
Idiopathic Infertility as a Feature of Genome Instability


{kind=link}
Abstract
1. Introduction
2. Methodology
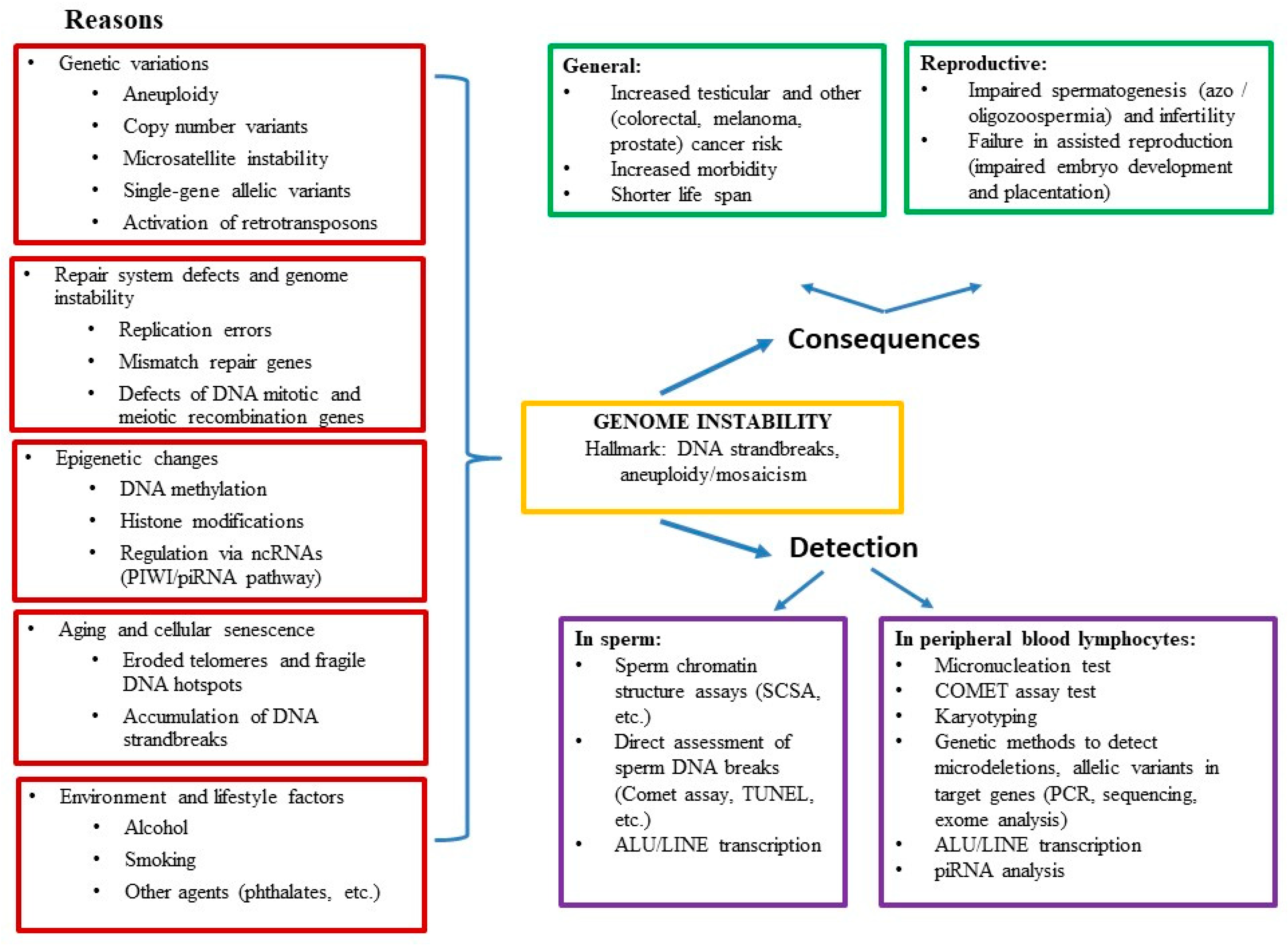
3. Reasons and Consequences of Genome Instability
3.1. Genetic Variations
3.1.1. Aneuploidy
3.1.2. Copy Number Variants (CNVs)
3.1.3. Microsatellite Instability (MSI)
3.1.4. Single-Gene Allelic Variants
3.1.5. Activation of Retrotransposons
3.2. Repair System Defects, Genome Instability and Male Infertility
3.2.1. Replication Errors
3.2.2. Mismatch Repair Genes
3.2.3. Defects of DNA Mitotic and Meiotic Repair Genes
3.3. Epigenetic Changes and Male Infertility
3.3.1. DNA Methylation
3.3.2. Histone Modifications
3.3.3. PIWI/piRNA Pathway
3.4. Aging and Cellular Senescence
3.5. Environment and Lifestyle Factors and Their Influence on the Genome and Spermatogenesis
4. Assessment of Genome Instability
4.1. Assessment of Genome Instability in Sperm
4.2. Evaluation of ALU/LINE Activity
4.3. Assessment of Genome Instability in Peripheral Blood Leukocytes
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Acknowledgments
Conflicts of Interest
References
- Nieschlag, E. Scope and Goals of Andrology. In Andrology: Male Reproductive Health and Dysfunction; Nieschlag, E., Behre, H.M., Eds.; Springer: Berlin/Heidelberg, Germany, 2001; pp. 1–8. [Google Scholar] [CrossRef]
- Punab, M.; Poolamets, O.; Paju, P.; Vihljajev, V.; Pomm, K.; Ladva, R.; Korrovits, P.; Laan, M. Causes of male infertility: A 9-year prospective monocentre study on 1737 patients with reduced total sperm counts. Hum. Reprod. 2016, 32, 18–31. [Google Scholar] [CrossRef]
- Carlsen, E.; Giwercman, A.; Keiding, N.; Skakkebaek, N.E. Evidence for decreasing quality of semen during past 50 years. BMJ 1992, 305, 609–613. [Google Scholar] [CrossRef] [PubMed]
- Sengupta, P.; Borges, E.; Dutta, S.; Krajewska-Kułak, E. Decline in sperm count in European men during the past 50 years. Hum. Exp. Toxicol. 2017, 37, 247–255. [Google Scholar] [CrossRef]
- Guzick, D.S.; Overstreet, J.W.; Factor-Litvak, P.; Brazil, C.K.; Nakajima, S.T.; Coutifaris, C.; Carson, S.A.; Cisneros, P.; Steinkampf, M.P.; Hill, J.A.; et al. Sperm Morphology, Motility, and Concentration in Fertile and Infertile Men. New Engl. J. Med. 2001, 345, 1388–1393. [Google Scholar] [CrossRef]
- Cao, X.W.; Lin, K.; Li, C.Y.; Yuan, C.W. A review of WHO Laboratory Manual for the Examination and Processing of Human Semen (5th edition)]. Zhonghua Nan Ke Xue 2011, 17, 1059–1063. [Google Scholar] [PubMed]
- van den Hoven, L.; Hendriks, J.C.; Verbeet, J.G.; Westphal, J.R.; Wetzels, A.M. Status of sperm morphology assessment: An evaluation of methodology and clinical value. Fertil. Steril. 2015, 103, 53–58. [Google Scholar] [CrossRef] [PubMed]
- O’Neill, C.L.; Parrella, A.; Keating, D.; Cheung, S.; Rosenwaks, Z.; Palermo, G.D. A treatment algorithm for couples with unexplained infertility based on sperm chromatin assessment. J. Assist. Reprod. Genet. 2018, 35, 1911–1917. [Google Scholar] [CrossRef]
- Evenson, D.P.; Jost, L.K.; Marshall, D.; Zinaman, M.J.; Clegg, E.; Purvis, K.; De Angelis, P.; Claussen, O.P. Utility of the sperm chromatin structure assay as a diagnostic and prognostic tool in the human fertility clinic. Hum. Reprod. 1999, 14, 1039–1049. [Google Scholar] [CrossRef]
- Bungum, M.; Humaidan, P.; Axmon, A.; Spano, M.; Bungum, L.; Erenpreiss, J.; Giwercman, A. Sperm DNA integrity assessment in prediction of assisted reproduction technology outcome. Hum. Reprod. 2007, 22, 174–179. [Google Scholar] [CrossRef]
- Erenpreiss, J.; Je, E.; Giwercman, A.; Tsarev, I.; Erenpreisa, J.; Spano, M. Toluidine blue cytometry test for sperm DNA conformation: Comparison with the flow cytometric sperm chromatin structure and TUNEL assays. Hum. Reprod. 2004, 19, 2277–2282. [Google Scholar] [CrossRef] [PubMed]
- Tsarev, I.; Bungum, M.; Giwercman, A.; Erenpreisa, J.; Ebessen, T.; Ernst, E.; Erenpreiss, J. Evaluation of male fertility potential by Toluidine Blue test for sperm chromatin structure assessment. Hum. Reprod. 2009, 24, 1569–1574. [Google Scholar] [CrossRef]
- Kim, H.-S.; Kang, M.J.; Kim, S.A.; Oh, S.K.; Kim, H.; Ku, S.-Y.; Kim, S.H.; Moon, S.Y.; Choi, Y.M. The utility of sperm DNA damage assay using toluidine blue and aniline blue staining in routine semen analysis. Clin. Exp. Reprod. Med. 2013, 40, 23–28. [Google Scholar] [CrossRef]
- Erenpreiss, J.; Bars, J.; Lipatnikova, V.; Erenpreisa, J.; Zalkalns, J. Comparative study of cytochemical tests for sperm chromatin integrity. J. Androl. 2001, 22, 45–53. [Google Scholar] [PubMed]
- Erenpreiss, J.; Spano, M.; Bungum, M.; Giwercman, A. Sperm chromatin structure and male fertility: Biological and clinical aspects. Asian J. Androl. 2006, 8, 11–29. [Google Scholar] [CrossRef]
- Osman, A.; Alsomait, H.; Seshadri, S.; El-Toukhy, T.; Khalaf, Y. The effect of sperm DNA fragmentation on live birth rate after IVF or ICSI: A systematic review and meta-analysis. Reprod. Biomed. Online 2015, 30, 120–127. [Google Scholar] [CrossRef] [PubMed]
- Pourmasumi, S.; Khoradmehr, A.; Rahiminia, T.; Sabeti, P.; Talebi, A.R.; Ghasemzadeh, J. Evaluation of Sperm Chromatin Integrity Using Aniline Blue and Toluidine Blue Staining in Infertile and Normozoospermic Men. J. Reprod. Infertil. 2019, 20, 95–101. [Google Scholar]
- Borini, A.; Tarozzi, N.; Bizzaro, D.; Bonu, M.; Fava, L.; Flamigni, C.; Coticchio, G. Sperm DNA fragmentation: Paternal effect on early post-implantation embryo development in ART. Hum. Reprod. 2006, 21, 2876–2881. [Google Scholar] [CrossRef]
- Collins, J.A.; Barnhart, K.T.; Schlegel, P.N. Do sperm DNA integrity tests predict pregnancy with in vitro fertilization? Fertil. Steril. 2008, 89, 823–831. [Google Scholar] [CrossRef] [PubMed]
- Lewis, S.E.; Aitken, R.J.; Conner, S.J.; De Iuliis, G.; Evenson, D.P.; Henkel, R.; Giwercman, A.; Gharagozloo, P. The impact of sperm DNA damage in assisted conception and beyond: Recent advances in diagnosis and treatment. Reprod. Biomed. Online 2013, 27, 325–337. [Google Scholar] [CrossRef]
- Aguilera, A.; Gómez-González, B. Genome instability: A mechanistic view of its causes and consequences. Nat. Rev. Genet. 2008, 9, 204–217. [Google Scholar] [CrossRef] [PubMed]
- Vijg, J.; Montagna, C. Genome instability and aging: Cause or effect? Transl. Med. Aging 2017, 1, 5–11. [Google Scholar] [CrossRef]
- Iourov, I.Y.; Vorsanova, S.G.; Yurov, Y.B.; Kutsev, S.I. Ontogenetic and Pathogenetic Views on Somatic Chromosomal Mosaicism. Genes 2019, 10, 379. [Google Scholar] [CrossRef] [PubMed]
- Jensen, T.K.; Jacobsen, R.; Christensen, K.; Nielsen, N.C.; Bostofte, E. Good Semen Quality and Life Expectancy: A Cohort Study of 43,277 Men. Am. J. Epidemiol. 2009, 170, 559–565. [Google Scholar] [CrossRef] [PubMed]
- Salonia, A.; Matloob, R.; Gallina, A.; Abdollah, F.; Saccà, A.; Briganti, A.; Suardi, N.; Colombo, R.; Rocchini, L.; Guazzoni, G.F.; et al. Are Infertile Men Less Healthy than Fertile Men? Results of a Prospective Case-Control Survey. Eur. Urol. 2009, 56, 1025–1032. [Google Scholar] [CrossRef] [PubMed]
- Aston, K.I.; Carrell, D.T. Emerging evidence for the role of genomic instability in male factor infertility. Syst. Biol. Reprod. Med. 2011, 58, 71–80. [Google Scholar] [CrossRef] [PubMed]
- Cheung, S.; Parrella, A.; Rosenwaks, Z.; Palermo, G.D. Genetic and epigenetic profiling of the infertile male. PLoS ONE 2019, 14, e0214275. [Google Scholar] [CrossRef]
- Morris, J.K.; Alberman, E.; Scott, C.; Jacobs, P. Is the prevalence of Klinefelter syndrome increasing? Eur. J. Hum. Genet. 2007, 16, 163–170. [Google Scholar] [CrossRef]
- Roux, C.; Tripogney, C.; Morel, F.; Joanne, C.; Fellmann, F.; Clavequin, M.; Bresson, J. Segregation of chromosomes in sperm of Robertsonian translocation carriers. Cytogenet. Genome Res. 2005, 111, 291–296. [Google Scholar] [CrossRef]
- Benet, J.; Oliver-Bonet, M.; Cifuentes, P.; Templado, C.; Navarro, J. Segregation of chromosomes in sperm of reciprocal translocation carriers: A review. Cytogenet. Genome Res. 2005, 111, 281–290. [Google Scholar] [CrossRef]
- Oud, M.S.; Volozonoka, L.; Smits, R.M.; Vissers, L.E.L.M.; Ramos, L.; A Veltman, J. A systematic review and standardized clinical validity assessment of male infertility genes. Hum. Reprod. 2019, 34, 932–941. [Google Scholar] [CrossRef] [PubMed]
- Jobling, M. Copy number variation on the human Y chromosome. Cytogenet. Genome Res. 2008, 123, 253–262. [Google Scholar] [CrossRef]
- Krausz, C.; Chianese, C.; Giachini, C.; Guarducci, E.; LaFace, I.; Forti, G. The Y chromosome-linked copy number variations and male fertility. J. Endocrinol. Investig. 2011, 34, 376–382. [Google Scholar] [CrossRef] [PubMed]
- Skaletsky, H.; Kuroda-Kawaguchi, T.; Minx, P.J.; Cordum, H.S.; Hillier, L.; Brown, L.G.; Repping, S.; Pyntikova, T.; Ali, J.; Bieri, T.; et al. The male-specific region of the human Y chromosome is a mosaic of discrete sequence classes. Nature 2003, 423, 825–837. [Google Scholar] [CrossRef]
- Jobling, M.A.; Tyler-Smith, C. The human Y chromosome: An evolutionary marker comes of age. Nat. Rev. Genet. 2003, 4, 598–612. [Google Scholar] [CrossRef]
- Vogt, P.H. Genomic heterogeneity and instability of the AZF locus on the human Y chromosome. Mol. Cell. Endocrinol. 2004, 224, 1–9. [Google Scholar] [CrossRef]
- Krausz, C. Y chromosome and male infertility: Update, 2006. Front. Biosci. 2006, 11, 3049–3061. [Google Scholar] [CrossRef] [PubMed]
- Krausz, C.; Forti, G.; McElreavey, K. The Y chromosome and male fertility and infertility1. Int. J. Androl. 2003, 26, 70–75. [Google Scholar] [CrossRef] [PubMed]
- Colaco, S.; Modi, D. Genetics of the human Y chromosome and its association with male infertility. Reprod. Biol. Endocrinol. 2018, 16, 1–24. [Google Scholar] [CrossRef]
- Vilar, E.; Gruber, S.B. Microsatellite instability in colorectal cancer—The stable evidence. Nat. Rev. Clin. Oncol. 2010, 7, 153–162. [Google Scholar] [CrossRef]
- Zheng, Y.-M.; Li, L.; Zhou, L.-M.; Le, F.; Cai, L.-Y.; Yu, P.; Zhu, Y.-R.; Liu, X.-Z.; Wang, L.-Y.; Li, L.-J.; et al. Alterations in the frequency of trinucleotide repeat dynamic mutations in offspring conceived through assisted reproductive technology. Hum. Reprod. 2013, 28, 2570–2580. [Google Scholar] [CrossRef]
- Erenpreiss, J.; Tsarev, I.; Giwercman, A.; Giwercman, Y. The impact of androgen receptor polymorphism and parental ethnicity on semen quality in young men from Latvia. Int. J. Androl. 2008, 31, 477–482. [Google Scholar] [CrossRef] [PubMed]
- Grigorova, M.; Punab, M.; Kahre, T.; Ivandi, M.; Tõnisson, N.; Poolamets, O.; Vihljajev, V.; Žilaitienė, B.; Erenpreiss, J.; Matulevičius, V.; et al. The number of CAG and GGN triplet repeats in the Androgen Receptor gene exert combinatorial effect on hormonal and sperm parameters in young men. Andrology 2017, 5, 495–504. [Google Scholar] [CrossRef]
- Bolton, E.C.; So, A.Y.; Chaivorapol, C.; Haqq, C.M.; Li, H.; Yamamoto, K.R. Cell- and gene-specific regulation of primary target genes by the androgen receptor. Genes Dev. 2007, 21, 2005–2017. [Google Scholar] [CrossRef]
- Weng, H.; Li, S.; Huang, J.-Y.; He, Z.-Q.; Meng, X.-Y.; Cao, Y.; Fang, C.; Zeng, X.-T. Androgen receptor gene polymorphisms and risk of prostate cancer: A meta-analysis. Sci. Rep. 2017, 7, 40554. [Google Scholar] [CrossRef] [PubMed]
- Yatsenko, A.N.; Georgiadis, A.P.; Röpke, A.; Berman, A.J.; Jaffe, T.; Olszewska, M.; Westernströer, B.; Sanfilippo, J.; Kurpisz, M.; Rajkovic, A.; et al. X-Linked TEX11 Mutations, Meiotic Arrest, and Azoospermia in Infertile Men. N. Engl. J. Med. 2015, 372, 2097–2107. [Google Scholar] [CrossRef] [PubMed]
- Bracke, A.; Peeters, K.; Punjabi, U.; Hoogewijs, D.; Dewilde, S. A search for molecular mechanisms underlying male idiopathic infertility. Reprod. Biomed. Online 2018, 36, 327–339. [Google Scholar] [CrossRef]
- Wang, Z.; Pan, Y.; He, L.; Song, X.; Chen, H.; Pan, C.; Qu, L.; Zhu, H.; Lan, X. Multiple morphological abnormalities of the sperm flagella (MMAF)-associated genes: The relationships between genetic variation and litter size in goats. Gene 2020, 753, 144778. [Google Scholar] [CrossRef] [PubMed]
- Solyom, S.; Ewing, A.D.; Hancks, D.C.; Takeshima, Y.; Awano, H.; Matsuo, M.; Jr, H.H.K. Pathogenic orphan transduction created by a nonreference LINE-1 retrotransposon. Hum. Mutat. 2012, 33, 369–371. [Google Scholar] [CrossRef]
- Pace, J.K.; Feschotte, C. The evolutionary history of human DNA transposons: Evidence for intense activity in the primate lineage. Genome Res. 2007, 17, 422–432. [Google Scholar] [CrossRef]
- Smit, A.F.; Riggs, A. Tiggers and DNA transposon fossils in the human genome. Proc. Natl. Acad. Sci. USA 1996, 93, 1443–1448. [Google Scholar] [CrossRef] [PubMed]
- Hedges, D.; Deininger, P. Inviting instability: Transposable elements, double-strand breaks, and the maintenance of genome integrity. Mutat. Res. Mol. Mech. Mutagen. 2007, 616, 46–59. [Google Scholar] [CrossRef] [PubMed]
- Bourque, G.; Burns, K.H.; Gehring, M.; Gorbunova, V.; Seluanov, A.; Hammell, M.; Imbeault, M.; Izsvák, Z.; Levin, H.L.; Macfarlan, T.S.; et al. Ten things you should know about transposable elements. Genome Biol. 2018, 19, 1–12. [Google Scholar] [CrossRef]
- Kumar, A. Jump around: Transposons in and out of the laboratory. F1000Research 2020, 9, 135. [Google Scholar] [CrossRef] [PubMed]
- Belancio, V.P.; Deininger, P.L.; Roy-Engel, A.M. LINE dancing in the human genome: Transposable elements and disease. Genome Med. 2009, 1, 97–98. [Google Scholar] [CrossRef]
- Ernst, C.; Odom, D.T.; Kutter, C. The emergence of piRNAs against transposon invasion to preserve mammalian genome integrity. Nat. Commun. 2017, 8, 1–10. [Google Scholar] [CrossRef]
- Cioppi, F.; Casamonti, E.; Krausz, C. Age-Dependent De Novo Mutations During Spermatogenesis and Their Consequences. Adv. Exp. Med. Biol. 2019, 1166, 29–46. [Google Scholar] [CrossRef] [PubMed]
- Gunes, S.; Sertyel, S. Sperm DNA Damage and Oocyte Repair Capability BT. In A Clinician’s Guide to Sperm DNA and Chromatin Damage; Zini, A., Agarwal, A., Eds.; Springer International Publishing: Berlin/Heidelberg, Germany, 2018; pp. 321–346. [Google Scholar] [CrossRef]
- Agarwal, A.; Barbăroșie, C.; Ambar, R.; Finelli, R. The Impact of Single- and Double-Strand DNA Breaks in Human Spermatozoa on Assisted Reproduction. Int. J. Mol. Sci. 2020, 21, 3882. [Google Scholar] [CrossRef]
- Mukherjee, S.; Ridgeway, A.D.; Lamb, D.J. DNA mismatch repair and infertility. Curr. Opin. Urol. 2010, 20, 525–532. [Google Scholar] [CrossRef]
- Gunes, S.; Agarwal, A.; Henkel, R.; Mahmutoglu, A.M.; Sharma, R.; Esteves, S.C.; Aljowair, A.; Emirzeoglu, D.; Alkhani, A.; Pelegrini, L.; et al. Association between promoter methylation of MLH1 and MSH2 and reactive oxygen species in oligozoospermic men-A pilot study. Andrology 2018, 50, e12903. [Google Scholar] [CrossRef]
- Zhao, X.; Mu, C.; Ma, J.; Dai, X.; Jiao, H. The association of four SNPs in DNA mismatch repair genes with idiopathic male infertility in northwest China. Int. J. Immunogenet. 2019, 46, 451–458. [Google Scholar] [CrossRef]
- Lynch, H.T.; De La Chapelle, A. Hereditary Colorectal Cancer. N. Engl. J. Med. 2003, 348, 919–932. [Google Scholar] [CrossRef]
- Pommier, Y.; Barcelo, J.M.; Rao, V.A.; Sordet, O.; Jobson, A.G.; Thibaut, L.; Miao, Z.; Seiler, J.A.; Zhang, H.; Marchand, C.; et al. Repair of Topoisomerase I-Mediated DNA Damage. Prog. Nucleic Acid Res. Mol. Biol. 2006, 81, 179–229. [Google Scholar] [CrossRef] [PubMed]
- Gómez-Herreros, F. DNA Double Strand Breaks and Chromosomal Translocations Induced by DNA Topoisomerase II. Front. Mol. Biosci. 2019, 6, 141. [Google Scholar] [CrossRef]
- Mavragani, I.V.; Nikitaki, Z.; Kalospyros, S.A.; Georgakilas, A.G. Ionizing Radiation and Complex DNA Damage: From Prediction to Detection Challenges and Biological Significance. Cancers 2019, 11, 1789. [Google Scholar] [CrossRef]
- Helena Mangs, A.; Morris, B.J. The Human Pseudoautosomal Region (PAR): Origin, Function and Future. Curr. Genom. 2007, 8, 129–136. [Google Scholar] [CrossRef]
- Acquaviva, L.; Boekhout, M.; Karasu, M.E.; Brick, K.; Pratto, F.; Li, T.; van Overbeek, M.; Kauppi, L.; Camerini-Otero, R.D.; Jasin, M.; et al. Ensuring meiotic DNA break formation in the mouse pseudoautosomal region. Nat. Cell Biol. 2020, 582, 426–431. [Google Scholar] [CrossRef]
- Zhang, B.; Tang, Z.; Li, L.; Lu, L.-Y. NBS1 is required for SPO11-linked DNA double-strand break repair in male meiosis. Cell Death Differ. 2020, 27, 2176–2190. [Google Scholar] [CrossRef]
- Ren, Z.-J.; Ren, P.-W.; Yang, B.; Liao, J.; Liu, S.-Z.; Fang, K.; Ren, S.-Q.; Liu, L.-R.; Dong, Q. The SPO11-C631T gene polymorphism and male infertility risk: A meta-analysis. Ren. Fail. 2017, 39, 299–305. [Google Scholar] [CrossRef]
- Tran, T.N.; Schimenti, J.C. A segregating human allele of SPO11 modeled in mice disrupts timing and amounts of meiotic recombination, causing oligospermia and a decreased ovarian reserve†. Biol. Reprod. 2019, 101, 347–359. [Google Scholar] [CrossRef] [PubMed]
- Ribas-Maynou, J.; García-Peiró, A.; Fernandez-Encinas, A.; Amengual, M.J.; Prada, E.; Cortés, P.; Navarro, J.; Benet, J. Double Stranded Sperm DNA Breaks, Measured by Comet Assay, Are Associated with Unexplained Recurrent Miscarriage in Couples without a Female Factor. PLoS ONE 2012, 7, e44679. [Google Scholar] [CrossRef]
- Zeng, Y.; Chen, T. DNA Methylation Reprogramming during Mammalian Development. Genes 2019, 10, 257. [Google Scholar] [CrossRef] [PubMed]
- Vanneste, E.; Voet, T.; Le Caignec, C.; Ampe, M.; Konings, P.; Melotte, C.; Debrock, S.; Amyere, M.; Vikkula, M.; Schuit, F.; et al. Chromosome instability is common in human cleavage-stage embryos. Nat. Med. 2009, 15, 577–583. [Google Scholar] [CrossRef]
- Cavé, T.; Desmarais, R.; Lacombe-Burgoyne, C.; Boissonneault, G. Genetic Instability and Chromatin Remodeling in Spermatids. Genes 2019, 10, 40. [Google Scholar] [CrossRef]
- Rathke, C.; Baarends, W.M.; Awe, S.; Renkawitz-Pohl, R. Chromatin dynamics during spermiogenesis. Biochim. Biophys. Acta (BBA) Bioenerg. 2014, 1839, 155–168. [Google Scholar] [CrossRef] [PubMed]
- Martiyanov, I.; Brancorsini, S.; Catena, R.; Gansmuller, A.; Kotaja, N.; Parvinen, M.; Sassone-Corsi, P.; Davidson, I. Polar nuclear localization of H1T2, a histone H1 variant, required for spermatid elongation and DNA condensation during spermiogenesis. Proc. Natl. Acad. Sci. USA 2005, 102, 2808–2813. [Google Scholar] [CrossRef] [PubMed]
- Hempfling, A.L.; Lim, S.L.; Adelson, D.L.; Evans, J.; E O’Connor, A.; Qu, Z.P.; Kliesch, S.; Weidner, W.; O’Bryan, M.K.; Bergmann, M. Expression patterns of HENMT1 and PIWIL1 in human testis: Implications for transposon expression. Reprod. 2017, 154, 363–374. [Google Scholar] [CrossRef]
- Jodar, M.; Selvaraju, S.; Sendler, E.; Diamond, M.; Krawetz, S.A. The presence, role and clinical use of spermatozoal RNAs. Hum. Reprod. Updat. 2013, 19, 604–624. [Google Scholar] [CrossRef] [PubMed]
- Ghasemi, H.; Khodadadi, I.; Fattahi, A.; Moghimbeigi, A.; Tavilani, H. Polymorphisms of DNA repair genes XRCC1 and LIG4 and idiopathic male infertility. Syst. Biol. Reprod. Med. 2017, 63, 382–390. [Google Scholar] [CrossRef]
- McHugh, D.; Gil, J. Senescence and aging: Causes, consequences, and therapeutic avenues. J. Cell Biol. 2017, 217, 65–77. [Google Scholar] [CrossRef]
- Evenson, D.P.; Djira, G.; Kasperson, K.; Christianson, J. Relationships between the age of 25,445 men attending infertility clinics and sperm chromatin structure assay (SCSA®) defined sperm DNA and chromatin integrity. Fertil. Steril. 2020, 114, 311–320. [Google Scholar] [CrossRef]
- Santana, V.P.; Furtado, C.; Pedroso, D.C.C.; Eiras, M.C.; Vasconcelos, M.A.C.; Ramos, E.; Calado, R.T.; Ferriani, R.A.; Esteves, S.C.; Dos Reis, R.M. The relationship among sperm global DNA methylation, telomere length, and DNA fragmentation in varicocele: A cross-sectional study of 20 cases. Syst. Biol. Reprod. Med. 2019, 65, 95–104. [Google Scholar] [CrossRef]
- Takemoto, M.; Mori, S.; Kuzuya, M.; Yoshimoto, S.; Shimamoto, A.; Igarashi, M.; Tanaka, Y.; Miki, T.; Yokote, K. Diagnostic criteria for Werner syndrome based on Japanese nationwide epidemiological survey. Geriatr. Gerontol. Int. 2013, 13, 475–481. [Google Scholar] [CrossRef]
- Oshima, J.; Sidorova, J.; Monnat, R.J. Werner syndrome: Clinical features, pathogenesis and potential therapeutic interventions. Ageing Res. Rev. 2017, 33, 105–114. [Google Scholar] [CrossRef] [PubMed]
- Shimamoto, A.; Yokote, K.; Tahara, H. Werner Syndrome-specific induced pluripotent stem cells: Recovery of telomere function by reprogramming. Front. Genet. 2015, 6. [Google Scholar] [CrossRef]
- Shamanna, R.A.; Lu, H.; De Freitas, J.K.; Tian, J.; Croteau, D.L.; Bohr, V. WRN regulates pathway choice between classical and alternative non-homologous end joining. Nat. Commun. 2016, 7, 13785. [Google Scholar] [CrossRef]
- Opresko, P. Werner syndrome and the function of the Werner protein; what they can teach us about the molecular aging process. Carcinogenesis 2003, 24, 791–802. [Google Scholar] [CrossRef] [PubMed]
- Poustka, A.; Dietrich, A.; Langenstein, G.; Toniolo, D.; Warren, S.T.; Lehrach, H. Physical map of human Xq27-qter: Localizing the region of the fragile X mutation. Proc. Natl. Acad. Sci. USA 1991, 88, 8302–8306. [Google Scholar] [CrossRef]
- Tashiro, S.; Nishihara, Y.; Kugou, K.; Ohta, K.; Kanoh, J. Subtelomeres constitute a safeguard for gene expression and chromosome homeostasis. Nucleic Acids Res. 2017, 45, 10333–10349. [Google Scholar] [CrossRef] [PubMed]
- Donkin, I.; Barrès, R. Sperm epigenetics and influence of environmental factors. Mol. Metab. 2018, 14, 1–11. [Google Scholar] [CrossRef]
- Kumar, D.; Salian, S.R.; Kalthur, G.; Uppangala, S.; Kumari, S.; Challapalli, S.; Chandraguthi, S.G.; Krishnamurthy, H.; Jain, N.; Kumar, P.; et al. Semen Abnormalities, Sperm DNA Damage and Global Hypermethylation in Health Workers Occupationally Exposed to Ionizing Radiation. PLoS ONE 2013, 8, e69927. [Google Scholar] [CrossRef]
- Zhang, W.; Li, M.; Sun, F.; Xu, X.; Zhang, Z.; Liu, J.; Sun, X.; Zhang, A.; Shen, Y.; Xu, J.; et al. Association of Sperm Methylation at LINE-1, Four Candidate Genes, and Nicotine/Alcohol Exposure With the Risk of Infertility. Front. Genet. 2019, 10. [Google Scholar] [CrossRef] [PubMed]
- Al Khaled, Y.; Tierling, S.; Laqqan, M.; Porto, C.L.; Hammadeh, M.E. Cigarette smoking induces only marginal changes in sperm DNA methylation levels of patients undergoing intracytoplasmic sperm injection treatment. Andrology 2018, 50, e12818. [Google Scholar] [CrossRef]
- Wang, Z.; Schwacke, R.; Kunze, R. DNA Damage-Induced Transcription of Transposable Elements and Long Non-coding RNAs in Arabidopsis Is Rare and ATM-Dependent. Mol. Plant 2016, 9, 1142–1155. [Google Scholar] [CrossRef]
- Lazaros, L.; Kitsou, C.; Kostoulas, C.; Bellou, S.; Hatzi, E.; Ladias, P.; Stefos, T.; Markoula, S.; Galani, V.; Vartholomatos, G.; et al. Retrotransposon expression and incorporation of cloned human and mouse retroelements in human spermatozoa. Fertil. Steril. 2017, 107, 821–830. [Google Scholar] [CrossRef]
- Reiner, B.C.; Doyle, G.A.; Weller, A.E.; Levinson, R.N.; Namoglu, E.; Pigeon, A.; Perea, E.D.; Weickert, C.S.; Turecki, G.; Mash, D.C.; et al. Restriction Enzyme Based Enriched L1Hs Sequencing (REBELseq): A Scalable Technique for Detection of Ta Subfamily L1Hs in the Human Genome. G3 Genes Genomes Genet. 2020, 10, 1647–1655. [Google Scholar] [CrossRef]
- Vondrak, T.; Robledillo, L. Ávila; Novák, P.; Koblížková, A.; Neumann, P.; Macas, J. Characterization of repeat arrays in ultra-long nanopore reads reveals frequent origin of satellite DNA from retrotransposon-derived tandem repeats. Plant J. 2019, 101, 484–500. [Google Scholar] [CrossRef]
- Xiao, T.; Zhou, W. The third generation sequencing: The advanced approach to genetic diseases. Transl. Pediatr. 2020, 9, 163–173. [Google Scholar] [CrossRef]
- Heather, J.M.; Chain, B. The sequence of sequencers: The history of sequencing DNA. Genomics 2016, 107, 1–8. [Google Scholar] [CrossRef] [PubMed]
- Förster, I.; Brockmann, M.; Schildgen, O.; Schildgen, V. Microsatellite instability testing in colorectal cancer using the QiaXcel advanced platform. BMC Cancer 2018, 18, 1–6. [Google Scholar] [CrossRef]
- Salipante, S.J.; Scroggins, S.M.; Hampel, H.L.; Turner, E.; Pritchard, C.C. Microsatellite Instability Detection by Next Generation Sequencing. Clin. Chem. 2014, 60, 1192–1199. [Google Scholar] [CrossRef]
- Cellerino, A.; Sanguanini, M. Next-generation RNA sequencing. In Transcriptome Analysis; Springer: Washington, DC, USA, 2018; pp. 11–25. [Google Scholar] [CrossRef]
- Luzhna, L.; Ekathiria, P.; Ekovalchuk, O. Micronuclei in genotoxicity assessment: From genetics to epigenetics and beyond. Front. Genet. 2013, 4, 131. [Google Scholar] [CrossRef] [PubMed]
- Morales-Ramírez, P.; Vallarino-Kelly, T.; Cruz-Vallejo, V.L. The OECD’s micronucleus test guideline for single exposure to an agent and the genotox-kinetic alternative. Mutagenesis 2017, 32, 411–415. [Google Scholar] [CrossRef] [PubMed][Green Version]
- Tice, R.R.; Agurell, E.; Anderson, D.; Burlinson, B.; Hartmann, A.; Kobayashi, H.; Miyamae, Y.; Rojas, E.; Ryu, J.C.; Sasaki, Y.F. Single cell gel/comet assay: Guidelines for in vitro and in vivo genetic toxicology testing. Environ. Mol. Mutagen. 2000, 35, 206–221. [Google Scholar] [CrossRef]

Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Puzuka, A.; Alksere, B.; Gailite, L.; Erenpreiss, J. Idiopathic Infertility as a Feature of Genome Instability. Life 2021, 11, 628. https://doi.org/10.3390/life11070628
Puzuka A, Alksere B, Gailite L, Erenpreiss J. Idiopathic Infertility as a Feature of Genome Instability. Life. 2021; 11(7):628. https://doi.org/10.3390/life11070628
Chicago/Turabian StylePuzuka, Agrita, Baiba Alksere, Linda Gailite, and Juris Erenpreiss. 2021. "Idiopathic Infertility as a Feature of Genome Instability" Life 11, no. 7: 628. https://doi.org/10.3390/life11070628
APA StylePuzuka, A., Alksere, B., Gailite, L., & Erenpreiss, J. (2021). Idiopathic Infertility as a Feature of Genome Instability. Life, 11(7), 628. https://doi.org/10.3390/life11070628