
Early Salivary miRNA Expression in Extreme Low Gestational Age Newborns

 , ,
, , 
Abstract
1. Introduction
2. Materials and Methods
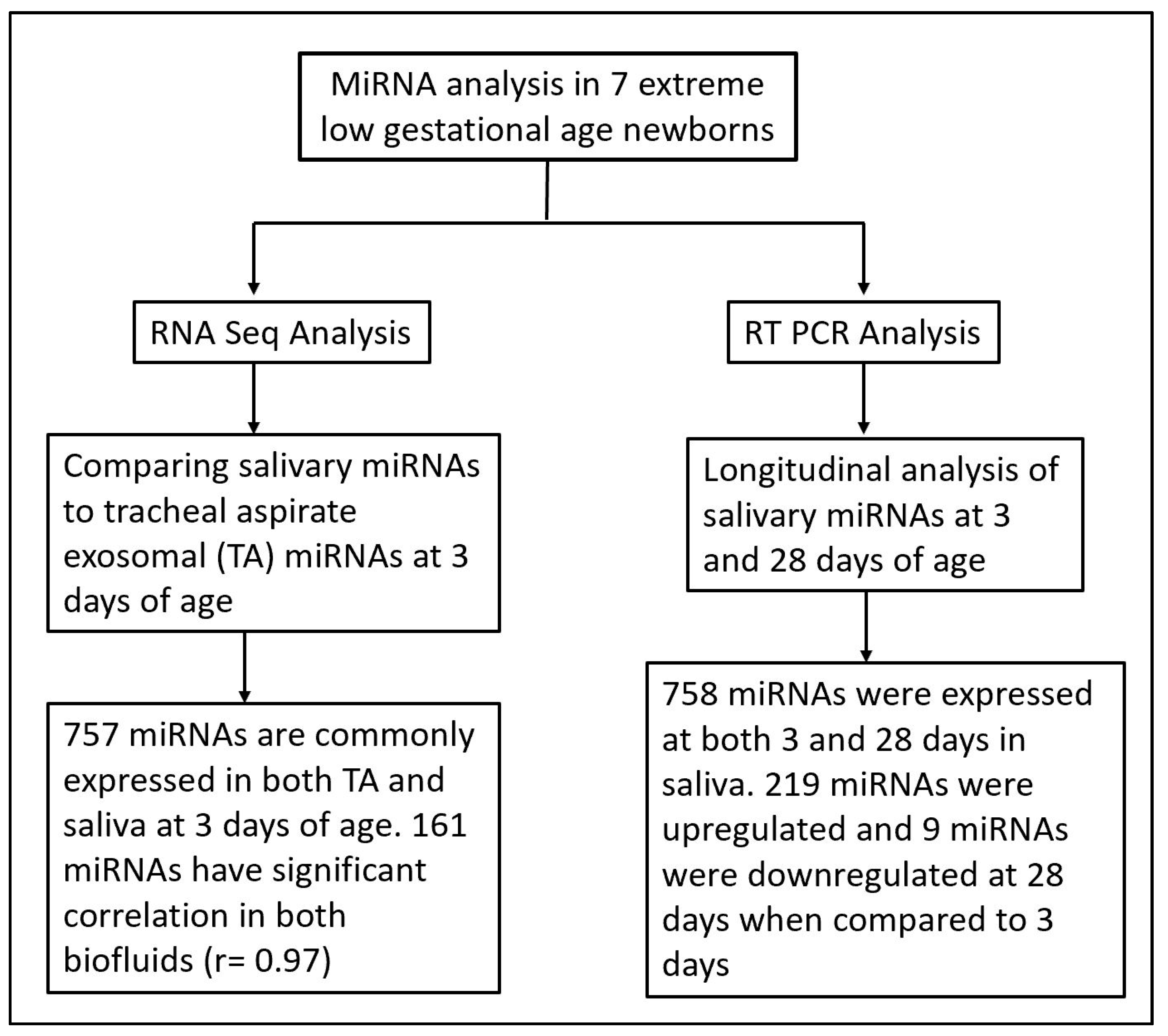
3. Results
3.1. Patient Demographics
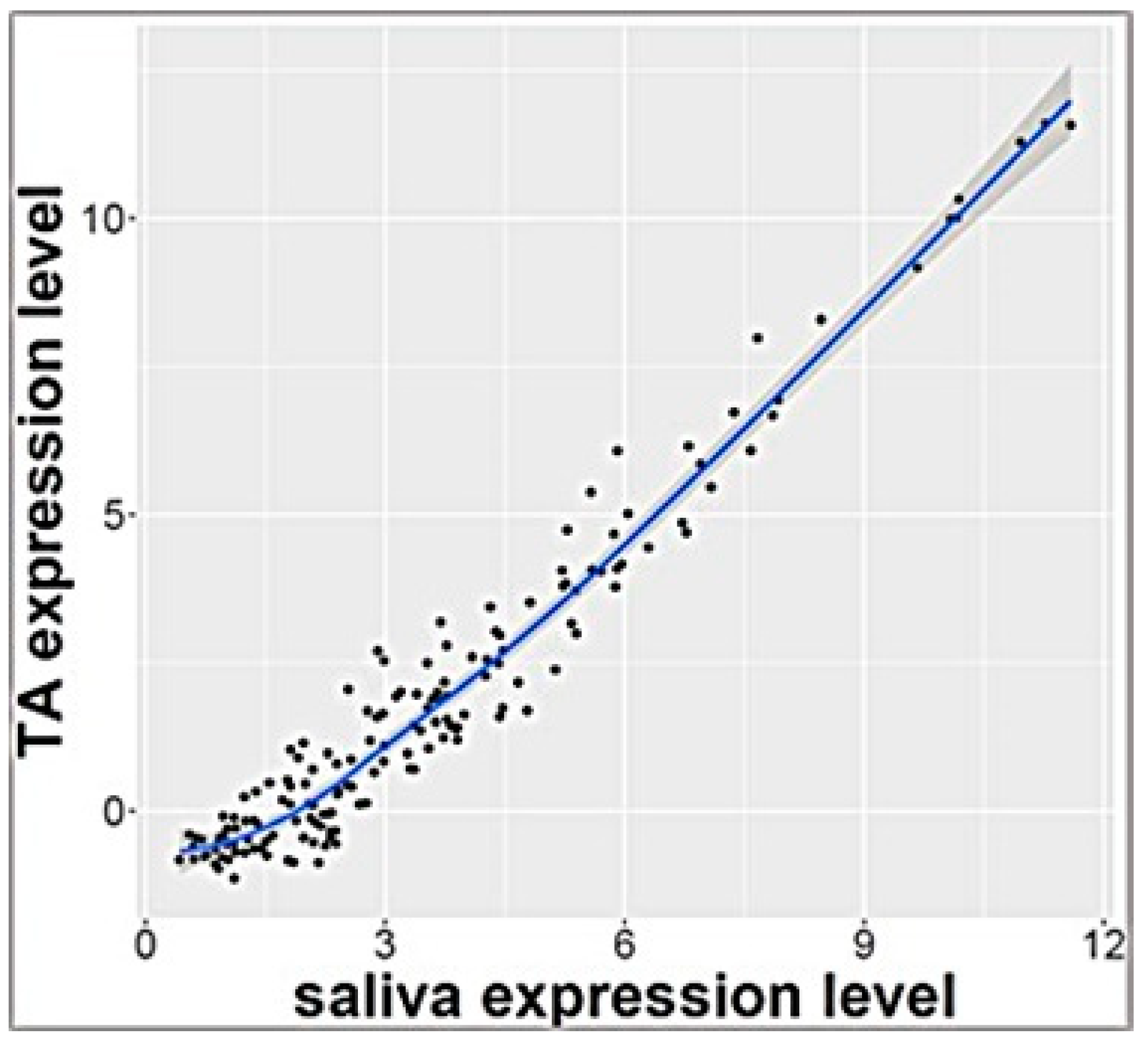
3.2. Salivary and Tracheal Aspirate miRNA Expression at 3 Days of Age
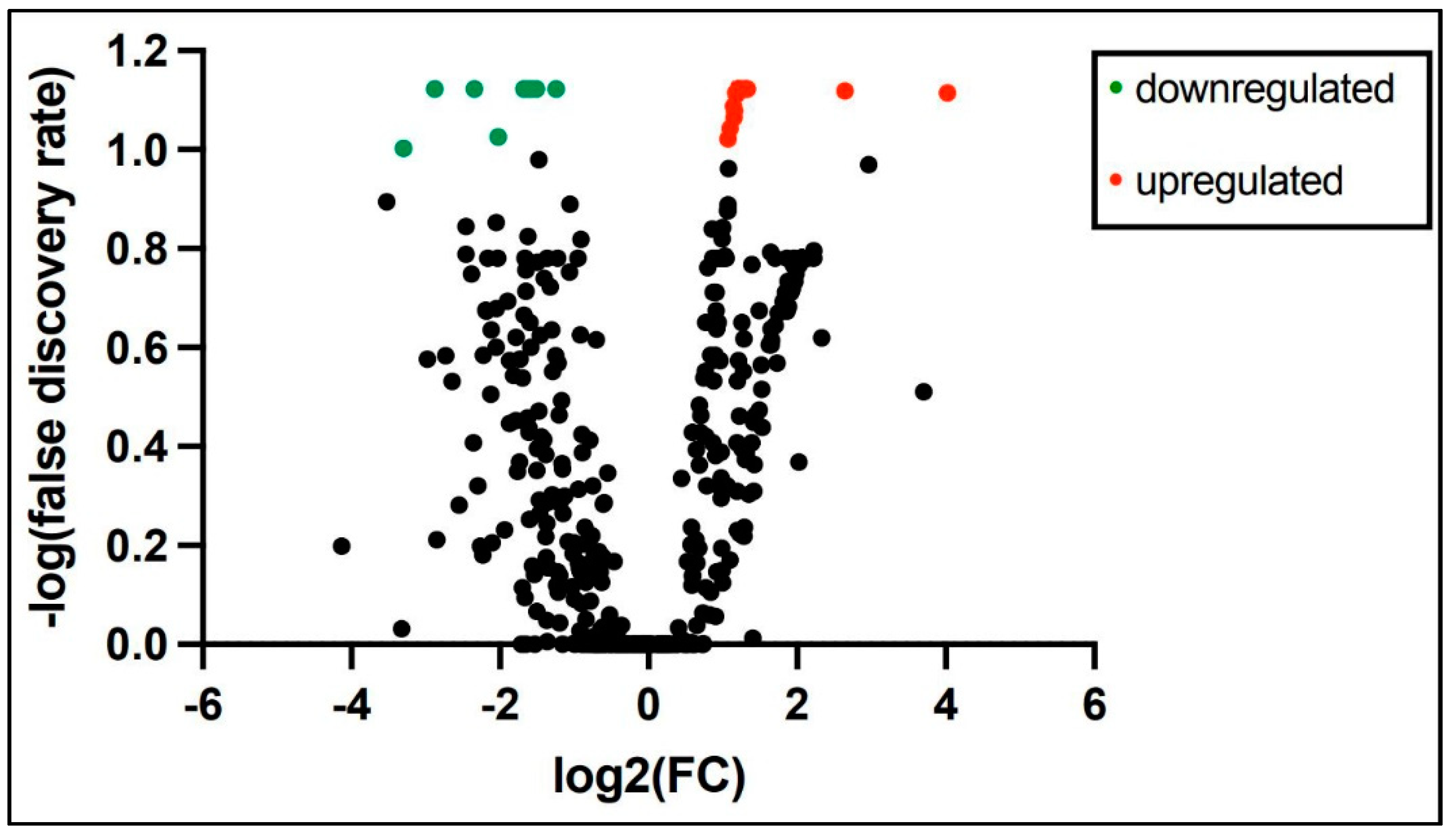
3.3. Longitudinal, Dynamic Salivary miRNA Expression at 3 and 28 Days of Age
3.4. Pathway Analysis of Differentially Expressed miRNAs in Saliva at 3 and 28 Days of Age
4. Discussion
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Mohr, A.M.; Mott, J.L. Overview of MicroRNA biology. Semin. Liver Dis. 2015, 35, 3–11. [Google Scholar] [CrossRef] [PubMed]
- Lal, C.V.; Olave, N.; Travers, C.; Rezonzew, G.; Dolma, K.; Simpson, A.; Halloran, B.; Aghai, Z.; Das, P.; Sharma, N.; et al. Exosomal microRNA predicts and protects against severe bronchopulmonary dysplasia in extremely premature infants. JCI Insight 2018, 3, e93994. [Google Scholar] [CrossRef] [PubMed]
- Schneider, A. Exosomes: Novel biomarkers for the clinical diagnosis of neurodegenerative diseases. J. Neural. Transm. 2019, 126, 646. [Google Scholar]
- Coalson, J.J. Pathology of bronchopulmonary dysplasia. Semin. Perinatol. 2006, 30, 179–184. [Google Scholar] [CrossRef] [PubMed]
- Lal, C.V.; Ambalavanan, N. Genetic predisposition to bronchopulmonary dysplasia. Semin. Perinatol. 2015, 39, 584–591. [Google Scholar] [CrossRef] [PubMed]
- Shah, D.; Sandhu, K.; Das, P.; Aghai, Z.H.; Andersson, S.; Pryhuber, G.; Bhandari, V. miR-184 mediates hyperoxia-induced injury by targeting cell death and angiogenesis signalling pathways in the developing lung (Retraction of 2020). Eur. Respir. J. 2021, 58, 1901789. [Google Scholar] [CrossRef]
- Oji-Mmuo, C.N.; Siddaiah, R.; Montes, D.T.; Pham, M.A.; Spear, D.; Donnelly, A.; Fuentes, N.; Imamura-Kawasawa, Y.; Howrylak, J.A.; Thomas, N.J.; et al. Tracheal aspirate transcriptomic and miRNA signatures of extreme premature birth with bronchopulmonary dysplasia. J. Perinatol. 2021, 41, 551–561. [Google Scholar] [CrossRef]
- Siddaiah, R.; Oji-Mmuo, C.N.; Montes, D.T.; Fuentes, N.; Spear, D.; Donnelly, A.; Silveyra, P. MicroRNA signatures associated with bronchopulmonary dysplasia severity in tracheal aspirates of preterm infants. Biomedicines 2021, 9, 257. [Google Scholar] [CrossRef]
- Kaczor-Urbanowicz, K.E.; Carreras-Presas, C.M.; Aro, K.; Tu, M.; Garcia-Godoy, F.; Wong, D.T.W. Saliva diagnostics—Current views and directions. Exp. Biol. Med. 2017, 242, 459–472. [Google Scholar] [CrossRef]
- Segal, A.; Wong, D.T. Salivary diagnostics: Enhancing disease detection and making medicine better. Eur. J. Dent. Educ. 2008, 12, 22–29. [Google Scholar]
- Martina, E.; Campanati, A.; Diotallevi, F.; Offidani, A. Saliva and oral diseases. J. Clin. Med. 2020, 9, 466. [Google Scholar] [CrossRef] [PubMed]
- Sripad, S. Salivaomics in systemic disease: An evolving science- an interesting prospect for ’lab-on-chip’ diagnostics in coronary artery and other systemic diseases. Indian J. Thorac. Cardiovasc. Surg. 2018, 34, 95–97. [Google Scholar] [CrossRef]
- Maron, J.L. Exploring the neonatal salivary transcriptome: Technical optimization and clinical applications. Clin. Biochem. 2011, 44, 467–468. [Google Scholar] [CrossRef] [PubMed]
- Romano-Keeler, J.; Wynn, J.L.; Maron, J.L. Great expectorations: The potential of salivary ‘omic’ approaches in neonatal intensive care. J. Perinatol. 2014, 34, 169–173. [Google Scholar] [CrossRef] [PubMed][Green Version]
- Hicks, S.D.; Carpenter, R.L.; Wagner, K.E.; Pauley, R.; Barros, M.; Tierney-Aves, C.; Barns, S.; Greene, C.D.; Middleton, F.A. Saliva microRNA differentiates children with autism from peers with typical and atypical development. J. Am. Acad. Child Adolesc. Psychiatry 2020, 59, 296–308. [Google Scholar] [CrossRef]
- Hicks, S.D.; Onks, C.; Kim, R.Y.; Zhen, K.J.; Loeffert, J.; Loeffert, A.C.; Olympia, R.P.; Fedorchak, G.; DeVita, S.; Rangnekar, A.; et al. Diagnosing mild traumatic brain injury using saliva RNA compared to cognitive and balance testing. Clin. Transl. Med. 2020, 10, e197. [Google Scholar] [CrossRef]
- Weaver, K.R.; Melkus, G.D.; Fletcher, J.; Henderson, W.A. Relevance of sex and subtype in patients with IBS: An exploratory study of gene expression. Biol. Res. Nurs. 2020, 22, 13–23. [Google Scholar] [CrossRef]
- Wang, L.; Feng, Z.; Wang, X.; Wang, X.; Zhang, X. DEGseq: An R package for identifying differentially expressed genes from RNA-seq data. Bioinformatics 2010, 26, 136–138. [Google Scholar] [CrossRef]
- Reiner, A.; Yekutieli, D.; Benjamini, Y. Identifying differentially expressed genes using false discovery rate controlling procedures. Bioinformatics 2003, 19, 368–375. [Google Scholar] [CrossRef]
- Abrahamsson, A.; Dabrosin, C. Tissue specific expression of extracellular microRNA in human breast cancers and normal human breast tissue in vivo. Oncotarget 2015, 6, 22959–22969. [Google Scholar] [CrossRef]
- Ambroise, J.; Badir, J.; Nienhaus, L.; Robert, A.; Dekairelle, A.F.; Gala, J.L. Using a constraint-based regression method for relative quantification of somatic mutations in pyrosequencing signals: A case for NRAS analysis. Algorithms Mol. Biol. 2016, 11, 24. [Google Scholar] [CrossRef] [PubMed]
- D’Haene, B.; Mestdagh, P.; Hellemans, J.; Vandesompele, J. miRNA expression profiling: From reference genes to global mean normalization. Methods Mol. Biol. 2012, 822, 261–272. [Google Scholar]
- Zhao, Y.; Srivastava, D. A developmental view of microRNA function. Trends Biochem. Sci. 2007, 32, 189–197. [Google Scholar] [CrossRef] [PubMed]
- Wang, J.; Yin, J.; Wang, X.; Liu, H.; Hu, Y.; Yan, X.; Zhuang, B.; Yu, Z.; Han, S. Changing expression profiles of mRNA, lncRNA, circRNA, and miRNA in lung tissue reveal the pathophysiological of bronchopulmonary dysplasia (BPD) in mouse model. J. Cell Biochem. 2019, 120, 9369–9380. [Google Scholar] [CrossRef] [PubMed]
- Yang, Y.; Qiu, J.; Kan, Q.; Zhou, X.G.; Zhou, X.Y. MicroRNA expression profiling studies on bronchopulmonary dysplasia: A systematic review and meta-analysis. Genet. Mol. Res. 2013, 12, 5195–5206. [Google Scholar] [CrossRef]
- Naeem, A.; Ahmed, I.; Silveyra, P. Bronchopulmonary dysplasia: An update on experimental therapeutics. Eur. Med. J. 2019, 4, 20–29. [Google Scholar]
- Rivera, L.; Siddaiah, R.; Oji-Mmuo, C.; Silveyra, G.R.; Silveyra, P. Biomarkers for bronchopulmonary dysplasia in the preterm infant. Front. Pediatr. 2016, 4, 33. [Google Scholar] [CrossRef]
- Zhou, Y.; Liu, S.; Wang, W.; Sun, Q.; Lv, M.; Yang, S.; Tong, S.; Guo, S. The miR-204-5p/FOXC1/GDF7 axis regulates the osteogenic differentiation of human adipose-derived stem cells via the AKT and p38 signalling pathways. Stem Cell Res. Ther. 2021, 12, 64. [Google Scholar] [CrossRef]
- Yao, S.; Yin, Y.; Jin, G.; Li, D.; Li, M.; Hu, Y.; Feng, Y.; Liu, Y.; Bian, Z.; Wang, X.; et al. Exosome-mediated delivery of miR-204-5p inhibits tumor growth and chemoresistance. Cancer Med. 2020, 9, 5989–5998. [Google Scholar] [CrossRef]
- Jung, H.M.; Hu, C.T.; Fister, A.M.; Davis, A.E.; Castranova, D.; Pham, V.N.; Price, L.M.; Weinstein, B.M. MicroRNA-mediated control of developmental lymphangiogenesis. Elife 2019, 8, e46007. [Google Scholar] [CrossRef]
- Zhu, J.; Liu, B.; Wang, Z.; Wang, D.; Ni, H.; Zhang, L.; Wang, Y. Exosomes from nicotine-stimulated macrophages accelerate atherosclerosis through miR-21-3p/PTEN-mediated VSMC migration and proliferation. Theranostics 2019, 9, 6901–6919. [Google Scholar] [CrossRef] [PubMed]
- Xu, N.; He, D.; Shao, Y.; Qu, Y.; Ye, K.; Memet, O.; Zhang, L.; Shen, J. Lung-derived exosomes in phosgene-induced acute lung injury regulate the functions of mesenchymal stem cells partially via miR-28-5p. Biomed. Pharmacother. 2020, 121, 109603. [Google Scholar] [CrossRef] [PubMed]
- Abdallah, F.; Henriet, E.; Suet, A.; Arar, A.; Clemençon, R.; Malinge, J.M.; Lecellier, G.; Baril, P.; Pichon, C. miR-21-3p/IL-22 axes are major drivers of psoriasis pathogenesis by modulating keratinocytes proliferation-survival balance and inflammatory response. Cells 2021, 10, 2547. [Google Scholar] [CrossRef]
- Zhao, G.; Wang, T.; Huang, Q.K.; Pu, M.; Sun, W.; Zhang, Z.C.; Ling, R.; Tao, K.S. MicroRNA-548a-5p promotes proliferation and inhibits apoptosis in hepatocellular carcinoma cells by targeting Tg737. World J. Gastroenterol. 2016, 22, 5364–5373. [Google Scholar] [CrossRef] [PubMed]
- He, W.; Pang, L.; Gong, S.; Wang, X.; Hou, L. Nei Endonuclease VIII-like 2 gene rs8191670 polymorphism affects the sensitivity of non-small cell lung cancer to cisplatin by binding with MiR-548a. J. Cancer 2020, 11, 4801–4809. [Google Scholar] [CrossRef]
- Lou, G.; Chen, L.; Xia, C.; Wang, W.; Qi, J.; Li, A.; Zhao, L.; Chen, Z.; Zheng, M.; Liu, Y. MiR-199a-modified exosomes from adipose tissue-derived mesenchymal stem cells improve hepatocellular carcinoma chemosensitivity through mTOR pathway. J. Exp. Clin. Cancer Res. 2020, 39, 4. [Google Scholar] [CrossRef]
- Zuo, Y.; Shen, W.; Wang, C.; Niu, N.; Pu, J. Circular RNA circ-ZNF609 promotes lung adenocarcinoma proliferation by modulating miR-1224-3p/ETV1 signaling. Cancer Manag. Res. 2020, 12, 2471–2479. [Google Scholar] [CrossRef]
- Zhang, Y.; Dai, J.; Deng, H.; Wan, H.; Liu, M.; Wang, J.; Li, S.; Li, X.; Tang, H. miR-1228 promotes the proliferation and metastasis of hepatoma cells through a p53 forward feedback loop. Br. J. Cancer 2015, 112, 365–374. [Google Scholar] [CrossRef]




{kind=link}
{kind=link}
{kind=link}
| Patient Demographics | |
|---|---|
| Gestational Age | 25 4/7 (23 5/7–28 3/7) |
| Birth weight in grams | 617 (38–1010) |
| Gender | M:F: 3:4 |
| Race: n | White: 3 Other: 3 Asian: 1 |
| AGA: SGA | 3:4 |
| Mode of delivery C-section: Vaginal birth | 6:1 |
| miRNAs | Fold Change | Adjusted p Value | Expression Intensity |
|---|---|---|---|
| has-miR-204 | 0.885151 | 0.080909 | 9254.185 |
| hashsa-miR-21-3p | −1.22323 | 0.063979 | 6147.197 |
| hsa-miR-28-5p | −0.67488 | 0.027228 | 1980.699 |
| hsa-miR-218-2-3p | −0.46345 | 0.055106 | 1350.412 |
| hsa-miR-3 has5p | −1.03647 | 0.003002 | 1326.227 |
| hsa-mihas0 e-5p | −1.52103 | 0.013793 | 1310.715 |
| hsa-has-3679-5p | −1.01302 | 0.001325 | 1219.705 |
| has-miR-363-3p | 0.840106 | 0.001739 | 1219.248 |
| hsa-miR-3153 | 0.52835 | 0.010092 | 1030.863 |
| hsa-miR-449 b-5p | −1.15254 | 0.000161 | 974.736 |
| miRNAs | Fold Change | Adjusted p Value | Up Ohasown Regulated |
|---|---|---|---|
| hsa-miR-548a | 16.236 | 0.076833 | Up-regulated |
| hsa-miR-199a | 6.245 | 0.076064 | Up-regulated |
| hsa-miR-1224-3P | 2.513 | 0.07549 | Up-regulated |
| hsa-miR-1288 | 2.482 | 0.07549 | Up-regulated |
| hsa-miR-337-3p | 2.474 | 0.07549 | Up-regulated |
| hsa-miR-182 | 2.437 | 0.07549 | Up-regulated |
| hsa-miR-554 | 2.347 | 0.07549 | Up-regulated |
| hsa-let-7c | 2.333 | 0.07549 | Up-regulated |
| hsa-let-7a | 2.32 | 0.07549 | Up-regulated |
| hsa-let-7b | 2.32 | 0.07549 | Up-regulated |
| hsa-let-7e | 2.32 | 0.07549 | Up-regulated |
| Top Molecular and Cellular Functions | p Value Range |
| Cellular Development | 4.83 × 10−2–2.39 × 10−8 |
| Cellular Movement | 3.77 × 10−2–2.74 × 10−7 |
| Cellular Growth and Proliferation | 4.83 × 10−2–9.35 × 10−7 |
| Cell Cycle | 4.83 × 10−2–1.24 × 10−5 |
| Cell Death and Survival | 4.72 × 10−2–1.06 × 10−4 |
| Top Physiological System Development and Function | p Value Range |
| Organismal development | 4.58 × 10−2–3.96 × 10−6 |
| Digestive System Development and Function | 2.17 × 10−4–2.17 × 10−4 |
| Hepatic System Development and Function | 2.17 × 10−4–2.17 × 10−4 |
| Organ Development | 2.73 × 10−2–2.17 × 10−4 |
| Embryonic Development | 1.96 × 10−2–8.60 × 10−4 |
| Top Molecular and Cellular Functions | p Value Range |
| Cellular Movement | 3.55 × 10−2–5.78 × 10−11 |
| Cellular Development | 4.90 × 10−2–7.99 × 10−11 |
| Cellular growth and Proliferation | 4.92 × 10−2–7.99 × 10−11 |
| Cell Death and Survival | 4.92 × 10−2–5.06 × 10−7 |
| Top Physiological System Development and Function | p Value Range |
| Embryonic Development | 4.19 × 10−2–1.97 × 10−6 |
| Organismal Development | 4.50 × 10−2–2.38 × 10−6 |
| Digestive System Development and Function | 4.79 × 10−6–4.79 × 10−6 |
| Hepatic System Development and Function | 4.79 × 10−6–4.79 × 10−6 |
| Organ Development | 4.19 × 10−2–4.79 × 10−6 |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Siddaiah, R.; Emery, L.; Stephens, H.; Donnelly, A.; Erkinger, J.; Wisecup, K.; Hicks, S.D.; Kawasawa, Y.I.; Oji-Mmuo, C.; Amatya, S.; et al. Early Salivary miRNA Expression in Extreme Low Gestational Age Newborns. Life 2022, 12, 506. https://doi.org/10.3390/life12040506
Siddaiah R, Emery L, Stephens H, Donnelly A, Erkinger J, Wisecup K, Hicks SD, Kawasawa YI, Oji-Mmuo C, Amatya S, et al. Early Salivary miRNA Expression in Extreme Low Gestational Age Newborns. Life. 2022; 12(4):506. https://doi.org/10.3390/life12040506
Chicago/Turabian StyleSiddaiah, Roopa, Lucy Emery, Heather Stephens, Ann Donnelly, Jennifer Erkinger, Kimberly Wisecup, Steven D. Hicks, Yuka Imamura Kawasawa, Christiana Oji-Mmuo, Shaili Amatya, and et al. 2022. "Early Salivary miRNA Expression in Extreme Low Gestational Age Newborns" Life 12, no. 4: 506. https://doi.org/10.3390/life12040506
APA StyleSiddaiah, R., Emery, L., Stephens, H., Donnelly, A., Erkinger, J., Wisecup, K., Hicks, S. D., Kawasawa, Y. I., Oji-Mmuo, C., Amatya, S., & Silveyra, P. (2022). Early Salivary miRNA Expression in Extreme Low Gestational Age Newborns. Life, 12(4), 506. https://doi.org/10.3390/life12040506