The Analysis of Human Serum N-Glycosylation in Patients with Primary and Metastatic Brain Tumors

 ,
, 
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
1. Introduction
2. Materials and Methods
2.1. Chemicals
2.2. Patient Samples
2.3. N-Glycan Release, Labeling, and Clean-Up
2.4. UPLC-FLR-MS Analysis
2.5. Data Analysis
3. Results and Discussion
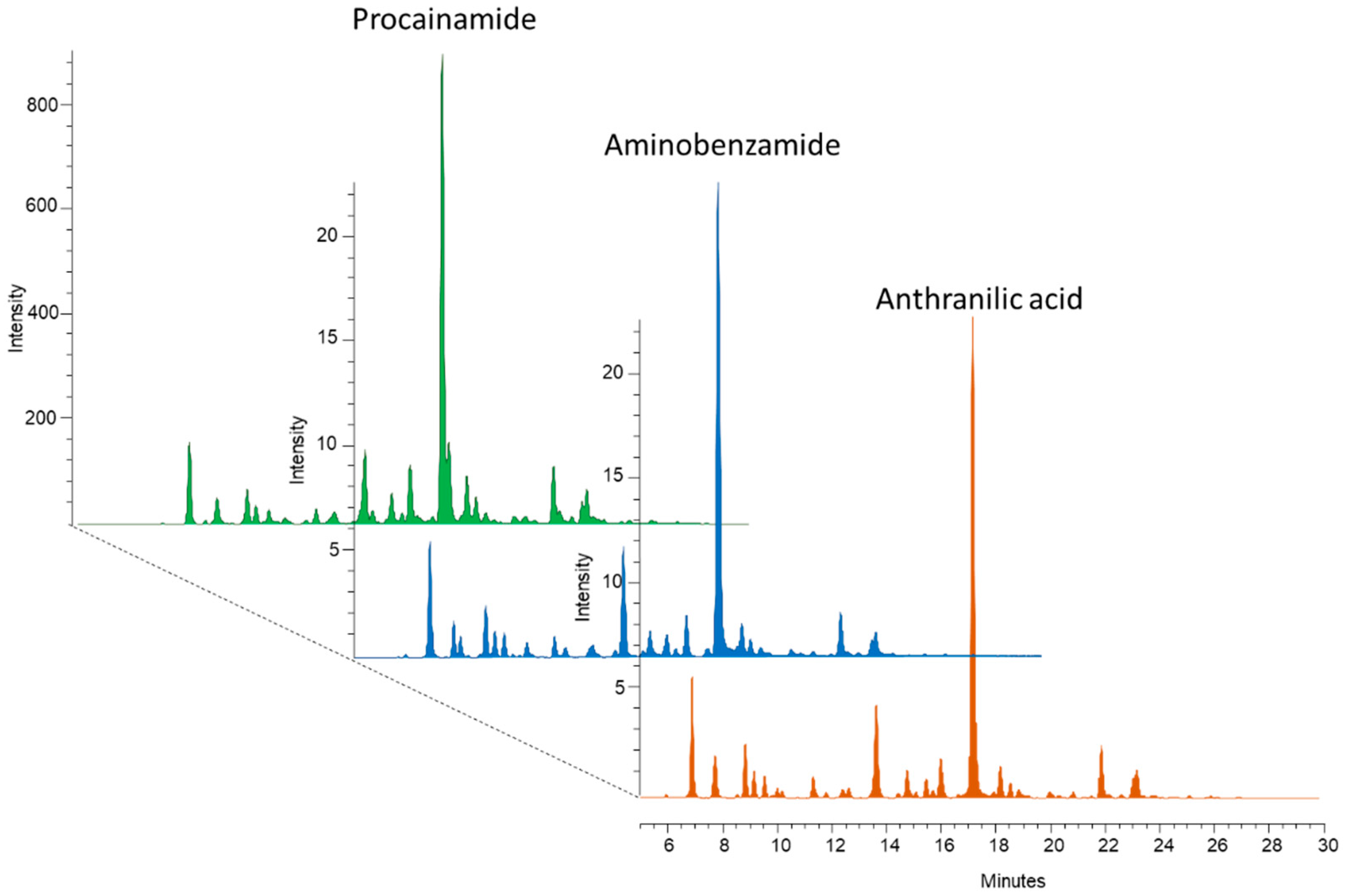
3.1. Magnetic-Nanoparticle-Based Sample Preparation
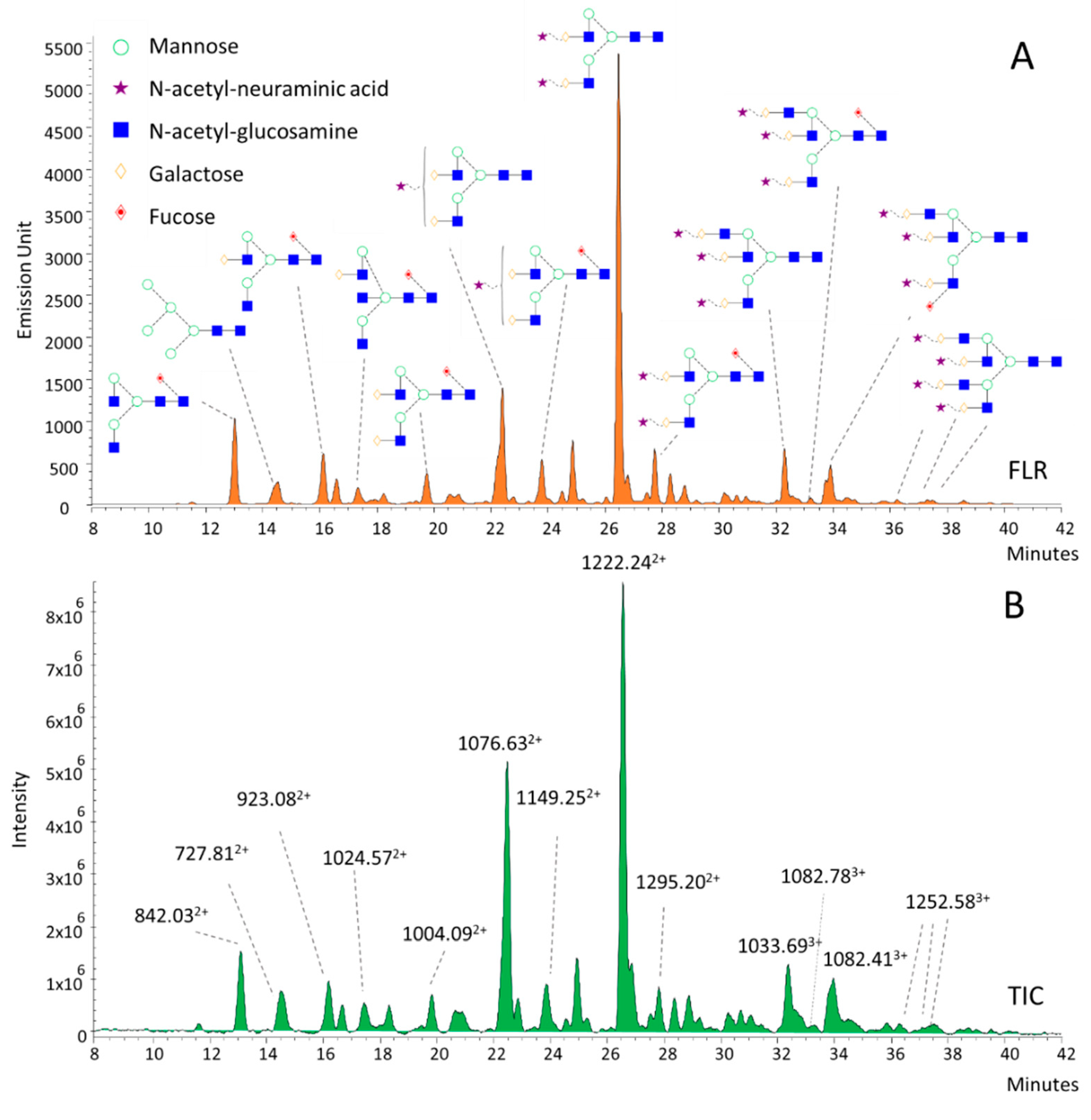
3.2. Structural Annotation of Serum N-Glycome
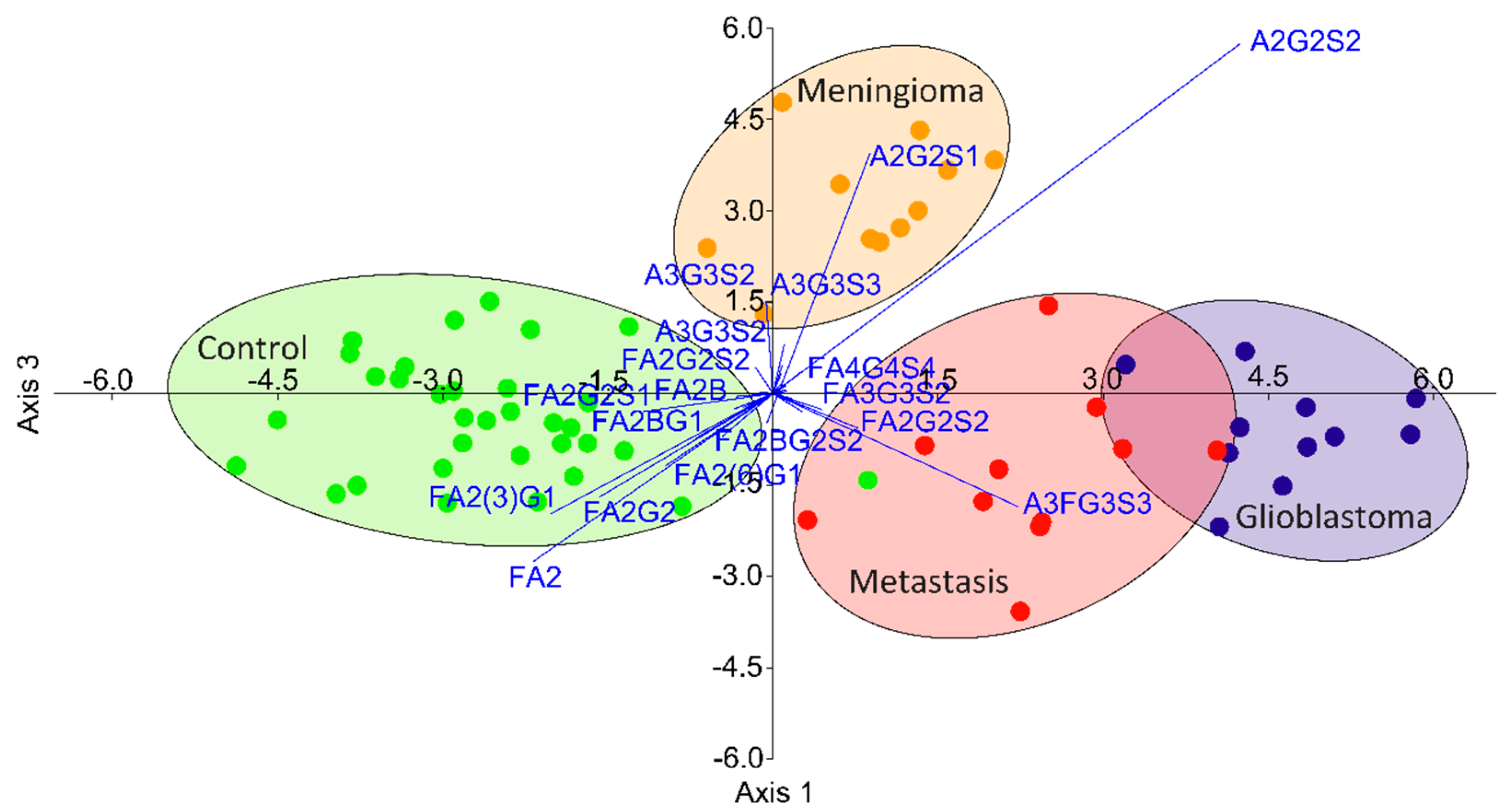
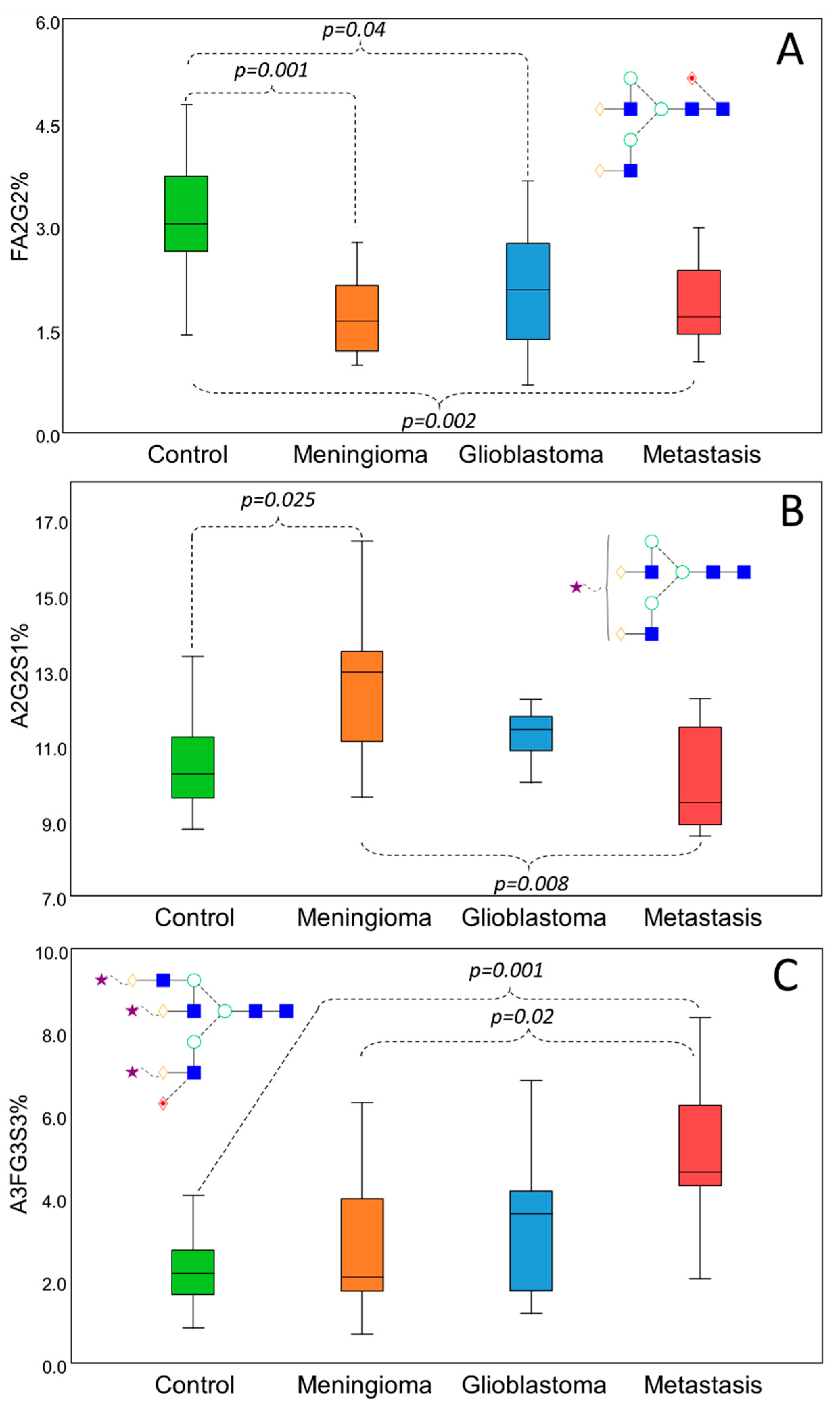
3.3. Data Analysis
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- GBD 2016 Brain and Other CNS Cancer Collaborators. Global, regional, and national burden of brain and other CNS cancer, 1990–2016: A systematic analysis for the Global Burden of Disease Study 2016. Lancet Neurol. 2019, 18, 376–393. [Google Scholar] [CrossRef]
- Arora, R.S.; Alston, R.D.; Eden, T.O.B.; Estlin, E.J.; Moran, A.; Birch, J.M. Age-incidence patterns of primary CNS tumors in children, adolescents, and adults in England. Neuro-Oncology 2009, 11, 403–413. [Google Scholar] [CrossRef]
- Nabors, L.B.; Ammirati, M.; Bierman, P.J.; Brem, H.; Butowski, N.; Chamberlain, M.C.; DeAngelis, L.M.; Fenstermaker, R.A.; Friedman, A.; Gilbert, M.R.; et al. Central nervous system cancers. J. Natl. Compr. Cancer Netw. 2013, 11, 1114–1151. [Google Scholar] [CrossRef]
- Thambi, R.; Kandamuthan, S.; Sainulabdeen, S.; Vilasiniamma, L.; Abraham, T.R.; Balakrishnan, P.K. Histopathological Analysis of Brain Tumours- A Seven Year Study from a Tertiary Care Centre in South India. J. Clin. Diagn. Res. 2017, 11, EC05–EC08. [Google Scholar] [CrossRef]
- Kuratsu, J.-I.; Kochi, M.; Ushio, Y. Incidence and clinical features of asymptomatic meningiomas. J. Neurosurg. 2000, 92, 766. [Google Scholar] [CrossRef] [PubMed]
- Dolecek, T.A.; Dressler, E.V.M.; Thakkar, J.P.; Liu, M.; Al-Qaisi, A.; Villano, J.L. Epidemiology of meningiomas post-Public Law 107-206: The Benign Brain Tumor Cancer Registries Amendment Act. Cancer 2015, 121, 2400–2410. [Google Scholar] [CrossRef] [PubMed]
- Gladson, C.L.; Prayson, R.A.; Liu, W.M. The pathobiology of glioma tumors. Annu. Rev. Pathol. 2010, 5, 33–50. [Google Scholar] [CrossRef] [PubMed]
- Walid, M.S. Prognostic factors for long-term survival after glioblastoma. Perm. J. 2008, 12, 45–48. [Google Scholar] [CrossRef] [PubMed]
- Leibold, A.T.; Monaco, G.N.; Dey, M. The role of the immune system in brain metastasis. Curr. Neurobiol. 2019, 10, 33–48. [Google Scholar]
- Henry, N.L.; Hayes, D.F. Cancer biomarkers. Mol. Oncol. 2012, 6, 140–146. [Google Scholar] [CrossRef]
- Gornik, O.; Lauc, G. Glycosylation of serum proteins in inflammatory diseases. Dis. Markers 2008, 25, 267–278. [Google Scholar] [CrossRef] [PubMed]
- Pinho, S.S.; Reis, C.A. Glycosylation in cancer: Mechanisms and clinical implications. Nat. Rev. Cancer 2015, 15, 540–555. [Google Scholar] [CrossRef] [PubMed]
- Stowell, S.R.; Ju, T.; Cummings, R.D. Protein glycosylation in cancer. Annu. Rev. Pathol. 2015, 10, 473–510. [Google Scholar] [CrossRef] [PubMed]
- Shan, M.; Yang, D.; Dou, H.; Zhang, L. Chapter Four—Fucosylation in cancer biology and its clinical applications. In Progress in Molecular Biology and Translational Science; Zhang, L., Ed.; Academic Press: Cambridge, MA, USA, 2019; Volume 162, pp. 93–119. [Google Scholar]
- Mereiter, S.; Balmaña, M.; Campos, D.; Gomes, J.; Reis, C.A. Glycosylation in the Era of Cancer-Targeted Therapy: Where Are We Heading? Cancer Cell 2019, 36, 6–16. [Google Scholar] [CrossRef] [PubMed]
- Kirwan, A.; Utratna, M.; O’Dwyer, M.E.; Joshi, L.; Kilcoyne, M. Glycosylation-Based Serum Biomarkers for Cancer Diagnostics and Prognostics. BioMed Res. Int. 2015, 2015, 490531. [Google Scholar] [CrossRef]
- Váradi, C.; Sikora, E.; Vanyorek, L.; Viskolcz, B. Purification of Fluorescently Derivatized N-Glycans by Magnetic Iron Nanoparticles. Nanomaterials 2019, 9, 1480. [Google Scholar] [CrossRef]
- Hammer, O.; Harper, D.; Ryan, P. PAST: Paleontological Statistics Software Package for Education and Data Analysis. Palaeontol. Electron. 2001, 4, 1–9. [Google Scholar]
- Harvey, D.J.; Merry, A.H.; Royle, L.; Campbell, M.P.; Dwek, R.A.; Rudd, P.M. Proposal for a standard system for drawing structural diagrams of N-and O-linked carbohydrates and related compounds. Proteomics 2009, 9, 3796–3801. [Google Scholar] [CrossRef]
- Ruhaak, L.R.; Zauner, G.; Huhn, C.; Bruggink, C.; Deelder, A.M.; Wuhrer, M. Glycan labeling strategies and their use in identification and quantification. Anal. Bioanal. Chem. 2010, 397, 3457–3481. [Google Scholar] [CrossRef]
- Keser, T.; Pavić, T.; Lauc, G.; Gornik, O. Comparison of 2-Aminobenzamide, Procainamide and RapiFluor-MS as Derivatizing Agents for High-Throughput HILIC-UPLC-FLR-MS N-glycan Analysis. Front. Chem. 2018, 6, 324. [Google Scholar] [CrossRef]
- Saldova, R.; Asadi Shehni, A.; Haakensen, V.D.; Steinfeld, I.; Hilliard, M.; Kifer, I.; Helland, Å.; Yakhini, Z.; Børresen-Dale, A.-L.; Rudd, P.M. Association of N-Glycosylation with Breast Carcinoma and Systemic Features Using High-Resolution Quantitative UPLC. J. Proteome Res. 2014, 13, 2314–2327. [Google Scholar] [CrossRef] [PubMed]
- Adamczyk, B.; Stöckmann, H.; O’Flaherty, R.; Karlsson, N.G.; Rudd, P.M. High-Throughput Analysis of the Plasma N-Glycome by UHPLC. In High-Throughput Glycomics and Glycoproteomics: Methods and Protocols; Lauc, G., Wuhrer, M., Eds.; Springer: New York, NY, USA, 2017; pp. 97–108. [Google Scholar] [CrossRef]
- Reiding, K.R.; Bondt, A.; Hennig, R.; Gardner, R.A.; O’Flaherty, R.; Trbojević-Akmačić, I.; Shubhakar, A.; Hazes, J.M.W.; Reichl, U.; Fernandes, D.L.; et al. High-throughput Serum N-Glycomics: Method Comparison and Application to Study Rheumatoid Arthritis and Pregnancy-associated Changes. Mol. Cell Proteom. 2019, 18, 3–15. [Google Scholar] [CrossRef] [PubMed]
- Li, F.; Ding, J. Sialylation is involved in cell fate decision during development, reprogramming and cancer progression. Protein Cell 2019, 10, 550–565. [Google Scholar] [CrossRef] [PubMed]
- Fuster, M.M.; Esko, J.D. The sweet and sour of cancer: Glycans as novel therapeutic targets. Nat. Rev. Cancer 2005, 5, 526–542. [Google Scholar] [CrossRef] [PubMed]
- Hu, Y.; Mayampurath, A.; Khan, S.; Cohen, J.K.; Mechref, Y.; Volchenboum, S.L. N-linked glycan profiling in neuroblastoma cell lines. J. Proteome Res. 2015, 14, 2074–2081. [Google Scholar] [CrossRef]
- Gatchev, O.; Råstam, L.; Lindberg, G.; Gullberg, B.; Eklund, G.A.; Törnberg, S. Tumours of the central nervous system and serum sialic acid concentration in men and women. Br. J. Cancer 1993, 68, 425–427. [Google Scholar] [CrossRef][Green Version]
- Gokmen, S.S.; Kazezoglu, C.; Tabakoglu, E.; Altiay, G.; Gungor, O.; Ture, M. Serum total sialic acid levels in lung cancer patients of different histological types with and no extrapulmonary metastases. Turk. J. Biochem. 2004, 29, 262–267. [Google Scholar]
- Kumar, S.; Saxena, M.; Srinivas, K.; Singh, V.K. Fucose: A biomarker in grading of oral cancer. Natl. J. Maxillofac. Surg. 2015, 6, 176–179. [Google Scholar] [CrossRef]
- Osuga, T.; Takimoto, R.; Ono, M.; Hirakawa, M.; Yoshida, M.; Okagawa, Y.; Uemura, N.; Arihara, Y.; Sato, Y.; Tamura, F.; et al. Relationship Between Increased Fucosylation and Metastatic Potential in Colorectal Cancer. JNCI 2016, 108. [Google Scholar] [CrossRef]
- Zipin, A.; Israeli-Amit, M.; Meshel, T.; Sagi-Assif, O.; Yron, I.; Lifshitz, V.; Bacharach, E.; Smorodinsky, N.I.; Many, A.; Czernilofsky, P.A.; et al. Tumor-microenvironment interactions: The fucose-generating FX enzyme controls adhesive properties of colorectal cancer cells. Cancer Res. 2004, 64, 6571–6578. [Google Scholar] [CrossRef]
- Liang, Y.; Han, P.; Wang, T.; Ren, H.; Gao, L.; Shi, P.; Zhang, S.; Yang, A.; Li, Z.; Chen, M. Stage-associated differences in the serum N-and O-glycan profiles of patients with non-small cell lung cancer. Clin. Proteom. 2019, 16, 20. [Google Scholar] [CrossRef] [PubMed]




Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Váradi, C.; Hajdu, V.; Farkas, F.; Gilányi, I.; Oláh, C.; Viskolcz, B. The Analysis of Human Serum N-Glycosylation in Patients with Primary and Metastatic Brain Tumors. Life 2021, 11, 29. https://doi.org/10.3390/life11010029
Váradi C, Hajdu V, Farkas F, Gilányi I, Oláh C, Viskolcz B. The Analysis of Human Serum N-Glycosylation in Patients with Primary and Metastatic Brain Tumors. Life. 2021; 11(1):29. https://doi.org/10.3390/life11010029
Chicago/Turabian StyleVáradi, Csaba, Viktória Hajdu, Flóra Farkas, Ibolya Gilányi, Csaba Oláh, and Béla Viskolcz. 2021. "The Analysis of Human Serum N-Glycosylation in Patients with Primary and Metastatic Brain Tumors" Life 11, no. 1: 29. https://doi.org/10.3390/life11010029
APA StyleVáradi, C., Hajdu, V., Farkas, F., Gilányi, I., Oláh, C., & Viskolcz, B. (2021). The Analysis of Human Serum N-Glycosylation in Patients with Primary and Metastatic Brain Tumors. Life, 11(1), 29. https://doi.org/10.3390/life11010029