The Emerging Role of BDNF/TrkB Signaling in Cardiovascular Diseases

 ,
, 
 and
and
Abstract
1. Introduction
2. BDNF/TrkB-Related Signaling Pathways
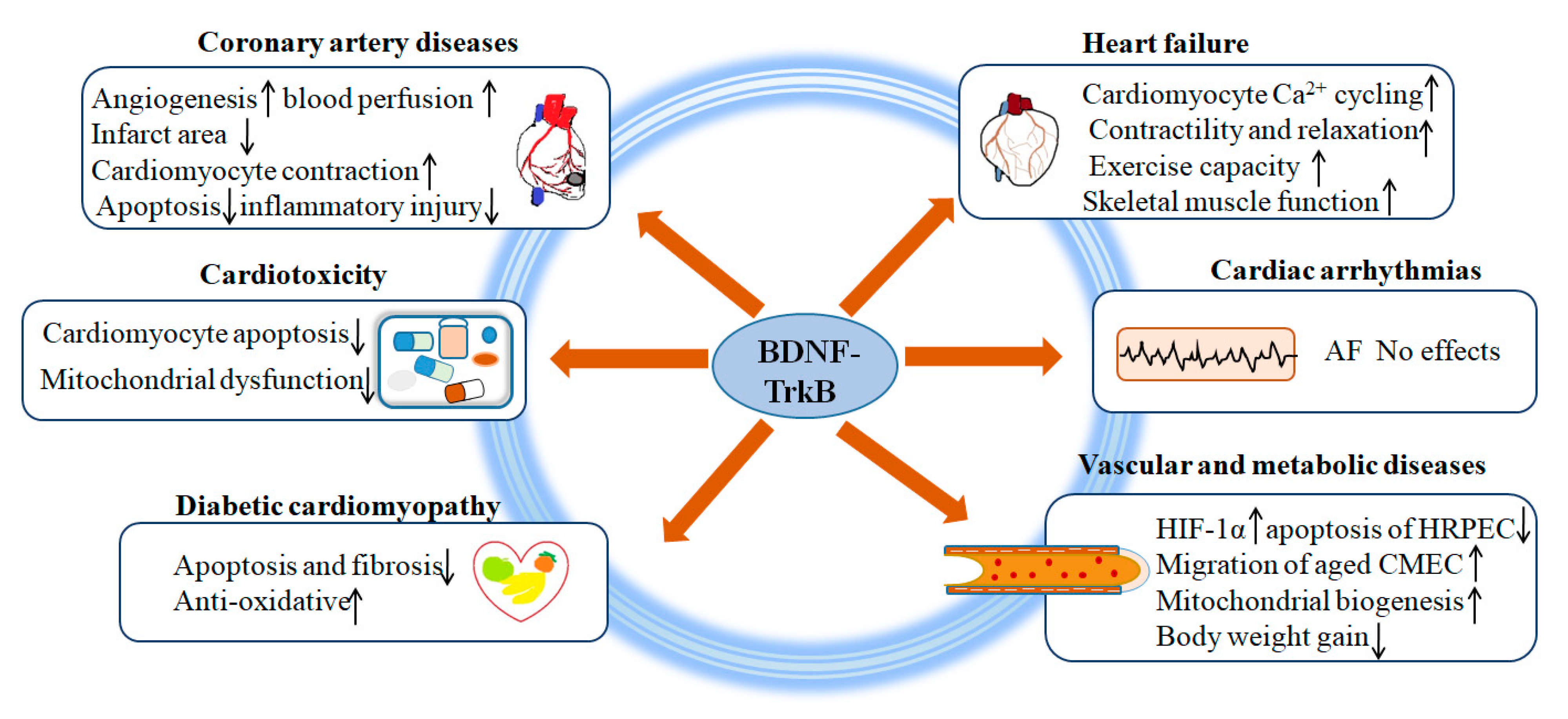
3. Role of BDNF/TrkB in Coronary Artery Diseases
4. Role of BDNF/TrkB in Heart Failure (HF)
5. Role of BDNF/TrkB in Cardiotoxicity
6. Role of BDNF/TrkB in Diabetes
7. Role of BDNF/TrkB in Cardiac Arrhythmias
8. Role of BDNF/TrkB in Vascular and Metabolic Diseases
9. Role of BDNF/TrkB in Noncardiac Cells
10. Role of Circulating BDNF in CVD
11. Concluding Remarks
Author Contributions
Funding
Conflicts of Interest
Abbreviations
| BDNF | brain-derived neurotrophic factor |
| CVD | cardiovascular diseases |
| DOX | doxorubicin |
| 7,8-DHF | 7,8-dihydroxyflavone |
| ERK | extracellular regulated protein kinase |
| HF | heart failure |
| MAPK | mitogen-activated protein kinase |
| MI | myocardial infarction |
| NF-κB | nuclear factor κB |
| NGF | neuronal growth factor |
| NT | neurotrophin |
| TrkB | tropomyosin-related kinase receptor B |
References
- Chao, M.V.; Rajagopal, R.; Lee, F.S. Neurotrophin signalling in health and disease. Clin. Sci. 2006, 110, 167–173. [Google Scholar] [CrossRef] [PubMed]
- Barde, Y.A.; Edgar, D.; Thoenen, H. Purification of a new neurotrophic factor from mammalian brain. EMBO J. 1982, 1, 549–553. [Google Scholar] [CrossRef] [PubMed]
- Scarisbrick, I.A.; Jones, E.G.; Isackson, P.J. Coexpression of mRNAs for NGF, BDNF, and NT-3 in the cardiovascular system of the pre- and postnatal rat. J. Neurosci. 1993, 13, 875–893. [Google Scholar] [CrossRef] [PubMed]
- Numakawa, T.; Suzuki, S.; Kumamaru, E.; Adachi, N.; Richards, M.; Kunugi, H. BDNF function and intracellular signaling in neurons. Histol. Histopathol. 2010, 25, 237–258. [Google Scholar] [PubMed]
- László, A.; Lénárt, L.; Illésy, L.; Fekete, A.; Nemcsik, J. The role of neurotrophins in psychopathology and cardiovascular diseases: Psychosomatic connections. J. Neural Transm. 2019, 126, 265–278. [Google Scholar] [CrossRef]
- Gupta, V.; You, Y.; Gupta, V.B.; Klistorner, A.; Graham, S.L. TrkB Receptor Signalling: Implications in Neurodegenerative, Psychiatric and Proliferative Disorders. Int. J. Mol. Sci. 2013, 14, 10122–10142. [Google Scholar] [CrossRef]
- Halievski, K.; Xu, Y.; Haddad, Y.W.; Tang, Y.P.; Yamada, S.; Katsuno, M.; Adachi, H.; Sobue, G.; Breedlove, S.M.; Jordan, C.L. Muscle BDNF improves synaptic and contractile muscle strength in Kennedy’s disease mice in a muscle-type specific manner. J. Physiol. 2020, 598, 2719–2739. [Google Scholar] [CrossRef]
- Gupta, V.K.; You, Y.; Li, J.C.; Klistorner, A.; Graham, S.L. Protective Effects of 7,8-Dihydroxyflavone on Retinal Ganglion and RGC-5 Cells Against Excitotoxic and Oxidative Stress. J. Mol. Neurosci. 2013, 49, 96–104. [Google Scholar] [CrossRef]
- Kang, J.S.; Choi, I.-W.; Han, M.H.; Kim, G.-Y.; Hong, S.H.; Park, C.; Hwang, H.J.; Kim, C.M.; Kim, B.W.; Choi, Y.H. The cytoprotective effects of 7,8-dihydroxyflavone against oxidative stress are mediated by the upregulation of Nrf2-dependent HO-1 expression through the activation of the PI3K/Akt and ERK pathways in C2C12 myoblasts. Int. J. Mol. Med. 2015, 36, 501–510. [Google Scholar] [CrossRef]
- Huang, H.-M.; Huang, C.-C.; Tsai, M.-H.; Poon, L.Y.-C.; Chang, Y.-C. Systemic 7,8-Dihydroxyflavone Treatment Protects Immature Retinas Against Hypoxic-Ischemic Injury via Müller Glia Regeneration and MAPK/ERK Activation. Investig. Opthalmol. Vis. Sci. 2018, 59, 3124–3135. [Google Scholar] [CrossRef]
- Tsai, T.; Klausmeyer, A.; Conrad, R.; Gottschling, C.; Leo, M.; Faissner, A.; Wiese, S. 7,8-Dihydroxyflavone leads to survival of cultured embryonic motoneurons by activating intracellular signaling pathways. Mol. Cell. Neurosci. 2013, 56, 18–28. [Google Scholar] [CrossRef] [PubMed]
- Park, H.Y.; Kim, G.Y.; Hyun, J.W.; Hwang, H.J.; Kim, N.D.; Kim, B.W.; Choi, Y.H. 7,8-Dihydroxyflavone exhibits anti-inflammatory properties by downregulating the NF-kappaB and MAPK signaling pathways in lipopolysaccharide-treated RAW264.7 cells. Int. J. Mol. Med. 2012, 29, 1146–1152. [Google Scholar] [PubMed]
- Han, X.; Cheng, M.-N.; Chen, L.; Fang, H.; Wang, L.-J.; Li, X.-T.; Qu, Z.-Q. 7,8-Dihydroxyflavone protects PC12 cells against 6-hydroxydopamine-induced cell death through modulating PI3K/Akt and JNK pathways. Neurosci. Lett. 2014, 581, 85–88. [Google Scholar] [CrossRef] [PubMed]
- Zhao, J.; Du, J.; Pan, Y.; Chen, T.; Zhao, L.; Zhu, Y.; Chen, Y.; Zheng, Y.; Liu, Y.; Sun, L.; et al. Activation of cardiac TrkB receptor by its small molecular agonist 7,8-dihydroxyflavone inhibits doxorubicin-induced cardiotoxicity via enhancing mitochondrial oxidative phosphorylation. Free Radic. Biol. Med. 2019, 130, 557–567. [Google Scholar] [CrossRef] [PubMed]
- Wang, Z.; Wang, S.-P.; Shao, Q.; Li, P.-F.; Sun, Y.; Luo, L.-Z.; Yan, X.-Q.; Fan, Z.-Y.; Hu, J.; Zhao, J.; et al. Brain-derived neurotrophic factor mimetic, 7,8-dihydroxyflavone, protects against myocardial ischemia by rebalancing optic atrophy 1 processing. Free Radic. Biol. Med. 2019, 145, 187–197. [Google Scholar] [CrossRef]
- Liu, Y.; Sun, L.; Huan, Y.; Zhao, H.; Deng, J. Application of bFGF and BDNF to Improve Angiogenesis and Cardiac Function. J. Surg. Res. 2006, 136, 85–91. [Google Scholar] [CrossRef]
- Schäbitz, W.R.; Steigleder, T.; Cooper-Kuhn, C.M.; Schwab, S.; Sommer, C.; Schneider, A.; Kuhn, H.G. Intravenous Brain-Derived Neurotrophic Factor Enhances Poststroke Sensorimotor Recovery and Stimulates Neurogenesis. Stroke 2007, 38, 2165–2172. [Google Scholar] [CrossRef]
- Hiltunen, J.O.; Laurikainen, A.; Meri, S.; Saarma, M. Nerve growth factor and brain-derived neurotrophic factor mRNAs are regulated in distinct cell populations of rat heart after ischaemia and reperfusion. J. Pathol. 2001, 194, 247–253. [Google Scholar] [CrossRef]
- Ejiri, J.; Inoue, N.; Kobayashi, S.; Shiraki, R.; Otsui, K.; Honjo, T.; Takahashi, M.; Ohashi, Y.; Ichikawa, S.; Terashima, M.; et al. Possible Role of Brain-Derived Neurotrophic Factor in the Pathogenesis of Coronary Artery Disease. Circulation 2005, 112, 2114–2120. [Google Scholar] [CrossRef]
- Okada, S.; Yokoyama, M.; Toko, H.; Tateno, K.; Moriya, J.; Shimizu, I.; Nojima, A.; Ito, T.; Yoshida, Y.; Kobayashi, Y.; et al. Brain-Derived Neurotrophic Factor Protects Against Cardiac Dysfunction After Myocardial Infarction via a Central Nervous System–Mediated Pathway. Arter. Thromb. Vasc. Biol. 2012, 32, 1902–1909. [Google Scholar] [CrossRef]
- Bai, H.-L.; Lu, Z.-F.; Zhao, J.-J.; Ma, X.; Li, X.-H.; Xu, H.; Wu, S.-G.; Kang, C.-M.; Lu, J.-B.; Xu, Y.-J.; et al. Microarray profiling analysis and validation of novel long noncoding RNAs and mRNAs as potential biomarkers and their functions in atherosclerosis. Physiol. Genom. 2019, 51, 644–656. [Google Scholar] [CrossRef] [PubMed]
- Feng, N.; Huke, S.; Zhu, G.; Tocchetti, C.G.; Shi, S.; Aiba, T.; Kaludercic, N.; Hoover, D.B.; Beck, S.E.; Mankowski, J.L.; et al. Constitutive BDNF/TrkB signaling is required for normal cardiac contraction and relaxation. Proc. Natl. Acad. Sci. USA 2015, 112, 1880–1885. [Google Scholar] [CrossRef] [PubMed]
- Fulgenzi, G.; Tomassoni-Ardori, F.; Babini, L.; Becker, J.; Barrick, C.; Puverel, S.; Tessarollo, L. BDNF modulates heart contraction force and long-term homeostasis through truncated TrkB.T1 receptor activation. J. Cell Biol. 2015, 210, 1003–1012. [Google Scholar] [CrossRef] [PubMed]
- Hang, P.; Zhao, J.; Cai, B.; Tian, S.; Huang, W.; Guo, J.; Sun, C.; Li, Y.; Du, Z. Brain-derived neurotrophic factor regulates TRPC3/6 channels and protects against myocardial infarction in rodents. Int. J. Biol. Sci. 2015, 11, 536–545. [Google Scholar] [CrossRef] [PubMed]
- Cao, L.; Zhang, L.; Chen, S.; Yuan, Z.; Liu, S.; Shen, X.; Zheng, X.; Qi, X.; Lee, K.K.; Chan, J.Y.-H.; et al. BDNF-mediated migration of cardiac microvascular endothelial cells is impaired during ageing. J. Cell. Mol. Med. 2012, 16, 3105–3115. [Google Scholar] [CrossRef]
- Halade, G.V.; Ma, Y.; Ramirez, T.A.; Zhang, J.; Dai, Q.; Hensler, J.G.; Lopez, E.F.; Ghasemi, O.; Jin, Y.-F.; Lindsey, M.L. Reduced BDNF attenuates inflammation and angiogenesis to improve survival and cardiac function following myocardial infarction in mice. Am. J. Physiol. Circ. Physiol. 2013, 305, H1830–H1842. [Google Scholar] [CrossRef]
- Cai, D.; Holm, J.M.; Duignan, I.J.; Zheng, J.; Xaymardan, M.; Chin, A.; Ballard, V.L.T.; Bella, J.N.; Edelberg, J.M. BDNF-mediated enhancement of inflammation and injury in the aging heart. Physiol. Genom. 2006, 24, 191–197. [Google Scholar] [CrossRef]
- Zhang, Z.; Wang, B.; Fei, A. BDNF contributes to the skeletal muscle anti-atrophic effect of exercise training through AMPK-PGC1α signaling in heart failure mice. Arch. Med. Sci. 2018, 15, 214–222. [Google Scholar] [CrossRef]
- Lee, H.W.; Ahmad, M.; Wang, H.-W.; Leenen, F.H.H. Effects of exercise training on brain-derived neurotrophic factor in skeletal muscle and heart of rats post myocardial infarction. Exp. Physiol. 2017, 102, 314–328. [Google Scholar] [CrossRef]
- Matsumoto, J.; Takada, S.; Kinugawa, S.; Furihata, T.; Nambu, H.; Kakutani, N.; Tsuda, M.; Fukushima, A.; Yokota, T.; Tanaka, S.; et al. Brain-Derived Neurotrophic Factor Improves Limited Exercise Capacity in Mice With Heart Failure. Circulation 2018, 138, 2064–2066. [Google Scholar] [CrossRef]
- Matsumoto, J.; Takada, S.; Furihata, T.; Nambu, H.; Kakutani, N.; Maekawa, S.; Mizushima, W.; Nakano, I.; Fukushima, A.; Yokota, T.; et al. Brain-Derived Neurotrophic Factor Improves Impaired Fatty Acid Oxidation Via the Activation of Adenosine Monophosphate-activated Protein Kinase-α—Proliferator-Activated Receptor-r Coactivator-1α Signaling in Skeletal Muscle of Mice With Heart Failure. Circ. Heart Fail. 2020. [Google Scholar] [CrossRef] [PubMed]
- Nakano, I.; Kinugawa, S.; Hori, H.; Fukushima, A.; Yokota, T.; Takada, S.; Kakutani, N.; Obata, Y.; Yamanashi, K.; Anzai, T. Serum Brain-Derived Neurotrophic Factor Levels Are Associated with Skeletal Muscle Function but Not with Muscle Mass in Patients with Heart Failure. Int. Heart J. 2020, 61, 96–102. [Google Scholar] [CrossRef]
- Lee, H.W.; Ahmad, M.; Wang, H.W.; Leenen, F.H.H. Effects of exercise on BDNF-TrkB signaling in the paraventricular nucleus and rostral ventrolateral medulla in rats post myocardial infarction. Neuropeptides 2020, 82, 102058. [Google Scholar] [CrossRef] [PubMed]
- Hang, P.; Zhao, J.; Sun, L.; Li, M.; Han, Y.; Du, Z.; Li, Y. Brain-derived neurotrophic factor attenuates doxorubicin-induced cardiac dysfunction through activating Akt signalling in rats. J. Cell. Mol. Med. 2016, 21, 685–696. [Google Scholar] [CrossRef] [PubMed]
- Bathina, S.; Srinivas, N.; Das, U.N. BDNF protects pancreatic beta cells (RIN5F) against cytotoxic action of alloxan, streptozotocin, doxorubicin and benzo(a)pyrene in vitro. Metab. Clin. Exp. 2016, 65, 667–684. [Google Scholar] [CrossRef]
- Liao, D.; Zhang, C.; Liu, N.; Cao, L.; Wang, C.; Feng, Q.; Yao, D.; Long, M.; Jiang, P. Involvement of neurotrophic signaling in doxorubicin‑induced cardiotoxicity. Exp. Ther. Med. 2019, 19, 1129–1135. [Google Scholar] [CrossRef]
- Zhao, X.-Y.; Li, G.-Y.; Liu, Y.; Chai, L.-M.; Chen, J.-X.; Zhang, Y.; Du, Z.-M.; Lu, Y.-J.; Yang, B. Resveratrol protects against arsenic trioxide-induced cardiotoxicity in vitro and in vivo. Br. J. Pharmacol. 2008, 154, 105–113. [Google Scholar] [CrossRef]
- Zhao, X.; Feng, T.; Chen, H.; Shan, H.; Zhang, Y.; Lu, Y.; Yang, B. Arsenic Trioxide-Induced Apoptosis in H9c2 Cardiomyocytes: Implications in Cardiotoxicity. Basic Clin. Pharmacol. Toxicol. 2008, 102, 419–425. [Google Scholar] [CrossRef]
- Chan, C.B.; Ahuja, P.; Ye, K. Developing Insulin and BDNF Mimetics for Diabetes Therapy. Curr. Top. Med. Chem. 2019, 19, 2188–2204. [Google Scholar] [CrossRef]
- Sefidgari-Abrasi, S.; Roshangar, L.; Karimi, P.; Morshedi, M.; Rahimiyan-Heravan, M.; Saghafi-Asl, M. From the gut to the heart: L. plantarum and inulin administration as a novel approach to control cardiac apoptosis via 5-HT2B and TrkB receptors in diabetes. Clin. Nutr. 2021, 40, 190–201. [Google Scholar] [CrossRef]
- Gong, J.; Zhou, F.; Wang, S.X.X.; Xu, J.; Xiao, F. Caveolin-3 protects diabetic hearts from acute myocardial infarction/reperfusion injury through beta2AR, cAMP/PKA, and BDNF/TrkB signaling pathways. Aging 2020, 12, 14300–14313. [Google Scholar] [CrossRef] [PubMed]
- Xu, B.; Xie, X. Neurotrophic factor control of satiety and body weight. Nat. Rev. Neurosci. 2016, 17, 282–292. [Google Scholar] [CrossRef] [PubMed]
- Rios, M. Neurotrophins and the Regulation of Energy Balance and Body Weight. Lipid Signal. Hum. Dis. 2014, 220, 283–307. [Google Scholar] [CrossRef]
- Nakagawa, T.; Ono-Kishino, M.; Sugaru, E.; Yamanaka, M.; Taiji, M.; Noguchi, H. Brain-derived neurotrophic factor (BDNF) regulates glucose and energy metabolism in diabetic mice. Diabetes/Metab. Res. Rev. 2002, 18, 185–191. [Google Scholar] [CrossRef]
- Yamanaka, M.; Tsuchida, A.; Nakagawa, T.; Nonomura, T.; Ono-Kishino, M.; Sugaru, E.; Noguchi, H.; Taiji, M. Brain-derived neurotrophic factor enhances glucose utilization in peripheral tissues of diabetic mice. Diabetes Obes. Metab. 2007, 9, 59–64. [Google Scholar] [CrossRef]
- Rahman, F.; Himali, J.J.; Yin, X.; Beiser, A.S.; Ellinor, P.T.; Lubitz, S.A.; Vasan, R.S.; Magnani, J.W.; McManus, D.D.; Seshadri, S.; et al. Serum brain-derived neurotrophic factor and risk of atrial fibrillation. Am. Heart J. 2016, 183, 69–73. [Google Scholar] [CrossRef]
- Huai, R.; Han, X.; Wang, B.; Li, C.; Niu, Y.; Li, R.; Qu, Z. Vasorelaxing and Antihypertensive Effects of 7,8-Dihydroxyflavone. Am. J. Hypertens. 2013, 27, 750–760. [Google Scholar] [CrossRef]
- Helan, M.; Aravamudan, B.; Hartman, W.R.; Thompson, M.A.; Johnson, B.D.; Pabelick, C.M.; Prakash, Y.S. BDNF secretion by human pulmonary artery endothelial cells in response to hypoxia. J. Mol. Cell. Cardiol. 2014, 68, 89–97. [Google Scholar] [CrossRef]
- Wood, J.; Tse, M.C.L.; Yang, X.; Brobst, D.; Liu, Z.; Pang, B.P.S.; Chan, W.S.; Zaw, A.M.; Chow, B.K.; Ye, K.; et al. BDNF mimetic alleviates body weight gain in obese mice by enhancing mitochondrial biogenesis in skeletal muscle. Metab. Clin. Exp. 2018, 87, 113–122. [Google Scholar] [CrossRef]
- Chan, C.B.; Tse, M.C.L.; Liu, X.; Zhang, S.; Schmidt, R.; Otten, R.; Liu, L.; Ye, K. Activation of Muscular TrkB by its Small Molecular Agonist 7,8-Dihydroxyflavone Sex-Dependently Regulates Energy Metabolism in Diet-Induced Obese Mice. Chem. Biol. 2015, 22, 355–368. [Google Scholar] [CrossRef]
- Allen, R.S.; Hanif, A.M.; Gogniat, M.A.; Prall, B.C.; Haider, R.; Aung, M.H.; Prunty, M.C.; Mees, L.M.; Coulter, M.M.; Motz, C.T.; et al. TrkB signalling pathway mediates the protective effects of exercise in the diabetic rat retina. Eur. J. Neurosci. 2018, 47, 1254–1265. [Google Scholar] [CrossRef] [PubMed]
- Agrimi, J.; Spalletti, C.; Baroni, C.; Keceli, G.; Zhu, G.; Caragnano, A.; Matteucci, M.; Chelko, S.P.; Ramirez-Correa, G.A.; Bedja, D.; et al. Obese mice exposed to psychosocial stress display cardiac and hippocampal dysfunction associated with local brain-derived neurotrophic factor depletion. EBioMedicine 2019, 47, 384–401. [Google Scholar] [CrossRef] [PubMed]
- Cho, S.J.; Kang, K.A.; Piao, M.J.; Ryu, Y.S.; Fernando, P.D.S.M.; Zhen, A.X.; Hyun, Y.J.; Ahn, M.J.; Kang, H.K.; Hyun, J.W. 7,8-Dihydroxyflavone Protects High Glucose-Damaged Neuronal Cells against Oxidative Stress. Biomol. Ther. 2019, 27, 85–91. [Google Scholar] [CrossRef] [PubMed]
- Yu, X.; Liu, Q.; Wang, X.; Liu, H.; Wang, Y. 7,8-Dihydroxyflavone ameliorates high-glucose induced diabetic apoptosis in human retinal pigment epithelial cells by activating TrkB. Biochem. Biophys. Res. Commun. 2018, 495, 922–927. [Google Scholar] [CrossRef] [PubMed]
- Wang, Z.; Chen, Y.; Chen, X.; Zheng, X.; Xu, G.; Yuan, Z.; Zhao, H.; Chen, W.; Li, L.; Zheng, N.; et al. The TrkB-T1 receptor mediates BDNF-induced migration of aged cardiac microvascular endothelial cells by recruiting Willin. Aging Cell 2019, 18, e12881. [Google Scholar] [CrossRef] [PubMed]
- Cherubini, E.; Mariotta, S.; Scozzi, D.; Mancini, R.; Osman, G.; D’Ascanio, M.; Bruno, P.; Cardillo, G.; Ricci, A. BDNF/TrkB axis activation promotes epithelial–mesenchymal transition in idiopathic pulmonary fibrosis. J. Transl. Med. 2017, 15, 196. [Google Scholar] [CrossRef] [PubMed]
- Freeman, M.R.; Sathish, V.; Manlove, L.; Wang, S.; Britt, R.D.; Thompson, M.A.; Pabelick, C.M.; Prakash, Y.S. Brain-derived neurotrophic factor and airway fibrosis in asthma. Am. J. Physiol. Cell. Mol. Physiol. 2017, 313, L360–L370. [Google Scholar] [CrossRef]
- Choi, J.W.; Lee, J.; Park, Y.I. 7,8-Dihydroxyflavone attenuates TNF-α-induced skin aging in Hs68 human dermal fibroblast cells via down-regulation of the MAPKs/Akt signaling pathways. Biomed. Pharmacother. 2017, 95, 1580–1587. [Google Scholar] [CrossRef]
- He, B.; Qu, Z.; Tian, Z.; Zhao, K.; Wei, L.; Ma, L. 7,8-dihydroxyflavone enhanced cholinergic contraction of rat gastric smooth muscle via augmenting muscarinic M3 receptor expression. Clin. Exp. Pharmacol. Physiol. 2018, 45, 1170–1180. [Google Scholar] [CrossRef]
- Monisha, K.G.; Prabu, P.; Chokkalingam, M.; Murugesan, R.; Milenkovic, D.; Ahmed, S.S. Clinical utility of brain-derived neurotrophic factor as a biomarker with left ventricular echocardiographic indices for potential diagnosis of coronary artery disease. Sci. Rep. 2020, 10, 1–8. [Google Scholar] [CrossRef]
- Kaess, B.M.; Preis, S.R.; Lieb, W.; Beiser, A.S.; Yang, Q.; Chen, T.C.; Hengstenberg, C.; Erdmann, J.; Schunkert, H.; Seshadri, S.; et al. Circulating brain-derived neurotrophic factor concentrations and the risk of cardiovascular disease in the community. J. Am. Heart Assoc. 2015, 4, e001544. [Google Scholar] [CrossRef] [PubMed]
- Takashio, S.; Sugiyama, S.; Yamamuro, M.; Takahama, H.; Hayashi, T.; Sugano, Y.; Izumiya, Y.; Hokimoto, S.; Minamino, N.; Yasuda, S.; et al. Significance of Low Plasma Levels of Brain-Derived Neurotrophic Factor in Patients With Heart Failure. Am. J. Cardiol. 2015, 116, 243–249. [Google Scholar] [CrossRef] [PubMed]
- Fukushima, A.; Kinugawa, S.; Homma, T.; Masaki, Y.; Furihata, T.; Yokota, T.; Matsushima, S.; Takada, S.; Kadoguchi, T.; Oba, K.; et al. Serum Brain-Derived Neurotropic Factor Level Predicts Adverse Clinical Outcomes in Patients With Heart Failure. J. Card. Fail. 2015, 21, 300–306. [Google Scholar] [CrossRef] [PubMed]
- Kadowaki, S.; Shishido, T.; Honda, Y.; Narumi, T.; Otaki, Y.; Kinoshita, D.; Nishiyama, S.; Takahashi, H.; Arimoto, T.; Miyamoto, T.; et al. Additive clinical value of serum brain-derived neurotrophic factor for prediction of chronic heart failure outcome. Heart Vessel. 2015, 31, 535–544. [Google Scholar] [CrossRef] [PubMed]
- Shibata, A.; Hanatani, A.; Izumi, Y.; Kitada, R.; Iwata, S.; Yoshiyama, M. Serum brain-derived neurotrophic factor level and exercise tolerance complement each other in predicting the prognosis of patients with heart failure. Heart Vessel. 2018, 33, 1325–1333. [Google Scholar] [CrossRef]
- Fukushima, A.; Kinugawa, S.; Homma, T.; Masaki, Y.; Furihata, T.; Yokota, T.; Matsushima, S.; Abe, T.; Suga, T.; Takada, S.; et al. Decreased serum brain-derived neurotrophic factor levels are correlated with exercise intolerance in patients with heart failure. Int. J. Cardiol. 2013, 168, e142–e144. [Google Scholar] [CrossRef]
- Belviranlı, M.; Okudan, N. Exercise training increases cardiac, hepatic and circulating levels of brain-derived neurotrophic factor and irisin in young and aged rats. Horm. Mol. Biol. Clin. Investig. 2018, 36, 3. [Google Scholar] [CrossRef]
- Han, W.; Zhang, C.; Wang, H.; Yang, M.; Guo, Y.; Li, G.; Zhang, H.; Wang, C.; Chen, D.; Geng, C.; et al. Alterations of irisin, adropin, preptin and BDNF concentrations in coronary heart disease patients comorbid with depression. Ann. Transl. Med. 2019, 7, 298. [Google Scholar] [CrossRef]
- Prabu, P.; Poongothai, S.; Shanthirani, C.S.; Anjana, R.M.; Mohan, V.; Balasubramanyam, M. Altered circulatory levels of miR-128, BDNF, cortisol and shortened telomeres in patients with type 2 diabetes and depression. Acta Diabetol. 2020, 57, 799–807. [Google Scholar] [CrossRef]
- Pollock, K.; Dahlenburg, H.; Nelson, H.; Fink, K.D.; Cary, W.; Hendrix, K.; Annett, G.; Torrest, A.; Deng, P.; Gutierrez, J.; et al. Human Mesenchymal Stem Cells Genetically Engineered to Overexpress Brain-derived Neurotrophic Factor Improve Outcomes in Huntington’s Disease Mouse Models. Mol. Ther. J. Am. Soc. Gene Ther. 2016, 24, 965–977. [Google Scholar] [CrossRef]
- Li, Y.; Xu, J.; Xu, P.; Song, S.; Liu, P.; Chi, T.; Ji, X.; Jin, G.; Qiu, S.; Hou, Y.; et al. Xanthoceras sorbifolia extracts ameliorate dendritic spine deficiency and cognitive decline via upregulation of BDNF expression in a rat model of Alzheimer’s disease. Neurosci. Lett. 2016, 629, 208–214. [Google Scholar] [CrossRef] [PubMed]
- Jiang, B.; Wang, Y.-J.; Wang, H.; Song, L.; Huang, C.; Zhu, Q.; Wu, F.; Zhang, W. Antidepressant-like effects of fenofibrate in mice via the hippocampal brain-derived neurotrophic factor signalling pathway. Br. J. Pharmacol. 2016, 174, 177–194. [Google Scholar] [CrossRef] [PubMed]
- Zhang, X.Y.; da Chen, C.; Tan, Y.L.; Tan, S.; Luo, X.; Zuo, L.; Soares, J.C. BDNF Polymorphisms Are Associated With Cognitive Performance in Schizophrenia Patients Versus Healthy Controls. J. Clin. Psychiatry 2016, 77, 1011–1018. [Google Scholar] [CrossRef] [PubMed]
- Wu, Y.; Si, R.; Yang, S.; Xia, S.; He, Z.; Wang, L.; He, Z.; Wang, Q.; Tang, H. Depression induces poor prognosis associates with the down-regulation brain derived neurotrophic factor of serum in advanced small cell lung cancer. Oncotarget 2016, 7, 85975–85986. [Google Scholar] [CrossRef][Green Version]
- Wu, Y.; Luo, X.; Liu, X.; Liu, D.; Wang, X.; Guo, Z.; Zhu, L.; Tian, Q.; Yang, X.; Wang, J.-Z. Intraperitoneal Administration of a Novel TAT-BDNF Peptide Ameliorates Cognitive Impairments via Modulating Multiple Pathways in Two Alzheimer’s Rodent Models. Sci. Rep. 2015, 5, 15032. [Google Scholar] [CrossRef]
- Chen, C.; Wang, Z.; Zhang, Z.; Liu, X.; Kang, S.S.; Zhang, Y.; Ye, K. The prodrug of 7,8-dihydroxyflavone development and therapeutic efficacy for treating Alzheimer’s disease. Proc. Natl. Acad. Sci. USA 2018, 115, 578–583. [Google Scholar] [CrossRef]
- Bathina, S.; Das, U.N. Brain-derived neurotrophic factor and its clinical implications. Arch. Med. Sci. 2015, 6, 1164–1178. [Google Scholar] [CrossRef]
- Sanada, K.; Zorrilla, I.; Iwata, Y.; Bermúdez-Ampudia, C.; Graff-Guerrero, A.; Martínez-Cengotitabengoa, M.; González-Pinto, A. The Efficacy of Non-Pharmacological Interventions on Brain-Derived Neurotrophic Factor in Schizophrenia: A Systematic Review and Meta-Analysis. Int. J. Mol. Sci. 2016, 17, 1766. [Google Scholar] [CrossRef]


{kind=link}
| Diseases | Source | Description | References |
|---|---|---|---|
| Unstable angina | Plasma | Higher in unstable angina compared with the stable effort angina and noncoronary artery disease groups | Ejiri J et al. 2005 [19] |
| Coronary artery disease | Serum | BDNF was significantly lower in patients with coronary artery disease than control | Monisha K et al. 2020 [60] |
| Myocardial ischemia | Plasma | BDNF was markedly increased after myocardial ischemia | Okada S et al. 2012 [20] |
| Cardiovascular diseases | Serum | Higher BDNF level is associated with a decreased risk of cardiovascular diseases and mortality | Kaess B et al. 2015 [61] |
| Depression in coronary heart disease | Plasma | Plasma BDNF was associated with occurrence of depression in coronary heart disease | Han W et al. 2019 [68] |
| Heart failure | Plasma | Low BDNF level is associated with the severity of heart failure | Takashio S et al. 2015 [62] |
| Heart failure | Serum | BDNF predicts adverse clinical outcomes in patients with heart failure | Fukushima A et al. 2015 [63] |
| Heart failure | Serum | Low BDNF level is independently associated with an increased risk of cardiac events | Kadowaki S et al. 2015 [64] |
| Heart failure | Serum | BDNF level at discharge may be a useful biomarker of the prognosis in patients with heart failure | Shibata A et al. 2018 [65] |
| Heart failure | Serum | Decreased BDNF is correlated with exercise intolerance in patients with heart failure | Fukushima A et al. 2013 [66] |
| Heart failure | Serum | BDNF levels were associated with exercise capacity and skeletal muscle function, but not with muscle mass | Nakano I et al. 2020 [32] |
| Atrial fibrillation | Serum | No statistically significant association was found between serum BDNF levels and risk of incident atrial fibrillation | Rahman F et al. 2016 [46] |
| Aging | Plasma | Regular exercise improves aging-induced decrease in the cardiac, hepatic, and plasma BDNF | Belviranli M et al. 2018 [67] |
| T2DM with depression | Serum | Decreased levels of circulatory BDNF in patients with type 2 diabetes and depression | Prabu P et al. 2020 [69] |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Hang, P.-Z.; Zhu, H.; Li, P.-F.; Liu, J.; Ge, F.-Q.; Zhao, J.; Du, Z.-M. The Emerging Role of BDNF/TrkB Signaling in Cardiovascular Diseases. Life 2021, 11, 70. https://doi.org/10.3390/life11010070
Hang P-Z, Zhu H, Li P-F, Liu J, Ge F-Q, Zhao J, Du Z-M. The Emerging Role of BDNF/TrkB Signaling in Cardiovascular Diseases. Life. 2021; 11(1):70. https://doi.org/10.3390/life11010070
Chicago/Turabian StyleHang, Peng-Zhou, Hua Zhu, Pei-Feng Li, Jie Liu, Feng-Qin Ge, Jing Zhao, and Zhi-Min Du. 2021. "The Emerging Role of BDNF/TrkB Signaling in Cardiovascular Diseases" Life 11, no. 1: 70. https://doi.org/10.3390/life11010070
APA StyleHang, P.-Z., Zhu, H., Li, P.-F., Liu, J., Ge, F.-Q., Zhao, J., & Du, Z.-M. (2021). The Emerging Role of BDNF/TrkB Signaling in Cardiovascular Diseases. Life, 11(1), 70. https://doi.org/10.3390/life11010070