
Antibody Cross-Reactivity in Serodiagnosis of Lyme Disease

Abstract
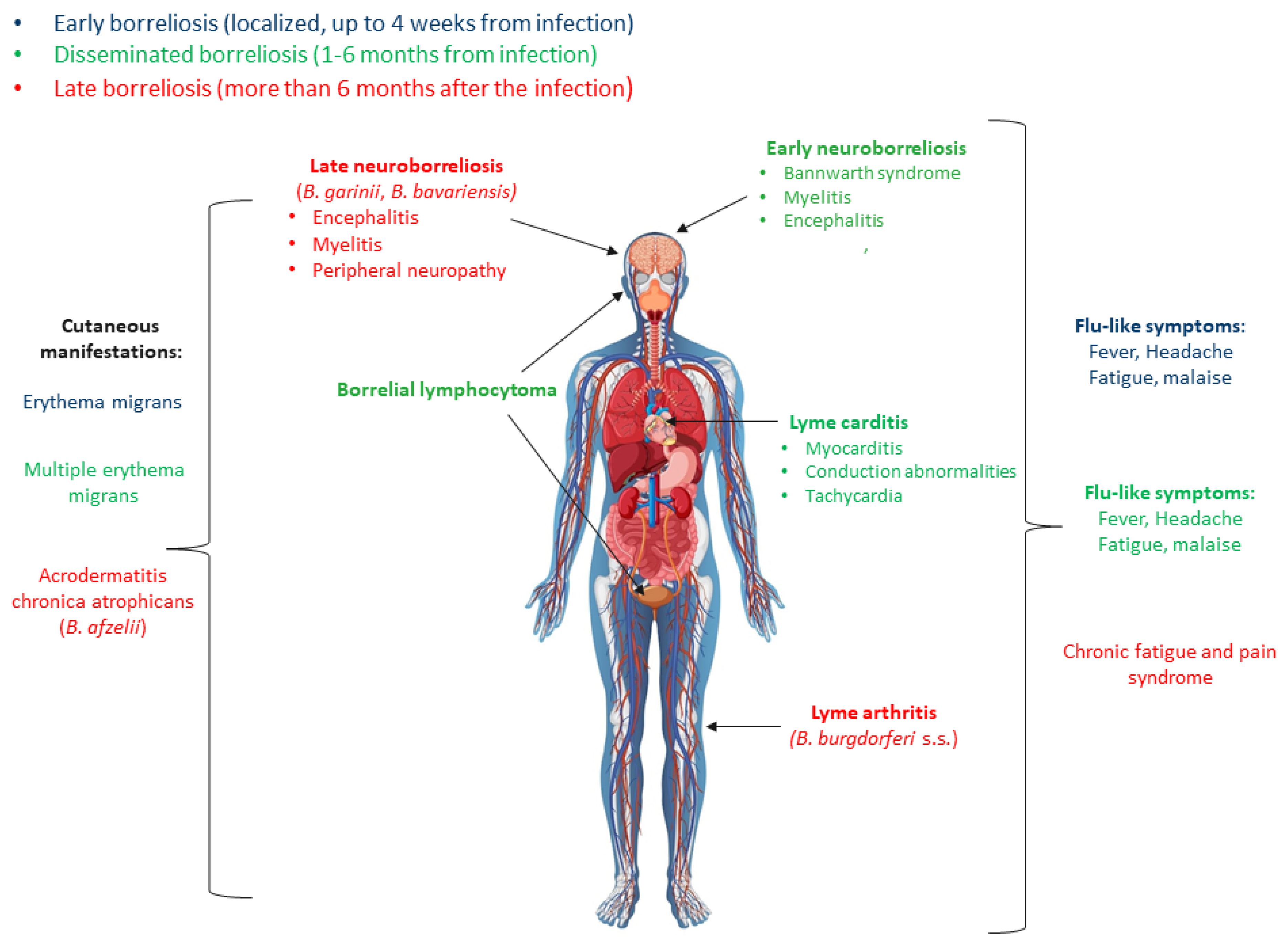
1. Introduction
2. Characteristic of B. burgdorferi s.l.
3. Diagnosis of Lyme Disease
4. Overdiagnosis of Lyme Disease
5. Disorders Most Often Confused with Lyme Disease
6. B. burgdorferi s.l. Cross-Reactive Proteins
7. Conclusions and Future Directions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Lohr, B.; Fingerle, V.; Norris, D.E.; Hunfeld, K.-P. Laboratory diagnosis of Lyme borreliosis: Current state of the art and future perspectives. Crit. Rev. Clin. Lab. Sci. 2018, 55, 219–245. [Google Scholar] [CrossRef] [PubMed]
- Pritt, B.S.; Respicio-Kingry, L.B.; Sloan, L.M.; Schriefer, M.E.; Replogle, A.J.; Bjork, J.; Liu, G.; Kingry, L.C.; Mead, P.S.; Neitzel, D.F.; et al. Borrelia mayonii sp. nov., a member of the Borrelia burgdorferi sensu lato complex, detected in patients and ticks in the upper midwestern United States. Int. J. Syst. Evol. Microbiol. 2016, 123, 4757–4763. [Google Scholar] [CrossRef] [PubMed]
- Rudenko, N.; Golovchenko, M.; Grubhoffer, L.; Oliver, J.H. Updates on Borrelia burgdorferi sensu lato complex with respect to public health. Ticks Tick-Borne Dis. 2011, 2, 123–128. [Google Scholar] [CrossRef] [PubMed]
- de Carvalho, I.L.; Fonseca, J.E.; Marques, J.G.; Ullmann, A.; Hojgaard, A.; Zeidner, N.; Núncio, M.S. Vasculitis-like syndrome associated with Borrelia lusitaniae infection. Clin. Rheumatol. 2008, 27, 1587–1591. [Google Scholar] [CrossRef]
- Margos, G.; Vollmer, S.A.; Ogden, N.H.; Fish, D. Population genetics, taxonomy, phylogeny and evolution of Borrelia burgdorferi sensu lato. Infect. Genet. Evol. 2011, 11, 1545–1563. [Google Scholar] [CrossRef]
- Eisen, L. Vector competence studies with hard ticks and Borrelia burgdorferi sensu lato spirochetes: A review. Ticks Tick-Borne Dis. 2020, 11, 101359. [Google Scholar] [CrossRef]
- Mannelli, A.; Bertolotti, L.; Gern, L.; Gray, J. Ecology of Borrelia burgdorferi sensu lato in Europe: Transmission dynamics in multi-host systems, influence of molecular processes and effects of climate change. FEMS Microbiol. Rev. 2011, 36, 837–861. [Google Scholar] [CrossRef]
- Allen, C.; Steere, M.; Stephen, E.; Malavista, M.; John, A.; Hardin, M.D.; Shaun Ruddy, M.; Philip, W.; Askenase, M.; Warren, A.; et al. Erythema chronicum migrans and Lyme arthritis. Ann. Agric. Environ. Med. 1977, 86, 685–698. [Google Scholar] [CrossRef]
- Burgdorfer, W.; Bargour, A.G.; Hayes, S.F.; Benach, J.; Grunwaldt, E.; Davis, J.P. Lyme Disease—A tick-borne spirochetosis ? Science 1982, 216, 1317–1319. [Google Scholar] [CrossRef]
- Strand, M.; Hönig, V.; Růžek, D.; Grubhoffer, L.; Regoa, R.O.M. Europe-wide meta-analysis of Borrelia burgdorferi sensu lato prevalence in questing Ixodes ricinus ticks. Appl. Environ. Microbiol. 2017, 83, e00609-17. [Google Scholar] [CrossRef]
- Blanchard, L.; Jones-Diette, J.; Lorenc, T.; Sutcliffe, K.; Sowden, A.; Thomas, J. Comparison of national surveillance systems for Lyme disease in humans in Europe and North America: A policy review. BMC Public Health 2022, 22, 1307. [Google Scholar] [CrossRef] [PubMed]
- Sykes, R.A.; Makiello, P. An estimate of Lyme borreliosis incidence in Western Europe. J. Public Health 2017, 39, 74–81. [Google Scholar] [CrossRef]
- Stevenson, B.; Krusenstjerna, A.C.; Castro-Padovani, T.N.; Savage, C.R.; Jutras, B.L.; Saylor, T.C. The consistent tick-vertebrate infectious cycle of the Lyme disease spirochete enables Borrelia burgdorferi to control protein expression by monitoring its physiological status. J. Bacteriol. 2022, 204, e00606-21. [Google Scholar] [CrossRef]
- Hofmann, H.; Fingerle, V.; Hunfeld, K.P.; Huppertz, H.I.; Krause, A.; Rauer, S.; Ruf, B. Cutaneous lyme borreliosis: Guideline of the german dermatology society. Ger. Med. Sci. 2017, 15, 1–31. [Google Scholar] [CrossRef]
- Krzyczmanik, D.; Sińczuk-Walczak, H.; Wittczak, T.; Cyran, A.; Pałczyński, C.; Walusiak-Skorupa, J. Borreliosis in occupational medicine practice. Med. Pr. 2012, 63, 483–492. [Google Scholar] [PubMed]
- Wang, G.; van Dam, A.P.; Schwartz, I.; Dankert, J. Molecular typing of Borrelia burgdorferi sensu lato: Taxonomic, epidemiological, and clinical implications. Clin. Microbiol. Rev. 1999, 12, 633–653. [Google Scholar] [CrossRef]
- Shapiro, E.D. Lyme disease. N. Engl. J. Med. 2014, 370, 1724–1731. [Google Scholar] [CrossRef]
- Stanek, G.; Wormser, G.P.; Gray, J.; Strle, F. Lyme borreliosis. Lancet 2012, 379, 461–473. [Google Scholar] [CrossRef]
- Aguero-Rosenfeld, M.; Wang, G.; Scwartz, I.; Wormser, G. Diagnosis of Lyme borreliosis. Clin. Microbiol. Rev. 2005, 18, 484–509. [Google Scholar] [CrossRef]
- Matyjasek, A.; Zdrojewski, Z. Borelioza—Najnowsze rekomendacje w diagnostyce i leczeniu Borreliosis—The latest recommendations on diagnosis. Forum Reumatol. 2016, 2, 58–64. [Google Scholar]
- Caine, J.A.; Coburn, J. Multifunctional and redundant roles of Borrelia burgdorferi outer surface proteins in tissue adhesion, colonization, and complement evasion. Front. Immunol. 2016, 7, 442. [Google Scholar] [CrossRef] [PubMed]
- van Dam, A.P.; Kuiper, H.; Vos, K.; Widjojokusumo, A.; de Jongh, B.M.; Spanjaard, L.; Ramselaar, A.C.; Kramer, M.D.; Dankert, J. Different genospecies of Borrelia burgdorferi are associated with distinct clinical manifestations of Lyme borreliosis. Clin. Infect. Dis. 1993, 17, 708–717. [Google Scholar] [CrossRef] [PubMed]
- Fingerle, V.; Schulte-Spechtel, U.C.; Ruzic-Sabljic, E.; Leonhard, S.; Hofmann, H.; Weber, K.; Pfister, K.; Strle, F.; Wilske, B. Epidemiological aspects and molecular characterization of Borrelia burgdorferi s.l. from southern Germany with special respect to the new species Borrelia spielmanii sp. nov. Int. J. Med. Microbiol. 2008, 298, 279–290. [Google Scholar] [CrossRef]
- Stanek, G.; Reiter, M. The expanding Lyme Borrelia complex-clinical significance of genomic species? Clin. Microbiol. Infect. 2011, 17, 487–493. [Google Scholar] [CrossRef]
- Schutzer, S.E.; Body, B.A.; Boyle, J.; Branson, B.M.; Dattwyler, R.J.; Fikrig, E.; Gerald, N.J.; Gomes-Solecki, M.; Kintrup, M.; Ledizet, M.; et al. Direct diagnostic tests for Lyme disease. Clin. Infect. Dis. 2019, 68, 1052–1057. [Google Scholar] [CrossRef] [PubMed]
- Barbour, A.G.; Hayes, S.F. Biology of Borrelia species. Microbiol. Rev. 1986, 50, 381–400. [Google Scholar] [CrossRef] [PubMed]
- Goettner, G.; Schulte-Spechtel, U.; Hillermann, R.; Liegl, G.; Wilske, B.; Fingerle, V. Improvement of Lyme borreliosis serodiagnosis by a newly developed recombinant immunoglobulin G (IgG) and IgM line immunoblot assay and addition of VlsE and DbpA homologues. J. Clin. Microbiol. 2005, 43, 3602–3609. [Google Scholar] [CrossRef]
- Goldstein, S.F.; Buttle, K.F.; Charon, N.W. Structural analysis of Leptospiraceae and Borrelia burgdorferi by high voltage electron microscopy. J. Bacteriol. 1996, 178, 6539–6545. [Google Scholar] [CrossRef][Green Version]
- Hyde, J.A. Borrelia burgdorferi keeps moving and carries on: A review of borrelial dissemination and invasion. Front. Immunol. 2017, 8, 114. [Google Scholar] [CrossRef]
- Casjens, S.R.; Gilcrease, E.B.; Vujadinovic, M.; Mongodin, E.F.; Luft, B.J.; Schutzer, S.E.; Fraser, C.M.; Qiu, W.G. Plasmid diversity and phylogenetic consistency in the Lyme disease agent Borrelia burgdorferi. BMC Genom. 2017, 18, 165. [Google Scholar] [CrossRef]
- Brisson, D.; Drecktrah, D.; Eggers, C.H.; Samuels, D.S. Genetics of Borrelia burgdorferi. Annu. Rev. Genet. 2012, 46, 515–536. [Google Scholar] [CrossRef]
- Ohnishi, J.; Piesman, J.; de Silva, A.M. Antigenic and genetic heterogeneity of Borrelia burgdorferi populations transmitted by ticks. Proc. Natl. Acad. Sci. USA 2001, 98, 670–675. [Google Scholar] [CrossRef] [PubMed]
- Ojaimi, C.; Brooks, C.; Casjens, S.; Rosa, P.; Elias, A.; Barbour, A.; Jasinskas, A.; Benach, J.; Katona, L.; Radolf, J.; et al. Profiling of temperature-induced changes in Borrelia burgdorferi gene expression by using whole genome arrays. Infect. Immun. 2003, 71, 1689–1705. [Google Scholar] [CrossRef] [PubMed]
- Kenedy, M.R.; Lenhart, T.R.; Akins, D.R. The role of Borrelia burgdorferi outer surface proteins. FEMS Immunol. Med. Microbiol. 2012, 66, 1–19. [Google Scholar] [CrossRef]
- Fraser, C.M.; Casjens, S.; Huang, W.M.; Sutton, G.G.; Clayton, R.; Lathigra, R.; White, O.; Ketchum, K.A.; Dodson, R.; Hickey, E.K.; et al. Genomic sequence of a Lyme disease spirochaete, Borrelia burgdorferi. Nature 1997, 390, 580–586. [Google Scholar] [CrossRef] [PubMed]
- Purser, J.E.; Norris, S.J. Correlation between plasmid content and infectivity in Borrelia burgdorferi. Proc. Natl. Acad. Sci. USA 2000, 97, 13865–13870. [Google Scholar] [CrossRef] [PubMed]
- Schwan, T.G.; Piesman, J.; Golde, W.T.; Dolan, M.C.; Rosa, P.A. Induction of an outer surface protein on Borrelia burgdorferi during tick feeding. Proc. Natl. Acad. Sci. USA 1995, 92, 2909–2913. [Google Scholar] [CrossRef]
- Yang, X.F.; Pal, U.; Alani, S.M.; Fikrig, E.; Norgard, M.V. Essential role for OspA/B in the life cycle of the Lyme disease spirochete. J. Exp. Med. 2004, 199, 641–648. [Google Scholar] [CrossRef]
- Norris, S.J. The dynamic proteome of Lyme disease Borrelia. Genome Biol. 2006, 7, 209. [Google Scholar] [CrossRef]
- Li, X.; Neelakanta, G.; Liu, X.; Beck, D.S.; Kantor, F.S.; Fish, D.; Anderson, J.F.; Fikrig, E. Role of outer surface protein D in the Borrelia burgdorferi life cycle. Infect. Immun. 2007, 75, 4237–4244. [Google Scholar] [CrossRef]
- Carrasco, S.E.; Troxell, B.; Yang, Y.; Brandt, S.L.; Li, H.; Sandusky, G.E.; Condon, K.W.; Serezani, C.H.; Frank, X. Outer surface protein OspC is an antiphagocytic factor that protects Borrelia burgdorderi from phagocytosis by macrophages. Am. Soc. Microbiol. 2015, 83, 4848–4860. [Google Scholar] [CrossRef]
- Schwan, T.G.; Piesman, J. Temporal changes in outer surface proteins A and C of the lyme disease- associated spirochete, Borrelia burgdorferi, during the chain of infection in ticks and mice. J. Clin. Microbiol. 2000, 38, 382–388. [Google Scholar] [CrossRef] [PubMed]
- Gilmore, R.D.; Howison, R.R.; Dietrich, G.; Patton, T.G.; Clifton, D.R.; Carroll, J.A. The bba64 gene of Borrelia burgdorferi, the Lyme disease agent, is critical for mammalian infection via tick bite transmission. Proc. Natl. Acad. Sci. USA 2010, 107, 7515–7520. [Google Scholar] [CrossRef] [PubMed]
- Brandt, K.S.; Patton, T.G.; Allard, A.S.; Caimano, M.J.; Radolf, J.D.; Gilmore, R.D. Evaluation of the Borrelia burgdorferi BBA64 protein as a protective immunogen in mice. Clin. Vaccine Immunol. 2014, 21, 526–533. [Google Scholar] [CrossRef] [PubMed]
- Schulte-Spechtel, U.; Fingerle, V.; Goettner, G.; Rogge, S.; Wilske, B. Molecular analysis of decorin-binding protein A (DbpA) reveals five major groups among European Borrelia burgdorferi sensu lato strains with impact for the development of serological assays and indicates lateral gene transfer of the dbpA gene. Int. J. Med. Microbiol. 2006, 296, 250–266. [Google Scholar] [CrossRef]
- Garcia, B.L.; Zhi, H.; Wager, B.; Höök, M.; Skare, J.T. Borrelia burgdorferi BBK32 inhibits the classical pathway by blocking activation of the C1 complement complex. PLoS Pathog. 2016, 12, e1005404. [Google Scholar] [CrossRef]
- Xie, J.; Zhi Id, H.; Garrigues Id, R.J.; Keightley, A.; Garcia, B.L.; Skare, J.T. Structural determination of the complement inhibitory domain of Borrelia burgdorferi BBK32 provides insight into classical pathway complement evasion by Lyme disease spirochetes. PLoS Pathog. 2019, 15, e1007659. [Google Scholar] [CrossRef]
- Panelius, J.; Lahdenne, P.; Saxén, H.; Carlsson, S.A.; Heikkilä, T.; Peltomaa, M.; Lauhio, A.; Seppälä, I. Diagnosis of Lyme neuroborreliosis with antibodies to recombinant proteins DbpA, BBK32, and OspC, and VIsE IR6peptide. J. Neurol. 2003, 250, 1318–1327. [Google Scholar] [CrossRef]
- Zhang, J.R.; Hardham, J.M.; Barbour, A.G.; Norris, S.J. Antigenic variation in Lyme disease borreliae by promiscuous recombination of VMP-like sequence cassettes. Cell 1997, 89, 275–285. [Google Scholar] [CrossRef]
- Roessler, D.; Hauser, U.; Wilske, B. Heterogeneity of BmpA (P39) among European isolates of Borrelia burgdorferi sensu lato and influence of interspecies variability on serodiagnosis. J. Clin. Microbiol. 1997, 35, 2752–2758. [Google Scholar] [CrossRef]
- Verma, A.; Brissette, C.A.; Bowman, A.; Stevenson, B. Borrelia burgdorferi BmpA is a laminin-binding protein. Infect. Immun. 2009, 77, 4940–4946. [Google Scholar] [CrossRef] [PubMed]
- Yang, X.; Izadi, H.; Coleman, A.S.; Wang, P.; Ma, Y.; Fikrig, E.; Anguita, J.; Pal, U. Borrelia burgdorferi lipoprotein BmpA activates pro-inflammatory responses in human synovial cells through a protein moiety. Microbes Infect. 2008, 10, 1300–1308. [Google Scholar] [CrossRef] [PubMed]
- Coburn, J.; Leong, J.; Chaconas, G. Illuminating the roles of the Borrelia burgdorferi adhesins. Trends Microbiol. 2013, 21, 372–379. [Google Scholar] [CrossRef] [PubMed]
- Stevenson, B.; El-Hage, N.; Hines, M.A.; Miller, J.C.; Babb, K. Differential binding of host complement inhibitor factor H by Borrelia burgdorferi Erp surface proteins: A possible mechanism underlying the expansive host range of Lyme disease spirochetes. Infect. Immun. 2002, 70, 491–497. [Google Scholar] [CrossRef]
- Kraiczy, P.; Hartmann, K.; Hellwage, J.; Skerka, C.; Kirschfink, M.; Brade, V.; Zipfel, P.F.; Wallich, R.; Stevenson, B. Immunological characterization of the complement regulator factor H-binding CRASP and Erp proteins of Borrelia burgdorferi. Int. J. Med. Microbiol. 2004, 293, 152–157. [Google Scholar] [CrossRef]
- Arnaboldi, P.M.; Dattwyler, R.J. Cross-reactive epitopes in Borrelia burgdorferi p66. Clin. Vaccine Immunol. 2015, 22, 840–843. [Google Scholar] [CrossRef]
- Wallich, R.; Simon, M.M.; Hofmann, H.; Moter, S.E.; Schaible, U.E.; Kramer, M.D. Molecular and immunological characterization of a novel polymorphic lipoprotein of Borrelia burgdorferi. Infect. Immun. 1993, 61, 4158–4166. [Google Scholar] [CrossRef]
- Signorino, G.; Arnaboldi, P.M.; Petzke, M.M.; Dattwyler, R.J. Identification of OppA2 linear epitopes as serodiagnostic markers for Lyme disease. Clin. Vaccine Immunol. 2014, 21, 704–711. [Google Scholar] [CrossRef]
- Ge, Y.; Li, C.; Corum, L.; Slaughter, C.A. Structure and expression of the FlaA periplasmic flagellar protein of Borrelia burgdorferi. J. Bacteriol. 1998, 180, 2418–2425. [Google Scholar] [CrossRef]
- Panelius, J.; Lahdenne, P.; Saxen, H.; Heikkila, T.; Seppala, I. Recombinant flagellin A proteins from Borrelia burgdorferi sensu stricto, B. afzelii, and B. garinii in serodiagnosis of Lyme borreliosis. J. Clin. Microbiol. 2001, 39, 4013–4019. [Google Scholar] [CrossRef]
- Johnson, B.J.B.; Robbins, K.E.; Bailey, R.E.; Cao, B.L.; Sviat, S.L.; Craven, R.B.; Mayer, L.W.; Dennis, D.T. Serodiagnosis of lyme disease: Accuracy of a two-step approach using a flagella-based ELISA and immunoblotting. J. Infect. Dis. 1996, 174, 346–353. [Google Scholar] [CrossRef] [PubMed]
- Wallich, R.; Moter, S.E.; Simon, M.M.; Ebnet, K.; Heiberger, A.; Kramer, M.D. The Borrelia burgdorferi flagellum-associated 41-kilodalton antigen (Flagellin): Molecular cloning, expression, and amplification of the gene. Infect. Immun. 1990, 58, 1711–1719. [Google Scholar] [CrossRef] [PubMed]
- Liveris, D.; Wang, G.; Girao, G.; Byrne, D.W.; Nowakowski, J.; McKenna, D.; Nadelman, R.; Wormser, G.P.; Schwartz, I. Quantitative detection of Borrelia burgdorferi in 2-millimeter skin samples of erythema migrans lesions: Correlation of results with clinical and laboratory findings. J. Clin. Microbiol. 2002, 40, 1249–1253. [Google Scholar] [CrossRef]
- van Dam, A.P. Molecular diagnosis of Borrelia bacteria for the diagnosis of Lyme disease. Expert Opin. Med. Diagn. 2011, 5, 135–149. [Google Scholar] [CrossRef] [PubMed]
- Engstrom, S.M.; Shoop, E.; Johnson, R.C. Immunoblot interpretation criteria for serodiagnosis of early Lyme disease. J. Clin. Microbiol. 1995, 33, 419–427. [Google Scholar] [CrossRef] [PubMed]
- Dressler, F.; Whalen, J.A.; Reinhardt, B.N.; Steere, A.C. Western Blotting in the serodiagnosis of Lyme-disease. J. Infect. Dis. 1993, 167, 392–400. [Google Scholar] [CrossRef]
- Wilske, B.; Fingerle, V.; Schulte-Spechtel, U. Microbiological and serological diagnosis of Lyme borreliosis. FEMS Immunol. Med. Microbiol. 2007, 49, 13–21. [Google Scholar] [CrossRef]
- Jonsson, M.; Noppa, L.; Barbour, A.G.; Bergstrom, S. Heterogeneity of outer membrane proteins in Borrelia burgdorferi: Comparison of osp operons of three isolates of different geographic origins. Infect. Immun. 1992, 60, 1845–1853. [Google Scholar] [CrossRef]
- Roberts, W.C.; Mullikin, B.A.; Lathigra, R.; Hanson, M.S. Molecular analysis of sequence heterogeneity among genes encoding decorin binding proteins A and B of Borrelia burgdorferi sensu lato. Infect. Immun. 1998, 66, 5275–5285. [Google Scholar] [CrossRef]
- Gilmore, R.D.; Mbow, M.L.; Stevenson, B. Analysis of Borrelia burgdorferi gene expression during life cycle phases of the tick vector Ixodes scapularis. Microbes Infect. 2001, 3, 799–808. [Google Scholar] [CrossRef]
- Liang, F.T.; Steere, A.C.; Marques, A.R.; Johnson, B.J.; Miller, J.N.; Philipp, M.T. Sensitive and specific serodiagnosis of Lyme disease by enzyme-linked immunosorbent assay with a peptide based on an immunodominant conserved region of Borrelia burgdorferi VlsE. J. Clin. Microbiol. 1999, 37, 3990–3996. [Google Scholar] [CrossRef] [PubMed]
- Norris, S.J. vls Antigenic variation systems of Lyme disease Borrelia: Eluding host immunity through both random, segmental gene conversion and framework heterogeneity. Microbiol. Spectr. 2015, 2, 471–489. [Google Scholar] [CrossRef]
- Centers for disease control and prevention (CDC) recommendations for test performance and interpretation from the second national conference on serologic diagnosis of Lyme disease. JAMA J. Am. Med. Assoc. 1995, 274, 937. [CrossRef]
- Branda, J.A.; Linskey, K.; Kim, Y.A.; Steere, A.C.; Ferraro, M.J. Two-tiered antibody testing for Lyme disease with use of 2 enzyme immunoassays, a whole-cell sonicate enzyme immunoassay followed by a vlse c6 peptide enzyme immunoassay. Clin. Infect. Dis. 2011, 53, 541–547. [Google Scholar] [CrossRef]
- Bacon, R.M.; Biggerstaff, B.J.; Schriefer, M.E.; Gilmore, R.D., Jr.; Philipp, M.T.; Steere, A.C.; Wormser, G.P.; Marques, A.R.; Johnson, B.J.B. Serodiagnosis of Lyme disease by kinetic enzyme-linked immunosorbent assay using recombinant VlsE1 or peptide antigens of Borrelia burgdorferi compared with 2-tiered testing using whole-cell lysates. J. Infect. Dis. 2003, 187, 1187–1199. [Google Scholar] [CrossRef] [PubMed]
- Hillerdal, H.; Henningsson, A.J. Serodiagnosis of Lyme borreliosis—Is IgM in serum more harmful than helpful? Eur. J. Clin. Microbiol. Infect. Dis. 2021, 40, 1161–1168. [Google Scholar] [CrossRef]
- Dessau, R.B.; van Dam, A.P.; Fingerle, V.; Gray, J.; Hovius, J.W.; Hunfeld, K.P.; Jaulhac, B.; Kahl, O.; Kristoferitsch, W.; Lindgren, P.E.; et al. To test or not to test? Laboratory support for the diagnosis of Lyme borreliosis: A position paper of ESGBOR, the ESCMID study group for Lyme borreliosis. Clin. Microbiol. Infect. 2018, 24, 118–124. [Google Scholar] [CrossRef]
- Lantos, P.M.; Lipsett, S.C.; Nigrovic, L.E. False positive Lyme disease IgM immunoblots in children. J. Pediatr. 2016, 174, 267–269. [Google Scholar] [CrossRef]
- Seriburi, V.; Ndukwe, N.; Chang, Z.; Cox, M.E.; Wormser, G.P. High frequency of false positive IgM immunoblots for Borrelia burgdorferi in clinical practice. Clin. Microbiol. Infect. 2012, 18, 1236–1240. [Google Scholar] [CrossRef]
- Cetin, E.; Sotoudeh, M.; Auer, H.; Stanek, G. Paradigm Burgenland: Risk of Borrelia burgdorferi sensu lato infection indicated by variable seroprevalence rates in hunters. Wien. Klin. Wochenschr. 2006, 118, 677–681. [Google Scholar] [CrossRef]
- Zając, V.; Pinkas, J.; Wójcik-Fatla, A.; Dutkiewicz, J.; Owoc, A.; Bojar, I. Prevalence of serological response to Borrelia burgdorferi in farmers from eastern and central Poland. Eur. J. Clin. Microbiol. Infect. Dis. 2017, 36, 437–446. [Google Scholar] [CrossRef] [PubMed]
- Kalish, R.; McHugh, G.; Granquist, J.; Shea, B.; Ruthazer, R.; Steere, A.C. Persistence of immunoglobulin M or immunoglobulin G antibody responses to Borrelia burgdorferi 10-20 years after active Lyme disease. Clin. Infect. Dis. 2001, 33, 780–785. [Google Scholar] [CrossRef] [PubMed]
- Wojciechowska-Koszko, I.; Kwiatkowski, P.; Sienkiewicz, M.; Kowalczyk, M.; Kowalczyk, E.; Dołęgowska, B. Cross-reactive results in serological tests for borreliosis in patients with active viral infections. Pathogens 2022, 11, 203. [Google Scholar] [CrossRef] [PubMed]
- Smismans, A.; Goossens, V.J.; Nulens, E.; Bruggeman, C.A. Comparison of five different immunoassays for the detection of Borrelia burgdorferi IgM and IgG antibodies. Clin. Microbiol. Infect. 2006, 12, 648–655. [Google Scholar] [CrossRef] [PubMed]
- Talagrand-Reboul, E.; Raffetin, A.; Zachary, P.; Jaulhac, B.; Eldin, C. Immunoserological diagnosis of human borrelioses: Current knowledge and perspectives. Front. Cell. Infect. Microbiol. 2020, 10, 241. [Google Scholar] [CrossRef]
- Magnarelli, L.A.; Lawrenz, M.; Fikrig, E.; Norris, S.J. Comparative reactivity of human sera to recombinant VlsE and other Borrelia burgdorferi antigens in class-specific enzyme-linked immunosorbent assays for Lyme borreliosis. J. Med. Microbiol. 2002, 51, 649–655. [Google Scholar] [CrossRef]
- Panelius, J.; Lahdenne, P.; Heikkila, T.; Peltomaa, M.; Oksi, J.; Seppala, I. Recombinant OspC from Borrelia burgdorferi sensu stricto B. afzelii and B. garinii in the serodiagnosis of Lyme borreliosis. J. Med. Microbiol. 2002, 51, 731–739. [Google Scholar] [CrossRef]
- Jakab, Á.; Kahlig, P.; Kuenzli, E.; Neumayr, A. Tick borne relapsing fever—A systematic review and analysis of the literature. PLoS Negl. Trop. Dis. 2022, 16, e0010212. [Google Scholar] [CrossRef]
- Magnarelli, L.A.; Miller, J.N.; Anderson, J.F.; Riviere, G.R. Cross-reactivity of nonspecific treponemal antibody in serologic tests for Lyme disease. J. Clin. Microbiol. 1990, 28, 1276–1279. [Google Scholar] [CrossRef]
- Patriquin, G.; LeBlanc, J.; Heinstein, C.; Roberts, C.; Lindsay, R.; Hatchette, T.F. Cross-reactivity between Lyme and syphilis screening assays: Lyme disease does not cause false-positive syphilis screens. Diagn. Microbiol. Infect. Dis. 2016, 84, 184–186. [Google Scholar] [CrossRef]
- Raoult, D.; Hechemy, K.E.; Baranton, G. Cross-reaction with Borrelia burgdorferi antigen of sera from patients with human immunodeficiency virus infection, syphillis, and leptospirosis. J. Clin. Microbiol. 1989, 27, 2152–2155. [Google Scholar] [CrossRef] [PubMed]
- Hunter, E.F.; RusselL, H.; Farshy, C.; Sampson, J. Evaluation of Sera from patients with Lyme disease in the fluorescent Treponemal antibody-absorption test for syphilis. Sex. Transm. Dis. 1986, 13, 232–236. [Google Scholar] [CrossRef] [PubMed]
- Obert, R.; Adelman, B.N.; Orowitz, A.W.H.; Ze -Chen, T.; Sieh, H.; Oseph, J.; Aria, M.; Guero-R Osenfeld, E.A.; Ra, I.; Chwartz, S.; et al. Simultaneous human granulocytic ehrlichiosis and Lyme borreliosis. N. Engl. J. Med. 2009, 337, 27–30. [Google Scholar] [CrossRef]
- Schicht, S.; Junge, S.; Schnieder, T.; Strube, C. Prevalence of Anaplasma phagocytophilum and coinfection with Borrelia burgdorferi sensu lato in the hard tick Ixodes ricinus in the city of Hanover (Germany). Vector-Borne Zoonotic Dis. 2011, 11, 1595–1597. [Google Scholar] [CrossRef]
- Magnarelli, L.A.; Dumler, J.S.; Anderson, J.F.; Johnson, R.C.; Fikrig, E. Coexistence of antibodies to tick-borne pathogens of babesiosis, ehrlichiosis, and Lyme borreliosis in human sera. J. Clin. Microbiol. 1995, 33, 3054–3057. [Google Scholar] [CrossRef]
- Rawlins, M.L.; Gerstner, C.; Hill, H.R.; Litwin, C.M. Evaluation of a Western blot method for the detection of Yersinia antibodies: Evidence of serological cross-reactivity between Yersinia outer membrane proteins and Borrelia burgdorferi. Clin. Diagn. Lab. Immunol. 2005, 12, 1269–1274. [Google Scholar] [CrossRef]
- Golkocheva-Markova, E.; Nenova, R.; Stoilov, R.; Christova, I.; Najdenski, H. Cross-reactivity between Yersinia outer membrane proteins and anti-Francisella and anti-Borrelia antibodies in serodiagnosis of Yersinia-triggered reactive arthritis. Comptes Rendus L’Academie Bulg. Sci. 2011, 64, 61–66. [Google Scholar]
- Wielkoszynski, T.; Moghaddam, A.; Bäckman, A.; Broden, J.; Piotrowski, R.; Mond-Paszek, R.; Kozarenko, A.; Ny, T.; Wilczynska, M. Novel diagnostic ELISA test for discrimination between infections with Yersinia enterocolitica and Yersinia pseudotuberculosis. Eur. J. Clin. Microbiol. Infect. Dis. 2018, 37, 2301–2306. [Google Scholar] [CrossRef]
- Nowalk, A.; Green, M. Epstein-Barr Virus. Microbiol. Spectr. 2016, 4, 1–8. [Google Scholar] [CrossRef]
- Cannon, M.J.; Schmid, D.S.; Hyde, T.B. Review of cytomegalovirus seroprevalence and demographic characteristics associated with infection. Rev. Med. Virol. 2010, 20, 202–213. [Google Scholar] [CrossRef]
- Goossens, H.; Nohlmans, M.K.; van den Bogaard, E. Epstein-Barr virus and cytomegalovirus infections cause false-positive results in IgM two-test protocol for early Lyme borreliosis. Infection 1999, 27, 231. [Google Scholar] [CrossRef] [PubMed]
- Vassilopoulos, D.; Calabrese, L.H. Virally associated arthritis 2008: Clinical, epidemiologic, and pathophysiologic considerations. Arthritis Res. Ther. 2008, 10, 215. [Google Scholar] [CrossRef] [PubMed]
- Tuuminen, T.; Hedman, K.; Söderlund-Venermo, M.; Seppälä, I. Acute parvovirus B19 infection causes nonspecificity frequently in Borrelia and less often in Salmonella and Campylobacter serology, posing a problem in diagnosis of infectious arthropathy. Clin. Vaccine Immunol. 2011, 18, 167–172. [Google Scholar] [CrossRef] [PubMed]
- Hsieh, Y.F.; Liu, H.W.; Hsu, T.C.; Wei, J.C.C.; Shih, C.M.; Krause, P.J.; Tsay, G.J. Serum reactivity against Borrelia burgdorferi OspA in patients with rheumatoid arthritis. Clin. Vaccine Immunol. 2007, 14, 1437–1441. [Google Scholar] [CrossRef] [PubMed]
- Renaudineau, Y.; JAMIN, C.; SARAUX, A.; PIERRE, Y. Rheumatoid factor on a daily basis. Autoimmunity 2005, 38, 11–16. [Google Scholar] [CrossRef]
- Magnarelli, L.A.; Ijdo, J.W.; Padula, S.J.; Flavell, R.A.; Fikrig, E. Serologic diagnosis of Lyme borreliosis by using enzyme-linked immunosorbent assays with recombinant antigens. J. Clin. Microbiol. 2000, 38, 1735–1739. [Google Scholar] [CrossRef] [PubMed]
- Tjernberg, I.; Krüger, G.; Eliasson, I. C6 peptide ELISA test in the serodiagnosis of Lyme borreliosis in Sweden. Eur. J. Clin. Microbiol. Infect. Dis. 2007, 26, 37–42. [Google Scholar] [CrossRef]
- Magnarelli, L.A.; Fikrig, E.; Padula, S.J.; Anderson, J.F.; Flavell, R.A. Use of recombinant antigens of Borrelia burgdorferi in serologic tests for diagnosis of Lyme borreliosis. J. Clin. Microbiol. 1996, 34, 237–240. [Google Scholar] [CrossRef]
- Luther, B.; Moskophidis, M. Antigenic Cross-reactivity between Borrelia burgdorferi, Borrelia recurrentis, Treponema pallidum, and Treponema phagedenis. Zentralblatt Bakteriol. 1990, 274, 214–226. [Google Scholar] [CrossRef]
- Bruckbauer, H.R.; Preac-Mursic, V.; Fuchs, R.; Wilske, B. Cross-reactive proteins of Borrelia burgdorferi. Eur. J. Clin. Microbiol. Infect. Dis. 1992, 11, 224–232. [Google Scholar] [CrossRef]
- Steere, A.C.; Drouin, E.E.; Glickstein, L.J. Relationship between immunity to Borrelia burgdorferi outer-surface protein A (OspA) and Lyme arthritis. Clin. Infect. Dis. 2011, 52, 259–265. [Google Scholar] [CrossRef] [PubMed]
- Ghosh, S.; Seward, R.; Costello, C.E.; Stollar, B.D.; Huber, B.T. Autoantibodies from synovial lesions in chronic, antibiotic treatment-resistant Lyme arthritis bind cytokeratin-10. J. Immunol. 2006, 177, 2486–2494. [Google Scholar] [CrossRef] [PubMed]
- Alaedini, A.; Latov, N. Antibodies against OspA epitopes of Borrelia burgdorferi cross-react with neural tissue. J. Neuroimmunol. 2005, 159, 192–195. [Google Scholar] [CrossRef] [PubMed]
- Arnaboldi, P.M.; Seedarnee, R.; Sambir, M.; Callister, S.M.; Imparato, J.A.; Dattwylera, R.J. Outer surface protein C peptide derived from Borrelia burgdorferi sensu stricto as a target for serodiagnosis of early Lyme disease. Clin. Vaccine Immunol. 2013, 20, 474–481. [Google Scholar] [CrossRef] [PubMed]
- Markowicz, M.; Reiter, M.; Gamper, J.; Stanek, G.; Stockinger, H. Persistent anti-Borrelia IgM antibodies without Lyme borreliosis in the clinical and immunological context. Microbiol. Spectr. 2021, 9, e01020-21. [Google Scholar] [CrossRef]
- Pallesen, L.; Hindersson, P. Cloning and sequencing of a Treponema pallidum gene encoding a 31.3-kilodalton endoflagellar subunit (FlaB2). Infect. Immun. 1989, 57, 2166–2172. [Google Scholar] [CrossRef]
- Hauser, U.; Wilske, B. Enzyme-linked immunosorbent assays with recombinant internal flagellin fragments derived from different species of Borrelia burgdorferi sensu lato for the serodiagnosis of Lyme neuroborreliosis. Med. Microbiol. Immunol. 1997 1862 1997, 186, 145–151. [Google Scholar] [CrossRef]
- Luft, B.J.; Dunn, J.J.; Dattwyler, R.J.; Gorgone, G.; Gorevic, P.D.; Schubach, W.H. Cross-reactive antigenic domains of the flagellin protein of Borrelia burgdorferi. Res. Microbiol. 1993, 144, 251–257. [Google Scholar] [CrossRef]
- Koetsveld, J.; Platonov, A.E.; Kuleshov, K.; Wagemakers, A.; Hoornstra, D.; Ang, W.; Szekeres, S.; van Duijvendijk, G.L.A.; Fikrig, E.; Embers, M.E.; et al. Borrelia miyamotoi infection leads to cross-reactive antibodies to the C6 peptide in mice and men. Clin. Microbiol. Infect. 2019, 26, 513.e1–513.e6. [Google Scholar] [CrossRef]
- Chandra, A.; Wormser, G.P.; Marques, A.R.; Latov, N.; Alaedini, A. Anti-Borrelia burgdorferi antibody profile in post-Lyme disease syndrome. Clin. Vaccine Immunol. 2011, 18, 767–771. [Google Scholar] [CrossRef]
- Eldin, C.; Raffetin, A.; Bouiller, K.; Hansmann, Y.; Roblot, F.; Raoult, D.; Parola, P. Review of European and American guidelines for the diagnosis of Lyme borreliosis. Med. Mal. Infect. 2019, 49, 121–132. [Google Scholar] [CrossRef] [PubMed]
- Mead, P.; Petersen, J.; Hinckley, A. Updated CDC recommendation for serologic diagnosis of Lyme disease. MMWR. Morb. Mortal. Wkly. Rep. 2019, 68, 703. [Google Scholar] [CrossRef] [PubMed]
- Burkot, T.R.; Schriefer, M.E.; Larsen, S.A. Cross-reactivity to Borrelia burgdorferi Proteins in serum samples from residents of a tropical country nonendemic for Lyme disease. J. Infect. Dis. 1997, 175, 466–469. [Google Scholar] [CrossRef] [PubMed]
- Golkocheva-Markova, E.; Christova, I.; Stoilov, R.; Najdenski, H. Cross-reaction between Yersinia outer membrane proteins and anti- Borrelia antibodies in sera of patients with Lyme disease. Clin. Microbiol. Infect. 2008, 14, 873–875. [Google Scholar] [CrossRef] [PubMed]
- Del-Rei, R.P.; Leony, L.M.; Celedon, P.A.F.; Zanchin, N.I.T.; dos Reis, M.G.; de Miranda Gomes, Y.; Schijman, A.G.; Longhi, S.A.; Santos, F.L.N. Detection of anti-Trypanosoma cruzi antibodies by chimeric antigens in chronic Chagas disease-individuals from endemic South American countries. PLoS ONE 2019, 14, e0215623. [Google Scholar] [CrossRef] [PubMed]
- Holec-Gąsior, L.; Ferra, B.; Drapała, D.; Lautenbach, D.; Kur, J. A new MIC1-MAG1 recombinant chimeric antigen can be used instead of the Toxoplasma gondii lysate antigen in serodiagnosis of human toxoplasmosis. Clin. Vaccine Immunol. 2012, 19, 57–63. [Google Scholar] [CrossRef]
- Grąźlewska, W.; Ferra, B.; Rudzińska, M.; Holec-Gąsior, L. Borrelia burgdorferi BmpA-BBK32 and BmpA-BBA64: New recombinant chimeric proteins with potential diagnostic value. Pathogens 2021, 10, 767. [Google Scholar] [CrossRef]
- Gomes-Solecki, M.J.C.; Dunn, J.J.; Luft, B.J.; Castillo, J.; Dykhuizen, D.E.; Yang, X.; Glass, D.; Dattwyler, R.J.; Glass, J.D. Recombinant chimeric Borrelia proteins for diagnosis of Lyme disease. J. Clin. Microbiol. 2000, 38, 2530–2535. [Google Scholar] [CrossRef]
- Bateman, A.; Martin, M.J.; Orchard, S.; Magrane, M.; Agivetova, R.; Ahmad, S.; Alpi, E.; Bowler-Barnett, E.H.; Britto, R.; Bursteinas, B.; et al. UniProt: The universal protein knowledgebase in 2021. Nucleic Acids Res. 2021, 49, 480–489. [Google Scholar] [CrossRef]
- Sharma, S.; Ciufo, S.; Starchenko, E.; Darji, D.; Chlumsky, L.; Karsch-Mizrachi, I.; Schoch, C.L. The NCBI BioCollections Database. Database 2018, 2018, 6. [Google Scholar] [CrossRef]
- Altschul, S.F.; Gish, W.; Miller, W.; Myers, E.W.; Lipman, D.J. Basic local alignment search tool. J. Mol. Biol. 1990, 215, 403–410. [Google Scholar] [CrossRef] [PubMed]
- Larkin, M.A.; Blackshields, G.; Brown, N.P.; Chenna, R.; Mcgettigan, P.A.; McWilliam, H.; Valentin, F.; Wallace, I.M.; Wilm, A.; Lopez, R.; et al. Clustal W and Clustal X version 2.0. Bioinformatics 2007, 23, 2947–2948. [Google Scholar] [CrossRef] [PubMed]
- Wormser, G.P.; Schriefer, M.; Aguero-Rosenfeld, M.E.; Levin, A.; Steere, A.C.; Nadelman, R.B.; Nowakowski, J.; Marques, A.; Johnson, B.J.B.; Dumler, J.S. Single-tier testing with the C6 peptide ELISA kit compared with two-tier testing for Lyme disease. Diagn. Microbiol. Infect. Dis. 2013, 75, 9–15. [Google Scholar] [CrossRef] [PubMed]
- Glatz, M.; Fingerle, V.; Wilske, B.; Ambros-Rudolph, C.; Kerl, H.; Müllegger, R.R. Immunoblot analysis of the seroreactivity to recombinant Borrelia burgdorferi sensu lato antigens, including VlsE, in the long-term course of treated patients with erythema migrans. Dermatology 2008, 216, 93–103. [Google Scholar] [CrossRef]
- Toumanios, C.; Prisco, L.; Dattwyler, R.J.; Arnaboldi, P.M. Linear B cell epitopes derived from the multifunctional surface lipoprotein BBK32 as targets for the serodiagnosis of Lyme disease. mSphere 2019, 4, 10–1128. [Google Scholar] [CrossRef]
- Arnaboldi, P.M.; Katseff, A.S.; Sambir, M.; Dattwyler, R.J. Linear peptide epitopes derived from ErpP, p35, and FlaB in the serodiagnosis of Lyme disease. Pathogens 2022, 11, 944. [Google Scholar] [CrossRef]
- Ahmad, T.A.; Eweida, A.E.; Sheweita, S.A. B-cell epitope mapping for the design of vaccines and effective diagnostics. Trials Vaccinol. 2016, 5, 71–83. [Google Scholar] [CrossRef]
- Potocnakova, L.; Bhide, M.; Pulzova, L.B. An Introduction to B-Cell epitope mapping and in silico epitope prediction. J. Immunol. Res. 2016, 2016, 6760830. [Google Scholar] [CrossRef]
- Maksimov, P.; Zerweck, J.; Maksimov, A.; Hotop, A.; Groß, U.; Pleyer, U.; Spekker, K.; Däubener, W.; Werdermann, S.; Niederstrasser, O.; et al. Peptide microarray analysis of in silico-predicted epitopes for serological diagnosis of Toxoplasma gondii infection in humans. Clin. Vaccine Immunol. 2012, 19, 865–874. [Google Scholar] [CrossRef]
- Heiss, K.; Heidepriem, J.; Fischer, N.; Weber, L.K.; Dahlke, C.; Jaenisch, T.; Loeffler, F.F. Rapid response to pandemic threats: Immunogenic epitope detection of pandemic pathogens for diagnostics and vaccine development using peptide microarrays. J. Proteome Res. 2020, 19, 4339–4354. [Google Scholar] [CrossRef]
- Bahrami, A.A.; Payandeh, Z.; Khalili, S.; Zakeri, A.; Bandehpour, M. Immunoinformatics: In silico approaches and computational design of a multi-epitope, immunogenic protein. Int. Rev. Immunol. 2019, 38, 307–322. [Google Scholar] [CrossRef] [PubMed]
- Gershoni, J.M.; Roitburd-Berman, A.; Siman-Tov, D.D.; Freund, N.T.; Weiss, Y. Epitope mapping: The first step in developing epitope-based vaccines. BioDrugs 2007, 21, 145–156. [Google Scholar] [CrossRef] [PubMed]
- Vigan-Womas, I.; Spadoni, J.L.; Poiret, T.; Taïeb, F.; Randrianarisaona, F.; Faye, R.; Mbow, A.A.; Gaye, A.; Dia, N.; Loucoubar, C.; et al. Linear epitope mapping of the humoral response against SARS-CoV-2 in two independent African cohorts. Sci. Rep. 2023, 13, 782. [Google Scholar] [CrossRef] [PubMed]


{kind=link}
| Bbsl Life Cycle Stage | Protein | Gene Localization | Information | Refs. |
|---|---|---|---|---|
| Ticks | OspA (p30) | Plasmid lp54 |
| [38,39] |
| OspB | Plasmid lp54 |
| [38,39] | |
| OspD (p28) | Plasmid lp38 |
| [40] | |
| Transmission | OspC (p23–25) | Plasmid cp26 |
| [21,41,42] |
| BBA64 (p35) | Plasmid lp54 |
| [43,44] | |
| Mammals | DbpA/B (p18) | Plasmid lp54 |
| [21,45] |
| BBK32 | Plasmid lp36 |
| [21,46,47] | |
| VlsE | Plasmid lp28-1 |
| [48,49] | |
| BmpA (p39) | Chromosome |
| [50,51,52] | |
| OspE-related proteins (OspF, ErpA, ErpC, ErpP) | Plasmid cp32 |
| [53,54,55] | |
| p66 | Chromosome |
| [56] | |
| LA7 (p22) | Chromosome |
| [33,57] | |
| OppA2 | Chromosome |
| [58] | |
| Ticks and mammals | FlaA (p37) | Chromosome |
| [59,60] |
| FlaB (p41) | Chromosome |
| [61,62] |
| WCL | LA7 (p22) | GroEL (p60) | FlaB (p41) | BBA64 (p35) | FlaA (p37) | BmpA (p39) | OppA2 | VlsE | pC6 | OspA (p30) | OspB | OspC (p23–25) | OspD (p28) | OspE | OspF | Refs. | |
|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|
| RFB | IgM+ IgG+ | Nd | Rabbit Ig | IgM+ IgG+ | IgM− IgG− | IgM+ IgG+ | IgM+ IgG+ | Nd | IgM+ IgG+ | Nd | Rabbit Ig | Rabbit Ig | IgM+ IgG+ | Nd | IgM− IgG− | IgM+ IgG+ | [86,110] |
| Syphylis | IgM+ IgG+ | IgM+ IgG+ | Nd | IgM+ IgG+ | IgM+ IgG+ | IgM+ IgG+ | IgM+ IgG− | IgM/ IgG+ | IgM− | Nd | IgM− IgG− | IgM− IgG− | IgM+ IgG− | Nd | IgM− IgG− | IgM+ IgG+ | [58,60,86,87,108] |
| Yersinia | Nd | Nd | Nd | IgM+ IgG+ | Nd | Nd | IgM+ IgG+ | Nd | Nd | Nd | IgMNd IgG+ | Nd | IgM+ IgG+ | IgMNd IgG− | Nd | Nd | [96] |
| HGA | IgM- IgG- | IgM+ IgG+ | Nd | IgM+ IgG+ | IgM+ IgG− | IgM+ IgG+ | IgM− IgG− | Nd | IgM+ IgG− | Nd | IgM+ IgG+ | IgM− IgG− | IgM+ IgG− | Nd | IgM− IgG− | IgM+ IgG+ | [86] |
| EBV | Nd | Nd | Nd | IgM+ IgGNd | Nd | IgM+ IgG+ | Nd | Nd | Nd | IgM/IgG+ | Nd | Nd | IgM+ IgG+ | Nd | Nd | Nd | [60,87,107] |
| CMV | IgM+ IgG+ | Nd | Nd | IgM+ IgG− | Nd | Nd | Nd | Nd | Nd | Nd | Nd | Nd | IgM+ IgG− | Nd | Nd | Nd | [83,84] |
| B19V | Nd | Nd | Nd | Nd | Nd | Nd | IgM+ IgGNd | Nd | IgM+ IgGNd | Nd | Nd | Nd | Nd | Nd | Nd | Nd | [103] |
| RA/RF | IgM+ IgG+ | IgM+ IgG+ | Nd | IgM− IgG− | IgM− IgG+ | IgM+ IgG+ | IgM+ IgG+ | IgM/IgG+ | Nd | IgM/IgG+ | IgM− IgG− | IgM− IgG− | IgM− IgG− | Nd | IgM+ IgG− | IgM− IgG− | [60,86,87,106,107,108] |
| HT | Nd | Nd | Nd | Nd | Nd | Nd | Nd | Nd | Nd | Nd | IgMNd IgG+ | Nd | Nd | Nd | Nd | Nd | [109,110] |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Grąźlewska, W.; Holec-Gąsior, L. Antibody Cross-Reactivity in Serodiagnosis of Lyme Disease. Antibodies 2023, 12, 63. https://doi.org/10.3390/antib12040063
Grąźlewska W, Holec-Gąsior L. Antibody Cross-Reactivity in Serodiagnosis of Lyme Disease. Antibodies. 2023; 12(4):63. https://doi.org/10.3390/antib12040063
Chicago/Turabian StyleGrąźlewska, Weronika, and Lucyna Holec-Gąsior. 2023. "Antibody Cross-Reactivity in Serodiagnosis of Lyme Disease" Antibodies 12, no. 4: 63. https://doi.org/10.3390/antib12040063
APA StyleGrąźlewska, W., & Holec-Gąsior, L. (2023). Antibody Cross-Reactivity in Serodiagnosis of Lyme Disease. Antibodies, 12(4), 63. https://doi.org/10.3390/antib12040063