

Characterization of an IDH1 R132H Rabbit Monoclonal Antibody, MRQ-67, and Its Applications in the Identification of Diffuse Gliomas
 ,
,
Abstract
1. Introduction
2. Materials and Methods
2.1. Tissue Specimens
2.2. IDH1/2 Peptides
2.3. ELISA
2.4. Dot Immunoassay
2.5. Cell Lines and Western Blotting
2.6. Cell Pellet Block Preparation
2.7. Immunohistochemical Testing and Result Assessment
2.8. IDH1 Genotyping
2.9. Statistical Analysis
3. Results
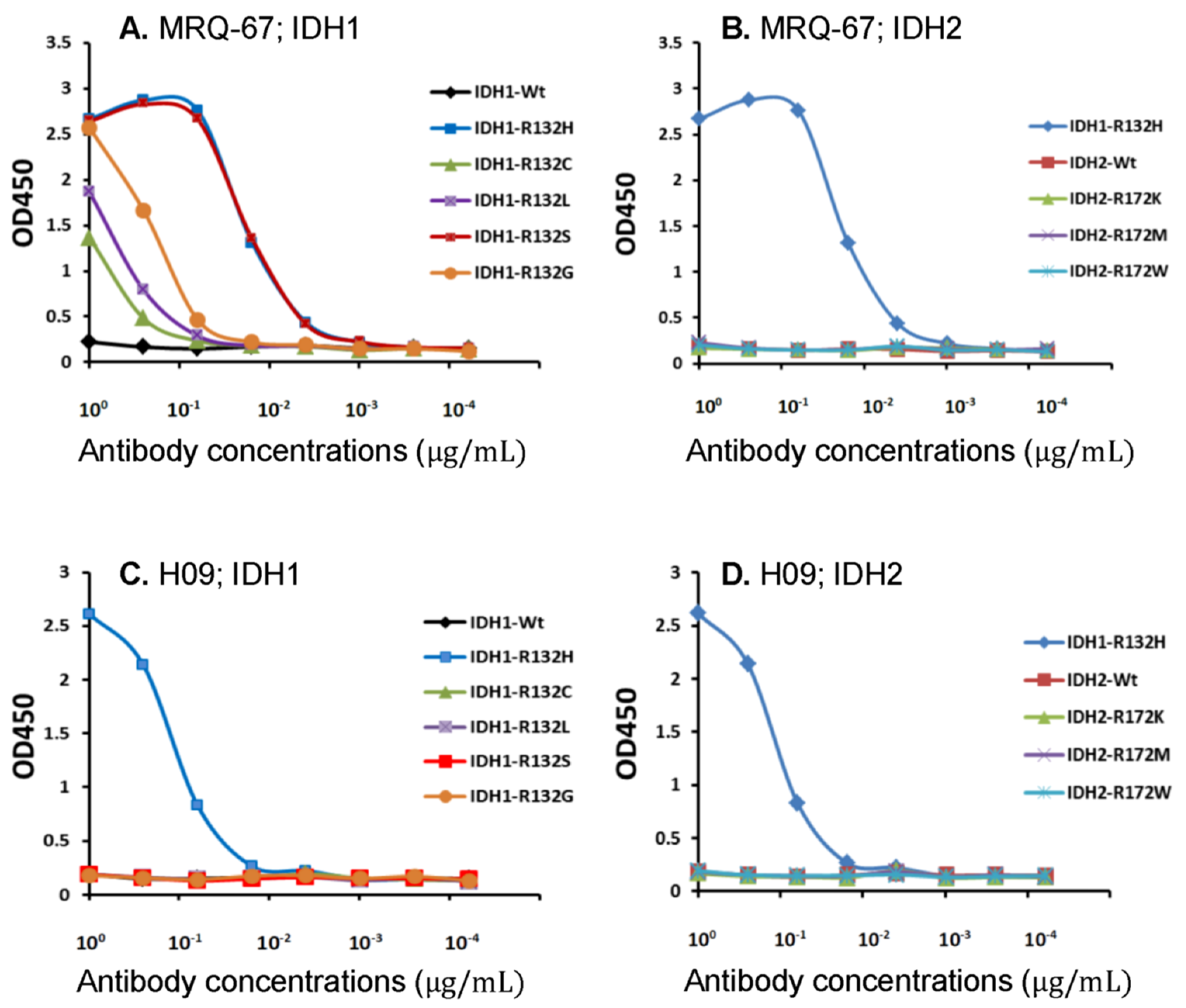
3.1. Binding of MRQ-67 to IDH1/2 Peptides as Demonstrated by ELISA
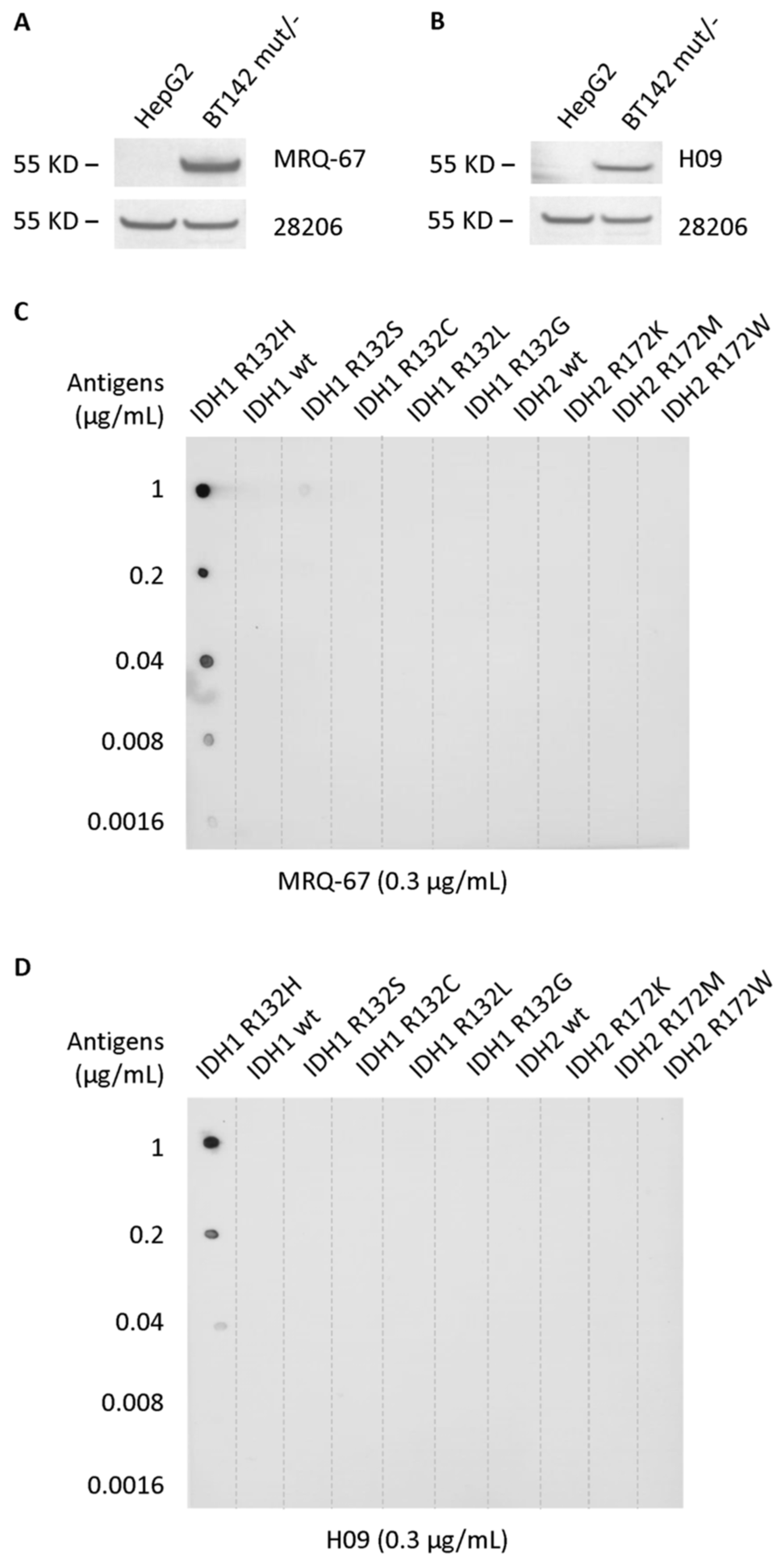
3.2. Specific Reaction of MRQ-67 to the IDH1 R132H Mutant Peptide on Blots
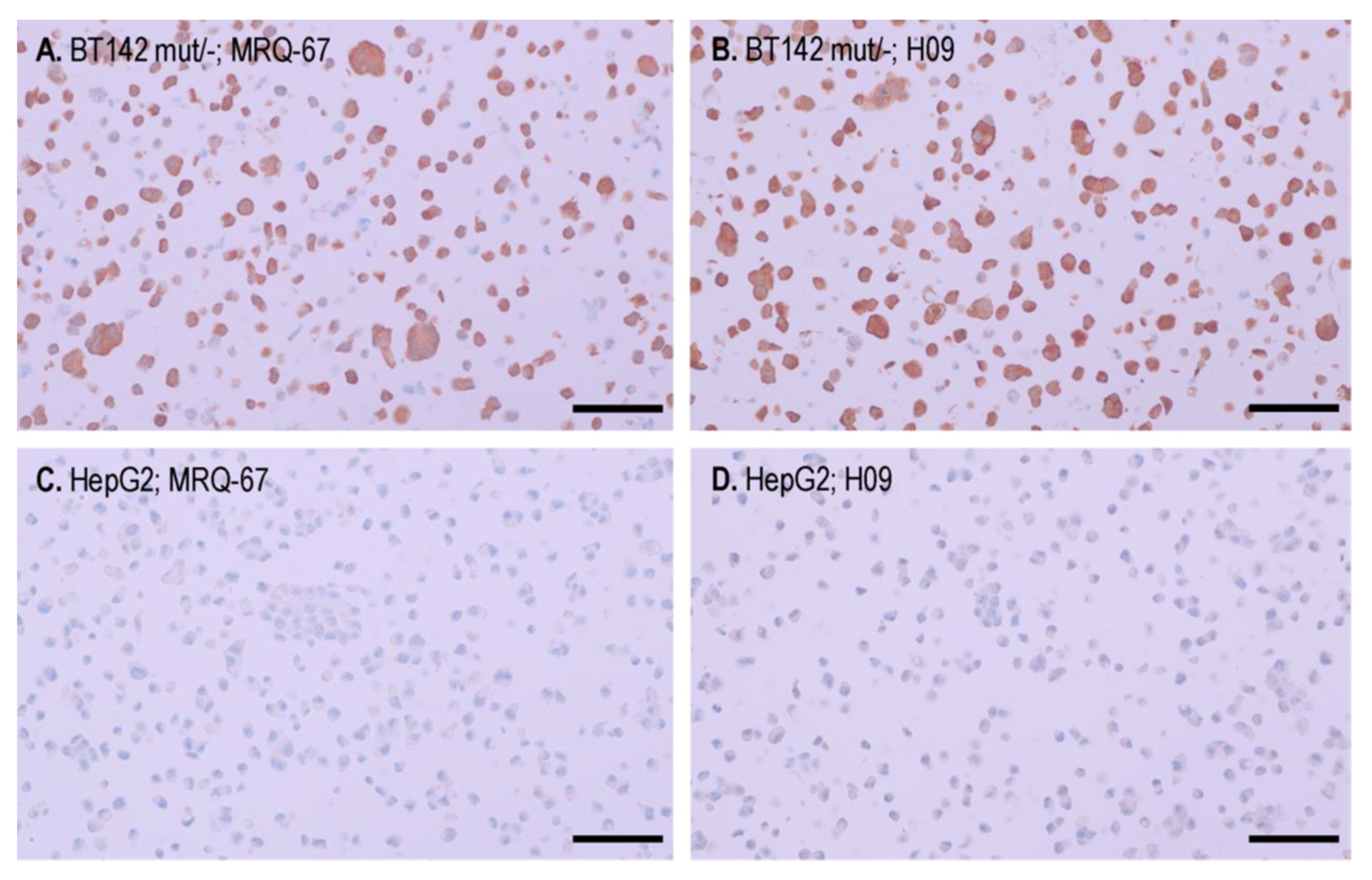
3.3. IHC Binding of MRQ-67 on the IDH1 R132H Mutant Tumor Cells in Cell Pellet Blocks
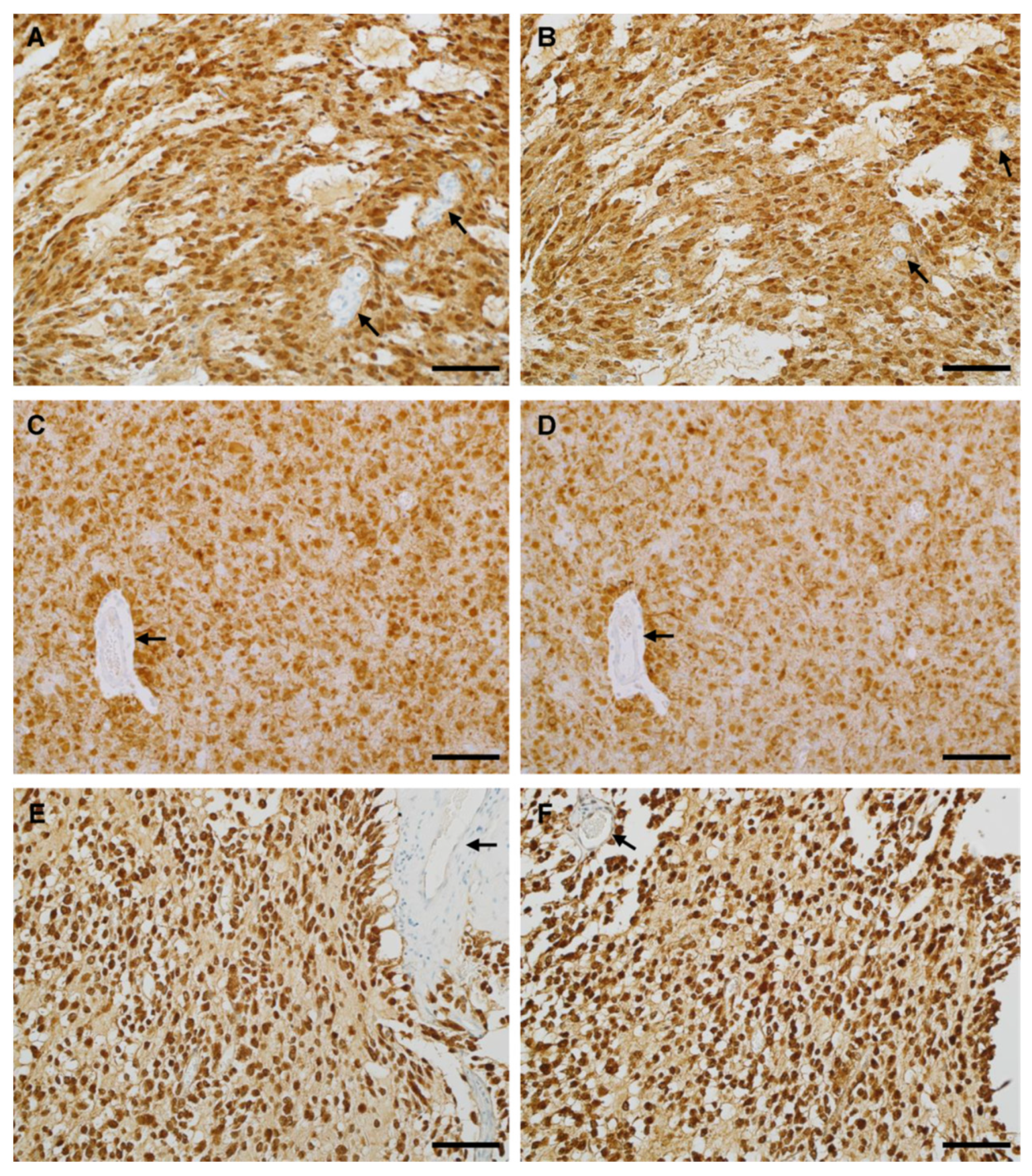
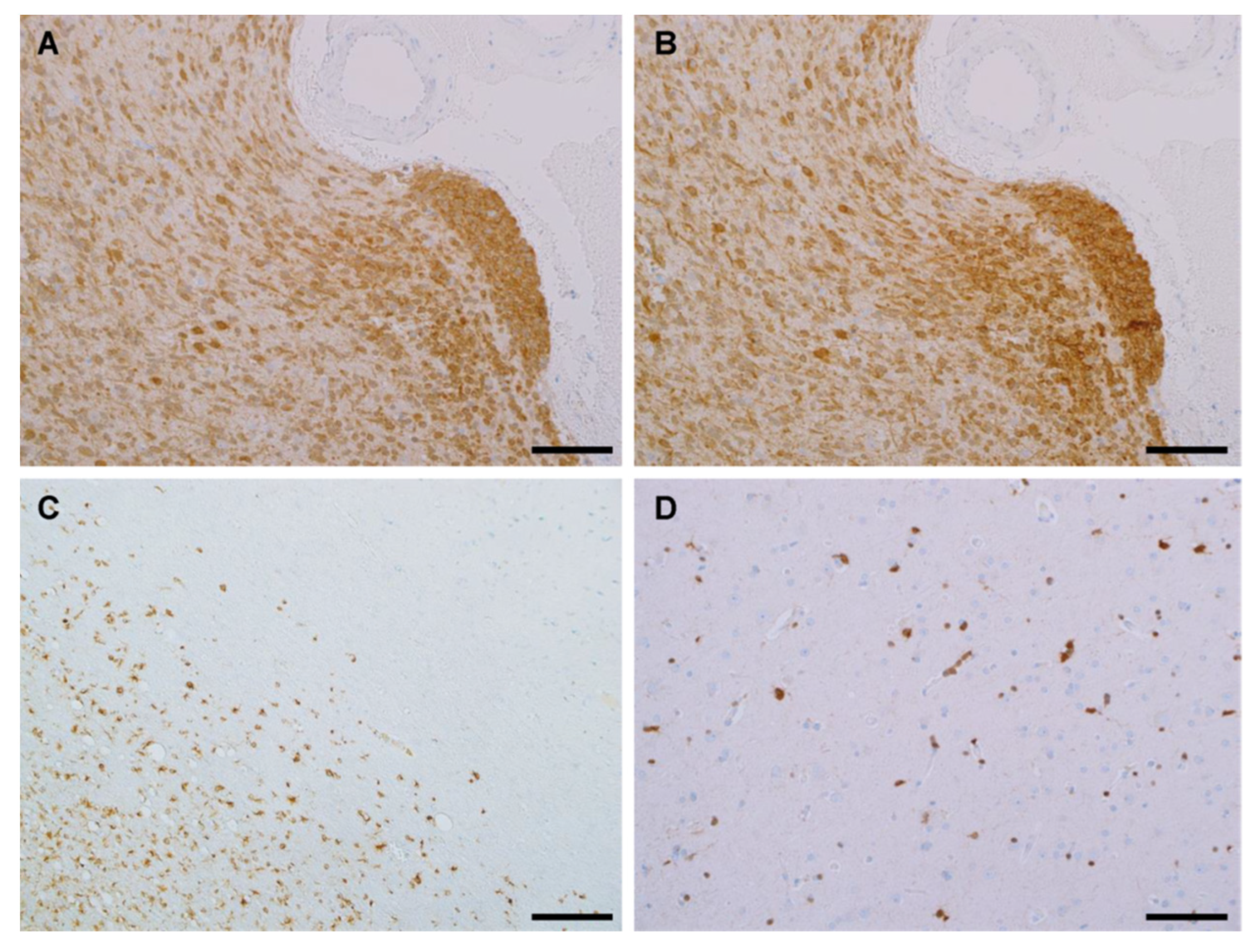
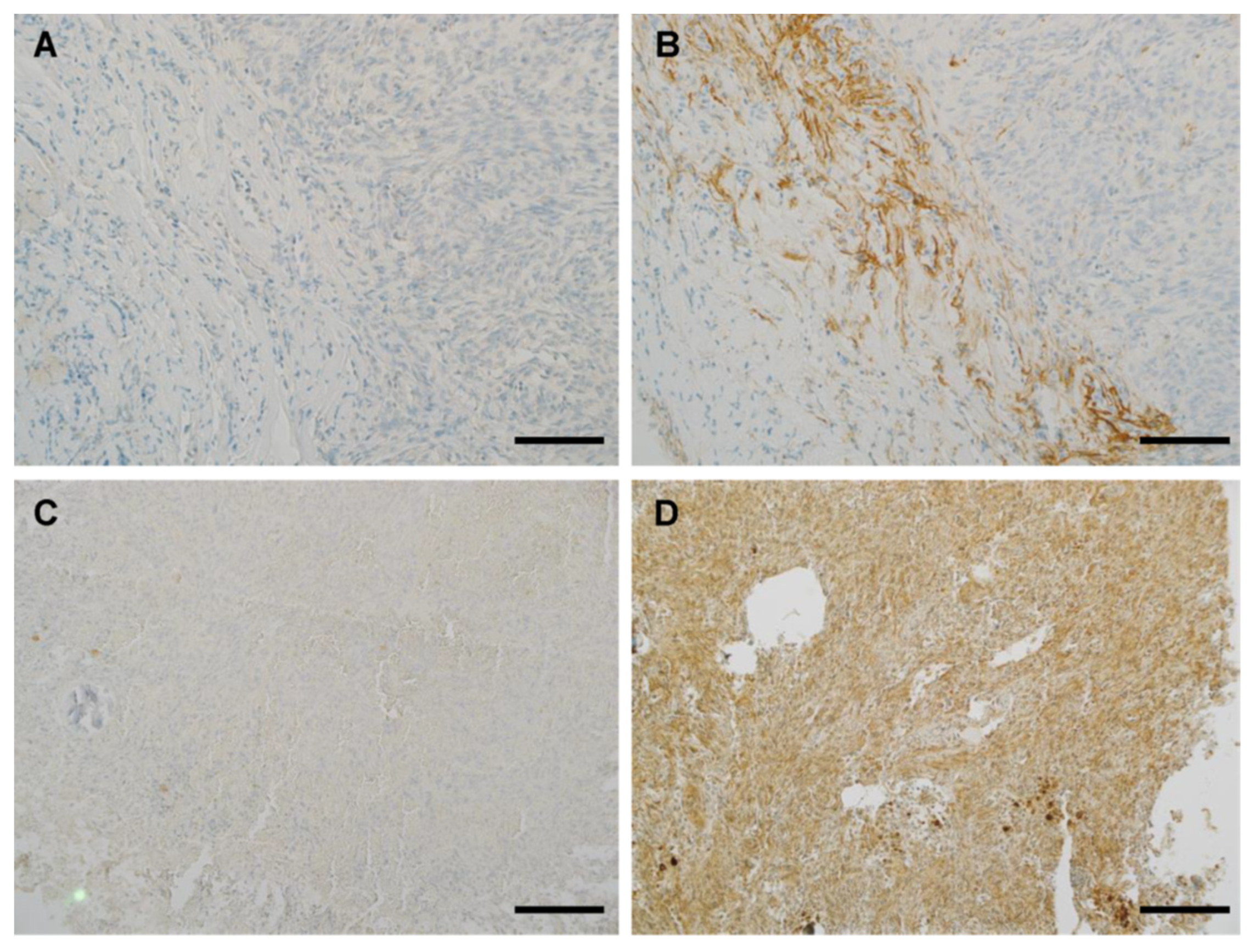
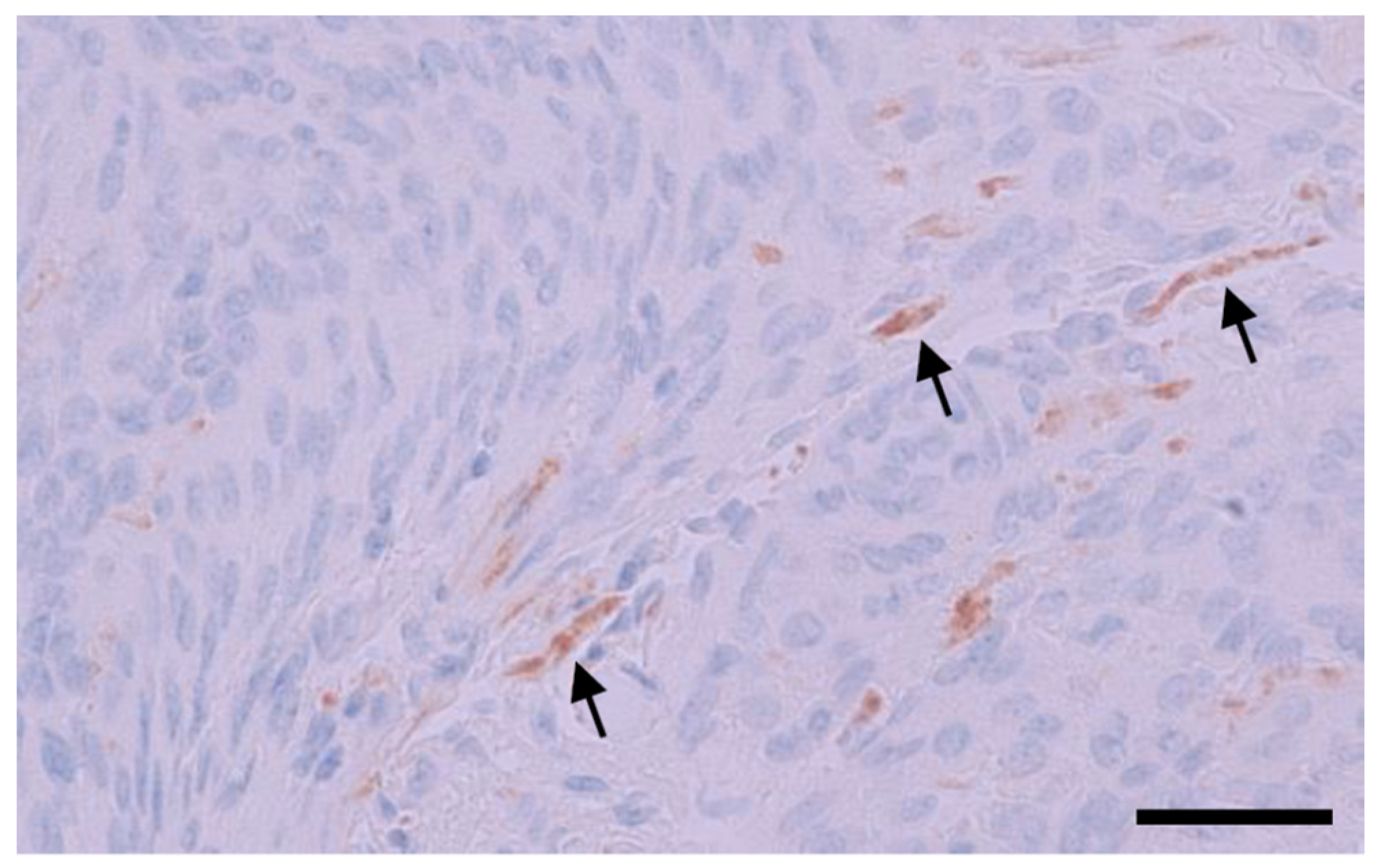
3.4. IHC Binding of MRQ-67 in Diffuse Gliomas
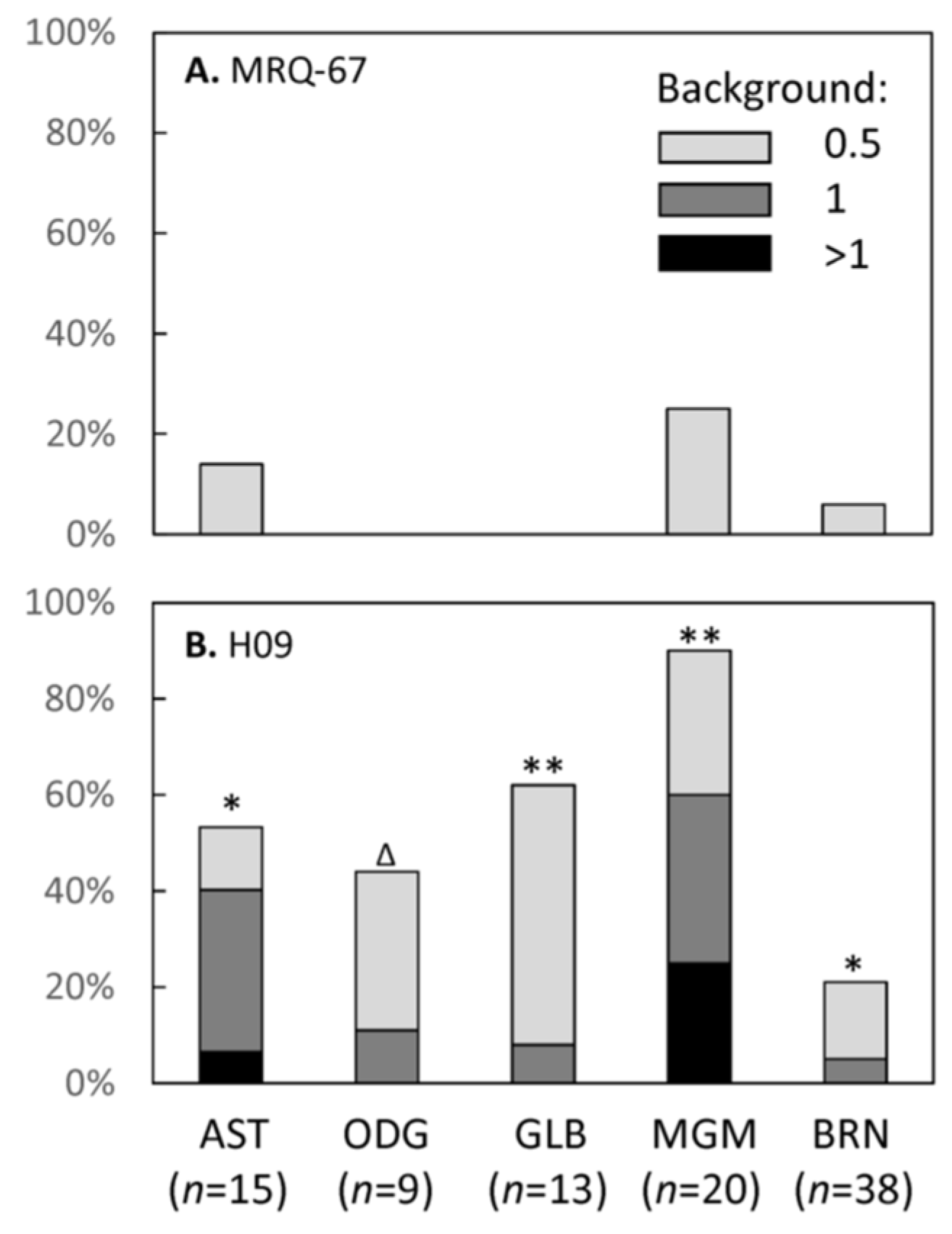
3.5. IHC Performance of MRQ-67 in Comparison with H09
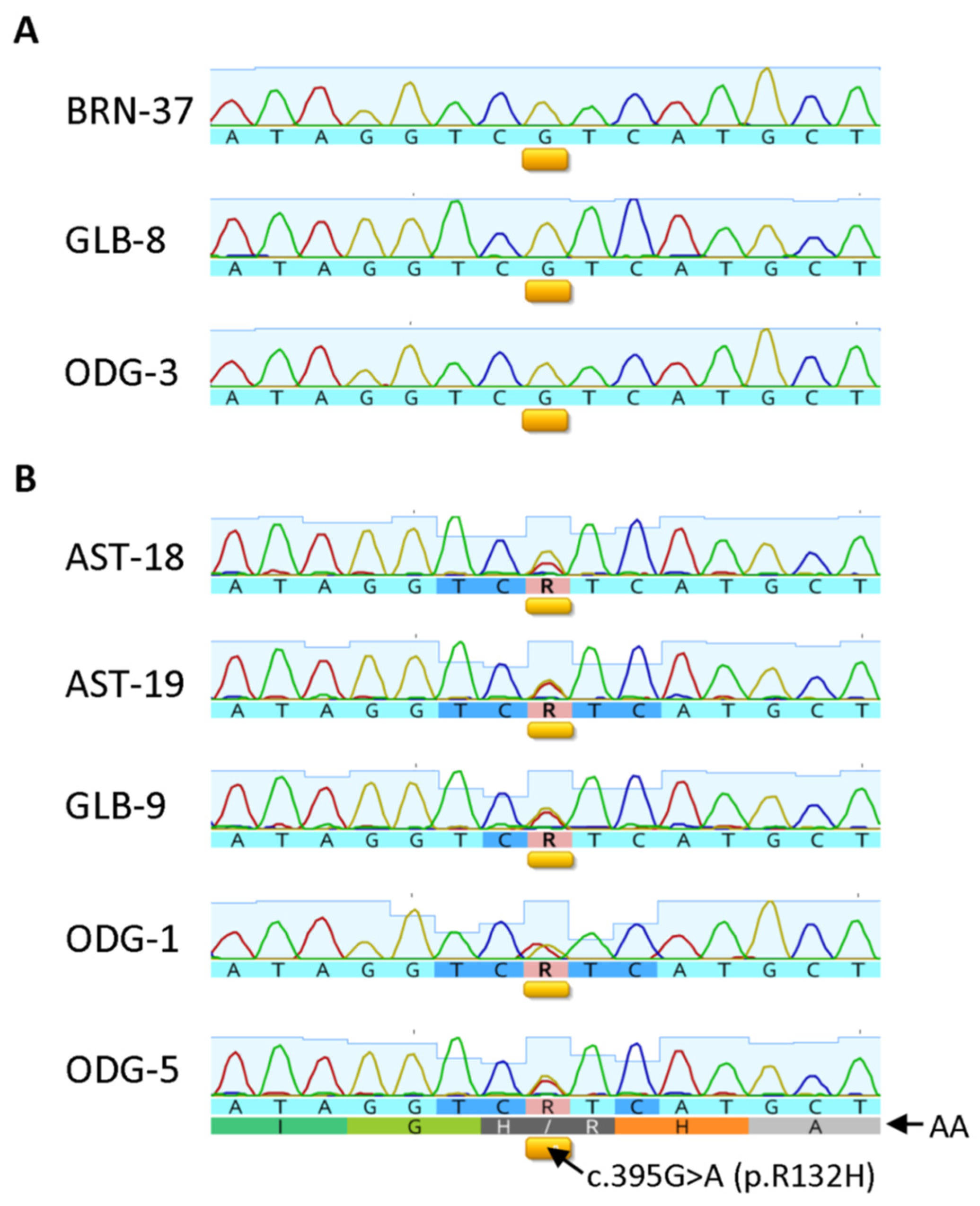
3.6. Correlation between IHC and Sequencing Results in Selective Cases
4. Discussion
5. Patents
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Louis, D.N.; von Deimling, A.; Cavenee, W.K. Diffuse astrocytic and oligodendroglial tumors—Introduction. In WHO Classification of Tumors of the Central Nervous System, 4th ed.; Louis, D.N., Ohgaki, H., Wiestler, O.D., Cavenee, W.K., Eds.; International Agency for Research on Cancer: Lyon, France, 2016; pp. 16–17. [Google Scholar]
- Louis, D.N.; Perry, A.; Reifenberger, G.; von Deimling, A.; Figarella-Branger, D.; Cavenee, W.K.; Ohgaki, H.; Wiestler, O.D.; Kleihues, P.; Ellison, D.W. The 2016 World Health Organization Classification of Tumors of the Central Nervous System: A summary. Acta Neuropathol. 2016, 131, 803–820. [Google Scholar] [CrossRef] [PubMed]
- Louis, D.N.; Perry, A.; Wesseling, P.; Brat, D.J.; Cree, I.A.; Figarella-Branger, D.; Hawkins, C.; Ng, H.K.; Pfister, S.M.; Reifenberger, G.; et al. The 2021 WHO Classification of Tumors of the Central Nervous System: A summary. Neuro Oncol. 2021, 23, 1231–1251. [Google Scholar] [CrossRef] [PubMed]
- Rushing, E.J. WHO classification of tumors of the nervous system: Preview of the upcoming 5th edition. MEMO 2021, 14, 188–191. [Google Scholar] [CrossRef]
- Komori, T. Grading of adult diffuse gliomas according to the 2021 WHO Classification of Tumors of the Central Nervous System. Lab. Investig. 2022, 102, 126–133. [Google Scholar] [CrossRef] [PubMed]
- Cancer Genome Atlas Research Network; Brat, D.J.; Verhaak, R.G.; Aldape, K.D.; Yung, W.K.; Salama, S.R.; Cooper, L.A.; Rheinbay, E.; Miller, C.R.; Vitucci, M.; et al. Comprehensive, integrative genomic analysis of diffuse lower-grade gliomas. N. Engl. J. Med. 2015, 372, 2481–2498. [Google Scholar] [CrossRef]
- Reuss, D.E.; Sahm, F.; Schrimpf, D.; Wiestler, B.; Capper, D.; Koelsche, C.; Schweizer, L.; Korshunov, A.; Jones, D.T.; Hovestadt, V.; et al. ATRX and IDH1-R132H immunohistochemistry with subsequent copy number analysis and IDH sequencing as a basis for an “integrated” diagnostic approach for adult astrocytoma, oligodendroglioma and glioblastoma. Acta Neuropathol. 2015, 129, 133–146. [Google Scholar] [CrossRef] [PubMed]
- Pekmezci, M.; Rice, T.; Molinaro, A.M.; Walsh, K.M.; Decker, P.A.; Hansen, H.; Sicotte, H.; Kollmeyer, T.M.; McCoy, L.S.; Sarkar, G.; et al. Adult infiltrating gliomas with WHO 2016 integrated diagnosis: Additional prognostic roles of ATRX and TERT. Acta Neuropathol. 2017, 133, 1001–1016. [Google Scholar] [CrossRef] [PubMed]
- DeWitt, J.C.; Jordan, J.T.; Frosch, M.P.; Samore, W.R.; Iafrate, A.J.; Louis, D.N.; Lennerz, J.K. Cost-effectiveness of IDH testing in diffuse gliomas according to the 2016 WHO classification of tumors of the central nervous system recommendations. Neuro Oncol. 2017, 19, 1640–1650. [Google Scholar] [CrossRef] [PubMed]
- Hartmann, C.; Meyer, J.; Balss, J.; Capper, D.; Mueller, W.; Christians, A.; Felsberg, J.; Wolter, M.; Mawrin, C.; Wick, W.; et al. Type and frequency of IDH1 and IDH2 mutations are related to astrocytic and oligodendroglial differentiation and age: A study of 1,010 diffuse gliomas. Acta Neuropathol. 2009, 118, 469–474. [Google Scholar] [CrossRef] [PubMed]
- Yan, H.; Parsons, D.W.; Jin, G.; McLendon, R.; Rasheed, B.A.; Yuan, W.; Kos, I.; Batinic-Haberle, I.; Jones, S.; Riggins, G.J.; et al. IDH1 and IDH2 mutations in gliomas. N. Engl. J. Med. 2009, 360, 765–773. [Google Scholar] [CrossRef] [PubMed]
- Chen, L.; Voronovich, Z.; Clark, K.; Hands, I.; Mannas, J.; Walsh, M.; Nikiforova, M.N.; Durbin, E.B.; Weiss, H.; Horbinski, C. Predicting the likelihood of an isocitrate dehydrogenase 1 or 2 mutation in diagnoses of infiltrative glioma. Neuro Oncol. 2014, 16, 1478–1483. [Google Scholar] [CrossRef] [PubMed]
- Chen, N.; Yu, T.; Gong, J.; Nie, L.; Chen, X.; Zhang, M.; Xu, M.; Tan, J.; Su, Z.; Zhong, J.; et al. IDH1/2 gene hotspot mutations in central nervous system tumours: Analysis of 922 Chinese patients. Pathology 2016, 48, 675–683. [Google Scholar] [CrossRef] [PubMed]
- Robinson, C.; Kleinschmidt-DeMasters, B.K. IDH1-mutation in diffuse gliomas in persons age 55 years and over. J. Neuropathol. Exp. Neurol. 2017, 76, 151–154. [Google Scholar] [CrossRef] [PubMed]
- Preusser, M.; Wöhrer, A.; Stary, S.; Höftberger, R.; Streubel, B.; Hainfellner, J.A. Value and limitations of immunohistochemistry and gene sequencing for detection of the IDH1-R132H mutation in diffuse glioma biopsy specimens. J. Neuropathol. Exp. Neurol. 2011, 70, 715–723. [Google Scholar] [CrossRef] [PubMed]
- Agarwal, S.; Sharma, M.C.; Jha, P.; Pathak, P.; Suri, V.; Sarkar, C.; Chosdol, K.; Suri, A.; Kale, S.S.; Mahapatra, A.K.; et al. Comparative study of IDH1 mutations in gliomas by immunohistochemistry and DNA sequencing. Neuro Oncol. 2013, 15, 718–726. [Google Scholar] [CrossRef]
- Macauly, R.J. Impending Impact of Molecular Pathology on Classifying Adult Diffuse Gliomas. Cancer Control 2015, 22, 200–205. [Google Scholar] [CrossRef] [PubMed]
- Capper, D.; Zentgraf, H.; Balss, J.; Hartmann, C.; von Deimling, A. Monoclonal antibody specific for IDH1 R132H mutation. Acta Neuropathol. 2009, 118, 599–601. [Google Scholar] [CrossRef] [PubMed]
- Kato, Y.; Jin, G.; Kuan, C.T.; McLendon, R.E.; Yan, H.; Bigner, D.D. A monoclonal antibody IMab-1 specifically recognizes IDH1R132H, the most common glioma-derived mutation. Biochem. Biophys. Res. Commun. 2009, 390, 547–551. [Google Scholar] [CrossRef]
- Kato, Y. Specific monoclonal antibodies against IDH1/2 mutations as diagnostic tools for gliomas. Brain Tumor Pathol. 2015, 32, 3–11. [Google Scholar] [CrossRef] [PubMed]
- Shayanfar, N.; Zare-Mirzaie, A.; Mohammadpour, M.; Jafari, E.; Mehrtash, A.; Emtiazi, N.; Tajik, F. Low expression of isocitrate dehydrogenase 1 (IDH1) R132H is associated with advanced pathological features in laryngeal squamous cell carcinoma. J. Cancer Res. Clin. Oncol. 2022, in press. [Google Scholar] [CrossRef] [PubMed]
- Capper, D.; Reuss, D.; Schittenhelm, J.; Hartmann, C.; Bremer, J.; Sahm, F.; Harter, P.N.; Jeibmann, A.; von Deimling, A. Mutation-specific IDH1 antibody differentiates oligodendrogliomas and oligoastrocytomas from other brain tumors with oligodendroglioma-like morphology. Acta Neuropathol. 2011, 121, 241–252. [Google Scholar] [CrossRef] [PubMed]
- Sahm, F.; Capper, D.; Meyer, J.; Hartmann, C.; Herpel, E.; Andrulis, M.; Mechtersheimer, G.; Petersen, I.; Paulus, W.; von Deimling, A. Immunohistochemical analysis of 1844 human epithelial and haematopoietic tumours and sarcomas for IDH1R132H mutation. Histopathology 2011, 58, 1167–1172. [Google Scholar] [CrossRef]
- Kurt, H.; Bueso-Ramos, C.E.; Khoury, J.D.; Routbort, M.J.; Kanagal-Shamanna, R.; Patel, U.V.; Jorgensen, J.L.; Wang, S.A.; Ravandi, F.; DiNardo, C.; et al. Characterization of IDH1 p.R132H Mutant Clones Using Mutation-specific Antibody in Myeloid Neoplasms. Am. J. Surg. Pathol. 2018, 42, 569–577. [Google Scholar] [CrossRef]
- Arita, H.; Narita, Y.; Yoshida, A.; Hashimoto, N.; Yoshimine, T.; Ichimura, K. IDH1/2 mutation detection in gliomas. Brain Tumor Pathol. 2015, 32, 79–89. [Google Scholar] [CrossRef] [PubMed]
- Ikota, H.; Nobusawa, S.; Arai, H.; Kato, Y.; Ishizawa, K.; Hirose, T.; Yokoo, H. Evaluation of IDH1 status in diffusely infiltrating gliomas by immunohistochemistry using anti-mutant and wild type IDH1 antibodies. Brain Tumor Pathol. 2015, 32, 237–244. [Google Scholar] [CrossRef] [PubMed]
- Nishikawa, T.; Watanabe, R.; Kitano, Y.; Yamamichi, A.; Motomura, K.; Ohka, F.; Aoki, K.; Hirano, M.; Kato, A.; Yamaguchi, J.; et al. Reliability of IDH1-R132H and ATRX and/or p53 immunohistochemistry for molecular subclassification of Grade 2/3 gliomas. Brain Tumor Pathol. 2022, 39, 14–24. [Google Scholar] [CrossRef]
- Capper, D.; Weissert, S.; Balss, J.; Habel, A.; Meyer, J.; Jäger, D.; Ackermann, U.; Tessmer, C.; Korshunov, A.; Zentgraf, H.; et al. Characterization of R132H mutation-specific IDH1 antibody binding in brain tumors. Brain Pathol. 2010, 20, 245–254. [Google Scholar] [CrossRef] [PubMed]
- Rashidian, J.; Copaciu, R.; Su, Q.; Merritt, B.; Johnson, C.; Yahyabeik, A.; French, E.; Cummings, K. Generation and Performance of R132H Mutant IDH1 Rabbit Monoclonal Antibody. Antibodies 2017, 6, 22. [Google Scholar] [CrossRef]
- Pennanen, M.; Tynninen, O.; Kytölä, S.; Ellonen, P.; Mustonen, H.; Heiskanen, I.; Haglund, C.; Arola, J. IDH1 Expression via the R132H Mutation-Specific Antibody in Adrenocortical Neoplasias-Prognostic Impact in Carcinomas. J. Endocr. Soc. 2020, 4, bvaa018. [Google Scholar] [CrossRef] [PubMed]
- Yoshida, A.; Satomi, K.; Ohno, M.; Matsushita, Y.; Takahashi, M.; Miyakita, Y.; Hiraoka, N.; Narita, Y.; Ichimura, K. Frequent false-negative immunohistochemical staining with IDH1 (R132H)-specific H09 antibody on frozen section control slides: A potential pitfall in glioma diagnosis. Histopathology 2019, 74, 350–354. [Google Scholar] [CrossRef]
- US Department of Health and Human Services Food and Drug Administration (FDA). Guidance for Submission of Immunohistochemistry Applications to the Food and Drug Administration—Final Guidance for Industry. 1998. Available online: https://www.fda.gov/media/73622/download (accessed on 2 October 2020).
- Su, Q.; Schröder, C.H.; Hofmann, W.J.; Otto, G.; Pichlmayr, R.; Bannasch, P. Expression of hepatitis B virus X protein in HBV-infected human livers and hepatocellular carcinomas. Hepatology 1998, 27, 1109–1120. [Google Scholar] [CrossRef] [PubMed]
- Luchman, H.A.; Stechishin, O.D.; Dang, N.H.; Blough, M.D.; Chesnelong, C.; Kelly, J.J.; Nguyen, S.A.; Chan, J.A.; Weljie, A.M.; Cairncross, J.G.; et al. An in vivo patient-derived model of endogenous IDH1-mutant glioma. Neuro Oncol. 2012, 14, 184–191. [Google Scholar] [CrossRef] [PubMed]
- Luchman, H.A.; Chesnelong, C.; Cairncross, J.G.; Weiss, S. Spontaneous loss of heterozygosity leading to homozygous R132H in a patient-derived IDH1 mutant cell line. Neuro Oncol. 2013, 15, 979–980. [Google Scholar] [CrossRef]
- Shechter, I.; Dai, P.; Huo, L.; Guan, G. IDH1 gene transcription is sterol regulated and activated by SREBP-1a and SREBP-2 in human hepatoma HepG2 cells: Evidence that IDH1 may regulate lipogenesis in hepatic cells. J. Lipid Res. 2003, 44, 2169–2180. [Google Scholar] [CrossRef]
- Lu, J.; Xu, L.; Zou, Y.; Yang, R.X.; Fan, Y.; Zhang, W.; Yu, D.; Yao, Y.G. IDH1 p.R132 mutations may not be actively involved in the carcinogenesis of hepatocellular carcinoma. Med. Sci. Monit. 2014, 20, 247–254. [Google Scholar] [CrossRef]
- Balassanian, R.; Wool, G.D.; Ono, J.C.; Olejnik-Nave, J.; Mah, M.M.; Sweeney, B.J.; Liberman, H.; Ljung, B.M.; Pitman, M.B. A superior method for cell block preparation for fine-needle aspiration biopsies. Cancer Cytopathol. 2016, 124, 508–518. [Google Scholar] [CrossRef]
- Saqi, A. The State of Cell Blocks and Ancillary Testing: Past, Present, and Future. Arch. Pathol. Lab. Med. 2016, 140, 1318–1322. [Google Scholar] [CrossRef]
- Mann, H.B.; Whitney, D.R. On a test of whether one of two random variables is stochastically larger than the other. Ann. Math. Statist. 1947, 18, 50–60. [Google Scholar] [CrossRef]
- Srinivasan, M.; Sedmak, D.; Jewell, S. Effect of Fixatives and Tissue Processing on the Content and Integrity of Nucleic Acids. Am. J. Pathol. 2002, 161, 1961–1971. [Google Scholar] [CrossRef]
- Do, H.; Dobrovic, A. Sequence Artifacts in DNA from Formalin-Fixed Tissues: Causes and Strategies for Minimization. Clin. Chem. 2015, 61, 64–71. [Google Scholar] [CrossRef]
- Zhang, P.; Lehmann, B.D.; Shyr, Y.; Guo, Y. The Utilization of Formalin Fixed-Paraffin-Embedded Specimens in High Throughput Genomic Studies. Int. J. Genom. 2017, 2017, 1926304. [Google Scholar] [CrossRef]
- Camelo-Piragua, S.; Jansen, M.; Ganguly, A.; Kim, J.C.; Louis, D.N.; Nutt, C.L. Mutant IDH1-specific immunohistochemistry distinguishes diffuse astrocytoma from astrocytosis. Acta Neuropathol. 2010, 119, 509–511. [Google Scholar] [CrossRef]
- Rao, S.; Rajkumar, A.; Sundaram, S.; Joyce, M.E.; Duvuru, P. Yet another utility for isocitrate dehydrogenase.1: Can it serve as an immunomarker to assess tumor margins in gliomas? J. Cancer Res. Ther. 2020, 16, 1476–1481. [Google Scholar] [CrossRef]
- Søndergaard, C.B.; Scheie, D.; Sehested, A.M.; Skjøth-Rasmussen, J. Finding of IDH1 R132H mutation in histologically non-neoplastic glial tissue changes surgical strategies, a case report. Childs Nerv. Syst. 2017, 33, 1217–1220. [Google Scholar] [CrossRef]
- Geramizadeh, B.; Kohandel-Shirazi, M.; Soltani, A. A simple panel of IDH1 and P53 in differential diagnosis between low-grade astrocytoma and reactive gliosis. Clin. Pathol. 2021, 14, 2632010X20986168. [Google Scholar] [CrossRef] [PubMed]
- Takano, S.; Kato, Y.; Yamamoto, T.; Kaneko, M.K.; Ishikawa, E.; Tsujimoto, Y.; Matsuda, M.; Nakai, K.; Yanagiya, R.; Morita, S.; et al. Immunohistochemical detection of IDH1 mutation, p53, and internexin as prognostic factors of glial tumors. J. Neurooncol. 2012, 108, 361–373. [Google Scholar] [CrossRef]
- Takano, S.; Kato, Y.; Yamamoto, T.; Liu, X.; Ishikawa, E.; Kaneko, M.K.; Ogasawara, S.; Matsuda, M.; Noguchi, M.; Matsumura, A. Diagnostic advantage of double immunohistochemistry using two mutation-specific anti-IDH antibodies (HMab-1 and MsMab-1) in gliomas. Brain Tumor Pathol. 2015, 32, 169–175. [Google Scholar] [CrossRef]
- Fujii, Y.; Ogasawara, S.; Oki, H.; Liu, X.; Kaneko, M.K.; Takano, S.; Kato, Y. A high-sensitive HMab-2 specifically detects IDH1-R132H, the most common IDH mutation in gliomas. Biochem. Biophys. Res. Commun. 2015, 466, 733–739. [Google Scholar] [CrossRef] [PubMed]
- Jabbar, K.J.; Luthra, R.; Patel, K.P.; Singh, R.R.; Goswami, R.; Aldape, K.D.; Medeiros, L.J.; Routbort, M.J. Comparison of next-generation sequencing mutation profiling with BRAF and IDH1 mutation-specific immunohistochemistry. Am. J. Surg. Pathol. 2015, 39, 454–461. [Google Scholar] [CrossRef]
- Catteau, A.; Girardi, H.; Monville, F.; Poggionovo, C.; Carpentier, S.; Frayssinet, V.; Voss, J.; Jenkins, R.; Boisselier, B.; Mokhtari, K.; et al. A new sensitive PCR assay for one-step detection of 12 IDH1/2 mutations in glioma. Acta Neuropathol. Commun. 2014, 2, 58. [Google Scholar] [CrossRef] [PubMed]
- Rossi, S.; Laurino, L.; Furlanetto, A.; Chinellato, S.; Orvieto, E.; Canal, F.; Facchetti, F.; Dei Tos, A.P. Rabbit monoclonal antibodies: A comparative study between a novel category of immunoreagents and the corresponding mouse monoclonal antibodies. Am. J. Clin. Pathol. 2005, 124, 295–302. [Google Scholar] [CrossRef] [PubMed]










{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Synthetic Peptides | Amino Acid Sequences |
|---|---|
| IDH1 wt | KPIIIGRHAYGD |
| IDH1 R132H | KPIIIGHHAYGD |
| IDH1 R132C | KPIIIGCHAYGD |
| IDH1 R132L | KPIIIGLHAYGD |
| IDH1 R132S | KPIIIGSHAYGD |
| IDH1 R132G | KPIIIGGHAYGD |
| IDH2 wt | KPITIGRHAHGD |
| IDH2 R172K | KPITIGKHAHGD |
| IDH2 R172M | KPITIGMHAHGD |
| IDH2 R172W | KPITIGWHAHGD |
| Tumor or Tissue Types (WHO Grades) | Number of Cases Tested | Number of Cases Positive (%) |
|---|---|---|
| Pilocytic astrocytoma (I) | 2 | 0 (0) |
| Diffuse astrocytoma (II) | 19 | 13 (68) |
| Anaplastic astrocytoma (III) | 3 | 3 (100) |
| Oligodendroglioma (II) | 12 | 7 (58) |
| Anaplastic oligodendroglioma (III) | 3 | 2 (67) |
| Primary glioblastoma (IV) | 24 | 0 (0) |
| Secondary glioblastoma (IV) a | 3 | 3 (100) |
| Meningioma (I, II) | 20 | 0 (0) |
| Nonneoplastic brain tissue | 38 | 0 (0) |
| Tissue Types | No. of Samples Tested | No. of Positive Samples |
|---|---|---|
| Cerebrum | 11 | 0 |
| Cerebellum | 4 | 0 |
| Adrenal gland | 3 | 0 |
| Ovary | 3 | 0 |
| Pancreas | 3 | 0 |
| Parathyroid | 3 | 0 |
| Pituitary gland | 3 | 0 |
| Testis | 4 | 0 |
| Thyroid | 3 | 0 |
| Breast | 3 | 0 |
| Spleen | 3 | 0 |
| Tonsil | 3 | 0 |
| Thymus | 3 | 0 |
| Bone marrow | 12 | 0 |
| Lung | 3 | 0 |
| Heart | 5 | 0 |
| Esophagus | 4 | 0 |
| Stomach | 3 | 0 |
| Small intestine | 3 | 0 |
| Colon | 3 | 0 |
| Liver | 5 | 0 |
| Salivary gland | 3 | 0 |
| Kidney | 3 | 0 a |
| Prostate | 3 | 0 |
| Uterus | 3 | 0 |
| Cervix | 3 | 0 |
| Skeletal muscle | 3 | 0 |
| Skin | 3 | 0 |
| Peripheral nerve | 3 | 0 |
| Mesothelial lining | 3 | 0 |
| Bladder | 3 | 0 |
| Tumor/Tissue Types (WHO grades) | No. of Cases Tested | No. Positive for MRQ-67 (%) | No. Positive for H09 (%) | No. with Background ≥ 1 for MRQ-67 (%) | No. with Background ≥ 1 for H09 (%) |
|---|---|---|---|---|---|
| Pilocytic astrocytoma (I) | 1 | 0 | 0 | 0 | 1 |
| Diffuse astrocytoma (II/III) | 14 a | 10 (71) | 10 (71) | 0 (0) | 6 (43) |
| Oligodendroglioma (II/III) | 9 b | 7 (78) | 7 (78) | 0 (0) | 1 (11) |
| Glioblastoma, primary (IV) | 10 | 0 (0) | 0 (0) | 0 (0) | 1 (10) |
| Glioblastoma, secondary (IV) | 3 | 3 (100) | 3 (100) | 0 (0) | 0 (0) |
| Meningioma (I/II) | 20 c | 0 (0) | 0 (0) | 0 (0) | 12 (60) |
| Non-neoplastic brain tissue | 38 | 0 (0) | 0 (0) | 0 (0) | 2 (5) |
| Lesion/Tissue Types and Case Codes | Scores with MRQ-67 (2.54 µg/mL) a | Scores with H09 (3.25 µg/mL) a | Scores with H09 (1.63 µg/mL) a |
|---|---|---|---|
| Pilocytic astrocytoma | |||
| AST-6 | 0/0.5 | 0/1 | 0/0 |
| Diffuse astrocytoma | |||
| AST-2 | 3/0 | 3.5/0 | 3/0 |
| AST-3 | 0/0 | 0/1 | 0/0 |
| AST-4 | 3.5/0 | 3.5/0 | 3/0 |
| AST-8 | 0/0 | 0/2 | 0/0 |
| AST-16 | 3.5/0 | 3.5/0 | 3/0 |
| AST-19 | 3.5/0 | 3.5/0.5 | 1/0 b |
| AST-22 | 4/0 | 4/0.5 | 3.5/0 |
| AST-25 | 3/0 | 3/0 | 2.5/0 |
| Oligodendroglioma | |||
| ODG-1 | 3.5/0 | 3/0 | 3.5/0 |
| ODG-2 | 3.5/0 | 3.5/0.5 | 2.5/0 b |
| ODG-3 | 0/0 | 0/0.5 | 0/0.5 |
| ODG-6 | 3.5/0 | 3/0 | 1/0 b |
| Primary glioblastoma | |||
| GLB-2 | 0/0 | 0/0.5 | 0/0 |
| GLB-3 | 0/0 | 0/0.5 | 0/0 |
| GLB-5 | 0/0 | 0/0 | 0/0 |
| GLB-6 | 0/0 | 0/0.5 | 0/0 |
| GLB-8 | 0/0 | 0/0.5 | 0/0 |
| GLB-10 | 0/0 | 0/0.5 | 0/0 |
| Secondary glioblastoma | |||
| GLB-9 | 4/0 | 4/0.5 | 2.5/0.5 b |
| Meningioma | |||
| MGM-4 | 0/0 | 0/1 | 0/1 |
| MGM-9 | 0/0 | 0/0 | 0/0 |
| MGM-11 | 0/0 | 0/1.5 | 0/0 |
| MGM-13 | 0/0 | 0/1 | 0/0 |
| MGM-14 | 0/0 | 0/0.5 | 0/0 |
| MGM-16 | 0/0.5 | 0/2 | 0/0 |
| MGM-17 | 0/0 | 0/2 | 0/0 |
| MGM-18 | 0/0 | 0/1.5 | 0/0.5 |
| MGM-19 | 0/0 | 0/1 | 0/0 |
| MGM-20 | 0/0 | 0/1 | 0/0 |
| MGM-21 | 0/0 | 0/0.5 | 0/0 |
| Brain tissue | |||
| BRN-20 | 0/0 | 0/0 | 0/0 |
| BRN-21 | 0/0 | 0/0 | 0/0 |
| BRN-25 | 0/0 | 0/0.5 | 0/0 |
| BRN-31 | 0/0 | 0/0 | 0/0 |
| BRN-36 | 0/0 | 0/0.5 | 0/0 |
| BRN-37 | 0/0 | 0/0 | 0/0 |
| BRN-41 | 0/0 | 0/0 | 0/0 |
| BRN-46 | 0/0 | 0/0 | 0/0 |
| BRN-53 | 0/0 | 0/0 | 0/0 |
| Case Codes | Lesion/Tissue Types | IHC Results a | Sequencing Results |
|---|---|---|---|
| AST-18 | Astrocytoma | Positive | R132H |
| AST-19 | Anaplastic astrocytoma | Positive | R132H |
| ODG-1 | Oligodendroglioma | Positive | R132H |
| ODG-5 | Oligodendroglioma | Positive | R132H |
| GLB-9 | Secondary glioblastoma | Positive | R132H |
| ODG-3 | Oligodendroglioma | Negative | Wild-type |
| GLB-8 | Primary glioblastoma | Negative | Wild-type |
| GLB-14 | Primary glioblastoma | Negative | Wild-type |
| MGM-1 | Meningioma | Negative | Wild-type |
| MGM-4 | Meningioma | Negative b,c | Wild-type |
| MGM-17 | Atypical meningioma | Negative b | Wild-type |
| MGM-18 | Meningioma | Negative b | Wild-type |
| MGM-19 | Atypical meningioma | Negative b | Wild-type |
| MGM-20 | Meningioma | Negative b | Wild-type |
| BRN-20 | Cerebellar tissue | Negative | Wild-type |
| BRN-32 | Cerebral tissue | Negative | Wild-type |
| BRN-37 | Cerebral tissue | Negative | Wild-type |
| BRNCEBL-14 | Cerebellar tissue | Negative | Wild-type |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Copaciu, R.; Rashidian, J.; Lloyd, J.; Yahyabeik, A.; McClure, J.; Cummings, K.; Su, Q. Characterization of an IDH1 R132H Rabbit Monoclonal Antibody, MRQ-67, and Its Applications in the Identification of Diffuse Gliomas. Antibodies 2023, 12, 14. https://doi.org/10.3390/antib12010014
Copaciu R, Rashidian J, Lloyd J, Yahyabeik A, McClure J, Cummings K, Su Q. Characterization of an IDH1 R132H Rabbit Monoclonal Antibody, MRQ-67, and Its Applications in the Identification of Diffuse Gliomas. Antibodies. 2023; 12(1):14. https://doi.org/10.3390/antib12010014
Chicago/Turabian StyleCopaciu, Raul, Juliet Rashidian, Joshua Lloyd, Aril Yahyabeik, Jennifer McClure, Kelsea Cummings, and Qin Su. 2023. "Characterization of an IDH1 R132H Rabbit Monoclonal Antibody, MRQ-67, and Its Applications in the Identification of Diffuse Gliomas" Antibodies 12, no. 1: 14. https://doi.org/10.3390/antib12010014
APA StyleCopaciu, R., Rashidian, J., Lloyd, J., Yahyabeik, A., McClure, J., Cummings, K., & Su, Q. (2023). Characterization of an IDH1 R132H Rabbit Monoclonal Antibody, MRQ-67, and Its Applications in the Identification of Diffuse Gliomas. Antibodies, 12(1), 14. https://doi.org/10.3390/antib12010014