple sclerosis symptoms. Issues Emerg. Health Technol. 2005, 72, 1–4. [Google Scholar]
- von Wrede, R.; Helmstaedter, C.; Surges, R. Cannabidiol in the Treatment of Epilepsy. Clin. Drug Investig. 2021, 41, 211–220. [Google Scholar] [CrossRef]
- Formato, M.; Crescente, G.; Scognamiglio, M.; Fiorentino, A.; Pecoraro, M.T.; Piccolella, S.; Catauro, M.; Pacifico, S. (−)-Cannabidiolic Acid, a Still Overlooked Bioactive Compound: An Introductory Review and Preliminary Research. Molecules 2020, 25, 2638. [Google Scholar] [CrossRef]
- Gupta, K.; Walton, R.; Kataria, S. Chemotherapy-Induced Nausea and Vomiting: Pathogenesis, Recommendations, and New Trends. Cancer Treat. Res. Commun. 2021, 26, 100278. [Google Scholar] [CrossRef]
- Brierley, D.I.; Samuels, J.; Duncan, M.; Whalley, B.J.; Williams, C.M. Neuromotor tolerability and behavioural characterisation of cannabidiolic acid, a phytocannabinoid with therapeutic potential for anticipatory nausea. Psychopharmacology 2016, 233, 243–254. [Google Scholar] [CrossRef]
- Alegre-Zurano, L.; Martín-Sánchez, A.; Valverde, O. Behavioural and molecular effects of cannabidiolic acid in mice. Life Sci. 2020, 259, 118271. [Google Scholar] [CrossRef]
- Viudez-Martinez, A.; García-Gutiérrez, M.S.; Medrano-Relinque, J.; Navarron, C.; Navarrete, F.; Manzanares, J. Cannabidiol does not display drug abuse potential in mice behavior. Acta Pharmacol. Sin. 2019, 40, 358–364. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Schleicher, E.M.; Ott, F.W.; Müller, M.; Silcher, B.; Sichler, M.E.; Löw, M.J.; Wagner, J.M.; Bouter, Y. Prolonged Cannabidiol Treatment Lacks on Detrimental Effects on Memory, Motor Performance and Anxiety in C57BL/6J Mice. Front. Behav. Neurosci. 2019, 13, 94. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Lueptow, L.M. Novel Object Recognition Test for the Investigation of Learning and Memory in Mice. J. Vis. Exp. 2017, 126, 55718. [Google Scholar] [CrossRef] [PubMed]
- Bourin, M.; Hascoët, M. The mouse light/dark box test. Eur. J. Pharmacol. 2003, 463, 55–65. [Google Scholar] [CrossRef]
- Kraeuter, A.K.; Guest, P.C.; Sarnyai, Z. The Elevated Plus Maze Test for Measuring Anxiety-Like Behavior in Rodents. Methods Mol. Biol. 2019, 1916, 69–74. [Google Scholar]
- Gómez-Nieto, R.; Hormigo, S.; López, D.E. Prepulse Inhibition of the Auditory Startle Reflex Assessment as a Hallmark of Brainstem Sensorimotor Gating Mechanisms. Brain Sci. 2020, 10, 639. [Google Scholar] [CrossRef]
- Peres, F.F.; Levin, R.; Suiama, M.A.; Diana, M.C.; Gouvêa, D.A.; Almeida, V.; Santos, C.M.; Lungato, L.; Zuardi, A.W.; Hallak, J.E.; et al. Cannabidiol Prevents Motor and Cognitive Impairments Induced by Reserpine in Rats. Front. Pharmacol. 2016, 7, 343. [Google Scholar] [CrossRef] [Green Version]
- Garcia-Arencibia, M.; González, S.; DE Lago, E.; Ramos, J.A.; Mechoulam, R.; Fernández-Ruiz, J. Evaluation of the neuroprotective effect of cannabinoids in a rat model of Parkinson’s disease: Importance of antioxidant and cannabinoid receptor-independent properties. Brain Res. 2007, 1134, 162–170. [Google Scholar] [CrossRef]
- Magen, I.; Avraham, Y.; Ackerman, Z.; Vorobiev, L.; Mechoulam, R.; Berry, E.M. Cannabidiol ameliorates cognitive and motor impairments in mice with bile duct ligation. J. Hepatol. 2009, 51, 528–534. [Google Scholar] [CrossRef]
- Geidl, W.; Gobster, C.; Streber, R.; Pfeifer, K. A systematic critical review of physical activity aspects in clinical guidelines for multiple sclerosis. Mult. Scler. Relat. Disord. 2018, 25, 200–207. [Google Scholar] [CrossRef]
- Jones, É.; Vlachou, S. A Critical Review of the Role of the Cannabinoid Compounds Δ9-Tetrahydrocannabinol (Δ9-THC) and Cannabidiol (CBD) and their Combination in Multiple Sclerosis Treatment. Molecules 2020, 25, 4930. [Google Scholar] [CrossRef] [PubMed]
- Rudroff, T.; Sosnoff, J. Cannabidiol to Improve Mobility in People with Multiple Sclerosis. Front. Neurol. 2018, 9, 183. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ciaccio, C.; Fontana, L.; Milani, D.; Tabano, S.; Miozzo, M.; Esposito, S. Fragile X syndrome: A review of clinical and molecular diagnoses. Ital. J. Pediatr. 2017, 43, 39. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Richter, J.D.; Zhao, X. The molecular biology of FMRP: New insights into fragile X syndrome. Nat. Rev. Neurosci. 2021, 22, 209–222. [Google Scholar] [CrossRef] [PubMed]
- Kim, L.; He, L.; Maaswinkel, H.; Zhu, L.; Sirotkin, H.; Weng, W. Anxiety, hyperactivity and stereotypy in a zebrafish model of fragile X syndrome and autism spectrum disorder. Prog. Neuro-Psychopharmacol. Biol. Psychiatry 2014, 55, 40–49. [Google Scholar] [CrossRef]
- Zieba, J.; Sinclair, D.; Sebree, T.; Bonn-Miller, M.; Gutterman, D.; Siegel, S.; Karl, T. Cannabidiol (CBD) reduces anxiety-related behavior in mice via an FMRP-independent mechanism. Pharmacol. Biochem. Behav. 2019, 181, 93–100. [Google Scholar] [CrossRef]
- Lok, K.; Zhao, H.; Shen, H.; Wang, Z.; Gao, X.; Zhao, W.; Yin, M. Characterization of the APP/PS1 mouse model of Alzheimer’s disease in senescence accelerated background. Neurosci. Lett. 2013, 557 Pt B, 84–89. [Google Scholar] [CrossRef]
- Powell, S.B.; Weber, M.; Geyer, M.A. Genetic models of sensorimotor gating: Relevance to neuropsychiatric disorders. Curr. Top. Behav. Neurosci. 2012, 12, 251–318. [Google Scholar]
- Coles, M.; Watt, G.; Kreilaus, F.; Karl, T. Medium-Dose Chronic Cannabidiol Treatment Reverses Object Recognition Memory Deficits of APPSwe/PS1ΔE9 Transgenic Female Mice. Front. Pharmacol. 2020, 11, 587604. [Google Scholar] [CrossRef]
- Florensa-Zanuy, E.; Garro-Martínez, E.; Adell, A.; Castro, E.; Díaz, Á.; Pazos, Á.; Mac-Dowell, K.S.; Martín-Hernández, D.; Pilar-Cuéllar, F. Cannabidiol antidepressant-like effect in the lipopolysaccharide model in mice: Modulation of inflammatory pathways. Biochem. Pharmacol. 2021, 185, 114433. [Google Scholar] [CrossRef]
- Taffe, M.A.; Creehan, K.M.; Vandewater, S.A. Cannabidiol fails to reverse hypothermia or locomotor suppression induced by Δ(9) -tetrahydrocannabinol in Sprague-Dawley rats. Br. J. Pharmacol. 2015, 172, 1783–1791. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Craft, R.M.; Wakley, A.A.; Tsutsui, K.T.; Laggart, J.D. Sex differences in cannabinoid 1 vs. cannabinoid 2 receptor-selective antagonism of antinociception produced by Δ9-tetrahydrocannabinol and CP55, 940 in the rat. J. Pharmacol. Exp. Ther. 2012, 340, 787–800. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Britch, S.C.; Wiley, J.L.; Yu, Z.; Clowers, B.H.; Craft, R.M. Cannabidiol-Δ9-tetrahydrocannabinol interactions on acute pain and locomotor activity. Drug Alcohol. Depend. 2017, 175, 187–197. [Google Scholar] [CrossRef] [PubMed]
- Javadi-Paydar, M.; Nguyen, J.D.; Kerr, T.M.; Grant, Y.; Vandewater, S.A.; Cole, M.; Taffe, M.A. Effects of Δ9-THC and cannabidiol vapor inhalation in male and female rats. Psychopharmacology 2018, 235, 2541–2557. [Google Scholar] [CrossRef] [PubMed]
- Whitey, S.L.; Kangas, B.D.; Charles, S.; Gumbert, A.B.; Eisold, J.E.; George, S.R.; Bergman, J.; Madras, B.K. Effects of daily Δ9-Tetrahydrocannabinol (THC) alone or combined with cannabidiol (CBD) on cognition-based behavior and activity in adolescent nonhuman primates. Drug Alcohol. Depend. 2021, 221, 108629. [Google Scholar]
- Jikomes, N.; Zoorob, M. The Cannabinoid Content of Legal Cannabis in Washington State Varies Systematically Across Testing Facilities and Popular Consumer Products. Sci. Rep. 2018, 8, 4519, Erratum in: Sci. Rep. 2020, 10, 14406. [Google Scholar] [CrossRef] [Green Version]
- Kasten, C.R.; Zhang, Y.; Boehm, S.L., 2nd. Acute Cannabinoids Produce Robust Anxiety-Like and Locomotor Effects in Mice, but Long-Term Consequences Are Age- and Sex-Dependent. Front. Behav. Neurosci. 2019, 13, 32. [Google Scholar] [CrossRef] [Green Version]
- Todd, S.M.; Arnold, J.C. Neural correlates of interactions between cannabidiol and Δ(9) -tetrahydrocannabinol in mice: Implications for medical cannabis. Br. J. Pharmacol. 2016, 173, 53–65. [Google Scholar] [CrossRef] [Green Version]
- Espejo-Porras, F.; Fernández-Ruiz, J.; Pertwee, R.G.; Mechoulam, R.; García, C. Motor effects of the non-psychotropic phytocannabinoid cannabidiol that are mediated by 5-HT1A receptors. Neuropharmacology 2013, 75, 155–163. [Google Scholar] [CrossRef]
- Pertwee, R.G.; Rock, E.M.; Guenther, K.; Limebeer, C.L.; Stevenson, L.A.; Haj, C.; Smoum, R.; Parker, L.A.; Mechoulam, R. Cannabidiolic acid methyl ester, a stable synthetic analogue of cannabidiolic acid, can produce 5-HT1A receptor-mediated suppression of nausea and anxiety in rats. Br. J. Pharmacol. 2018, 175, 100–112. [Google Scholar] [CrossRef] [Green Version]
- Chaves, Y.C.; Genaro, K.; Crippa, J.A.; da Cunha, J.M.; Zanoveli, J.M. Cannabidiol induces antidepressant and anxiolytic-like effects in experimental type-1 diabetic animals by multiple sites of action. Metab. Brain Dis. 2021, 36, 639–652. [Google Scholar] [CrossRef]
- Calapai, G.; Mannucci, C.; Chinou, I.; Cardia, L.; Calapai, F.; Sorbara, E.E.; Firenzuoli, B.; Ricca, V.; Gensini, G.F.; Firenzuoli, F. Preclinical and Clinical Evidence Supporting Use of Cannabidiol in Psychiatry. Evid. Based Complementary Altern. Med. 2019, 2019, 2509129. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Walsh, R.N.; Cummins, R.A. The Open-Field Test: A critical review. Psychol. Bull. 1976, 83, 482–504. [Google Scholar] [CrossRef] [PubMed]
- Reddy, D.S.; Kulkarni, S.K. Role of GABA-A and mitochondrial diazepam binding inhibitor receptors in the anti-stress activity of neurosteroids in mice. Psychopharmacology 1996, 128, 280–292. [Google Scholar] [CrossRef]
- Kajero, J.A.; Seedat, S.; Ohaeri, J.; Akindele, A.; Aina, O. Investigation of the effects of cannabidiol on vacuous chewing movements, locomotion, oxidative stress and blood glucose in rats treated with oral haloperidol. World J. Biol. Psychiatry. 2020, 21, 612–626. [Google Scholar] [CrossRef] [PubMed]
- Gasparyan, A.; Navarrete, F.; Rodríguez-Arias, M.; Miñarro, J.; Manzanares, J. Cannabidiol Modulates Behavioural and Gene Expression Alterations Induced by Spontaneous Cocaine Withdrawal. Neurotherapeutics 2020, 18, 615–623. [Google Scholar] [CrossRef] [PubMed]
- Galaj, E.; Bi, G.H.; Yang, H.J.; Xi, Z.X. Cannabidiol attenuates the rewarding effects of cocaine in rats by CB2, 5-HT1Aand TRPV1 receptor mechanisms. Neuropharmacology 2020, 167, 107740. [Google Scholar] [CrossRef]
- Brakatselos, C.; Delis, F.; Asprogerakas, M.Z.; Lekkas, P.; Tseti, I.; Tzimas, P.S.; Petrakis, E.A.; Halabalaki, M.; Skaltsounis, L.A.; Antoniou, K. Cannabidiol Modulates the Motor Profile and NMDA Receptor-related Alterations Induced by Ketamine. Neuroscience 2021, 454, 105–115. [Google Scholar] [CrossRef] [PubMed]
- Sartim, A.G.; Marques, J.; Silveira, K.M.; Gobira, P.H.; Guimarães, F.S.; Wegener, G.; Joca, S.R. Co-administration of cannabidiol and ketamine induces antidepressant-like effects devoid of hyperlocomotor side-effects. Neuropharmacology 2021, 195, 108679. [Google Scholar] [CrossRef]
- Alzeer, J.; Hadeed, K.A.; Basar, H.; Al-Razem, F.; Abdel-Wahhab, M.A.; Alhamdan, Y. Cannabis and Its Permissibility Status. Cannabis Cannabinoid Res. 2021, 6, 451–456. [Google Scholar] [CrossRef]
- Kaplan, J.S.; Stella, N.; Catterall, W.A.; Westenbroek, R.E. Cannabidiol attenuates seizures and social deficits in a mouse model of Dravet syndrome. Proc. Natl. Acad. Sci. USA 2017, 114, 11229–11234. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Rimmerman, N.; Ben-Hail, D.; Porat, Z.; Juknat, A.; Kozela, E.; Daniels, M.P.; Connelly, P.S.; Leishman, E.; Bradshaw, H.B.; Shoshan-Barmatz, V.; et al. Direct modulation of the outer mitochondrial membrane channel, voltage-dependent anion channel 1 (VDAC1) by cannabidiol: A novel mechanism for cannabinoid-induced cell death. Cell Death Dis. 2013, 4, e949. [Google Scholar] [CrossRef] [PubMed]
- Ericson, E.; Samuelsson, J.; Ahlenius, S. Photocell measurements of rat motor activity. A contribution to sensitivity and variation in behavioral observations. J. Pharmacol. Methods. 1991, 25, 111–122. [Google Scholar] [CrossRef]
- Peng, J.; Fan, M.; An, C.; Ni, F.; Huang, W.; Luo, J. A narrative review of molecular mechanism and therapeutic effect of cannabidiol (CBD). Basic Clin. Pharmacol. Toxicol. 2022, 130, 439–456. [Google Scholar] [CrossRef]
- Celorrio, M.; Rojo-Bustamante, E.; Fernández-Suárez, D.; Sáez, E.; Estella-Hermoso de Mendoza, A.; Müller, C.E.; Ramírez, M.J.; Oyarzábal, J.; Franco, R.; Aymerich, M.S. GPR55: A therapeutic target for Parkinson’s disease? Neuropharmacology 2017, 125, 319–332. [Google Scholar] [CrossRef]
- Davis, B.H.; Beasley, T.M.; Amaral, M.; Szaflarski, J.P.; Gaston, T.; Perry Grayson, L.; Standaert, D.G.; Bebin, E.M.; Limdi, N.A.; UAB CBD Study Group (includes all the investigators involved in the UAB EAP CBD program). Pharmacogenetic Predictors of Cannabidiol Response and Tolerability in Treatment-Resistant Epilepsy. Clin. Pharmacol Ther. 2021, 110, 1368–1380. [Google Scholar] [CrossRef]
- Shapiro, L.; Escayg, A.; Wong, J.C. Cannabidiol Increases Seizure Resistance and Improves Behavior in an Scn8a Mouse Model. Front. Pharmacol. 2022, 13, 815950. [Google Scholar] [CrossRef]
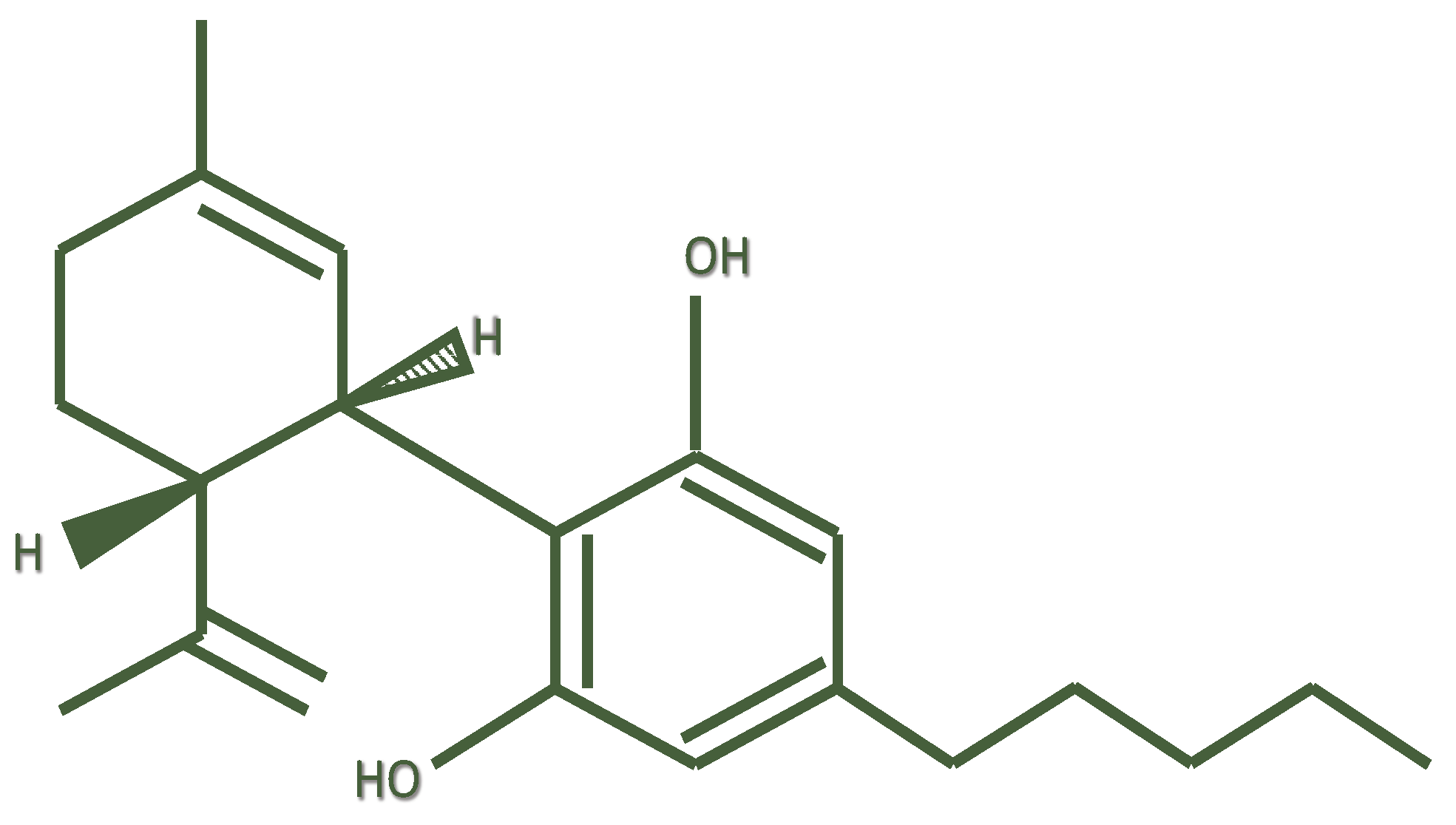
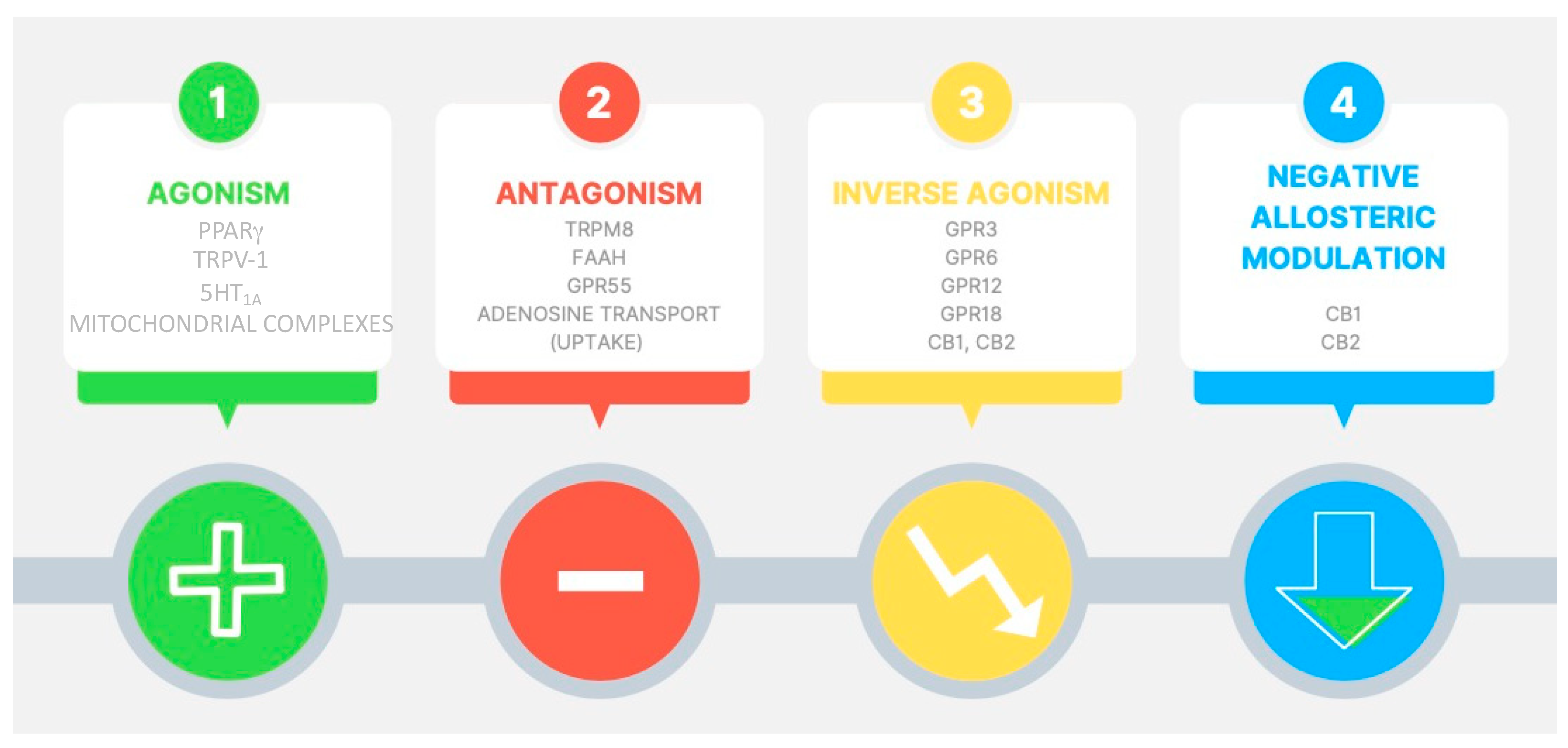
| Species | Experimental Model or Disease | CBD Administration | Locomotor Activity | Reference |
|---|---|---|---|---|
| Rat | Hypolocomotion associated with Parkinson’s disease induced by s.c. reserpine 1 mg/kg | Single i.p. 0.5 or 5 mg/kg or vehicle | Reduction of catalepsy and tardive dyskinesia | [47] |
| Mouse | Hepatic encephalopathy induced by bile-duct ligation | Daily i.p. 5 mg/kg or vehicle for four weeks | Reduction of hypolocomotion | [49] |
| Fmr1 knock out mouse | Fragile X syndrome | Single i.p. 5 or 20 mg/kg or vehicle | No effect on hyperlocomotion | [56] |
| APP/PS1 Mouse | Alzheimer disease | Daily i.p. 5 mg/kg or vehicle for 3 weeks before the beginning of experiments and during behavioral test assessment | No effect on locomotor activity | [59] |
| Mouse | Depressive and anxious-like behavior induced by LPS | Single i.p. 30 mg/kg or vehicle 30 min before LPS | No effect on changes in time spent in the center or distance travelled induced by LPS | [60] |
| Species | THC | CBD | Locomotor Activity | Reference |
|---|---|---|---|---|
| Rat | Single i.p. 10–30 mg/kg | Simultaneous single i.p. CBD 10–30 mg/kg with THC; i.p. CBD 10–30 mg/kg or 30–90 mg/kg, 30 min prior to THC | No effect on hypolocomotion induced by THC | [61] |
| Mouse | Single i.p. THC (0, 1.8, 3.2, 5.6 or 10 mg/kg) | Single i.p. CBD (30 mg/kg) 15 min before THC | CBD enhanced hypolocomotion induced by low to moderate THC doses | [63] |
| Single i.p. THC 1.8 mg/kg | Single i.p. CBD (0, 10 or 30 mg/kg) 13 h or 15 min before THC | CBD did not change significantly THC effects on locomotor activity. | ||
| Rat | Vaporized THC (12.5, 25, 50, 100, 200 mg/mL) 0.125 mL in a 40 min session | CBD (100, 400 mg/mL) 0.125 mL in a 40 min session | CBD alone when inhaled suppressed locomotor activity. THC and THC/CBD (1:4) inhalation decreased total locomotion in the center of the OF. | [64] |
| Monkey | Increasing i.m. THC dose (0.1–1 mg/kg for four months | Increasing i.m. CBD dose (0.3–3 mg/kg) for four months | Tolerance to hypolocomotion induced by THC appeared after weeks. CBD did not influence THC hypolocomotion or occurrence of tolerance. | [65] |
| Mouse | Single i.p. THC (10 mg/kg) | Simultaneous i.p. CBD (20 mg/kg) | CBD did not modify hypolocomotion induced by THC. | [67] |
| Mouse | Single i.p. THC (10 mg/kg) | Simultaneous single i.p. CBD (10 mg/kg) | CBD enhanced hypolocomotion induced by THC. | [68] |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Calapai, F.; Cardia, L.; Calapai, G.; Di Mauro, D.; Trimarchi, F.; Ammendolia, I.; Mannucci, C. Effects of Cannabidiol on Locomotor Activity. Life 2022, 12, 652. https://doi.org/10.3390/life12050652
Calapai F, Cardia L, Calapai G, Di Mauro D, Trimarchi F, Ammendolia I, Mannucci C. Effects of Cannabidiol on Locomotor Activity. Life. 2022; 12(5):652. https://doi.org/10.3390/life12050652
Chicago/Turabian StyleCalapai, Fabrizio, Luigi Cardia, Gioacchino Calapai, Debora Di Mauro, Fabio Trimarchi, Ilaria Ammendolia, and Carmen Mannucci. 2022. "Effects of Cannabidiol on Locomotor Activity" Life 12, no. 5: 652. https://doi.org/10.3390/life12050652
APA StyleCalapai, F., Cardia, L., Calapai, G., Di Mauro, D., Trimarchi, F., Ammendolia, I., & Mannucci, C. (2022). Effects of Cannabidiol on Locomotor Activity. Life, 12(5), 652. https://doi.org/10.3390/life12050652