
Persicaria minor (Huds.) Opiz Prevents In Vitro Atherogenesis by Attenuating Tumor Necrosis Factor-α-Induced Monocyte Adhesion to Human Umbilical Vein Endothelial Cells

 ,
,  , , and
, , and {kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
1. Introduction
2. Materials and Methods
2.1. Standardized Aqueous Extract of P. minor Leaves Preparation
2.2. HUVEC Isolation and Culture
2.3. Determination of Cytotoxicity and Optimal Dose of PM on HUVEC Viability by 3-(4,5-Dimethylthiazol-2-YI)-2,5-Diphenyltetrazolium Bromide (MTT) Assay
2.4. HUVEC Treatment Protocol
2.5. Measurement of ICAM-1 and VCAM-1 mRNA Expressions Using Quantitative Real-Time Polymerase Chain Reaction (qPCR)
2.6. Measurement of Protein Levels of ICAM-I and VCAM-1 in HUVEC Using Enzyme-Linked Immunosorbent Assay (ELISA)
2.7. Measurement of Monocyte Adhesion to HUVEC
2.8. Measurement of NF-kB p65 Levels in HUVEC
2.9. Data Analysis
3. Results
3.1. PM Improved the Viability of HUVEC Exposed to TNF-α
3.2. Effect of PM on ICAM-1 and VCAM-1 mRNA Expression and Protein Levels in HUVEC
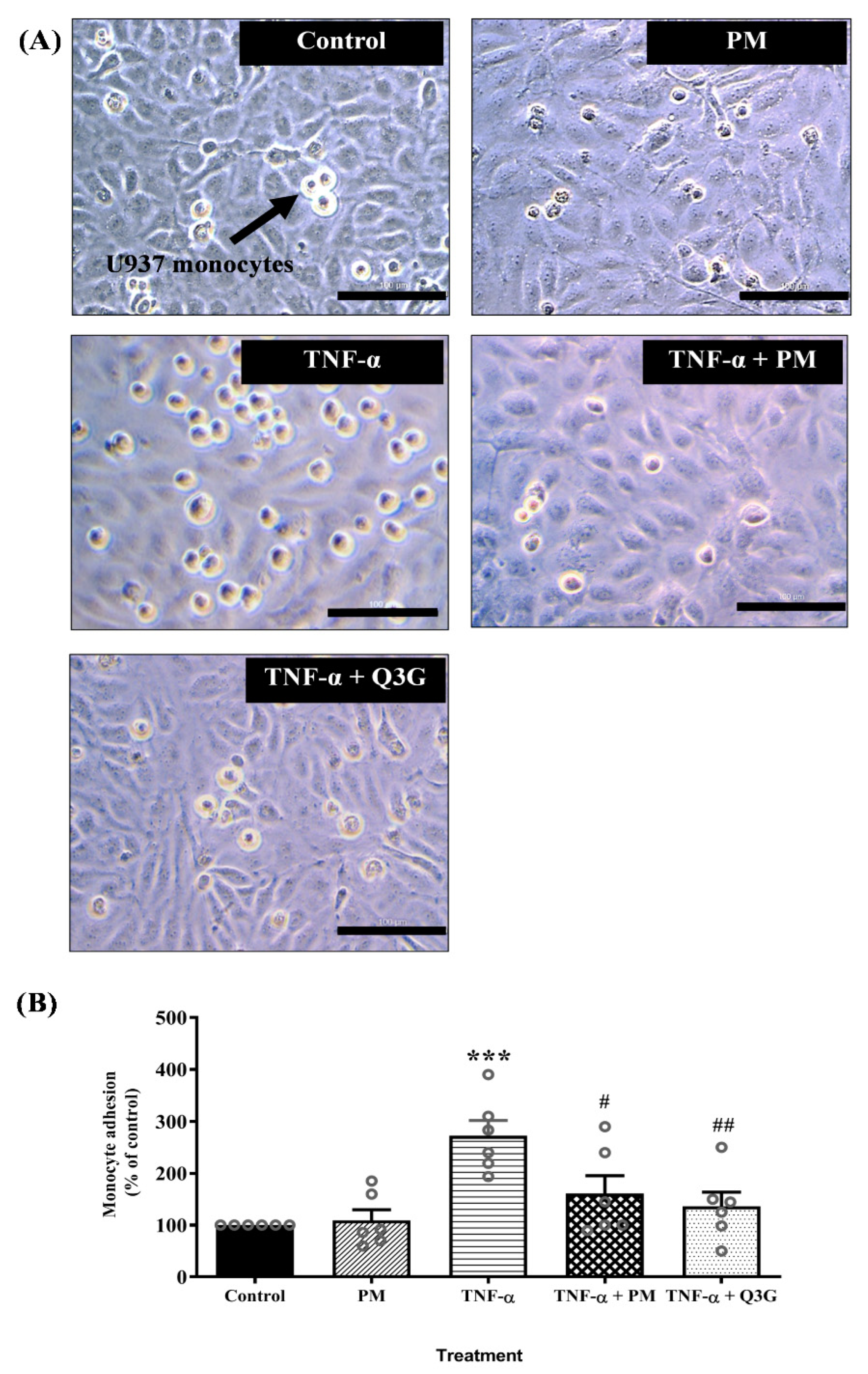
3.3. Effect of PM on U937 Monocyte Adhesion to HUVEC
3.4. Effect of PM on NF-κB p65 Level in HUVEC Nuclear Extract
4. Discussion
5. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Soehnlein, O.; Libby, P. Targeting Inflammation in Atherosclerosis—from Experimental Insights to the Clinic. Nat. Rev. Drug Discov. 2021, 20, 589–610. [Google Scholar] [CrossRef] [PubMed]
- Davignon, J.; Ganz, P. Role of Endothelial Dysfunction in Atherosclerosis. Circulation 2004, 109 (Suppl. S1), 23. [Google Scholar] [CrossRef] [PubMed]
- Čejková, S.; Králová-Lesná, I.; Poledne, R. Monocyte Adhesion to the Endothelium Is an Initial Stage of Atherosclerosis Development. Cor Vasa 2016, 58, e419–e425. [Google Scholar] [CrossRef]
- Botts, S.R.; Fish, J.E.; Howe, K.L. Dysfunctional Vascular Endothelium as a Driver of Atherosclerosis: Emerging Insights Into Pathogenesis and Treatment. Front. Pharmacol. 2021, 12, 787541. [Google Scholar] [CrossRef]
- Al Shahi, H.; Shimada, K.; Miyauchi, K.; Yoshihara, T.; Sai, E.; Shiozawa, T.; Naito, R.; Aikawa, T.; Ouchi, S.; Kadoguchi, T.; et al. Elevated Circulating Levels of Inflammatory Markers in Patients with Acute Coronary Syndrome. Int. J. Vasc. Med. 2015, 2015, 805375. [Google Scholar] [CrossRef]
- Ohta, H.; Wada, H.; Niwa, T.; Kirii, H.; Iwamoto, N.; Fujii, H.; Saito, K.; Sekikawa, K.; Seishima, M. Disruption of Tumor Necrosis Factor-α Gene Diminishes the Development of Atherosclerosis in ApoE-Deficient Mice. Atherosclerosis 2005, 180, 11–17. [Google Scholar] [CrossRef]
- Galkina, E.; Ley, K. Vascular Adhesion Molecules in Atherosclerosis. Arterioscler. Thromb. Vasc. Biol. 2007, 27, 2292–2301. [Google Scholar] [CrossRef]
- Habas, K.; Shang, L. Alterations in Intercellular Adhesion Molecule 1 (ICAM-1) and Vascular Cell Adhesion Molecule 1 (VCAM-1) in Human Endothelial Cells. Tissue Cell 2018, 54, 139–143. [Google Scholar] [CrossRef]
- Chen, X.; Li, H.; Wang, Z.; Zhou, Q.; Chen, S.; Yang, B.; Yin, D.; He, H.; He, M. Quercetin Protects the Vascular Endothelium against Iron Overload Damages via ROS/ADMA/DDAHⅡ/ENOS/NO Pathway. Eur. J. Pharmacol. 2020, 868, 172885. [Google Scholar] [CrossRef]
- Toma, L.; Sanda, G.M.; Niculescu, L.S.; Deleanu, M.; Sima, A.V.; Stancu, C.S. Phenolic Compounds Exerting Lipid-Regulatory, Anti-Inflammatory and Epigenetic Effects as Complementary Treatments in Cardiovascular Diseases. Biomolecules 2020, 10, 641. [Google Scholar] [CrossRef]
- Chen, C.; Luo, F.; Liu, X.; Lu, L.; Xu, H.; Yang, Q.; Xue, J.; Shi, L.; Li, J.; Zhang, A.; et al. NF-KB-Regulated Exosomal MiR-155 Promotes the Inflammation Associated with Arsenite Carcinogenesis. Cancer Lett. 2017, 388, 21–33. [Google Scholar] [CrossRef] [PubMed]
- Zinatizadeh, M.R.; Schock, B.; Chalbatani, G.M.; Zarandi, P.K.; Jalali, S.A.; Miri, S.R. The Nuclear Factor Kappa B (NF-KB) Signaling in Cancer Development and Immune Diseases. Genes Dis. 2021, 8, 287–297. [Google Scholar] [CrossRef] [PubMed]
- Burkill, I.H. A Dictionary of the Economic Products of the Malay Peninsula. Nature 1936, 137, 255. [Google Scholar] [CrossRef]
- Christapher, P.V.; Parasuraman, S.; Asmawi, M.Z.; Murugaiyah, V. Acute and Subchronic Toxicity Studies of Methanol Extract of Polygonum Minus Leaves in Sprague Dawley Rats. Regul. Toxicol. Pharmacol. 2017, 86, 33–41. [Google Scholar] [CrossRef] [PubMed]
- Christapher, P.; Vikneswaran, M.; Xin, T.; Yuan, G.; Parasuraman, S.; Leng, L.; Kiun, C.; Fu, N. Evaluation of Analgesic, Anti-Inflammatory, Antipyretic and Antiulcer Effect of Aqueous and Methanol Extracts of Leaves of Polygonum Minus Huds. (Polygonaceae) in Rodents. Arch. Med. Health Sci. 2015, 3, 12. [Google Scholar] [CrossRef]
- Abdullah, M.Z.; Mohd Ali, J.; Abolmaesoomi, M.; Abdul-Rahman, P.S.; Hashim, O.H. Anti-Proliferative, in Vitro Antioxidant, and Cellular Antioxidant Activities of the Leaf Extracts from Polygonum Minus Huds: Effects of Solvent Polarity. Int. J. Food Prop. 2017, 20, S846–S862. [Google Scholar] [CrossRef]
- Hamid, A.A.; Aminuddin, A.; Yunus, M.H.M.; Murthy, J.K.; Hui, C.K.; Ugusman, A. Antioxidative and Anti-Inflammatory Activities of Polygonum Minus: A Review of Literature. Rev. Cardiovasc. Med. 2020, 21, 275–288. [Google Scholar] [CrossRef]
- Sumazian, Y.; Syahida, A.; Hakiman, M.; Maziah, M. Antioxidant Activities, Flavonoids, Ascorbic Acid and Phenolic Contents of Malaysian Vegetables. J. Med. Plants Res. 2010, 4, 881–890. [Google Scholar] [CrossRef]
- Maizura, M.; Aminah, A.; Aida, W.M.W. Total Phenolic Content and Antioxidant Activity of Kesum (Polygonum Minus), Ginger (Zingiber Officinale) and Turmeric (Curcuma Longa) Extract. Int. Food Res. J. 2011, 18, 526–531. [Google Scholar]
- Wan Yahaya, W.A.; Abu Yazid, N.; Mohd Azman, N.A.; Almajano, M.P. Antioxidant Activities and Total Phenolic Content of Malaysian Herbs as Components of Active Packaging Film in Beef Patties. Antioxidants 2019, 8, 204. [Google Scholar] [CrossRef]
- Nurul, H.; Ruzita, A.; Aronal, A.P. The Antioxidant Effects of Cosmos Caudatus and Polygonum Minus in Refrigerated Duck Meatballs. Int. Food Res. J. 2010, 17, 893–904. [Google Scholar]
- Saputri, F.C.; Jantan, I. Effects of Selected Medicinal Plants on Human Low-Density Lipoprotein Oxidation, 2, 2-Diphenyl-1-Picrylhydrazyl (DPPH) Radicals and Human Platelet Aggregation. J. Med. Plant Res. 2011, 5, 6182–6191. [Google Scholar] [CrossRef]
- George, A.; Chinnappan, S.; Chintamaneni, M.; Kotak, C.V.; Choudhary, Y.; Kueper, T.; Radhakrishnan, A.K. Anti-Inflammatory Effects of Polygonum Minus (Huds) Extract (LineminusTM) in in-Vitro Enzyme Assays and Carrageenan Induced Paw Edema. BMC Complement. Altern. Med. 2014, 14, 355. [Google Scholar] [CrossRef]
- Kusumaningrum, P.A.; Yustinasari, L.R.; Sahrial, I.; Hamid; Sudjarwo, S.A.; Kuncoro Puguh Santoso, C.A. Effect of Polygonum Minus (KESUM) leaves ethanolic extract on histopathological changes on the wall aorta of mice (Mus Musculus) induced by cadmium chloride antioxidant. J. Basic Med. Vet. 2019, 8, 66–75. [Google Scholar] [CrossRef]
- George, A.; Ng, C.P.; O’Callaghan, M.; Jensen, G.S.; Wong, H.J. In Vitro and Ex-Vivo Cellular Antioxidant Protection and Cognitive Enhancing Effects of an Extract of Polygonum Minus Huds (LineminusTM) Demonstrated in a Barnes Maze Animal Model for Memory and Learning. BMC Complement. Altern. Med. 2014, 14, 161. [Google Scholar] [CrossRef]
- Sundar, U.M.; Ugusman, A.; Chua, H.K.; Latip, J.; Aminuddin, A. Piper Sarmentosum Promotes Endothelial Nitric Oxide Production by Reducing Asymmetric Dimethylarginine in Tumor Necrosis Factor-α-Induced Human Umbilical Vein Endothelial Cells. Front. Pharmacol. 2019, 10, 1033. [Google Scholar] [CrossRef]
- Ugusman, A.; Zakaria, Z.; Hui, C.K.; Megat Mohd Nordin, N.A. Piper Sarmentosum Inhibits ICAM-1 and Nox4 Gene Expression in Oxidative Stress-Induced Human Umbilical Vein Endothelial Cells. BMC Complement. Altern. Med. 2011, 11, 31. [Google Scholar] [CrossRef]
- Bradford, M.M. A Rapid and Sensitive Method for the Quantitation of Microgram Quantities of Protein Utilizing the Principle of Protein-Dye Binding. Anal. Biochem. 1976, 72, 248–254. [Google Scholar] [CrossRef]
- Chang, C.C.; Chu, C.F.; Wang, C.N.; Wu, H.T.; Bi, K.W.; Pang, J.H.S.; Huang, S.T. The Anti-Atherosclerotic Effect of Tanshinone IIA Is Associated with the Inhibition of TNF-α-Induced VCAM-1, ICAM-1 and CX3CL1 Expression. Phytomedicine 2014, 21, 207–216. [Google Scholar] [CrossRef]
- Ismail, S.M.; Sundar, U.M.; Hui, C.K.; Aminuddin, A.; Ugusman, A. Piper Sarmentosum Attenuates TNF-α-Induced VCAM-1 and ICAM-1 Expression in Human Umbilical Vein Endothelial Cells. J. Taibah Univ. Med. Sci. 2018, 13, 225–231. [Google Scholar] [CrossRef]
- Cook-Mills, J.M.; Marchese, M.E.; Abdala-Valencia, H. Vascular Cell Adhesion Molecule-1 Expression and Signaling during Disease: Regulation by Reactive Oxygen Species and Antioxidants. Antioxid. Redox Signal. 2011, 15, 1607–1638. [Google Scholar] [CrossRef] [PubMed]
- Maguire, E.M.; Pearce, S.W.A.; Xiao, Q. Foam Cell Formation: A New Target for Fighting Atherosclerosis and Cardiovascular Disease. Vascul. Pharmacol. 2019, 112, 54–71. [Google Scholar] [CrossRef] [PubMed]
- Tribolo, S.; Lodi, F.; Connor, C.; Suri, S.; Wilson, V.G.; Taylor, M.A.; Needs, P.W.; Kroon, P.A.; Hughes, D.A. Comparative Effects of Quercetin and Its Predominant Human Metabolites on Adhesion Molecule Expression in Activated Human Vascular Endothelial Cells. Atherosclerosis 2008, 197, 50–56. [Google Scholar] [CrossRef] [PubMed]
- Mochizuki, M.; Kajiya, K.; Terao, J.; Kaji, K.; Kumazawa, S.; Nakayama, T.; Shimoi, K. Effect of Quercetin Conjugates on Vascular Permeability and Expression of Adhesion Molecules. BioFactors 2004, 22, 201–204. [Google Scholar] [CrossRef]
- Winterbone, M.S.; Tribolo, S.; Needs, P.W.; Kroon, P.A.; Hughes, D.A. Physiologically Relevant Metabolites of Quercetin Have No Effect on Adhesion Molecule or Chemokine Expression in Human Vascular Smooth Muscle Cells. Atherosclerosis 2009, 202, 431–438. [Google Scholar] [CrossRef]
- Krüger-Genge, A.; Blocki, A.; Franke, R.P.; Jung, F. Vascular Endothelial Cell Biology: An Update. Int. J. Mol. Sci. 2019, 20, 4411. [Google Scholar] [CrossRef]
- Bhatt, D.; Ghosh, S. Regulation of the NF-ΚB-Mediated Transcription of Inflammatory Genes. Front. Immunol. 2014, 5, 71. [Google Scholar] [CrossRef]
- Milstone, D.S.; Ilyama, M.; Chen, M.; O’Donnell, P.; Davis, V.M.; Plutzky, J.; Brown, J.D.; Haldar, S.M.; Siu, A.; Lau, A.C.; et al. Differential Role of an NF-ΚB Transcriptional Response Element in Endothelial versus Intimal Cell VCAM-1 Expression. Circ. Res. 2015, 117, 166–177. [Google Scholar] [CrossRef]
- Ernst, O.; Vayttaden, S.J.; Fraser, I.D.C. Measurement of NF-ΚB Activation in TLR-Activated Macrophages. Methods Mol. Biol. 2018, 1714, 67–78. [Google Scholar] [CrossRef]
- Guo, X.D.; Zhang, D.Y.; Gao, X.J.; Parry, J.; Liu, K.; Liu, B.L.; Wang, M. Quercetin and Quercetin-3-O-Glucuronide Are Equally Effective in Ameliorating Endothelial Insulin Resistance through Inhibition of Reactive Oxygen Species-Associated Inflammation. Mol. Nutr. Food Res. 2013, 57, 1037–1045. [Google Scholar] [CrossRef]
- Chen, T.; Zhang, X.; Zhu, G.; Liu, H.; Chen, J.; Wang, Y.; He, X. Quercetin Inhibits TNF-α Induced HUVECs Apoptosis and Inflammation via Downregulating NF-KB and AP-1 Signaling Pathway in Vitro. Medicine 2020, 99, e22241. [Google Scholar] [CrossRef] [PubMed]





Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Hamid, A.A.; Aminuddin, A.; Anuar, N.N.M.; Mansor, N.I.; Ahmad, M.F.; Saleh, M.S.M.; Mokhtar, M.H.; Ugusman, A. Persicaria minor (Huds.) Opiz Prevents In Vitro Atherogenesis by Attenuating Tumor Necrosis Factor-α-Induced Monocyte Adhesion to Human Umbilical Vein Endothelial Cells. Life 2022, 12, 1462. https://doi.org/10.3390/life12101462
Hamid AA, Aminuddin A, Anuar NNM, Mansor NI, Ahmad MF, Saleh MSM, Mokhtar MH, Ugusman A. Persicaria minor (Huds.) Opiz Prevents In Vitro Atherogenesis by Attenuating Tumor Necrosis Factor-α-Induced Monocyte Adhesion to Human Umbilical Vein Endothelial Cells. Life. 2022; 12(10):1462. https://doi.org/10.3390/life12101462
Chicago/Turabian StyleHamid, Adila A., Amilia Aminuddin, Nur Najmi Mohamad Anuar, Nur Izzati Mansor, Mohd Faizal Ahmad, Mohammed S. M. Saleh, Mohd Helmy Mokhtar, and Azizah Ugusman. 2022. "Persicaria minor (Huds.) Opiz Prevents In Vitro Atherogenesis by Attenuating Tumor Necrosis Factor-α-Induced Monocyte Adhesion to Human Umbilical Vein Endothelial Cells" Life 12, no. 10: 1462. https://doi.org/10.3390/life12101462
APA StyleHamid, A. A., Aminuddin, A., Anuar, N. N. M., Mansor, N. I., Ahmad, M. F., Saleh, M. S. M., Mokhtar, M. H., & Ugusman, A. (2022). Persicaria minor (Huds.) Opiz Prevents In Vitro Atherogenesis by Attenuating Tumor Necrosis Factor-α-Induced Monocyte Adhesion to Human Umbilical Vein Endothelial Cells. Life, 12(10), 1462. https://doi.org/10.3390/life12101462