
Mechano-Immunomodulation in Space: Mechanisms Involving Microgravity-Induced Changes in T Cells

Abstract
1. The Importance of Human Health and Immunity for Space Exploration
2. Studying Cellular Responses to Altered Gravity
2.1. Microgravity Analogs
2.2. Microgravity Platforms
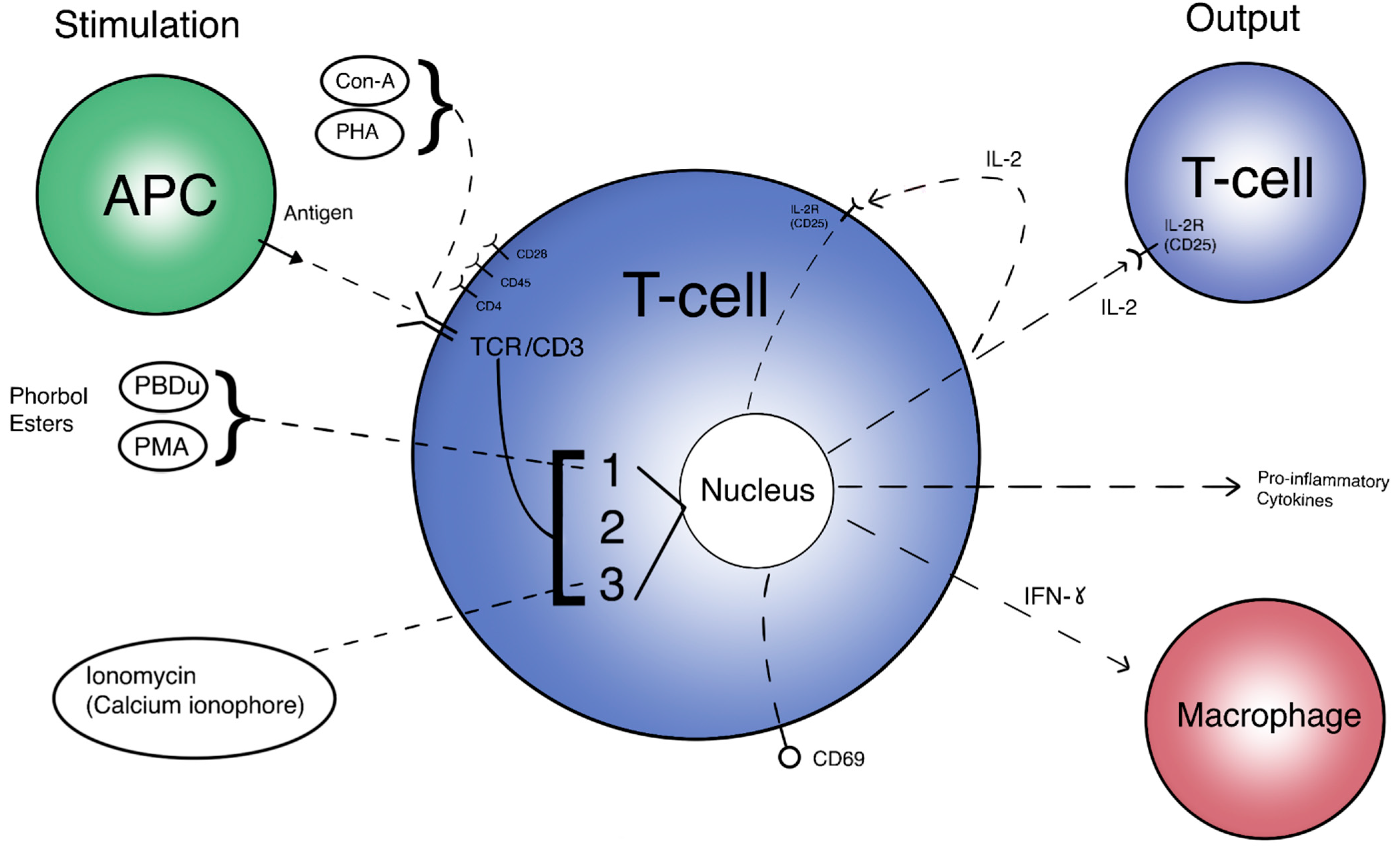
3. Mechanisms Involving Microgravity-Induced Changes in T Cell Signaling Pathways
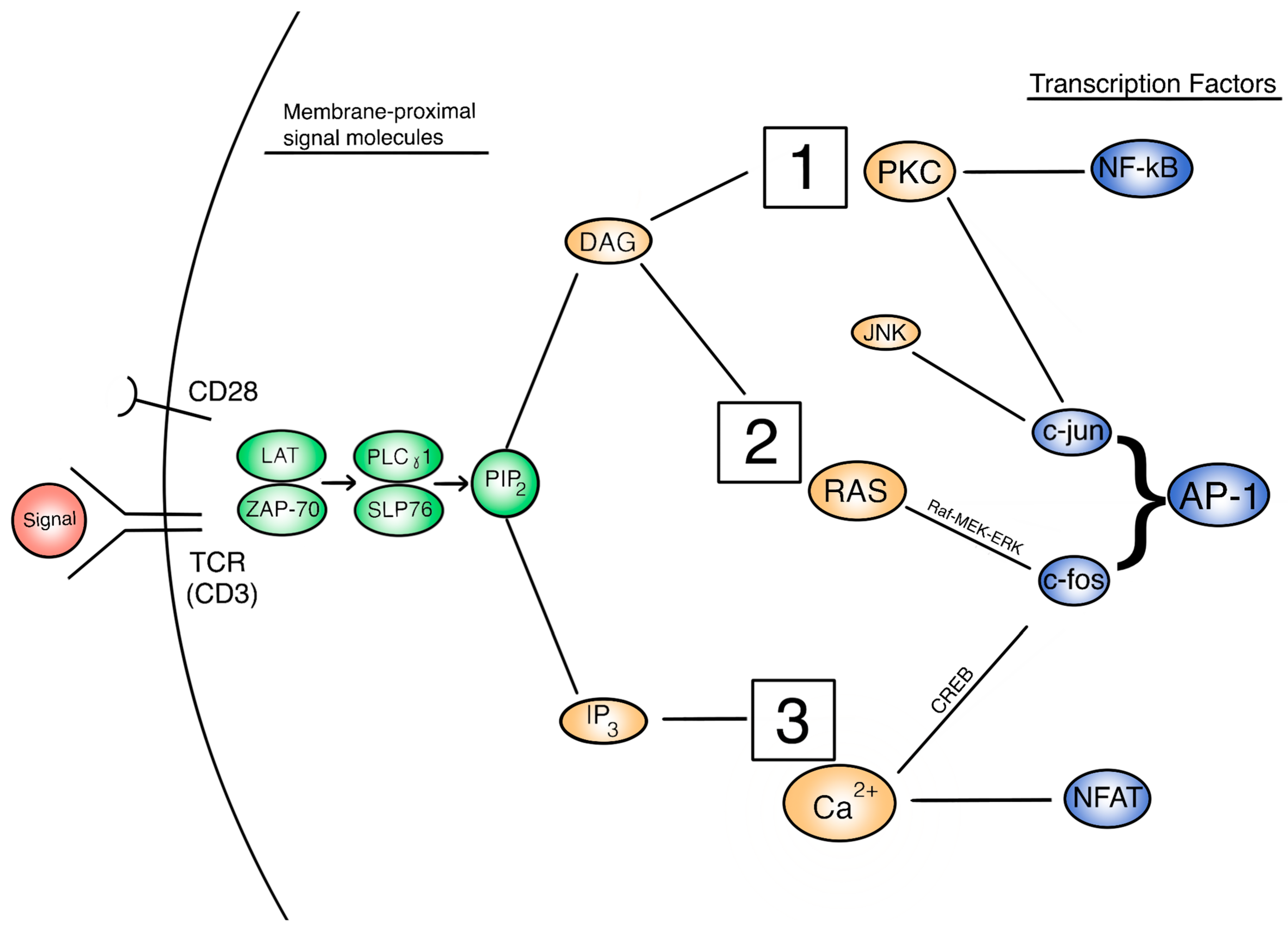
3.1. Signaling Pathways Responsible for T Cell Activation under Microgravity
3.1.1. PKC/NF-kB Pathways
3.1.2. Ras/AP-1 Pathways
3.1.3. Calcineurin/NFAT Pathways
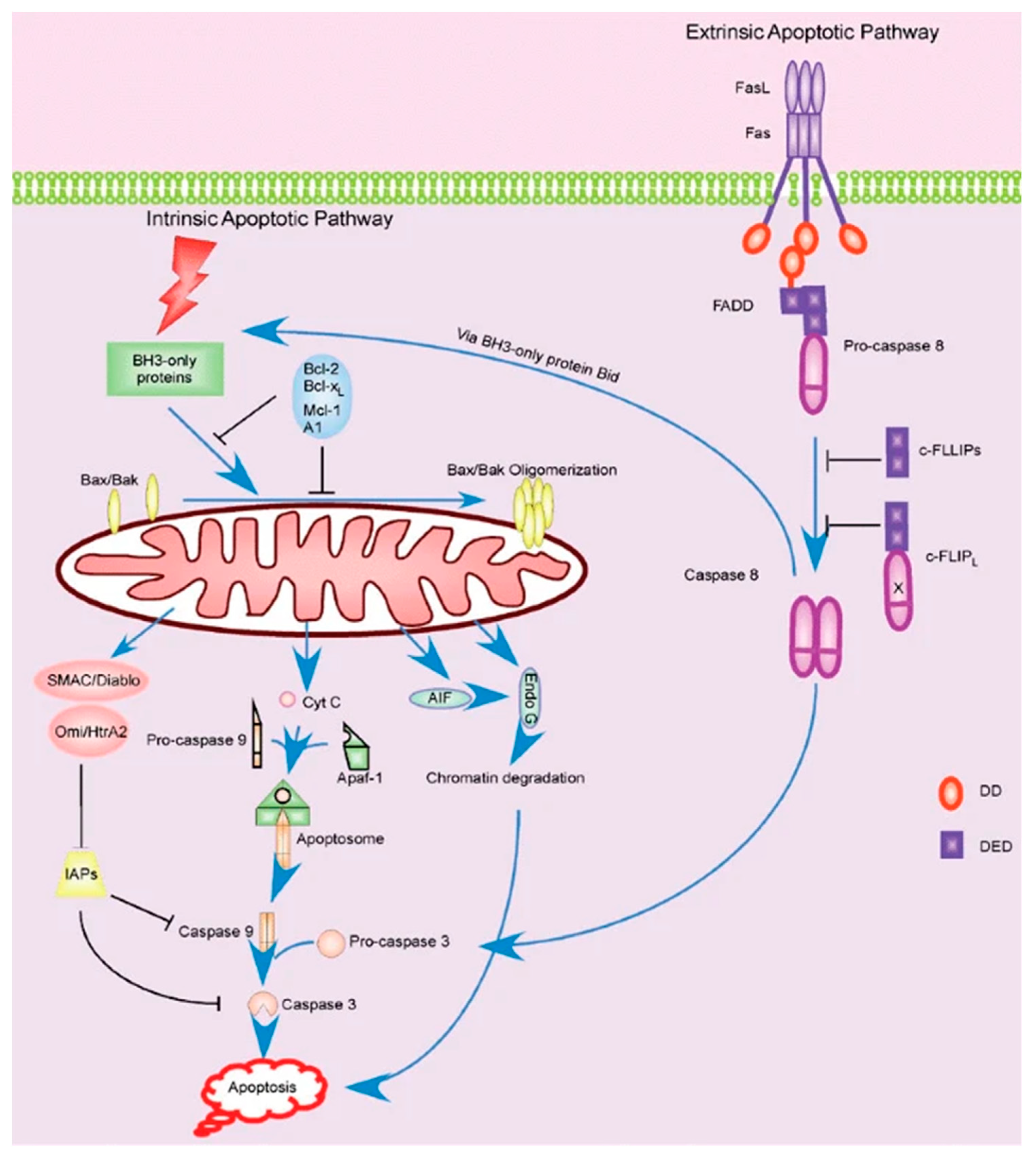
3.2. Signaling Pathways Responsible for T Cell Apoptosis under Microgravity
4. Future Directions
5. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- What is Artemis? Available online: https://www.nasa.gov/what-is-artemis (accessed on 1 August 2021).
- Hawkins, W.R.; Zieglschmid, J.F. Clinical aspects of crew health. Biomed. Res. Apollo 1975, NASA sp 368, 43–81. [Google Scholar]
- Bergman, S.A., Jr.; Johnson, R.L. Evaluation of the Electromechanical Properties of the Cardiovascular System After Prolonged Weightlessness. In Biomedical Results from Skylab; National Aeronautics and Space Administration: Washington, DC, USA, 1975; pp. 249–282. [Google Scholar]
- Konstantinova, I.V.; Antropova, Y.N.; Legenkov, V.I.; Zazhirey, V.D. Study of reactivity of blood lymphoid cells in crew members of the Soyuz-6, Soyuz-7 and Soyuz-8 spaceships before and after flight. Space Biol. Med. 1973, 7, 48–55. [Google Scholar]
- Klein-Nulend, J.; Bacabac, R.G.; Veldhuijzen, J.; Van Loon, J. Microgravity and bone cell mechanosensitivity. Adv. Space Res. 2003, 32, 1551–1559. [Google Scholar] [CrossRef]
- Ulbrich, C.; Wehland, M.; Pietsch, J.; Aleshcheva, G.; Wise, P.; Van Loon, J.; Magnusson, N.E.; Infanger, M.; Grosse, J.; Eilles, C.; et al. The Impact of Simulated and Real Microgravity on Bone Cells and Mesenchymal Stem Cells. BioMed Res. Int. 2014, 2014, 1–15. [Google Scholar] [CrossRef] [PubMed]
- Hughes-Fulford, M. Changes in gene expression and signal transduction in microgravity. J. Int. Soc. Gravit. Physiol. 2001, 8, 1–6. [Google Scholar]
- Lyndon B. Johnson Space Center. NASA Information—Muscle Atrophy. Available online: https://www.nasa.gov/pdf/64249main_ffs_factsheets_hbp_atrophy.pdf (accessed on 1 July 2021).
- Van Denburgh, H.; Chromiak, J.; Shansky, J.; Del Tatto, M.; Lemaire, J. Space travel directly induces skeletal muscle atrophy. FASEB J. 1999, 13, 1031–1038. [Google Scholar] [CrossRef] [PubMed]
- Droppert, P.M. A review of muscle atrophy in microgravity and during prolonged bed rest. J. Br. Interplanet. Soc. 1993, 46, 83–86. [Google Scholar]
- Goodwin, T.J.; Christofidou-Solomidou, M. Oxidative Stress and Space Biology: An Organ-Based Approach. Int. J. Mol. Sci. 2018, 19, 959. [Google Scholar] [CrossRef] [PubMed]
- Dunbar, B. Cardiovascular Health in Microgravity. Available online: https://www.nasa.gov/mission_pages/station/research/station-science-101/cardiovascular-health-in-microgravity/ (accessed on 1 August 2021).
- Roll, J.; Popov, K.; Gurfinkel, V.; Lipshits, M.; André-Deshays, C.; Gilhodes, J.; Quoniam, C. Sensorimotor and Perceptual Function of Muscle Proprioception in Microgravity. J. Vestib. Res. 1993, 3, 259–273. [Google Scholar] [CrossRef]
- Crucian, B.E.; Choukèr, A.; Simpson, R.; Mehta, S.; Marshall, G.; Smith, S.M.; Zwart, S.R.; Heer, M.; Ponomarev, S.; Whitmire, A.; et al. Immune System Dysregulation During Spaceflight: Potential Countermeasures for Deep Space Exploration Missions. Front. Immunol. 2018, 9, 1437. [Google Scholar] [CrossRef]
- Crucian, B.E.; Stowe, R.P.; Pierson, D.L.; Sams, C.F. Immune System Dysregulation Following Short- vs. Long-Duration Spaceflight. Aviat. Space Environ. Med. 2008, 79, 835–843. [Google Scholar] [CrossRef] [PubMed]
- Baqai, F.P.; Gridley, D.S.; Slater, J.M.; Luo-Owen, X.; Stodieck, L.S.; Ferguson, V.; Chapes, S.K.; Pecaut, M.J. Effects of spaceflight on innate immune function and antioxidant gene expression. J. Appl. Physiol. 2009, 106, 1935–1942. [Google Scholar] [CrossRef] [PubMed]
- Garrett-Bakelman, F.E.; Darshi, M.; Green, S.J.; Gur, R.C.; Lin, L.; Macias, B.R.; McKenna, M.J.; Meydan, C.; Mishra, T.; Nasrini, J.; et al. The NASA Twins Study: A multidimensional analysis of a year-long human spaceflight. Science 2019, 364, eaau8650. [Google Scholar] [CrossRef] [PubMed]
- Harland, D.M. Microgravity. Available online: https://www.britannica.com/science/microgravity (accessed on 7 July 2021).
- Freed, L.E.; Langer, R.; Martin, I.; Pellis, N.R.; Vunjak-Novakovic, G. Tissue engineering of cartilage in space. Proc. Natl. Acad. Sci. USA 1997, 94, 13885–13890. [Google Scholar] [CrossRef] [PubMed]
- Ulbrich, C.; Westphal, K.; Pietsch, J.; Winkler, H.; Leder, A.; Bauer, J.; Kossmehl, P.; Grosse, J.; Schoenberger, J.; Infanger, M.; et al. Characterization of Human Chondrocytes Exposed to Simulated Microgravity. Cell. Physiol. Biochem. 2010, 25, 551–560. [Google Scholar] [CrossRef]
- Delves, P.J. Overview of the Immune System. Available online: https://www.merckmanuals.com/professional/immunology-allergic-disorders/biology-of-the-immune-system/overview-of-the-immune-system (accessed on 1 July 2021).
- IQWiG (Institute for Quality and Efficiency in Health Care). The Innate and Adaptive Immune Systems. Available online: https://www.ncbi.nlm.nih.gov/books/NBK279396/ (accessed on 1 July 2021).
- Guéguinou, N.; Huin-Schohn, C.; Bascove, M.; Bueb, J.-L.; Tschirhart, E.; Legrand-Frossi, C.; Frippiat, J.-P. Could spaceflight-associated immune system weakening preclude the expansion of human presence beyond Earth’s orbit? J. Leukoc. Biol. 2009, 86, 1027–1038. [Google Scholar] [CrossRef]
- Cogoli, A.; Tschopp, A.; Fuchs-Bislin, P. Cell sensitivity to gravity. Science 1984, 225, 228–230. [Google Scholar] [CrossRef] [PubMed]
- Cooper, D.; Pride, M.W.; Brown, E.L.; Risin, D.; Pellis, N.R. Suppression of antigen-specific lymphocyte activation in modeled microgravity. Vitr. Cell. Dev. Biol. Anim. 2001, 37, 63–65. [Google Scholar] [CrossRef]
- Wardle, E.N. Guide to Signal Pathways in Immune Cells; Springer: Berlin, Germany, 2009; pp. 247–277. [Google Scholar] [CrossRef]
- Sundaresan, A.; Pellis, N.R. Cellular and Genetic Adaptation in Low-gravity Environments. Ann. N. Y. Acad. Sci. 2009, 1161, 135–146. [Google Scholar] [CrossRef]
- Hughes-Fulford, M. Function of the cytoskeleton in gravisensing during spaceflight. Adv. Space Res. 2003, 32, 1585–1593. [Google Scholar] [CrossRef]
- Ullrich, O.; Huber, K.; Lang, K. Signal transduction in cells of the immune system in microgravity. Cell Commun. Signal. 2008, 6, 9. [Google Scholar] [CrossRef]
- Fitzgerald, W.; Chen, S.; Walz, C.; Zimmerberg, J.; Margolis, L.; Grivel, J.-C. Immune suppression of human lymphoid tissues and cells in rotating suspension culture and onboard the International Space Station. Vitr. Cell. Dev. Biol. Anim. 2009, 45, 622–632. [Google Scholar] [CrossRef][Green Version]
- Hughes-Fulford, M.; Chang, T.T.; Martinez, E.M.; Li, C. Spaceflight alters expression of microRNA during T-cell activation. FASEB J. 2015, 29, 4893–4900. [Google Scholar] [CrossRef]
- Ratliff, D. The Next Frontier: Stem Cells and the Center for the Advancement of Science in Space. Stem Cells Dev. 2013, 22, 94–95. [Google Scholar] [CrossRef]
- Brungs, S.; Kolanus, W.; Hemmersbach, R. Syk phosphorylation—A gravisensitive step in macrophage signalling. Cell Commun. Signal. 2015, 13, 1–9. [Google Scholar] [CrossRef] [PubMed]
- Maier, J. Impact of Simulated Microgravity on Cell Cycle Control and Cytokine Release by U937 Cells. Int. J. Immunopathol. Pharmacol. 2006, 19, 279–286. [Google Scholar] [CrossRef] [PubMed]
- Wuest, S.L.; Richard, S.; Kopp, S.; Grimm, D.; Egli, M. Simulated Microgravity: Critical Review on the Use of Random Positioning Machines for Mammalian Cell Culture. BioMed Res. Int. 2015, 2015, 971474. [Google Scholar] [CrossRef] [PubMed]
- Herranz, R.; Anken, R.; Boonstra, J.; Braun, M.; Christianen, P.C.; De Geest, M.; Hauslage, J.; Hilbig, R.; Hill, R.J.; Lebert, M.; et al. Ground-Based Facilities for Simulation of Microgravity: Organism-Specific Recommendations for Their Use, and Recommended Terminology. Astrobiology 2013, 13, 1–17. [Google Scholar] [CrossRef]
- Albrecht-Buehler, G. The simulation of microgravity conditions on the ground. ASGSB Bull. Publ. Am. Soc. Gravit. Space Biol. 1992, 5, 3–10. [Google Scholar]
- Martinelli, L.; Russomano, T.; Dos Santos, M.; Falcao, F.; Bauer, M.; Machado, A.; Sundaresan, A. Effect of microgravity on immune cell viability and proliferation. IEEE Eng. Med. Boil. Mag. 2009, 28, 85–90. [Google Scholar] [CrossRef] [PubMed]
- Liu, H.; Wang, Z.-C.; Bai, Y.-G.; Cai, Y.; Yu, J.-W.; Zhang, H.-J.; Bao, J.-X.; Ren, X.-L.; Xie, M.-J.; Ma, J. Simulated microgravity promotes monocyte adhesion to rat aortic endothelium via nuclear factor-κB activation. Clin. Exp. Pharmacol. Physiol. 2015, 42, 510–519. [Google Scholar] [CrossRef]
- Morey-Holton, E.R.; Globus, R.K. Hindlimb unloading rodent model: Technical aspects. J. Appl. Physiol. 2002, 92, 1367–1377. [Google Scholar] [CrossRef] [PubMed]
- Chapes, S.K.; Mastro, A.M.; Sonnenfeld, G.; Berry, W.D. Antiorthostatic suspension as a model for the effects of spaceflight on the immune system. J. Leukoc. Biol. 1993, 54, 227–235. [Google Scholar] [CrossRef]
- Armstrong, J.W.; Nelson, K.A.; Simske, S.J.; Luttges, M.W.; Iandolo, J.J.; Chapes, S.K. Skeletal unloading causes organ-specific changes in immune cell responses. J. Appl. Physiol. 1993, 75, 2734–2739. [Google Scholar] [CrossRef]
- Crucian, B.E.; Stowe, R.P.; Mehta, S.K.; Yetman, D.L.; Leal, M.J.; Quiriarte, H.D.; Pierson, D.L.; Sams, C.F. Immune Status, Latent Viral Reactivation, and Stress During Long-Duration Head-Down Bed Rest. Aviat. Space Environ. Med. 2009, 80, A37–A44. [Google Scholar] [CrossRef]
- Choukèr, A.; Thiel, M.; Baranov, V.; Meshkov, D.; Kotov, A.; Peter, K.; Messmer, K.; Christ, F. Simulated microgravity, psychic stress, and immune cells in men: Observations during 120-day 6° HDT. J. Appl. Physiol. 2001, 90, 1736–1743. [Google Scholar] [CrossRef] [PubMed]
- Xu, X.; Tan, C.; Li, P.; Zhang, S.; Pang, X.; Liu, H.; Li, L.; Sun, X.; Zhang, Y.; Wu, H.; et al. Changes of Cytokines during a Spaceflight Analog a 45-Day Head-Down Bed Rest. PLoS ONE 2013, 8, e77401. [Google Scholar] [CrossRef] [PubMed]
- Kelsen, J.; Bartels, L.E.; Dige, A.; Hvas, C.L.; Frings-Meuthen, P.; Boehme, G.; Thomsen, M.K.; Fenger-Grøn, M.; Dahlerup, J.F. 21 Days head-down bed rest induces weakening of cell-mediated immunity Some spaceflight findings confirmed in a ground-based analog. Cytokine 2012, 59, 403–409. [Google Scholar] [CrossRef] [PubMed]
- Hoff, P.; Belavy, D.; Huscher, D.; Lang, A.; Hahne, M.; Kuhlmey, A.-K.; Maschmeyer, P.; Armbrecht, G.; Fitzner, R.; Perschel, F.H.; et al. Effects of 60-day bed rest with and without exercise on cellular and humoral immunological parameters. Cell. Mol. Immunol. 2014, 12, 483–492. [Google Scholar] [CrossRef] [PubMed]
- Schmitt, D.; Schwarzenberg, M.; Tkaczuk, J.; Hebrard, S.; Brandenberger, G.; Mauco, G.; Cogoli-Greuter, M.; Abbal, M. Head-down tilt bed rest and immune responses. Pflugers Arch. 2000, 441, R79–R84. [Google Scholar] [CrossRef]
- Gmünder, F.K.; Baisch, F.; Bechler, B.; Cogoli, A.; Cogoli, M.; Joller, P.W.; Maass, H.; Müller, J.; Ziegler, W.H. Effect of head-down tilt bedrest (10 days) on lymphocyte reactivity. Acta Physiol. Scand. Suppl. 1992, 604, 131–141. [Google Scholar]
- Paulsen, K.; Thiel, C.; Timm, J.; Schmidt, P.M.; Huber, K.; Tauber, S.; Hemmersbach, R.; Seibt, D.; Kroll, H.; Grote, K.-H.; et al. Microgravity-induced alterations in signal transduction in cells of the immune system. Acta Astronaut. 2010, 67, 1116–1125. [Google Scholar] [CrossRef]
- Tauber, S.; Hauschild, S.; Paulsen, K.; Gutewort, A.; Raig, C.; Hürlimann, E.; Biskup, J.; Philpot, C.; Lier, H.; Engelmann, F.; et al. Signal Transduction in Primary Human T Lymphocytes in Altered Gravity During Parabolic Flight and Clinostat Experiments. Cell. Physiol. Biochem. 2015, 35, 1034–1051. [Google Scholar] [CrossRef]
- Hashemi, B.B.; Penkala, J.E.; Vens, C.; Huls, H.; Cubbage, M.; Sams, C.F. T cell activation responses are differentially regulated during clinorotation and in spaceflight. FASEB J. 1999, 13, 2071–2082. [Google Scholar] [CrossRef] [PubMed]
- Thiel, C.S.; Huge, A.; Hauschild, S.; Tauber, S.; Lauber, B.A.; Polzer, J.; Paulsen, K.; Lier, H.; Engelmann, F.; Schmitz, B.; et al. Stability of gene expression in human T cells in different gravity environments is clustered in chromosomal region 11p15.4npj. Microgravity 2017, 3, 22. [Google Scholar] [CrossRef] [PubMed]
- Thiel, C.S.; Christoffel, S.; Tauber, S.; Vahlensieck, C.; De Zélicourt, D.; Layer, L.E.; Lauber, B.; Polzer, J.; Ullrich, O. Rapid Cellular Perception of Gravitational Forces in Human Jurkat T Cells and Transduction into Gene Expression Regulation. Int. J. Mol. Sci. 2020, 21, 514. [Google Scholar] [CrossRef]
- Thiel, C.S.; Paulsen, K.; Bradacs, G.; Lust, K.; Tauber, S.; Dumrese, C.; Hilliger, A.; Schoppmann, K.; Biskup, J.; Gölz, N.; et al. Rapid alterations of cell cycle control proteins in human T lymphocytes in microgravity. Cell Commun. Signal. 2012, 10, 1. [Google Scholar] [CrossRef]
- Wang, C.; Li, N.; Zhang, C.; Sun, S.; Gao, Y.; Long, M. Effects of Simulated Microgravity on Functions of Neutrophil-like HL-60 Cells. Microgravity Sci. Technol. 2015, 27, 515–527. [Google Scholar] [CrossRef]
- Ritz, B.W.; Lelkes, P.I.; Gardner, E.M. Functional recovery of peripheral blood mononuclear cells in modeled microgravity. FASEB J. 2005, 20, 305–307. [Google Scholar] [CrossRef][Green Version]
- Spatz, J.M.; Fulford, M.H.; Tsai, A.; Gaudilliere, D.; Hedou, J.; Ganio, E.; Angst, M.; Aghaeepour, N.; Gaudilliere, B. Human immune system adaptations to simulated microgravity revealed by single-cell mass cytometry. Sci. Rep. 2021, 11, 11872. [Google Scholar] [CrossRef]
- Singh, K.P.; Kumari, R.; DuMond, J.W. Simulated microgravity-induced epigenetic changes in human lymphocytes. J. Cell. Biochem. 2010, 111, 123–129. [Google Scholar] [CrossRef] [PubMed]
- Wei, L.; Liu, C.; Kang, L.; Liu, Y.; Shi, S.; Wu, Q.; Li, Y. Experimental Study on Effect of Simulated Microgravity on Structural Chromosome Instability of Human Peripheral Blood Lymphocytes. PLoS ONE 2014, 9, e100595. [Google Scholar] [CrossRef]
- Simons, D.M.; Gardner, E.M.; Lelkes, P.I.; Cabrales, P.; Tsai, A.G.; Johnson, P.C.; Intaglietta, M. Dynamic culture in a rotating-wall vessel bioreactor differentially inhibits murine T-lymphocyte activation by mitogenic stimuli upon return to static conditions in a time-dependent manner. J. Appl. Physiol. 2006, 100, 1287–1292. [Google Scholar] [CrossRef]
- Bakos, A.; Varkonyi, A.; Minarovits, J.; Batkai, L. Effect of simulated microgravity on the production of IL-12 by PBMCs. J. Int. Soc. Gravit. Physiol. 2002, 9, 335–336. [Google Scholar]
- Walther, I.; Cogoli, A.; Pippia, P.A.; Meloni, M.; Cossu, G.; Cogoli, M.; Schwarzenberg, M.; Turrini, F.; Mannu, F. Human immune cells as space travelers. Eur. J. Med. Res. 1999, 4, 361–363. [Google Scholar]
- Luo, H.; Wang, C.; Feng, M.; Zhao, Y. Microgravity Inhibits Resting T Cell Immunity in an Exposure Time-Dependent Manner. Int. J. Med. Sci. 2014, 11, 87–96. [Google Scholar] [CrossRef] [PubMed]
- Sundaresan, A.; Risin, D.; Pellis, N.R. Loss of Signal Transduction and inhibition of Lymphocyte Locomotion in a ground-based model of microgravity. Vitr. Cell. Dev. Biol. Anim. 2002, 38, 118. [Google Scholar] [CrossRef]
- Cooper, D.; Pellis, N.R. Suppressed PHA activation of T lymphocytes in simulated microgravity is restored by direct activation of protein kinase C. J. Leukoc. Biol. 1998, 63, 550–562. [Google Scholar] [CrossRef]
- Boonyaratanakornkit, J.B.; Cogoli, A.; Li, C.; Schopper, T.; Pippia, P.; Galleri, G.; Meloni, M.A.; Hughes-Fulford, M. Key gravity-sensitive signaling pathways drive T-cell activation. FASEB J. 2005, 19, 2020–2022. [Google Scholar] [CrossRef] [PubMed]
- Walther, I.; Pippia, P.; Meloni, M.A.; Turrini, F.; Mannu, F.; Cogoli, A. Simulated microgravity inhibits the genetic expression of interleukin-2 and its receptor in mitogen-activated T lymphocytes. FEBS Lett. 1998, 436, 115–118. [Google Scholar] [CrossRef]
- Martinez, E.M.; Yoshida, M.C.; Candelario, T.L.T.; Hughes-Fulford, M. Spaceflight and simulated microgravity cause a significant reduction of key gene expression in early T-cell activation. Am. J. Physiol. Integr. Comp. Physiol. 2015, 308, 480–488. [Google Scholar] [CrossRef]
- Galleri, G.; Meloni, M.A.; Camboni, M.G.; Deligios, M.; Cogoli, A.; Pippia, P. Signal transduction in T lymphocites under sim-ulated microgravity conditions: Involvement of PKC isoforms. J. Gravit. Physiol. 2002, 9, 289–290. [Google Scholar]
- Van Walleghem, M.; Tabury, K.; Fernandez-Gonzalo, R.; Janssen, A.; Buchheim, J.-I.; Choukèr, A.; Baatout, S.; Moreels, M. Gravity-Related Immunological Changes in Human Whole Blood Cultured Under Simulated Microgravity Using an In Vitro Cytokine Release Assay. J. Interferon Cytokine Res. 2017, 37, 531–540. [Google Scholar] [CrossRef] [PubMed]
- Hoson, T.; Kamisaka, S.; Masuda, Y.; Yamashita, M.; Buchen, B. Evaluation of the three-dimensional clinostat as a simulator of weightlessness. Planta 1997, 203, 187–197. [Google Scholar] [CrossRef] [PubMed]
- Nomura, J.; Arase, Y.; Sugita, T.; Sugaya, S.; Takahashi, S.; Kita, K.; Yamamori, H.; Sekiya, S.; Suzuki, N. Increased and de-creased expression of CD69 and CD23, respectively, in gravity-stressed lymphocytes. Aviat. Space Environ. Med. 2001, 72, 727–732. [Google Scholar] [PubMed]
- Thiel, C.S.; Hauschild, S.; Huge, A.; Tauber, S.; Lauber, B.A.; Polzer, J.; Paulsen, K.; Lier, H.; Engelmann, F.; Schmitz, B.; et al. Dynamic gene expression response to altered gravity in human T cells. Sci. Rep. 2017, 7, 5204. [Google Scholar] [CrossRef]
- Bechler, B.; Cogoli, A.; Cogoli-Greuter, M.; Müller, O.; Hunzinger, E.; Criswell, S.B. Activation of microcarrier-attached lymphocytes in microgravity. Biotechnol. Bioeng. 1992, 40, 991–996. [Google Scholar] [CrossRef]
- Moser, D.; Sun, S.J.; Li, N.; Biere, K.; Hoerl, M.; Matzel, S.; Feuerecker, M.; Buchheim, J.-I.; Strewe, C.; Thiel, C.S.; et al. Cells’ Flow and Immune Cell Priming under alternating g-forces in Parabolic Flight. Sci. Rep. 2019, 9, 11276. [Google Scholar] [CrossRef]
- Vogel, J.; Thiel, C.S.; Tauber, S.; Stockmann, C.; Gassmann, M.; Ullrich, O. Expression of Hypoxia-Inducible Factor 1α (HIF-1α) and Genes of Related Pathways in Altered Gravity. Int. J. Mol. Sci. 2019, 20, 436. [Google Scholar] [CrossRef]
- Stervbo, U.; Roch, T.; Kornprobst, T.; Sawitzki, B.; Grütz, G.; Wilhelm, A.; Lacombe, F.; Allou, K.; Kaymer, M.; Pacheco, A.; et al. Gravitational stress during parabolic flights reduces the number of circulating innate and adaptive leukocyte subsets in human blood. PLoS ONE 2018, 13, e0206272. [Google Scholar] [CrossRef]
- Tauber, S.; Christoffel, S.; Thiel, C.S.; Ullrich, O. Transcriptional Homeostasis of Oxidative Stress-Related Pathways in Altered Gravity. Int. J. Mol. Sci. 2018, 19, 2814. [Google Scholar] [CrossRef] [PubMed]
- Chang, T.T.; Walther, I.; Li, C.-F.; Boonyaratanakornkit, J.; Galleri, G.; Meloni, M.A.; Pippia, P.; Cogoli, A.; Hughes-Fulford, M. The Rel/NF-κB pathway and transcription of immediate early genes in T cell activation are inhibited by microgravity. J. Leukoc. Biol. 2012, 92, 1133–1145. [Google Scholar] [CrossRef]
- Cogoli, M.; Bechler, B.; Cogoli, A.; Arena, N.; Barni, S.; Pippia, P.; Sechi, G.; Valora, N.; Monti, R. Lymphocytes on sounding rockets. Adv. Space Res. 1992, 12, 141–144. [Google Scholar] [CrossRef]
- Paulsen, K.; Tauber, S.; Dumrese, C.; Bradacs, G.; Simmet, D.M.; Gölz, N.; Hauschild, S.; Raig, C.; Engeli, S.; Gutewort, A.; et al. Regulation of ICAM-1 in Cells of the Monocyte/Macrophage System in Microgravity. BioMed Res. Int. 2015, 2015, 1–18. [Google Scholar] [CrossRef]
- Why Do Science in Microgravity? Available online: https://www.nasa.gov/mission_pages/station/research/station-science-101/why-do-science-in-microgravity/ (accessed on 1 August 2021).
- Crucian, B.; Stowe, R.P.; Mehta, S.; Quiriarte, H.; Pierson, D.; Sams, C. Alterations in adaptive immunity persist during long-duration spaceflightnpj. Microgravity 2015, 1, 15013. [Google Scholar] [CrossRef]
- Novoselova, E.; Lunin, S.; Khrenov, M.; Parfenyuk, S.; Novoselova, T.; Shenkman, B.; Fesenko, E. Changes in immune cell signalling, apoptosis and stress response functions in mice returned from the BION-M1 mission in space. Immunobiology 2015, 220, 500–509. [Google Scholar] [CrossRef] [PubMed]
- Rykova, M.; Antropova, E.; Larina, I.; Morukov, B. Humoral and cellular immunity in cosmonauts after the ISS missions. Acta Astronaut. 2008, 63, 697–705. [Google Scholar] [CrossRef]
- Ortega, M.T.; Lu, N.; Chapes, S.K. Evaluation of in vitro macrophage differentiation during space flight. Adv. Space Res. 2012, 49, 1441–1455. [Google Scholar] [CrossRef]
- Risso, A.; Tell, G.; Vascotto, C.; Costessi, A.; Arena, S.; Scaloni, A.; Cosulich, M.E. Activation of human T lymphocytes under conditions similar to those that occur during exposure to microgravity: A proteomics study. Proteomics 2005, 5, 1827–1837. [Google Scholar] [CrossRef]
- Gasperi, V.; Rapino, C.; Battista, N.; Bari, M.; Mastrangelo, N.; Angeletti, S.; Dainese, E.; Maccarrone, M. A Functional Interplay between 5-Lipoxygenase andμ-Calpain Affects Survival and Cytokine Profile of Human Jurkat T Lymphocyte Exposed to Simulated Microgravity. BioMed Res. Int. 2014, 2014, 782390. [Google Scholar] [CrossRef]
- Li, X.; Liu, C.-T.; Zhou, H. The influence of leptin on the activity of lung lymphocytes under simulated microgravity. Graefe’s Arch. Clin. Exp. Ophthalmol. 2009, 107, 335–344. [Google Scholar] [CrossRef]
- Bradley, J.H.; Stein, R.; Randolph, B.; Molina, E.; Arnold, J.P.; Gregg, R.K. T cell resistance to activation by dendritic cells requires long-term culture in simulated microgravity. Life Sci. Space Res. 2017, 15, 55–61. [Google Scholar] [CrossRef]
- Rinaldi, S.; Meloni, M.A.; Galleri, G.; Maioli, M.; Pigliaru, G.; Cugia, G.; Santaniello, S.; Castagna, A.; Fontani, V. Radio Electric Asymmetric Conveyer (REAC) technology to obviate loss of T cell responsiveness under simulated microgravity. PLoS ONE 2018, 13, e0200128. [Google Scholar] [CrossRef] [PubMed]
- Limouse, M.; Manié, S.; Konstantinova, I.; Ferrua, B.; Schaffar, L. Inhibition of phorbol ester-induced cell activation in microgravity. Exp. Cell Res. 1991, 197, 82–86. [Google Scholar] [CrossRef]
- Pippia, P.; Sciola, L.; Cogoli-Greuter, M.; Meloni, M.A.; Spano, A.; Cogoli, A. Activation signals of T lymphocytes in microgravity. J. Biotechnol. 1996, 47, 215–222. [Google Scholar] [CrossRef]
- Nash, P.V.; Mastro, A. Variable lymphocyte responses in rats after space flight. Exp. Cell Res. 1992, 202, 125–131. [Google Scholar] [CrossRef]
- Chapes, S.K.; Morrison, D.R.; Guikema, J.A.; Lewis, M.L.; Spooner, B.S. Cytokine secretion by immune cells in space. J. Leukoc. Biol. 1992, 52, 104–110. [Google Scholar] [CrossRef]
- Grove, D.S.; Pishak, S.A.; Mastro, A. The Effect of a 10-Day Space Flight on the Function, Phenotype, and Adhesion Molecule Expression of Splenocytes and Lymph Node Lymphocytes. Exp. Cell Res. 1995, 219, 102–109. [Google Scholar] [CrossRef]
- Hwang, S.-A.; Crucian, B.; Sams, C.; Actor, J.K. Post-Spaceflight (STS-135) Mouse Splenocytes Demonstrate Altered Activation Properties and Surface Molecule Expression. PLoS ONE 2015, 10, e0124380. [Google Scholar] [CrossRef]
- Nash, P.V.; Konstantinova, I.V.; Fuchs, B.B.; Rakhmilevich, A.L.; Lesnyak, A.T.; Mastro, A.M. Effect of spaceflight on lymphocyte proliferation and interleukin-2 production. J. Appl. Physiol. 1992, 73, S186–S190. [Google Scholar] [CrossRef]
- Crucian, B.; Stowe, R.; Mehta, S.; Uchakin, P.; Quiriarte, H.; Pierson, D.; Sams, C. Immune System Dysregulation Occurs During Short Duration Spaceflight on Board the Space Shuttle. J. Clin. Immunol. 2012, 33, 456–465. [Google Scholar] [CrossRef]
- Chang, T.T.; Spurlock, S.M.; Candelario, T.L.T.; Grenon, S.M.; Hughes-Fulford, M. Spaceflight impairs antigen-specific tolerance induction in vivo and increases inflammatory cytokines. FASEB J. 2015, 29, 4122–4132. [Google Scholar] [CrossRef]
- Crucian, B.E.; Cubbage, M.L.; Sams, C.F. Altered Cytokine Production by Specific Human Peripheral Blood Cell Subsets Immediately Following Space Flight. J. Interferon Cytokine Res. 2000, 20, 547–556. [Google Scholar] [CrossRef] [PubMed]
- Konstantinova, I.V.; Sonnenfeld, G.; Lesnyak, A.T.; Shaffar, L.; Mandel, A.; Rykova, M.P.; Antropova, E.N.; Ferrua, B. Cellular immunity and lymphokine production during spaceflights. Physiology 1991, 34, 52–56. [Google Scholar]
- Stowe, R.P.; Sams, C.F.; Pierson, D.L. Adrenocortical and immune responses following short- and long-duration spaceflight. Aviat. Space Environ. Med. 2011, 82, 627–634. [Google Scholar] [CrossRef] [PubMed]
- Irina, V.; Konstantinova, M. Immune resistance of man in space flights. Acta Astronaut. 1991, 23, 123–127. [Google Scholar] [CrossRef]
- Perbal, B. Communication is the key. Cell Commun. Signal. 2003, 1, 3. [Google Scholar] [CrossRef]
- Bradshaw, R.A.; Dennis, E.A. Cell Signaling: Yesterday, Today, and Tomorrow. In Handbook of Cell Signaling, 2nd ed.; Academic Press: Cambridge, MA, USA, 2009. [Google Scholar]
- O’Connor, C.; Adams, J.U.; Fairman, J.E. Essentials of Cell Biology Unit 4: How Do Cells Sense Their Environment? Available online: https://www.nature.com/scitable/ebooks/essentials-of-cell-biology-14749010/how-do-cells-sense-their-environment-14751787/ (accessed on 10 August 2021).
- Ludtka, C.; Silberman, J.; Moore, E.; Allen, J.B. Macrophages in microgravity: The impact of space on immune cells. npj Microgravity 2021, 7, 13. [Google Scholar] [CrossRef]
- Woods, C.C.; Banks, K.E.; Gruener, R.; DeLuca, D. Loss of T cell precursors after spaceflight and exposure to vector-averaged gravity. FASEB J. 2003, 17, 1–17. [Google Scholar] [CrossRef] [PubMed]
- Woods, C.C.; Banks, K.E.; Lebsack, T.W.; White, T.C.; Anderson, G.A.; Maccallum, T.; Gruener, R.; DeLuca, D. Use of a microgravity organ culture dish system to demonstrate the signal dampening effects of modeled microgravity during T cell development. Dev. Comp. Immunol. 2005, 29, 565–582. [Google Scholar] [CrossRef] [PubMed]
- Sonnenfeld, G.; Mandel, A.D.; Konstantinova, I.V.; Berry, W.D.; Taylor, G.R.; Lesnyak, A.T.; Fuchs, B.B.; Rakhmilevich, A.L. Spaceflight alters immune cell function and distribution. J. Appl. Physiol. 1992, 73, S191–S195. [Google Scholar] [CrossRef] [PubMed]
- Simons, D.M.; Gardner, E.M.; Lelkes, P.I. Sub-mitogenic phorbol myristate acetate co-stimulation rescues the PHA-induced activation of both naïve and memory T cells cultured in the rotating-wall vessel bioreactor. Cell Biol. Int. 2009, 33, 882–886. [Google Scholar] [CrossRef] [PubMed]
- Pecaut, M.J.; Nelson, G.A.; Peters, L.L.; Kostenuik, P.J.; Bateman, T.A.; Morony, S.; Stodieck, L.S.; Lacey, D.L.; Simske, S.J.; Gridley, D.S. Selected Contribution: Effects of spaceflight on immunity in the C57BL/6 mouse. I. Immune population distributions. J. Appl. Physiol. 2003, 94, 2085–2094. [Google Scholar] [CrossRef]
- Häggström, M. Medical gallery of Mikael Häggström 2014. WikiJ. Med. 2014, 1, 1–53. [Google Scholar] [CrossRef]
- Tauber, S.; Hauschild, S.; Crescio, C.; Secchi, C.; Paulsen, K.; Pantaleo, A.; Saba, A.; Buttron, I.; Thiel, C.S.; Cogoli, A.; et al. Signal transduction in primary human T lymphocytes in altered gravity results of the MASER-12 suborbital space flight mission. Cell Commun. Signal. 2013, 11, 32. [Google Scholar] [CrossRef]
- Schmitt, D.A.; Hatton, J.P.; Emond, C.; Chaput, D.; Paris, H.; Levade, T.; Cazenave, J.; Schaffar, L. The distribution of protein kinase C in human leukocytes is altered in microgravity. FASEB J. 1996, 10, 1627–1634. [Google Scholar] [CrossRef]
- Simons, D.; Gardner, E.; Lelkes, P. Intact T cell receptor signaling by CD4+T cells cultured in the rotating wall-vessel bioreactor. J. Cell. Biochem. 2010, 109, 1201–1209. [Google Scholar] [CrossRef] [PubMed]
- Huse, M. The T-cell-receptor signaling network. J. Cell Sci. 2009, 122, 1269–1273. [Google Scholar] [CrossRef]
- Morrow, M.A. Clinorotation Differentially Inhibits T-lymphocyte transcription factor activation. Vitr. Cell. Dev. Biol. Anim. 2006, 42, 153. [Google Scholar] [CrossRef]
- JNK Signaling Pathway. Available online: https://www.creative-diagnostics.com/JNK-Signaling-Pathway.htm (accessed on 21 July 2021).
- Wen, A.Y.; Sakamoto, K.M.; Miller, L.S. The Role of the Transcription Factor CREB in Immune Function. J. Immunol. 2010, 185, 6413–6419. [Google Scholar] [CrossRef] [PubMed]
- Jain, J.; McCafffrey, P.G.; Miner, Z.; Kerppola, T.; Lambert, J.N.; Verdine, G.L.; Curran, T.; Rao, A. The T-cell transcription factor NFATp is a substrate for calcineurin and interacts with Fos and Jun. Nature 1993, 365, 352–355. [Google Scholar] [CrossRef]
- Rao, A.; Luo, C.; Hogan, P.G. Transcription factors of the nfat family: Regulation and Function. Annu. Rev. Immunol. 1997, 15, 707–747. [Google Scholar] [CrossRef] [PubMed]
- Boehm, U.; Klamp, T.; Groot, A.M.; Howard, J. Cellular responses to interferon-γ. Annu. Rev. Immunol. 1997, 15, 749–795. [Google Scholar] [CrossRef]
- Licato, L.L.A.; Grimm, E. Multiple interleukin-2 signaling pathways differentially regulated by microgravity. Immunopharmacology 1999, 44, 273–279. [Google Scholar] [CrossRef]
- Renehan, A.G.; Booth, C.; Potten, C.S. What is apoptosis, and why is it important? BMJ 2001, 322, 1536–1538. [Google Scholar] [CrossRef]
- Ekert, P.; Vaux, D. Apoptosis and the immune system. Br. Med. Bull. 1997, 53, 591–603. [Google Scholar] [CrossRef]
- Lewis, M.L.; Reynolds, J.; Cubano, L.A.; Hatton, J.P.; Lawless, B.D.; Piepmeier, E.H. Spaceflight alters microtubules and increases apoptosis in human lymphocytes (Jurkat). FASEB J. 1998, 12, 1007–1018. [Google Scholar] [CrossRef] [PubMed]
- Cubano, L.A.; Lewis, M.L. Fas/APO-1 protein is increased in spaceflown lymphocytes (Jurkat). Exp. Gerontol. 2000, 35, 389–400. [Google Scholar] [CrossRef]
- Sokolovskaya, A.; Korneeva, E.; Zaichenko, D.; Virus, E.; Kolesov, D.; Moskovtsev, A.; Kubatiev, A. Changes in the Surface Expression of Intercellular Adhesion Molecule 3, the Induction of Apoptosis, and the Inhibition of Cell-Cycle Progression of Human Multidrug-Resistant Jurkat/A4 Cells Exposed to a Random Positioning Machine. Int. J. Mol. Sci. 2020, 21, 855. [Google Scholar] [CrossRef] [PubMed]
- Schatten, H.; Lewis, M.L.; Chakrabarti, A. Spaceflight and clinorotation cause cytoskeleton and mitochondria changes and increases in apoptosis in cultured cells. Acta Astronaut. 2001, 49, 399–418. [Google Scholar] [CrossRef]
- Maccarrone, M.; Battista, N.; Meloni, M.; Bari, M.; Galleri, G.; Pippia, P.; Cogoli, A.; Finazzi-Agrò, A. Creating conditions similar to those that occur during exposure of cells to microgravity induces apoptosis in human lymphocytes by 5-lipoxygenase-mediated mitochondrial uncoupling and cytochrome c release. J. Leukoc. Biol. 2003, 73, 472–481. [Google Scholar] [CrossRef] [PubMed]
- Zhang, N.; Hartig, H.; Dzhagalov, I.; Draper, D.; He, Y.W. The role of apoptosis in the development and function of T lymphocytes. Cell Res. 2005, 15, 749–769. [Google Scholar] [CrossRef] [PubMed]
- Zhan, Y.; Carrington, E.M.; Zhang, Y.; Heinzel, S.; Lew, A. Life and Death of Activated T Cells: How Are They Different from Naïve T Cells? Front. Immunol. 2017, 8, 1809. [Google Scholar] [CrossRef]
- Risin, D.; Pellis, N.R. Modeled microgravity inhibits apoptosis in peripheral blood lymphocytes1. Vitr. Cell. Dev. Biol. Anim. 2001, 37, 66. [Google Scholar] [CrossRef]
- Kumari, R.; Singh, K.P.; DuMond, J.W., Jr. Simulated microgravity decreases DNA repair capacity and induces DNA damage in human lymphocytes. J. Cell. Biochem. 2009, 107, 723–731. [Google Scholar] [CrossRef] [PubMed]
- Nollen, E.A.A.; Morimoto, R.I. Chaperoning signaling pathways: Molecular chaperones as stress-sensing ‘heat shock’ proteins. J. Cell Sci. 2002, 115, 2809–2816. [Google Scholar] [CrossRef] [PubMed]
- Liu, C.S.C.; Raychaudhuri, D.; Paul, B.; Chakrabarty, Y.; Ghosh, A.R.; Rahaman, O.; Talukdar, A.; Ganguly, D. Cutting Edge: Piezo1 Mechanosensors Optimize Human T Cell Activation. J. Immunol. 2018, 200, 1255–1260. [Google Scholar] [CrossRef]




{kind=link}
{kind=link}
{kind=link}
| Pro-Inflammatory | Anti-Inflammatory | ||||||||||||||||
|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|
| Microgravity Exposure | Time | T Cell Source | IL-1B | IL-1 | IL-2 | IL-5 | IL-12 | IL-17/A | IL-22 | IFN-y | TNF-a | IL-4 | IL-6 | IL-8 | IL-10 | TGF-B1 | Study |
| Headdown Bedrest | 21 days | Human blood | ↡ | ↡ | ↡ | ↡ | [46] | ||||||||||
| 45 days | Human blood | / | ↡ | / | ↡ | / | / | / | ↑ | ↑ | [45] | ||||||
| 120 days | Human blood | ↟ | [44] | ||||||||||||||
| 2D Clinostat | 12 h | PBMC | ↑ | ↓ | [88] | ||||||||||||
| 48 h | PBMC | ↡ | [68] | ||||||||||||||
| 48 h | PBMC | ↡ | [68] | ||||||||||||||
| Rotary Wall Vessel | 3.5 h | Mice | ↡ | ↡ | [69] | ||||||||||||
| 18 h | PBMC | / | ↡ | [58] | |||||||||||||
| 24 h | Rats | ↡ | ↡ | [64] | |||||||||||||
| 24 h | Rats | ↡ | ↡ | [64] | |||||||||||||
| 48 h | PBMC | ↟ | [62] | ||||||||||||||
| 48 h | Jurkat T Cells | ↡ | ↟ | ↡ | / | / | [89] | ||||||||||
| 3 days | PBMC | ↑ | ↓ | ↓ | ↑ | [66] | |||||||||||
| 72 h | Rats | ↡ | [90] | ||||||||||||||
| 72 h | PBMC | ↓ | / | [57] | |||||||||||||
| 72 h | PBMC | ↡ | [27] | ||||||||||||||
| 72 h | Mice | ↡ | [61] | ||||||||||||||
| 120 h | OTII-TCH cells | ↓ | [91] | ||||||||||||||
| 3D Clinostat | 4 h | PBMC | ↡ | ↡ | ↡ | [67] | |||||||||||
| 12 h | PBMC | ↡ | [92] | ||||||||||||||
| 12 h | PBMC | ↑ | ↓ | [68] | |||||||||||||
| 24 h | PBMC | / | ↡ | ↟ | / | [71] | |||||||||||
| 26.5 h | Mice | ↡ | ↡ | [69] | |||||||||||||
| Parabolic Flight | 48 h | PBMC | / | / | / | / | [76] | ||||||||||
| Spaceflight | 6 h | PBMC | / | ↑ | [75] | ||||||||||||
| 14 h | Jurkat T Cells | ↓ | ↓ | [93] | |||||||||||||
| 72 h | PBMC | ↟ | ↡ | ↡ | / | [94] | |||||||||||
| 4 days | Rats | / | [95] | ||||||||||||||
| 9 days | Mice | ↟ | [96] | ||||||||||||||
| 10 days | Human blood | ↡ | [80] | ||||||||||||||
| 10 days | Mice | ↓ | [97] | ||||||||||||||
| 13 days | Mice | / | / | ↟ | / | [98] | |||||||||||
| 13 days 19 h | Mice | / | [99] | ||||||||||||||
| 14 days | Human blood | ↟ | ↡ | / | ↡ | ↡ | ↡ | ↡ | ↡ | [100] | |||||||
| 14 days | Human blood | / | ↡ | / | ↡ | ↡ | ↡ | ↡ | ↡ | [100] | |||||||
| 14 days | Human blood | / | / | / | / | ↟ | / | [100] | |||||||||
| 15 days | Mice | ↟ | ↡ | ↟ | ↑ | ↑ | / | ↑ | ↑ | [101] | |||||||
| 15 days | Mice | ↡ | ↡ | [69] | |||||||||||||
| 18 days | Human blood | ↡ | ↡ | [102] | |||||||||||||
| 30 days | Mice | ↡ | / | ↡ | [85] | ||||||||||||
| 175 days | Human blood | / | / | [103] | |||||||||||||
| 180 days | Human blood | / | / | ↟ | [104] | ||||||||||||
| 6 months | Human blood | / | ↡ | ↡ | / | / | ↡ | / | ↡ | [84] | |||||||
| 6 months | Human blood | ↓ | ↡ | ↡ | ↡ | ↡ | ↡ | ↡ | ↡ | [84] | |||||||
| 6 months | Human blood | ↓ | / | / | ↑ | ↓ | [84] | ||||||||||
| 11 months | Human blood | / | / | [105] | |||||||||||||
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Dhar, S.; Kaeley, D.K.; Kanan, M.J.; Yildirim-Ayan, E. Mechano-Immunomodulation in Space: Mechanisms Involving Microgravity-Induced Changes in T Cells. Life 2021, 11, 1043. https://doi.org/10.3390/life11101043
Dhar S, Kaeley DK, Kanan MJ, Yildirim-Ayan E. Mechano-Immunomodulation in Space: Mechanisms Involving Microgravity-Induced Changes in T Cells. Life. 2021; 11(10):1043. https://doi.org/10.3390/life11101043
Chicago/Turabian StyleDhar, Sarit, Dilpreet Kaur Kaeley, Mohamad Jalal Kanan, and Eda Yildirim-Ayan. 2021. "Mechano-Immunomodulation in Space: Mechanisms Involving Microgravity-Induced Changes in T Cells" Life 11, no. 10: 1043. https://doi.org/10.3390/life11101043
APA StyleDhar, S., Kaeley, D. K., Kanan, M. J., & Yildirim-Ayan, E. (2021). Mechano-Immunomodulation in Space: Mechanisms Involving Microgravity-Induced Changes in T Cells. Life, 11(10), 1043. https://doi.org/10.3390/life11101043