
Role of HLA-I Structural Variants and the Polyreactive Antibodies They Generate in Immune Homeostasis

 , ,
, ,
Abstract
1. Introduction
- (i)
- Unique diversity of HLA polymorphism, which is reflected in the differences of the amino acid sequences in different domains, in the basic structural configurations, designated as HLA variants or Faces, and their role in immune homeostasis;
- (ii)
- Comparison of the diversity in allele specificity of HLA Abs in normal healthy individuals and in the commercial preparations of IVIg;
- (iii)
- Diversity of HLA monospecific and polyreactive monoclonal Abs (mAbs) generated by HLA molecules.
- (iv)
- Immune homeostatic role of HLA polyreactive Abs by binding
- (a)
- to intact HLA (Face-1) (e.g., mAb W6/32); and
- (b)
- to the monomer HC of HLA (Face-2) (e.g., TFL-006 & TFL-007); and
- (v)
- Immune homeostatic role of HLA-monospecific Abs.
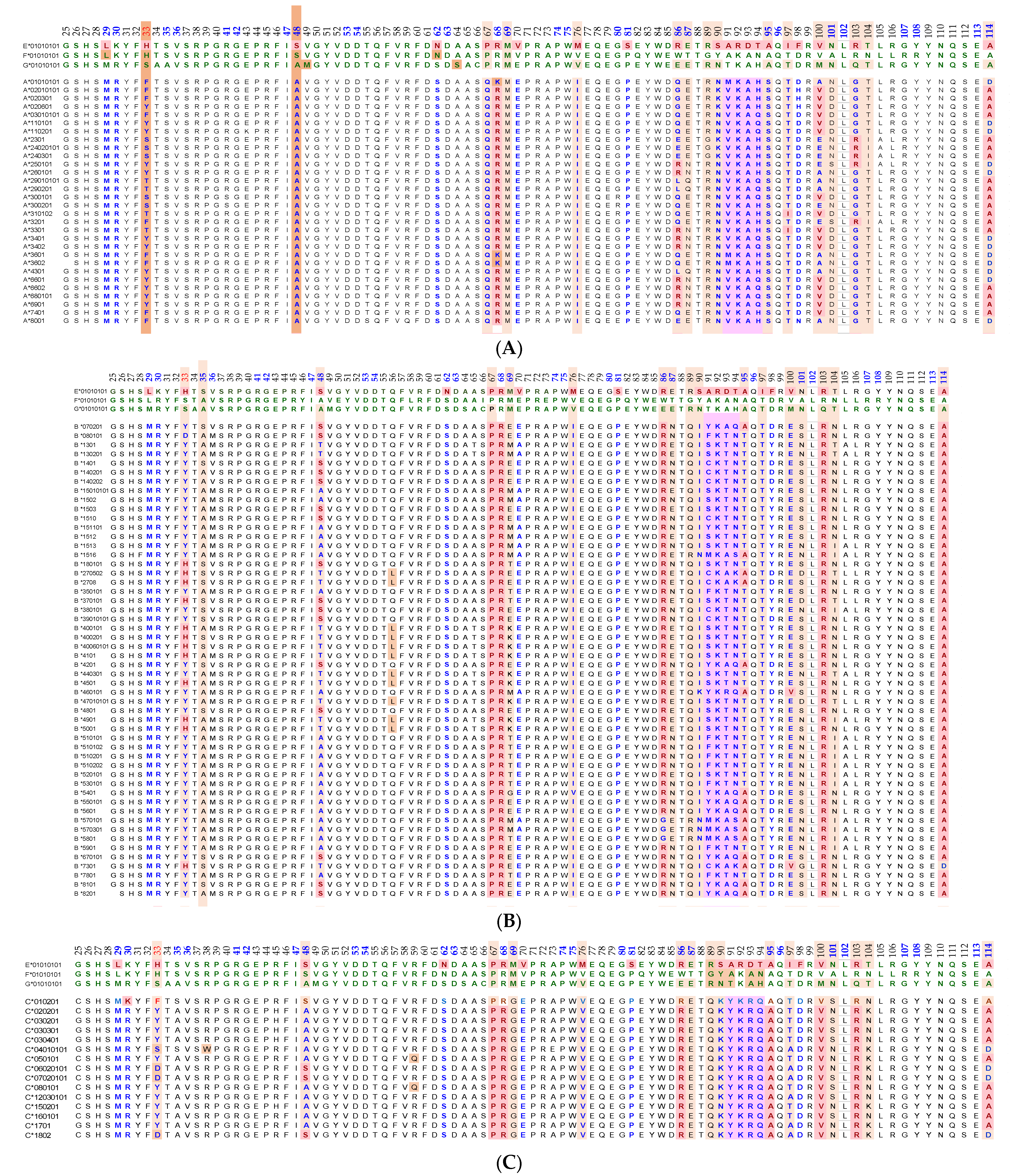
2. The Diversity of HLA Polymorphisms
2.1. Diversity in the HLA Molecules and the Domains of Their Polypeptides
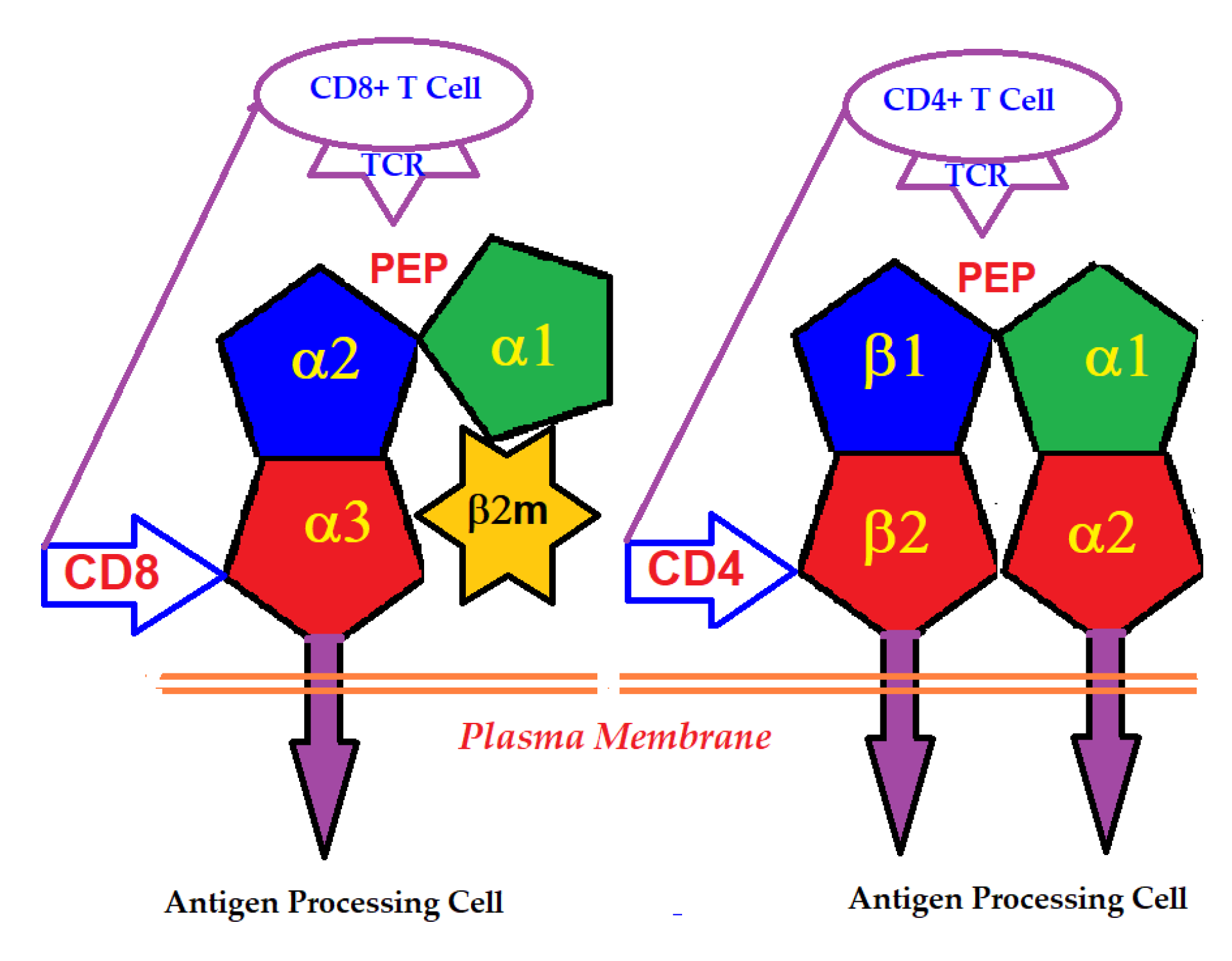
2.2. The Domains Involved in the Primary Function of HLA
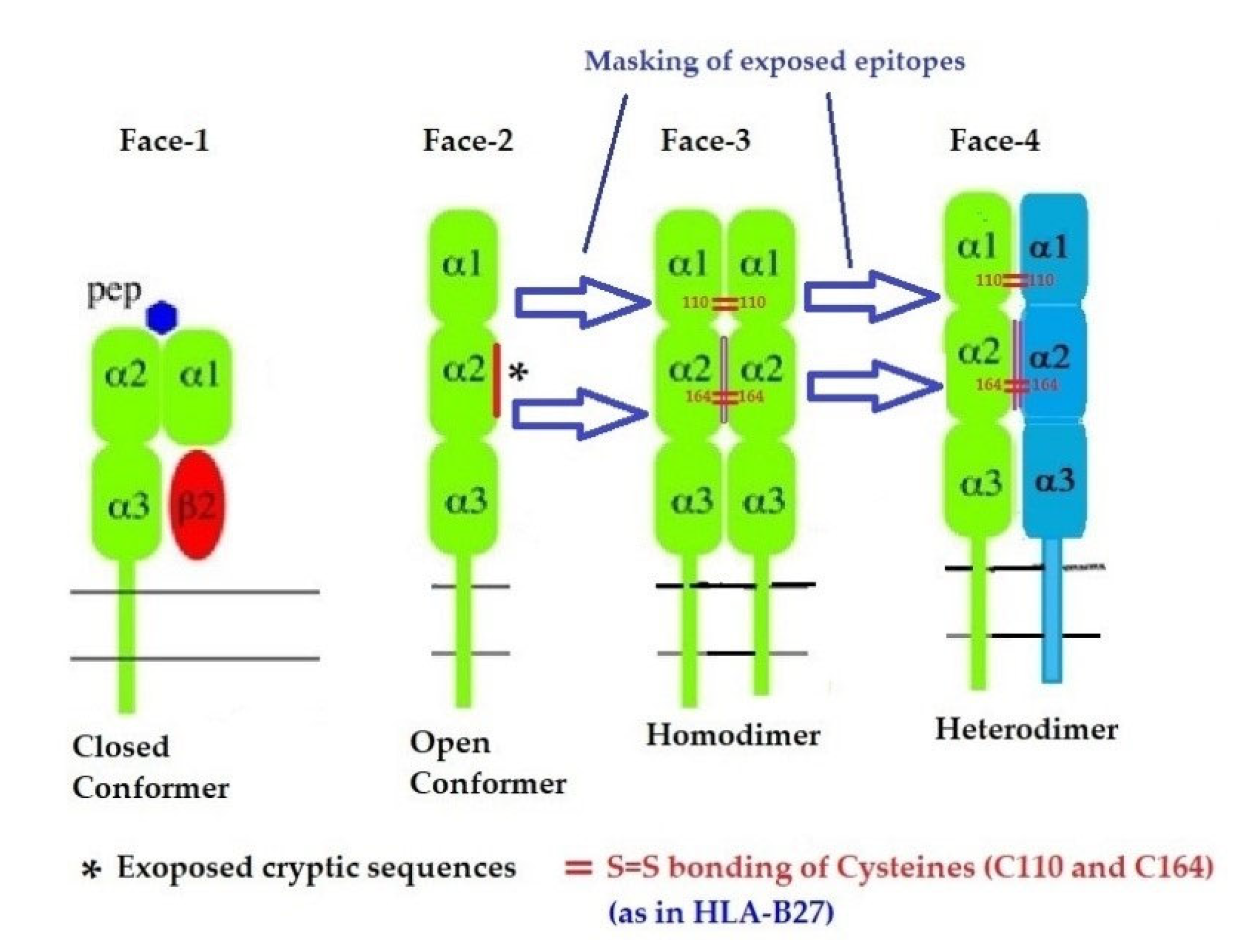
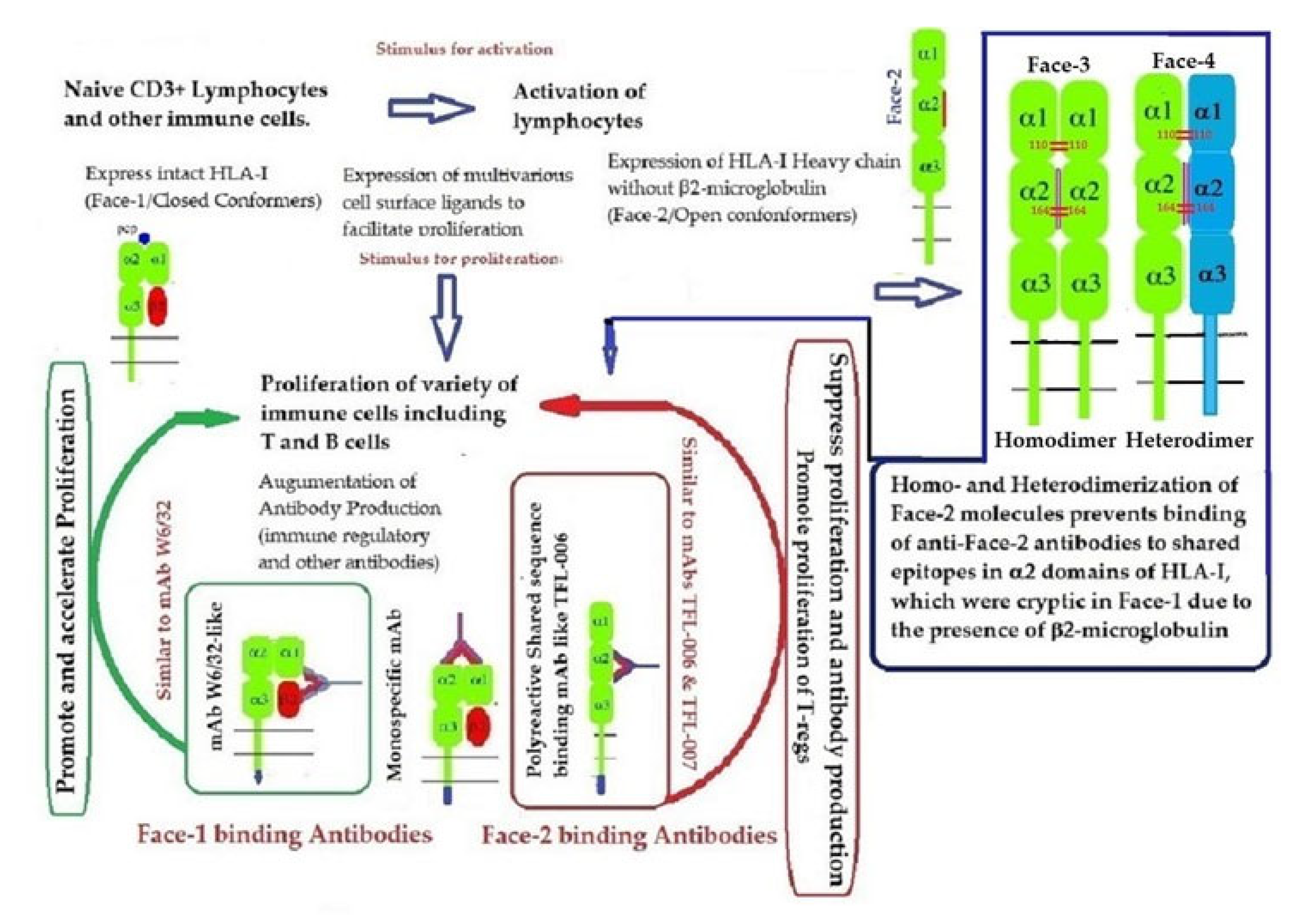
2.3. The Four Structural Variants of HLA-I
- Face-1: The intact HLA-I that consists of an α-HC non-covalently linked to β2m;
- Face-2: HLA-I monomeric α-HC devoid of β2m, exposing immunogenic cryptic sequences, such as 141AYDGKDY147, 152EDLARSWTA159, and 161DTAAQI166.
- Face 3: Homodimers of Face-2: Dimers of two α-HCs of the same allele formed by S-S linkage of cysteines at position 67, as observed in HLA-B27.
- Face-4: Heterodimers of Face-2: Two α-HCs of different HLA-I monomers formed by linkages of cysteines at positions 67, 101, 164, 203, or 43, as in HLA-G, or by salt linkage, H-bonding, or van der Waal forces, as observed in HLA-F and HLA-C.

2.4. Transitory Expression of Face-2 on Activated Immune Cells
2.5. Do the Structural Variants of HLA Have a Role in Immune Homeostasis?
- I.
- Is the dimerization of Face-2 a natural mechanism to prevent the exposure of cryptic epitopes otherwise masked by β2m?
- II.
- If so, what is the purpose of ephemeral exposure of these cryptic epitopes?
- III.
- Does the transient-exposure of cryptic epitopes of Face-2 generate polyreactive Abs following immune recognition by B cells? If so, what kinds of polyreactive HLA Abs are being produced?
- IV.
- Do the naturally occurring polyreactive Abs bind to the ephemeral exposure of cryptic epitopes on Face-2 to elicit salient immunoregulatory functions?
- V.
- How do the immunoregulatory effects differ between the binding of Abs directed against Face-1, such as that of mAb W6/32, and that of Abs directed against Face-2, such as that of TFL-006 and TFL-007?
3. The Diversity of HLA-I Antibodies
3.1. The Diversity of Natural Anti-HLA-I Abs in the Normal Human Sera
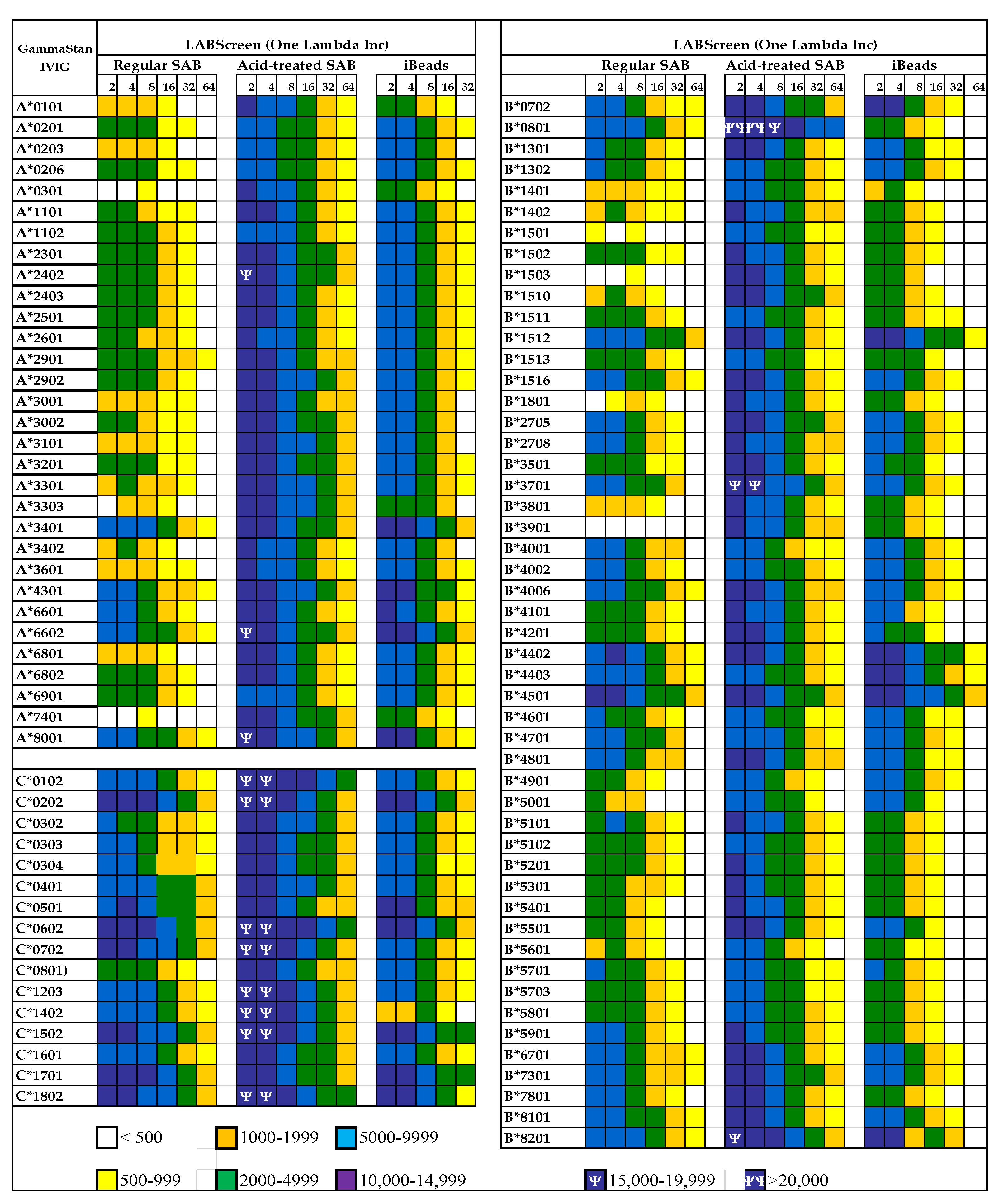
3.2. The Diversity of Abs in Intravenous Immunoglobulin
4. The Pattern of Diversity of HLA Monoclonal Abs (mAbs)
4.1. Is the mAb W6/32 Specific for Closed Conformers (Face-1) of HLA-I?
4.2. Are There True HLA-I Polyreactive mAbs That Recognize Face-2 of All HLA-I Isoforms and Their Alleles?
5. The Immunoregulatory Potential of Anti-HLA-I mAbs unravels the Roles of the HLA-I Face-1 and Face-2 and Their Polyreactive Abs in Immune Homeostasis
5.1. The Immunopotential of Anti-HLA Face-1 mAb W6/32
- (i)
- (ii)
- Tran et al. [103] reported that “although it (W6/32) does recognize in a monomorphic fashion all HLA-A, B, and C molecules when present in their native state (Face-1) on the cell-surface, under partially denaturalized conditions, it recognizes an epitope preserved in free nonreduced α chains of most HLA-B antigens but not in other HLA class-I α chains. Essentially identical results were also obtained with another pan-HLA class-I mAb MEM-147 which cross-blocks W6/32” (p. 443, column 1, para 3). Therefore, one should be cautious while interpreting the results stemming from using W6/32 on activated cells, which express Face-2.
5.1.1. Impact of the mAb W6/32 on Endothelial, Smooth Muscle and Epithelial Cells
5.1.2. Impact of the mAb W6/32 on Immune Cells
5.2. The Immune-Reactive Potential of HLA-I Shared-Epitope Polyreactive mAbs (TFL-006/TFL-007) Directed against Anti-HLA Open Conformers (Face-2)
5.2.1. HLA-I Face-2 Polyreactive mAbs (TFL-006/TFL-007) Mimic IVIg HLA-I Reactivity and Regulate Activated Lymphocytes
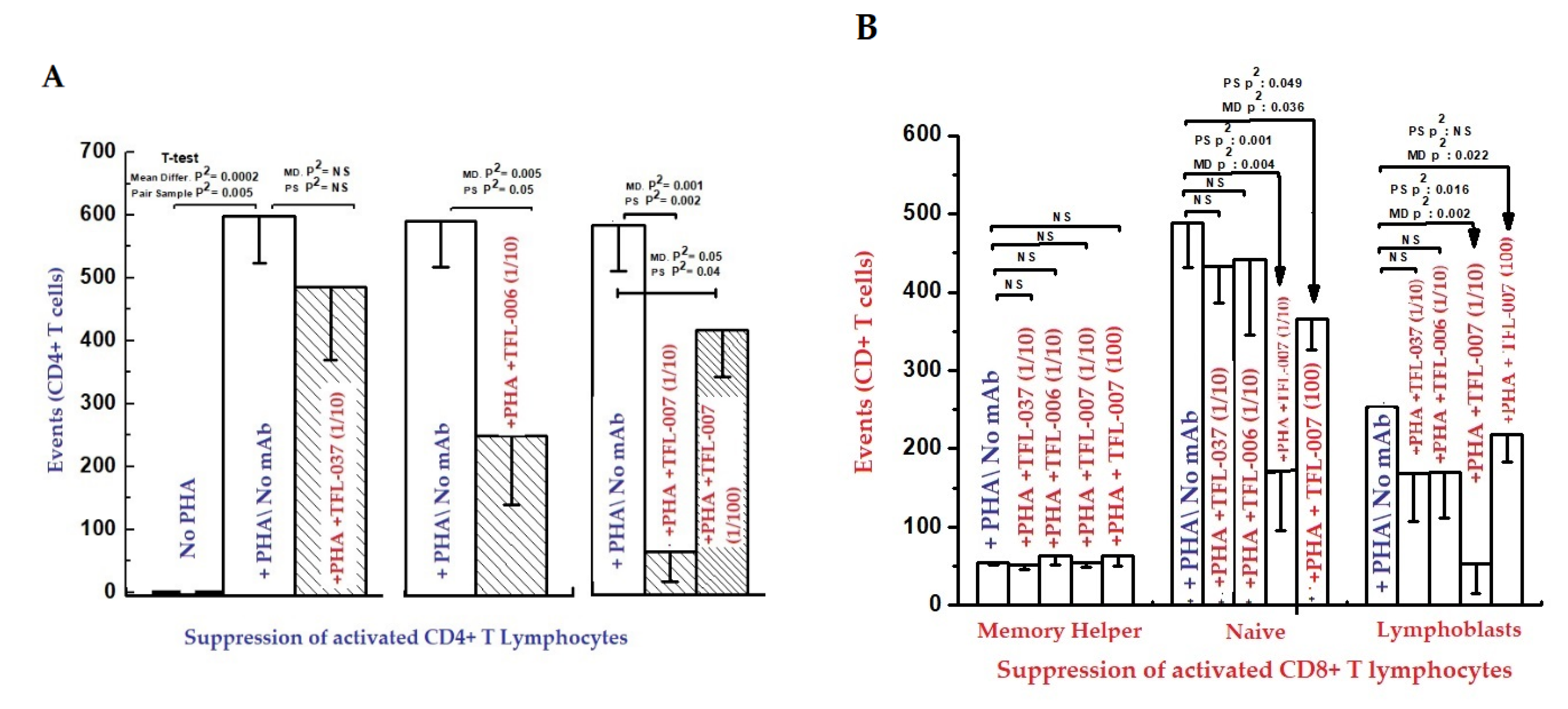
- (i)
- The impact of the mAbs (TFL-006 and TFL-007) (Experimental) and controls (TFL-033 and TFL-037) (Controls) on the blastogenesis in PHA-treated and untreated T-cells was determined by counting lymphoblasts after culturing purified lymphocytes from donors for 72 h. Lymphoblasts were recognized by flow cytometry based on size (side-scatter) and granularity (forward scatter). The blastogenesis and suppression were compared with different commercial preparations of IVIg.
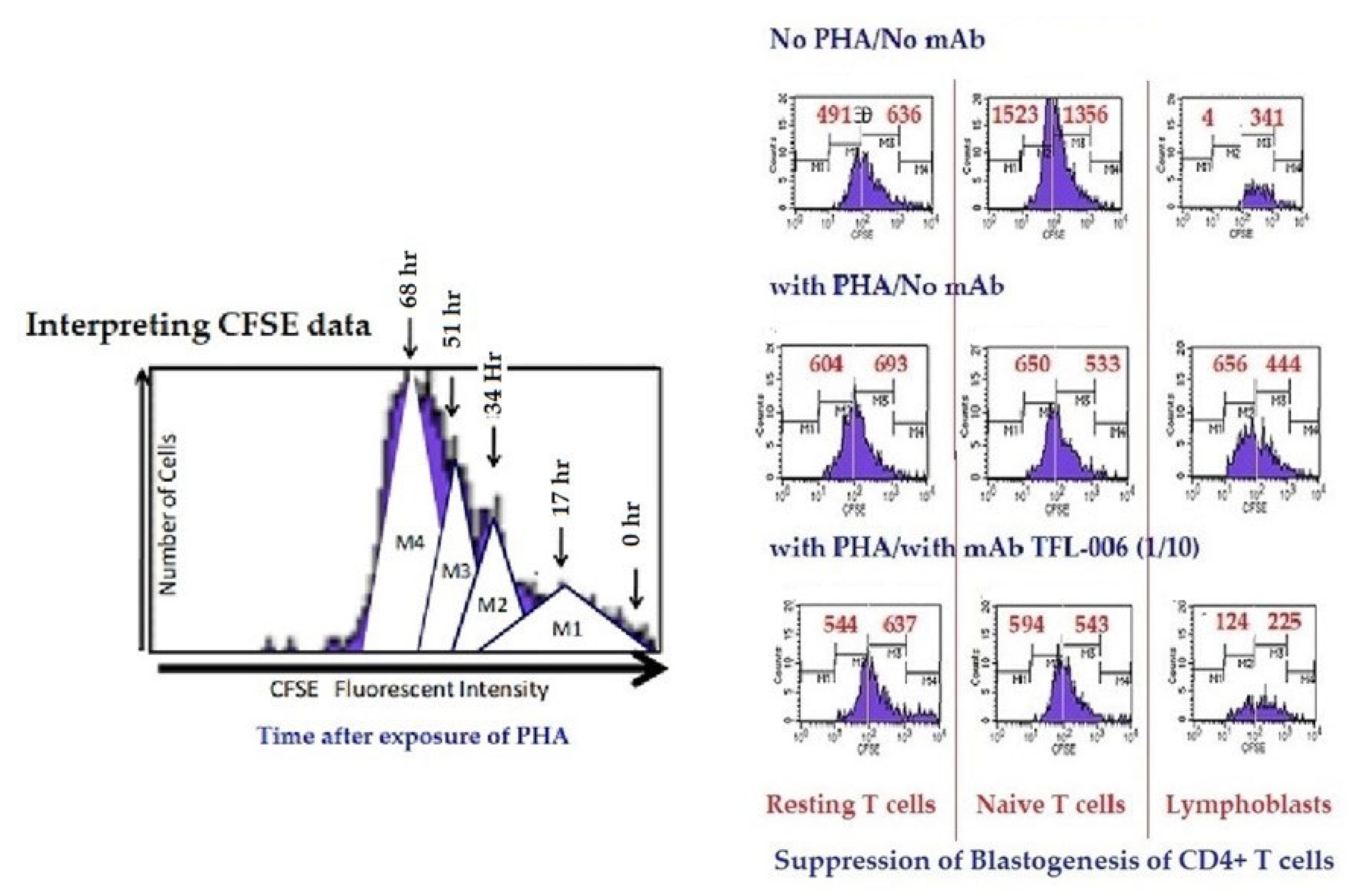
- (ii)
- The impact of the above listed mAbs was examined for enhancement or suppression of proliferation by labelling the purified lymphocytes during PHA activation with the intracellular fluorescent dye carboxyfluorescein succinimidyl ester (CFSE). Using flow cytometry, the mitotic activity was measured by the successive twofold reductions in fluorescent intensity of the T-cells placed in culture for 72 h. PHA-treated T-cells undergo, on average, four to six divisions by 72 h. The suppression of blastogenesis and proliferation can be visualized by cessation of progression of mitotic activity, as measured by the successive twofold reductions in the fluorescent intensity after 72 h of treatment.
5.2.2. HLA-I Face-2-Polyreactive mAbs (TFL-006/TFL-007) May Simulate IVIg in Suppressing allo-HLA Abs Produced by Human B Cells
5.2.3. The Polyreactive mAbs (TFL-006/TFL-007) Mimic IVIg in Upregulating CD4+/CD25+/Foxp3+ Regulatory T-Cells (T-Regs)
6. The Major Players in HLA-I-Mediated Cellular and Humoral Immune Homeostasis
6.1. HLA-I Variants as Receptors in Immune Regulation
6.2. Antibodies Formed against Face-1 and Face-2 as Major Players in HLA-Mediated Immune Homeostasis
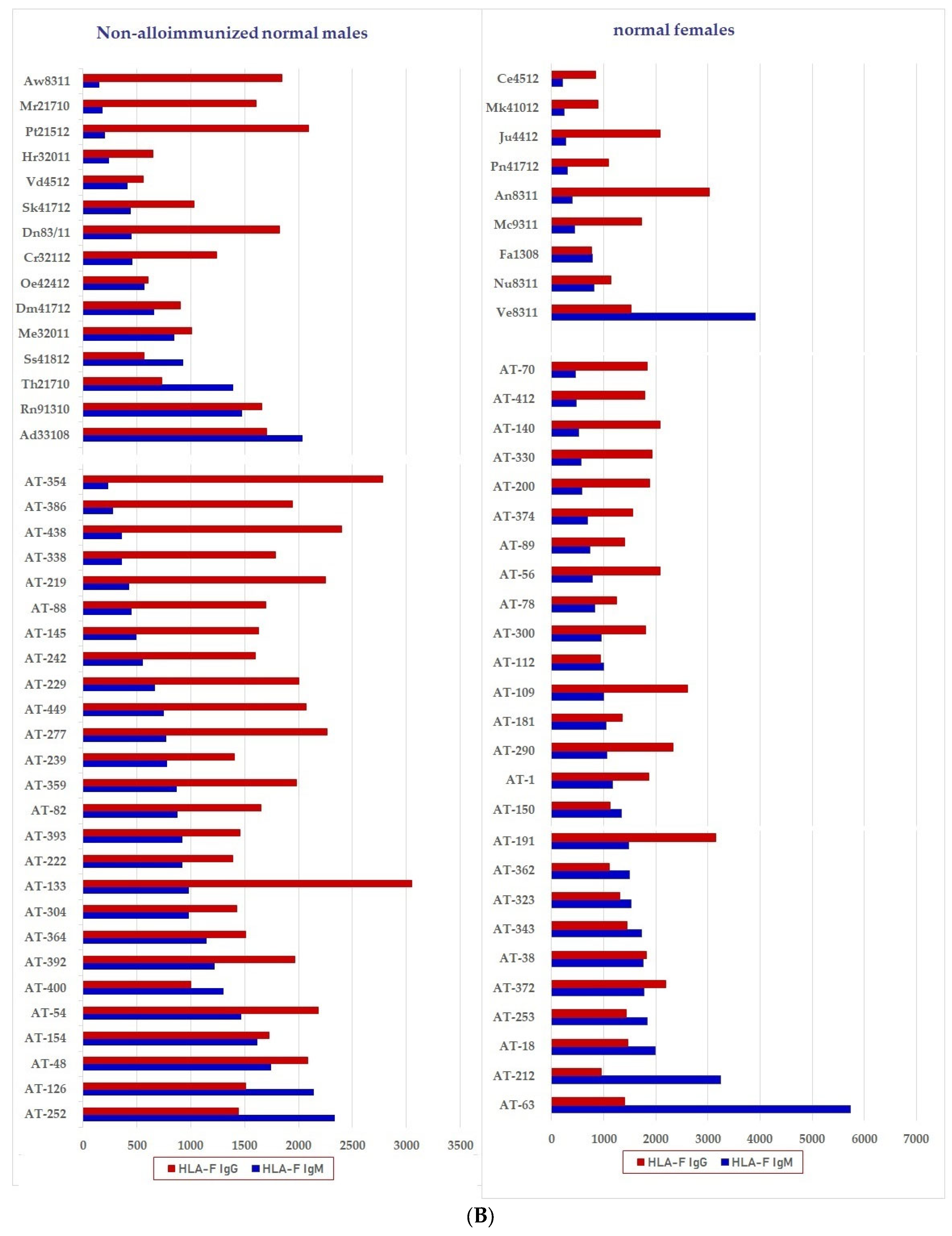
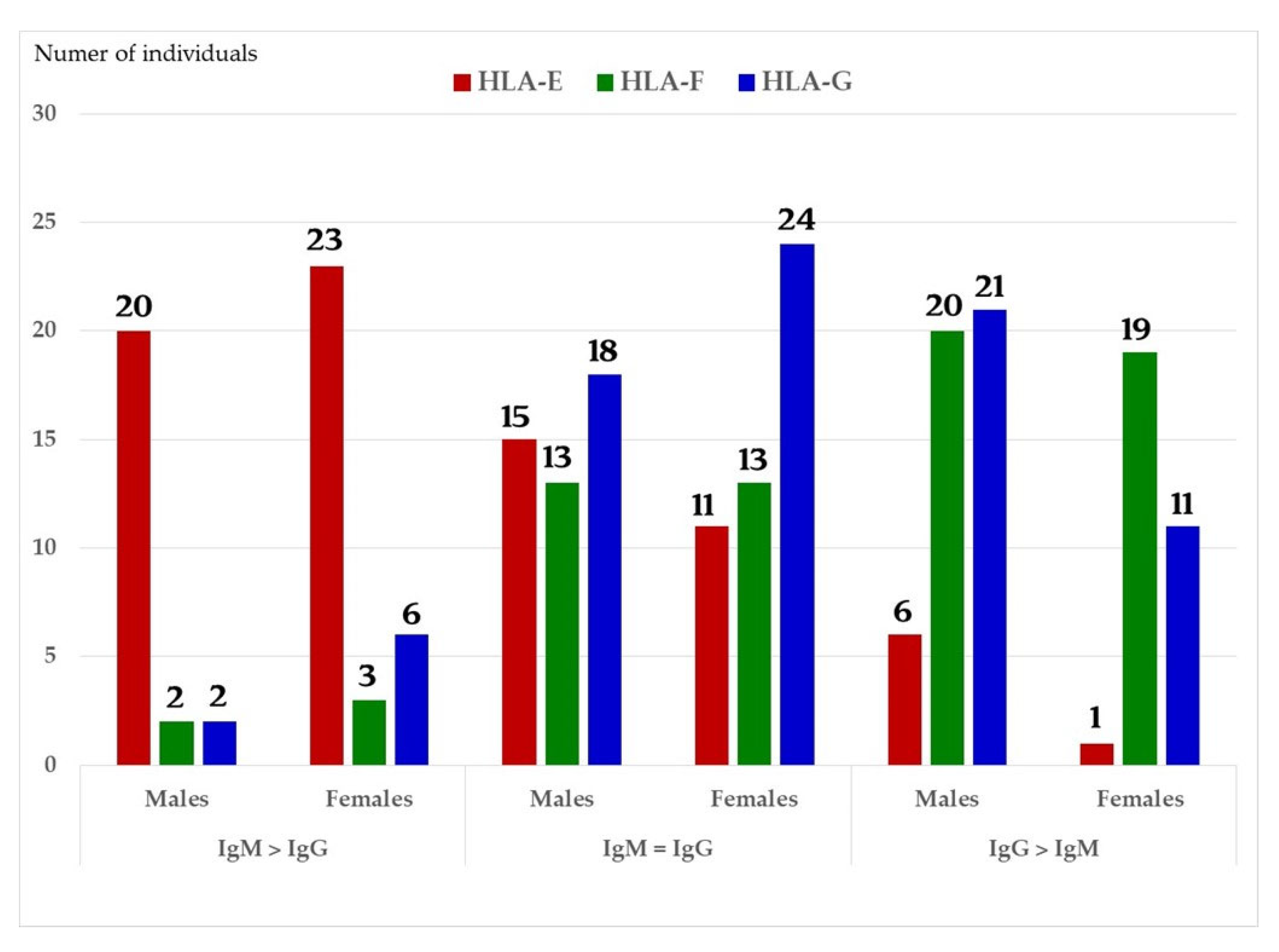
6.2.1. Profiles of HLA Abs in Sera of Normal Females and Non-Alloimmunized Males
6.2.2. Functional Potentials of Antibodies Directed against Face-2 as Evidenced from the Observations Made on IVIg
- (1)
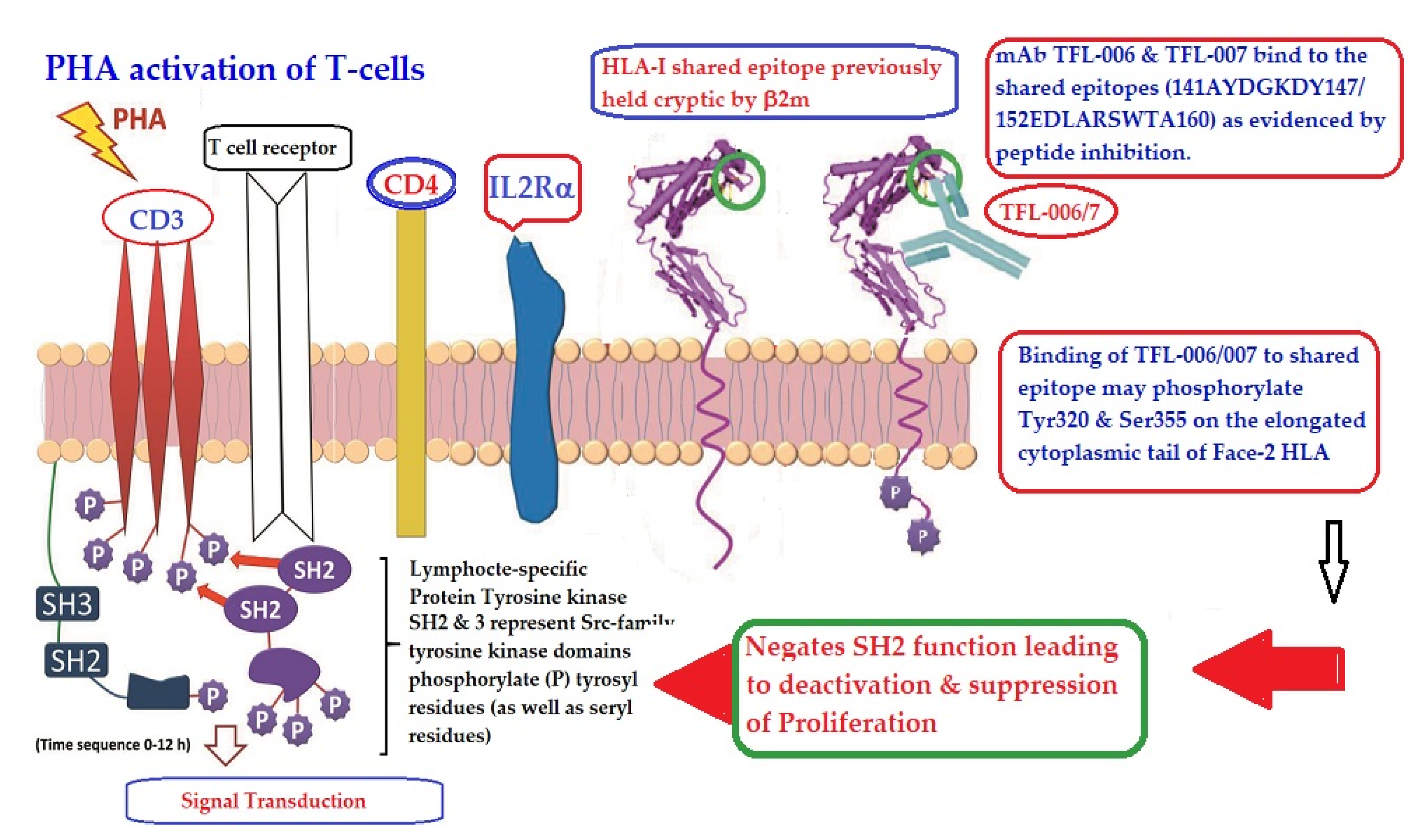
- The difference in HLA variants found in normal and activated CD3+ lymphocytes. In normal healthy individuals, CD3+ lymphocytes express intact HLA-I (Face-1). The lymphocytes may get activated naturally upon inflammation and injury under different pathological conditions. In addition, other exogenous agents can induce activation, such as mAb 9.1 [125,126,127,128], phorbol myristate acetate (PMA), anti-CD3 antibody, phytohemagglutinin (PHA), brefeldin A (PFA), and IFN-γ [15]. Such activated T-cells initiate activation of transcription factors [64], production of cell-surface molecules such as IL-2Rα (Figure 13) [45,46,47], and Face-2 of HLA-I [12,15,16,17,115,116,117,118,119,120,121].
- (2)
- This unique shift in the cell-surface expression of Face-1 HLA-I on normal cells to Face-2 upon activation exposes epitopes that remain cryptic in Face-1 due to masking of amino acid sequences in the α2 domain by β2m.
- (3)
- This domain (Figure 2, Figure 3, Figure 4D–F Figure 5) exposes several sequences shared by HLA-A, -B. -C, -E, -F and -G. (Table 3). When all HLA alleles of all HLA isomers are seen on one screen, it is easy to pinpoint which sequences were cryptic and how they are shared by almost all alleles of all HLA isomers. The uniquely shared epitopes are important to understand and appreciate the role of HLA variants in immune homeostasis.
- (4)
- The exposure of cryptic epitopes may involve immediate immune recognition resulting in the formation of Abs against those shared and cryptic epitopes. This is exemplified by TFL-006 and TFL-007 and by their role in several immunoregulatory functions. Essentially, they are capable of suppressing proliferation of T and B-cells while promoting proliferation of T-regulatory cells (Figure 14).
- (5)
- Figure 13 clarifies the events that occur after the binding by mAbs TFL-006 and TFL-007 to Face-2 on the cell-surface of activated T-cells. The mAb binding to the exposed cryptic epitopes on Face-2 may involve a reversal of activation of T-cells mediated by signal transduction. In support of such an inference, it is documented that the elongation of the cytoplasmic tail of the HLA class-I open conformers will expose otherwise cryptic tyrosine320 [137,138,139] and serine335 [140] with a provision for phosphorylation (Figure 13). Although serine335 is generally considered the primary site of phosphorylation in this tail, phosphorylation of tyrosine320 has been documented by others [141,143]. Either way, what is most important is that the open conformers in activated normal human T-cells are associated with tyrosine phosphorylation and can enable cis interactions with cell-surface receptors or other signalling molecules [141,142,143,144]. The phosphorylation mediated by TFL mAbs may result in dephosphorylation of the cytoplasmic tails of CD3 molecules by activating phosphatases, leading to the arrest of transcription factors and synthesis of the proteins involved in blastogenesis and mitosis.
- (6)
- Again, a homeostatic function of the HLA variant Face-2 is the transient exposure of unique shared epitopes which is terminated by homo- and/or heterodimerization. Face-3, the homodimeric variant of Face-2, and Face-4, the heterodimeric variant of Face-2, recapitulate the function previously performed by β2-m in Face-1 HLA-I (Figure 14). The exposure of cryptic domains is blocked from receptor-ligand interactions and recognition by immunoregulatory Abs such as TFL-006 and TFL-007.
6.2.3. Are Immunoregulatory Roles Restricted Only to HLA-polyreactive mAbs?
7. Conclusions
- (1)
- Exposures of cryptic epitopes in β2-m free HC (Face-2) of HLA upon activation of cells. Such activation may occur during inflammation, injury, malignancy, and other pathological conditions, such as arthritis, autoimmune diseases and cancer.
- (2)
- Several of these exposed cryptic epitopes are shared by almost all HLA-I isomers.
- (3)
- Such exposure invites immune recognition by Abs and various other ligands.
- (4)
- It appears that such exposure may not last long, and it is prevented by homo- or heterodimerization of Face-2.
- (5)
- The Abs generated by the exposed cryptic epitopes are uniquely polyreactive with Face-2 of all HLA isomers since the epitopes are shared by all HLA isomers.
- (6)
- Binding of these Abs is capable of suppressing the proliferation and blastogenesis of CD4+ and CD8+ T-cells.
- (7)
- Binding of these Abs enhances the proliferation of T-regulatory cells, which are capable of several immunoregulatory functions.
- (8)
- The function of these Abs directed against cryptic epitopes on Face-2 is different from that of mAb W6/32 which is capable of binding to Face-1 of all loci (and Face-2 of B alleles) and promoting the proliferation of immune and non-immune cells.
Author Contributions
Funding
Data Availability Statement
Acknowlegments
Conflicts of Interest
References
- Marsh, S.G.E.; Parham, P.; Barber, L.D. The HLA, Facts Book; Facts Book Series; Academic Press: San Diego, CA, USA, 2000; ISBN 0-12-545025-7. [Google Scholar]
- Butler, E.A.; Flynn, F.V.; Harris, H.; Robson, E.B.A. A study of urine proteins by two-dimensional electrophoresis with special reference to the proteinuria of renal tubular disorders. Clin. Chim. Acta 1962, 7, 34–41. [Google Scholar] [CrossRef]
- Greeth, J.M.; Kekwick, R.A.; Flynn, F.V.; Harris, H.; Robson, E.B. An ultracentrifuge study of urine proteins with particular reference to the proteinuria of renal tubular disorders. Clin. Chim. Acta 1963, 8, 406414. [Google Scholar] [CrossRef]
- Berggård, I.; Bearn, A.G. Isolation and properties of a low molecular weight beta-2-globulin occurring in human biological fluids. J. Biol. Chem. 1968, 243, 4095–4103. [Google Scholar] [CrossRef]
- Cresswell, P.; Turner, M.J.; Strominger, J.L. 1973. Papain solubilized HLA antigens from cultured human lymphocytes contain two peptide fragments. Proc. Natl. Acad. Sci. USA 1973, 70, 1603–1607. [Google Scholar] [CrossRef]
- Tanigaki, N.; Nakamuro, K.; Appella, E.; Poulik, M.D.; Pressman, D. Identity of the HL-A common portion fragment and human beta2-microglobulin. Biochem. Biophys. Res. Commun. 1973, 55, 1234–1239. [Google Scholar] [CrossRef]
- Chou, P.Y.; Fasman, G.D. Prediction of the secondary structure of proteins from their amino acid sequence. Adv. Enzymol. Relat. Areas Mol. Biol. 1978, 47, 45–148. [Google Scholar] [CrossRef]
- Kolaskar, A.S.; Tongaonkar, P.C. A semi-empirical method for prediction of antigenic determinants on protein antigens. FEBS Lett. 1990, 276, 172–174. [Google Scholar] [CrossRef]
- Karplus, P.A.; Schulz, G.E. Prediction of chain flexibility in proteins—A tool for the selection of peptide antigens. Naturwissenschaft 1985, 72, 212–213. [Google Scholar] [CrossRef]
- Parker, J.M.; Guo, D.; Hodges, R.S. New hydrophilicity scale derived from high-performance liquid chromatography peptide retention data: Correlation of predicted surface residues with antigenicity and X-ray-derived accessible sites. Biochemistry 1986, 25, 5425–5432. [Google Scholar] [CrossRef]
- Ravindranath, M.H.; Ravindranath, N.M.; Selvan, S.R.; Filippone, E.J.; Amato-Menker, C.J.; El Hilali, F. Four Faces of Cell-Surface HLA Class-I: Their Antigenic and Immunogenic Divergence Generating Novel Targets for Vaccines. Vaccines 2022, 10, 339. [Google Scholar] [CrossRef]
- Schnabl, E.; Stockinger, H.; Majdic, O.; Gaugitsch, H.; Lindley, I.J.; Maurer, D.; Hajek-Rosenmayr, A.; Knapp, W. Activated human T lymphocytes express MHC class I heavy chains not associated with beta 2-microglobulin. J. Exp. Med. 1990, 171, 1431–1442. [Google Scholar] [CrossRef]
- Lancet, D.; Parham, P.; Strominger, J.L. Heavy chain ofHLA-A and HLA-B antigens is conformationally labile: A possible role for beta2-microglobulin. Proc. Natl. Acad. Sci. USA 1979, 76, 3844–3848. [Google Scholar] [CrossRef]
- Raine, T.; Brown, D.; Bowness, P.; Hill Gaston, J.S.; Moffett, A.; Trowsdale, J.; Allen, R.L. Consistent patterns of expression of HLA class I free heavy chains in healthy individuals and raised expression in spondyloarthropathy patients point to physiological and pathological roles. Rheumatology 2006, 45, 1338–1344. [Google Scholar] [CrossRef]
- Demaria, S.; Schwab, R.; Bushkin, Y. The origin and fate of beta 2m-free MHC class I molecules induced on activated T-cells. Cell Immunol. 1992, 142, 103–113. [Google Scholar] [CrossRef]
- Aiuti, A.; Forte, P.; Simeoni, L.; Lino, M.; Pozzi, L.; Fattorossi, A.; Giacomini, P.; Ginelli, E.; Beretta, A.; Siccardi, A. Membrane expression of HLA-Cw4 free chains in activated T-cells of transgenic mice. Immunogenetics 1995, 42, 368–375. [Google Scholar] [CrossRef]
- Arosa, F.A.; Santos, S.G.; Powis, S.J. Open conformers: The hidden Face of MHC-I molecules. Trends Immunol. 2007, 28, 115–123. [Google Scholar] [CrossRef]
- Dangoria, N.S.; De Lay, M.L.; Kingsbury, D.J.; Mear, M.P.; Uchanska-Ziegler, B.; Ziegler, A.; Colbert, R.A. HLA-B27 misfolding is associated with aberrant intermolecular disulfide bond formation (dimerization) in the endoplasmic reticulum. J. Biol. Chem. 2002, 277, 23459–23468. [Google Scholar] [CrossRef]
- Allen, H.J.; Fraser, J.; Flyer, D.; Calvin, S.; Flavell, R. Beta 2-microglobulin is not required for cell-surface expression of the routine class I histocompatibility antigen H-2Db. Proc. Natl. Acad. Sci. USA 1986, 83, 7447–7451. [Google Scholar] [CrossRef]
- Goodridge, J.P.; Burian, A.; Lee, N.; Geraghty D., E. HLA-F complex without peptide binds to MHC class I protein in the open conformer form. J. Immunol. 2010, 184, 6199–6208. [Google Scholar] [CrossRef]
- Algarra, I.; Garcia-Lora, A.; Cabrera, T.; Ruiz-Cabello, F.; Garrido, F. The selection of tumor variants with altered expression of classical and nonclassical MHC class I molecules: Implications for tumor immune escape. Cancer Immunol. Immunother. 2004, 53, 904–910. [Google Scholar] [CrossRef]
- Stam, N.J.; Spits, H.; Ploegh, H.L. Monoclonal antibodies raised against denatured HLA-B locus heavy chains permit biochemical characterization of certain HLA-C locus products. J. Immunol. 1986, 137, 2299–2306. [Google Scholar] [PubMed]
- Grassi, F.; Meneveri, R.; Gullberg, M.; Lopalco, L.; Rossi, G.B.; Lanza, P.; De Santis, C.; Bratsand, G.; Butto, S.; Ginelli, E.; et al. Human immunodeficiency virus type 1 gpl20 mimics a hidden monomorphic epitope borne by class I major histocompatibility complex heavy chains. J. Exp. Med. 1990, 174, 53–62. [Google Scholar] [CrossRef] [PubMed]
- Marozzi, A.; Meneveri, R.; Bunone, G.; De Santis, C.; Lopalco, L.; Beretta, A.; Siccardi, A.G.; Della Valle, G.; Ginelli, E. Expression of beta 2m-free HLA class I heavy chains in neuroblastoma cell lines. Scand. J. Immunol. 1993, 37, 661–667. [Google Scholar] [CrossRef] [PubMed]
- Martayan, A.; Fiscella, M.; Setini, A.; Ciccarelli, G.; Gambari, R.; Feriotto, G.; Beretta, A.; Siccardi, A.G.; Appella, E.; Giacomini, P. Conformation and surface expression of free HLA-CW1 heavy chains in the absence of beta 2-microglobulin. Hum. Immunol. 1997, 53, 23–33. [Google Scholar] [CrossRef]
- Giacomini, P.; Beretta, A.; Nicotra, M.R.; Ciccarelli, G.; Martayan, A.; Cerboni, C.; Lopalco, L.; Bini, D.; Delfino, L.; Ferrara, G.B.; et al. HLA-C heavy chains free of β2-microglobulin: Distribution in normal tissues and neoplastic lesions of non-lymphoid origin and interferon-γ responsiveness. Tissue Antigens 1997, 50, 555–566. [Google Scholar] [CrossRef]
- Avrameas, S.; Guilbert, B.; Dighiero, B. Natural antibodies against tubulin. actin, myoglobin. Ihyroglobtilin. fetuin, albumin and transferin arc present in normal human sera, and monoclonal immunoglobulins from multiple myclomc and Waldenstrom macroglobulinemia may express similar antibody specificities. Ann. Immunol. 1981, 132, 103–113. [Google Scholar] [CrossRef]
- Collins, Z.V.; Arnold, P.F.; Peetoom, F.; Smith, G.S.; Walford, R.L. A naturally occurring monospecific anti-HLA8 isoantibody. Tissue Antigens 1973, 3, 358–363. [Google Scholar] [CrossRef]
- Tongio, M.M.; Falkenrodt, A.; Mitsuishi, Y.; Urlacher, A.; Bergerat, J.P.; North, M.L.; Mayer, S. Natural HLA antibodies. Tissue Antigens 1985, 26, 271–285. [Google Scholar] [CrossRef]
- Zhou, B.; Saito, S.; Nakazawa, Y.; Kobayashi, N.; Matsuda, M.; Matsumoto, Y.; Hosoyama, T.; Koike, K. Existence of an immunoglobulin G component of naturally occurring HLA class I antibodies that are not directed against self-antigens in human serum. Tissue Antigens 2008, 72, 98–104. [Google Scholar] [CrossRef]
- Ravindranath, M.H.; Terasaki, P.I.; Maehara, C.Y.; Jucaud, V.; Kawakita, S.; Pham, T.; Yamashita, W. Immunoglobulin (Ig)G purified from human sera mirrors intravenous Ig human leucocyte antigen (HLA) reactivity and recognizes one’s own HLA types, but may be masked by Fab complementarity-determining region peptide in the native sera. Clin. Exp. Immunol. 2015, 179, 309–328. [Google Scholar] [CrossRef]
- Morales-Buenrostro, L.E.; Terasaki, P.I.; Marino-Vazquez, L.A.; Lee, J.H.; El-Awar, N.; Alberú, J. “Natural” human leukocyte antigen antibodies found in nonalloimmunized healthy males. Transplantation 2008, 86, 1111–1115. [Google Scholar] [CrossRef]
- Ravindranath, M.H.; Jucaud, V.; Ferrone, S. Monitoring native HLA-I trimer specific antibodies in Luminex multiplex single antigen bead assay: Evaluation of beadsets from different manufacturers. J. Immunol. Methods 2017, 450, 73–80. [Google Scholar] [CrossRef]
- Jucaud, V.; Ravindranath, M.H.; Terasaki, P.I. Conformational Variants of the Individual HLA-I Antigens on Luminex Single Antigen Beads Used in Monitoring HLA Antibodies: Problems and Solutions. Transplantation 2017, 101, 764–777. [Google Scholar] [CrossRef]
- Sullivan, K.M.; Kopecky, K.J.; Jocom, J.; Fisher, L.; Buckner, C.D.; Meyers, J.D.; Counts, G.W.; Bowden, R.A.; Peterson, F.B.; Witherspoon, R.P.; et al. Immuno-modulatory and antimicrobial efficacy of intravenous immunoglobulin in bone marrow transplantation. N. Engl. J. Med. 1990, 323, 705–712. [Google Scholar] [CrossRef]
- Sullivan, K.M.; Storek, J.; Kopecky, K.J.; Jocom, J.; Longton, G.; Flowers, M.; Siadak, M.; Nims, J.; Witherspoon, R.P.; Anasetti, C.; et al. A controlled trial of long-term administration of intravenous immunoglobulin to prevent late infection and chronic graft-vs.-host disease after marrow transplantation: Clinical outcome and effect on subsequent immune recovery. Biol. Blood Marrow Transpl. 1996, 2, 44–53. [Google Scholar]
- Bayry, J.; Lacroix-Desmazes, S.; Kazatchkine, M.D.; Kaveri, S.V. Intravenous immunoglobulin for infectious diseases: Back to the pre-antibiotic and passive prophylaxis era? Trends Pharmacol. Sci. 2004, 25, 306–310. [Google Scholar] [CrossRef]
- Molica, S.; Musto, P.; Chiurazzi, F.; Specchia, G.; Brugiatelli, M.; Cicoira, L.; Levato, D.; Nobile, F.; Carotenuto, M.; Liso, V.; et al. Prophylaxis against infections with low-dose intravenous immunoglobulins (IVIg) in chronic lymphocytic leukemia. Results of a crossover study. Haematologica 1996, 81, 121–126. [Google Scholar]
- Elluru, S.R.; Kaveri, S.V.; Bayry, J. The protective role of immunoglobulins in fungal infections and inflammation. Semin. Immunopathol. 2015, 37, 187–197. [Google Scholar] [CrossRef]
- Dalakas, M.C. Intravenous immunoglobulin in autoimmune neuromuscular diseases. JAMA 2004, 291, 2367–2375. [Google Scholar] [CrossRef]
- Sibéril, S.; Elluru, S.; Graff-Dubois, S.; Negi, V.S.; Delignat, S.; Mouthon, L.; Lacroix-Desmazes, S.; Kazatchkine, M.D.; Bayry, J.; Kaveri, S.V. Intravenous immunoglobulins in autoimmune and inflammatory diseases: A mechanistic perspective. Ann. N. Y. Acad. Sci. 2007, 1110, 497–506. [Google Scholar] [CrossRef]
- Galeotti, C.; Kaveri, S.V.; Bayry, J. IVIG-mediated effector functions in autoimmune and inflammatory diseases. Int. Immunol. 2017, 29, 491–498. [Google Scholar] [CrossRef]
- Hoffmann, J.H.O.; Enk, A.H. High-Dose Intravenous Immunoglobulin in Skin Autoimmune Disease. Front. Immunol. 2019, 10, 1090. [Google Scholar] [CrossRef]
- Jordan, S.C.; Toyoda, M.; Vo, A.A. Regulation of immunity and inflammation by intravenous immunoglobulin: Relevance to solid organ transplantation. Expert Rev. Clin. Immunol. 2011, 7, 341–348. [Google Scholar] [CrossRef]
- Abu Jawdeh, B.G.; Cuffy, M.C.; Alloway, R.R.; Shields, A.R.; Woodle, E.S. Desensitization in kidney transplantation: Review and future perspectives. Clin. Transpl. 2014, 28, 494–507. [Google Scholar] [CrossRef] [PubMed]
- Picascia, A.; Grimaldi, V.; Paolillo, R.; Vasco, M.; Casamassimi, A.; De Luca, F.P.; Cavalca, F.; Schiano, C.; Napoli, C. Intravenous human immunoglobulin treatment of serum from HLA-sensitized patients in kidney transplantation. Ren. Fail. 2014, 36, 585–588. [Google Scholar] [CrossRef]
- Kornberg, A. Intravenous immunoglobulins in liver transplant patients: Perspectives of clinical immune modulation. World J. Hepatol. 2015, 7, 1494–1508. [Google Scholar] [CrossRef]
- Tedla, F.M.; Roche-Recinos, A.; Brar, A. Intravenous immunoglobulin in kidney transplantation. Curr. Opin. Organ. Transpl. 2015, 20, 630–637. [Google Scholar] [CrossRef]
- Matignon, M.; Leibler, C.; Moranne, O.; Salomon, L.; Charron, D.; Lang, P.; Jacquelinet, C.; Suberbielle, C.; Grimbert, P. Anti-HLA sensitization after kidney allograft nephrectomy: Changes one year post-surgery and beneficial effect of intravenous immunoglobulin. Clin. Transpl. 2016, 30, 731–740. [Google Scholar] [CrossRef] [PubMed]
- Matignon, M.; Pilon, C.; Commereuc, M.; Grondin, C.; Leibler, C.; Kofman, T.; Audard, V.; Cohen, J.; Canoui-Poitrine, F.; Grimbert, P. Intravenous immunoglobulin therapy in kidney transplant recipients with de novo DSA: Results of an observational study. PLoS ONE. 2017, 12, e0178572. [Google Scholar] [CrossRef]
- Glotz, D.; Rostaing, L.; Merville, P.; Squifflet, J.P.; Lebranchu, Y. Intravenous Immunoglobulins in the Prevention of Rejection of a Second or Third Kidney Graft. Transpl. Proc. 2018, 50, 70–71. [Google Scholar] [CrossRef]
- Bujnowska, A.; Michon, M.; Konopelski, P.; Hryniewiecka, E.; Jalbrzykowska, A.; Perkowska-Ptasinska, A.; Cieciura, T.; Zagozdzon, R.; Paczek, L.; Ciszek, M. Outcomes of Prolonged Treatment with Intravenous Immunoglobulin Infusions for Acute Antibody-mediated Rejection in Kidney Transplant Recipients. Transpl. Proc. 2018, 50, 1720–1725. [Google Scholar] [CrossRef]
- Hoang, J.; Krisl, J.; Moaddab, M.; Nguyen, D.T.; Graviss, E.A.; Hussain, I.; Kassi, M.; Yousefzai, R.; Kim, J.; Trachtenberg, B.; et al. Intravenous immunoglobulin in heart transplant recipients with mild to moderate hypogammaglobulinemia and infection. Clin. Transpl. 2022, 36, e14571. [Google Scholar] [CrossRef]
- Ulisses, L.R.S.; Paixão, J.O.; Agena, F.; Souza, P.S.; Paula, F.J.; Bezerra, G.; Rodrigues, H.; Panajotopolous, N.; David-Neto, E.; Castro, M.C.R. Desensitization using IVIG alone for living-donor kidney transplant: Impact on donor-specific antibodies. J. Bras. Nefrol. 2022, 8. [Google Scholar] [CrossRef]
- Vani, J.; Elluru, S.; Negi, V.S.; Lacroix-Desmazes, S.; Kazatchkine, M.D.; Bayry, J.; Kaveri, S.V. Role of natural antibodies in immune homeostasis: IVIg perspective. Autoimmun. Rev. 2008, 7, 440–444. [Google Scholar] [CrossRef]
- Sbrana, S.; Ruocco, L.; Vanacore, R.; Azzarà, A.; Ambrogi, F. In vitro effects of an immunoglobulin preparation for intravenous use (IVIG) on T-cells activation. Allergy Immunol. 1993, 25, 35–37. [Google Scholar]
- Hurez, V.; Kaveri, S.V.; Mouhoub, A.; Dietrich, G.; Mani, J.C.; Klatzmann, D.; Kazatchkine, M.D. Anti-CD4 activity of normal human immunoglobulin G for therapeutic use (intravenous immunoglobulin, IVIg). Ther. Immunol. 1994, 1, 269–277. [Google Scholar]
- Amran, D.; Renz, H.; Lack, G.; Bradley, K.; Gelfand, E.W. Suppression of cytokine-dependent human T-cell proliferation by intravenous immunoglobulin. Clin. Immunol. Immunopathol. 1994, 73, 180–186. [Google Scholar] [CrossRef]
- Andersson, J.; Skansén-Saphir, U.; Sparrelid, E.; Andersson, U. Intravenous immune globulin affects cytokine production in T lymphocytes and monocytes/macrophages. Clin. Exp. Immunol. 1996, 104 (Suppl. S1), 10–20. [Google Scholar] [CrossRef]
- Aktas, O.; Waiczies, S.; Grieger, U.; Wendling, U.; Zschenderlein, R.; Zipp, F. Polyspecific immunoglobulins (IVIg) suppress proliferation of human (auto) antigen-specific T-cells without inducing apoptosis. J. Neuroimmunol. 2001, 114, 160–167. [Google Scholar] [CrossRef]
- MacMillan, H.F.; Lee, T.; Issekutz, A.C. Intravenous immunoglobulin G-mediated inhibition of T-cell proliferation reflects an endogenous mechanism by which IgG modulates T-cell activation. Clin. Immunol. 2009, 132, 222–233. [Google Scholar] [CrossRef]
- Tha-In, T.; Metselaar, H.J.; Tilanus, H.W.; Boor, P.P.; Mancham, S.; Kuipers, E.J.; de Man, R.A.; Kwekkeboom, J. Superior immunomodulatory effects of intravenous immunoglobulins on human T-cells and dendritic cells: Comparison to calcineurin inhibitors. Transplantation 2006, 81, 1725–1734. [Google Scholar] [CrossRef] [PubMed]
- Van Schaik, I.N.; Lundkvist, I.; Vermeulen, M.; Brand, A. Polyvalent immunoglobulin for intravenous use interferes with cell proliferation in vitro. J. Clin. Immunol. 1992, 12, 325–334. [Google Scholar] [CrossRef] [PubMed]
- Prasad, N.K.; Papoff, G.; Zeuner, A.; Bonnin, E.; Kazatchkine, M.D.; Ruberti, G.; Kaveri, S.V. Therapeutic preparations of normal polyspecific IgG (IVIg) induce apoptosis in human lymphocytes and monocytes: A novel mechanism of action of IVIg involving the Fas apoptotic pathway. J. Immunol. 1998, 161, 3781–3790. [Google Scholar]
- Aubin, E.; Lemieux, R.; Bazin, R. Indirect inhibition of in vivo and in vitro T-cell responses by intravenous immunoglobulins due to impaired antigen presentation. Blood 2010, 115, 1727–1734. [Google Scholar] [CrossRef] [PubMed]
- Aubin, É.; Proulx, D.P.; Trépanier, P.; Lemieux, R.; Bazin, R. Prevention of T cell dependent native antigen presentation. Clin. Immunol. 2011, 141, 273–283. [Google Scholar] [CrossRef]
- Kaveri, S.; Vassilev, T.; Hurez, V.; Lengagne, R.; Lefranc, C.; Cot, S.; Pouletty, P.; Glotz, D.; Kazatchkine, M.D. Antibodies to a conserved region of HLA class I molecules, capable of modulating CD8 T cell-mediated function, are present in pooled normal immunoglobulin for therapeutic use. J. Clin. Investig. 1996, 97, 865–869. [Google Scholar] [CrossRef]
- Ravindranath, M.H.; Terasaki, P.I.; Pham, T.; Jucaud, V.; Kawakita, S. Therapeutic preparations of IVIg contain naturally occurring anti-HLA-E Abs that react with HLA-Ia (HLA-A/-B/-Cw) alleles. Blood 2013, 121, 2013–2028. [Google Scholar] [CrossRef]
- Cavallini, G.; Pozzan, T.; Selvatici, R.; Baricordi, R.; Mazzilli, C.; Gandini, E. Monoclonal antibodies against HLA class I antigens inhibit human lymphocyte proliferation but do not affect mitogen dependent second messenger generation. Biochem. Biophys. Res. Commun. 1988, 154, 712–718. [Google Scholar] [CrossRef]
- De Felice, M.; Turco, M.C.; Costanzo, F.; Corbo, L.; Ferrone, S.; Venuta, S. Inhibition by anti-HLA class I mAb of IL-2 and IL-2 receptor synthesis in lymphocytes stimulated with PHA-P. Cell Immunol. 1990, 126, 420–427. [Google Scholar] [CrossRef]
- Tanabe, M.; Sekimata, M.; Ferrone, S.; Takiguchi, M. Structural and functional analysis of monomorphic determinants recognized by monoclonal antibodies reacting with the HLA class I alpha 3 domain. J. Immunol. 1992, 148, 3202–3209. [Google Scholar]
- Smith, D.M.; Bluestone, J.A.; Jeyarajah, D.R.; Newberg, M.H.; Engelhard, V.H.; Thistlethwaite, J.R., Jr.; Woodle, E.S. Inhibition of T cell activation by a monoclonal antibody reactive against the alpha 3 domain of human MHC class I molecules. J. Immunol. 1994, 153, 1054–1067. [Google Scholar]
- Woodle, E.S.; Smith, D.M.; Zhou, N. Class I MHC mediates programmed cell death in human lymphoid cells. Transplantation 1997, 64, 140–146. [Google Scholar] [CrossRef]
- Woodle, E.S.; Smith, D.M.; Bluestone, J.A.; Kirkman, W.M., 3rd; Green, D.R.; Skowronski, E.W. Anti-human class I MHC antibodies induce apoptosis by a pathway that is distinct from the Fas antigen-mediated pathway. J. Immunol. 1997, 158, 2156–2164. [Google Scholar]
- Coupel, S.; Moreau, A.; Hamidou, M.; Horejsi, V.; Soulillou, J.-P.; Béatrice Charreau, B. Expression and release of soluble HLA-E is an immunoregulatory feature of endothelial cell activation. Blood 2007, 109, 2806–2814. [Google Scholar] [CrossRef]
- Harris, P.E.; Bian, H.; Reed, E.F. Induction of high-affinity fibroblast growth factor receptor expression and proliferation in human endothelial cells by anti-HLA antibodies: A possible mechanism for transplant atherosclerosis. J. Immunol. 1997, 159, 5697–5704. [Google Scholar]
- Bian, H.; Harris, P.E.; Mulder, A.; Reed, E.F. Anti-HLA antibody ligation to HLA class I molecules expressed by endothelial cells stimulates tyrosine phosphorylation, inositol phosphate generation, and proliferation. Hum. Immunol. 1997, 53, 90–97. [Google Scholar] [CrossRef]
- Bian, H.; Harris, P.E.; Reed, E.F. Ligation of HLA class I molecules on smooth muscle cells with anti-HLA antibodies induces tyrosine phosphorylation, fibroblast growth factor receptor expression and cell proliferation. Int. Immunol. 1998, 10, 1315–1323. [Google Scholar] [CrossRef]
- Bian, H.; Reed, E.F. Alloantibody-mediated class I signal transduction in endothelial cells and smooth muscle cells: Enhancement by IFN-γ and TNF-α. J. Immunol. 1999, 163, 1010–1018. [Google Scholar]
- Bian, H.; Reed, E.F. Anti-HLA antibodies transduce proliferative signals in endothelial cells and smooth muscle cells. Transpl. Proc. 1999, 31, 1924. [Google Scholar] [CrossRef]
- Nath, N.; Bian, H.; Reed, E.F.; Chellappan, S.P. HLA class I-mediated induction of cell proliferation involves cyclin E-mediated inactivation of Rb function and induction of E2F activity. J. Immunol. 1999, 162, 5351. [Google Scholar]
- Jin, Y.P.; Singh, R.P.; Du, Z.Y.; Rajasekaran, A.K.; Rozengurt, E.; Reed, E.F. Ligation of HLA class I molecules on endothelial cells induces phosphorylation of Src, paxillin, and focal adhesion kinase in an actin-dependent manner. J. Immunol. 2002, 168, 5415–5423. [Google Scholar] [CrossRef]
- Jin, Y.P.; Jindra, P.T.; Gong, K.W.; Lepin, E.J.; Reed, E.F. Anti-HLA class I antibodies activate endothelial cells and promote chronic rejection. Transplantation 2005, 15, S19–S21. [Google Scholar] [CrossRef]
- Jin, Y.P.; Fishbein, M.C.; Said, J.W.; Jindra, P.T.; Rajalingam, R.; Rozengurt, E.; Reed, E.F. Anti-HLA class I antibody-mediated activation of the PI3K/Akt signaling pathway and induction of Bcl-2 and Bcl-xL expression in endothelial cells. Hum. Immunol. 2004, 65, 291–302. [Google Scholar] [CrossRef]
- Jindra, P.T.; Zhang, X.; Mulder, A.; Claas, F.; Veale, J.; Jin, Y.P.; Reed, E.F. Anti-HLA antibodies can induce endothelial cell survival or proliferation depending on their concentration. Transplantation 2006, 82 (Suppl. S1), S33–S35. [Google Scholar] [CrossRef]
- Jindra, P.T.; Jin, Y.P.; Rozengurt, E.; Reed, E.F. HLA class I antibody-mediated endothelial cell proliferation via the mTOR pathway. J. Immunol. 2008, 180, 2357–2366. [Google Scholar] [CrossRef]
- Li, F.; Zhang, X.; Jin, Y.P.; Mulder, A.; Reed, E.F. Antibody ligation of human leukocyte antigen class I molecules stimulates migration and proliferation of smooth muscle cells in a focal adhesion kinase-dependent manner. Hum. Immunol. 2011, 72, 1150–1159. [Google Scholar] [CrossRef]
- Ziegler, M.E.; Souda, P.; Jin, Y.P.; Whitelegge, J.P.; Reed, E.F. Characterization of the endothelial cell cytoskeleton following HLA class I ligation. PLoS ONE 2012, 7, e29472. [Google Scholar] [CrossRef] [PubMed]
- Zhang, X.; Valenzuela, N.M.; Reed, E.F. HLA class I antibody-mediated endothelial and smooth muscle cell activation. Curr. Opin. Organ Transpl. 2012, 17, 446–451. [Google Scholar] [CrossRef] [PubMed]
- Lepin, E.J.; Bastin, J.M.; Allan, D.S.; Roncador, G.; Braud, V.M.; Mason, D.Y.; van der Merwe, P.A.; McMichael, A.J.; Bell, J.I.; Powis, S.H.; et al. Functional characterization of HLA-F and binding of HLA-F tetramers to ILT2 and ILT4 receptors. Eur. J. Immunol. 2000, 30, 3552–3561. [Google Scholar] [CrossRef]
- Barnstable, C.J.; Bodmer, W.F.; Brown, G.; Galfre, G.; Milstein, C.; Williams, A.F.; Ziegler, A. Production of monoclonal antibodies to group A erythrocytes HLA and other human cell-surface antigens-new tools for genetic analysis. Cell 1978, 14, 9–20. [Google Scholar] [CrossRef]
- Parham, P.; Barnstable, C.J.; Bodmer, W.F. Use of a monoclonal antibody (W6/32) in structural studies of HLA-A, B, C, antigens. J. Immunol. 1979, 123, 342–349. [Google Scholar]
- Brown, G.; Joshua, D.E.; Bastin, J.; Mason, D.Y.; Barnstable, C.J. Monoclonal antibodies to human cell-surface antigens. Haematol. Blood Transfus. 1979, 23, 365–368. [Google Scholar] [CrossRef]
- Smith, K.D.; Mace, B.E.; Valenzuela, A.; Vigna, J.L.; McCutcheon, J.A.; Barbosa, J.A.; Huczko, E.; Engelhard, V.H.; Lutz, C.T. Probing HLA-B7 conformational shifts induced by peptide-binding groove mutations and bound peptide with anti-HLA monoclonal antibodies. J. Immunol. 1996, 157, 2470–2478. [Google Scholar]
- Vigna, J.L.; Smith, K.D.; Lutz, C.T. Invariant chain association with MHC class I: Preference for HLA class I/beta 2-microglobulin heterodimers, specificity, and influence of the MHC peptide-binding groove. J. Immunol. 1996, 157, 4503–4510. [Google Scholar]
- Ivanyi, D.; van de Meugheuvel, W. A monomorphic HLA-specific monoclonal antibody, W6/32, reacts with the H-2Db molecule of normal mouse lymphocytes. Immunogenetics 1984, 20, 699–703. [Google Scholar] [CrossRef]
- Maziarz, R.T.; Fraser, J.; Strominger, J.L.; Burakoff, S.J. The human HLA-specific monoclonal antibody W6/32 recognizes a discontinuous epitope within the alpha 2 domain of murine H-2Db. Immunogenetics 1986, 24, 206–208. [Google Scholar] [CrossRef]
- Jefferies, W.A.; MacPherson, G.G. Expression of the W6/32 HLA epitope by cells of rat, mouse, human and other species: Critical dependence on the interaction of specific MHC heavy chains with human or bovine beta 2-microglobulin. Eur. J. Immunol. 1987, 17, 1257–1263. [Google Scholar] [CrossRef]
- Kahn-Perles, B.; Boyer, C.; Arnold, B.; Sanderson, A.R.; Ferrier, P.; Lemonnier, F.A. Acquisition of HLA class I W6/32 defined antigenic determinant by heavy chains from different species following association with bovine beta 2-microglobulin. J. Immunol. 1987, 138, 2190–2196. [Google Scholar]
- Kievits, F.; Ivanyi, P. Monomorphic anti-HLA monoclonal antibody (W6/32) recognizes polymorphic H-2 heavy-chain determinants exposed by association with bovine or human but not murine beta 2-microglobulin. Hum. Immunol. 1987, 20, 115–126. [Google Scholar] [CrossRef]
- Shields, M.J.; Ribaudo, R.K. Mapping of the monoclonal antibody W6/32: Sensitivity to the amino terminus of beta2-microglobulin. Tissue Antigens 1998, 51, 567–570. [Google Scholar] [CrossRef]
- Ladasky, J.J.; Shum, B.P.; Canavez, F.; Seuánez, H.N.; Parham, P. Residue 3 of beta2-microglobulin affects binding of class I MHC molecules by the W6/32 antibody. Immunogenetics 1999, 49, 312–320. [Google Scholar] [CrossRef] [PubMed]
- Tran, T.M.; Ivanyi, P.; Hilgert, I.; Brdicka, T.; Pla, M.; Breur, B.; Flieger, M.; Ivasková, E.; Horejsí, V. The epitope recognized by pan-HLA class I-reactive monoclonal antibody W6/32 and its relationship to unusual stability of the HLA-B27/beta2-microglobulin complex. Immunogenetics 2001, 53, 440–446. [Google Scholar] [CrossRef] [PubMed]
- Ravindranath, M.H.; Ravindranath, N.M.; El Hilali, F.; Selvan, S.R.; Filippone, E.J. Ramifications of the HLA-I Allelic Reactivity of Anti-HLA-E*01:01 and Anti-HLA-E*01:03 Heavy Chain Monoclonal Antibodies in Comparison with Anti-HLA-I IgG Reactivity in Non-Alloimmunized Males, Melanoma-Vaccine Recipients, and End-Stage Renal Disease Patients. Antibodies 2022, 11, 18. [Google Scholar] [CrossRef] [PubMed]
- Ravindranath, M.H.; Terasaki, P.I. Diagnostic and Therapeutic Potential of HLA-E Monospecific Monoclonal IgG Antibodies Directed against Tumor Cell-surface and Soluble HLA-E. U.S. Patent 10656156, 5 July 2012. [Google Scholar]
- Sasaki, T.; Ravindranath, M.H.; Terasaki, P.I.; Freitas, M.C.; Kawakita, S.; Jucaud, V. Gastric cancer progression may involve a shift in HLA-E profile from an intact heterodimer to β2-microglobulin-free monomer. Int. J. Cancer 2014, 134, 1558–1570. [Google Scholar] [CrossRef] [PubMed]
- Ravindranath, M.H.; Terasaki, P.I.; Pham, T.; Jucaud, V. The Monospecificity of Novel Anti-HLA-E Monoclonal Antibodies Enables Reliable Immunodiagnosis, Immunomodulation of HLA-E, and Upregulation of CD8+ T Lymphocytes. Monoclon. Antibodies Immunodiagn. Immunother. 2015, 34, 135–153. [Google Scholar] [CrossRef] [PubMed]
- Ravindranath, M.H.; Hopfield, J.; Ferrone, S. HLA-E restricted monoclonal antibodies: Therapeutic potential as a double-edged sword against tumor progression. Intern. Rev. Med. 2017, 3, 1–49. [Google Scholar]
- Ravindranath, M.H.; El Hilali, F. Monospecific and Polyreactive Monoclonal Antibodies against human Leukocyte Antigen-E: Diagnostic and Therapeutic relevance. In Monoclonal Antibodies; Rezaei, N., Ed.; Intech Open: London, UK, 2021; ISBN 978-1-83968-369-5. [Google Scholar]
- Ravindranath, M.H.; Terasaki, P.I. Anti-HLA Class-IB Antibodies Mimic Immunoreactivity and Immunomodulatory Functions of Intravenous Immunoglobulin (IVIG) Useful as Therapeutic IVIG Mimetics and Methods of Their Use. International Patent WO 2013/106586 A2, 19 July 2013. [Google Scholar]
- Zhu, D.; Ravindranath, M.H.; Terasaki, P.I.; Miyazaki, T.; Pham, T.; Jucaud, V. Suppression of allo-human leucocyteantigen (HLA) Abs secreted by B memory cells in vitro: Intravenous immunoglobulin (IVIg) versus a monoclonal anti-HLA-E IgG that mimics HLA-I reactivities of IVIg. Clin. Exp. Immunol. 2014, 177, 464–477. [Google Scholar] [CrossRef]
- Ravindranath, M.H.; Zhu, D.; Pham, T.; Jucaud, V.; Hopfield, J.; Kawakita, S.; Terasaki, P.I. Anti-HLA-E monoclonal antibodies reacting with HLA-la and lb alleles like IVIg as potential IVIg-immunomimetics: An evolving therapeutic concept. Clin. Transpl. 2013, 2013, 293–305. [Google Scholar]
- Ravindranath, M.H. HLA Class Ia and Ib Polyreactive Anti-HLA-E IgG2a Monoclonal Antibodies (TFL-006 and TFL-007) suppress Anti-HLA IgG Production by CD19+ B Cells and Proliferation of CD4+ T-cells While upregulating T-regs. J. Immunol. Res. 2017, 2017, 3475926. [Google Scholar] [CrossRef]
- Majdic, O.; Schnabl, E.; Stockinger, H.; Gadd, S.; Maurer, D.; Radaszkiewics, T. LA45, an activation-induced human lymphocyte antigen with strong homology to MHC class I molecules. In Leukocyte Typing IV; Oxford University Press: Oxford, UK, 1989; pp. 8–511. [Google Scholar]
- Schumacher, T.N.; Heemels, M.T.; Neefjes, J.J.; Kast, W.M.; Melief, C.J.; Ploegh, H.L. Direct binding of peptide to empty MHC class I molecules on intacT-cells and in vitro. Cell 1990, 62, 563–567. [Google Scholar] [CrossRef]
- Benjamin, R.J.; Madrigal, J.A.; Parham, P. Peptide binding to empty HLA-B27 molecules of viable human cells. Nature 1991, 351, 74–77. [Google Scholar] [CrossRef]
- Madrigal, J.A.; Belich, M.P.; Benjamin, R.J.; Little, A.M.; Hildebrand, W.H.; Mann, D.L.; Parham, P. Molecular definition of a polymorphic antigen (LA45) of free HLA-A and -B heavy chains found on the surfaces of activated B and T-cells. J. Exp. Med. 1991, 174, 1085–1095. [Google Scholar] [CrossRef]
- Demaria, S.; Bushkin, Y. CD8 and beta 2microglobulin-free MHC class I molecules in T cell immunoregulation. Int. J. Clin. Lab. Res. 1993, 23, 61–69. [Google Scholar] [CrossRef]
- Capps, G.G.; Robinson, B.E.; Lewis, K.D.; Zúñiga, M.C. In vivo dimeric association of class I MHC heavy chains. Possible relationship to class I MHC heavy chain-beta 2microglobulin dissociation. J. Immunol. 1993, 151, 159–169. [Google Scholar]
- Matko, J.; Bushkin, Y.; Wei, T.; Edidin, M. Clustering of class I HLA molecules on the surfaces of activated and transformed human cells. J. Immunol. 1994, 152, 3353–3360. [Google Scholar]
- Khare, S.D.; Hansen, J.; Luthra, H.S.; David, C.S. HLA-B27 heavy chains contribute to spontaneous inflammatory disease in B27/human beta2microglobulin (beta2m) double transgenic mice with disrupted mouse beta2m. J. Clin. Investig. 1996, 98, 2746–2755. [Google Scholar] [CrossRef]
- Narayanan, K.; Jendrisak, M.D.; Phelan, D.L.; Mohanakumar, T. HLA class I antibody mediated accommodation of endothelial cells via the activation of PI3K/cAMP dependent PKA pathway. Transpl. Immunol. 2006, 15, 187–197. [Google Scholar] [CrossRef]
- Reznik, S.I.; Jaramillo, A.; Zhang, L.; Patterson, G.A.; Cooper, J.D.; Mohanakumar, T. Anti-HLA antibody bindng to HLA class I molecules induces proliferation of airway epithelial cells: A potential mechanism for bronchiolitis obliterans syndrome. J. Thorac. Cardiovasc. Surg. 2000, 119, 39–45. [Google Scholar] [CrossRef]
- Jaramillo, A.; Smith, C.R.; Maruyama, T.; Zhang, L.; Patterson, G.A.; Mohanakumar, T. Anti-HLA class I antibody binding to airway epithelial cells induces production of fibrogenic growth factors and apoptotic cell death: A possible mechanism for bronchiolitis obliterans syndrome. Hum. Immunol. 2003, 64, 521–529. [Google Scholar] [CrossRef]
- Turco, M.C.; De Felice, M.; Corbo, L.; Giarrusso, P.C.; Yang, S.Y.; Ferrone, S.; Venuta, S. Enhancing effect of anti-HLA class I monoclonal antibodies on T cell proliferation induced via CD2 molecule. J. Immunol. 1988, 141, 2275–2281. [Google Scholar]
- Santoli, D.; Radka, S.F.; Kreider, B.L.; Ferrone, S. Regulatory role of monomorphic and polymorphic determinants of HLA class I antigens in the proliferation of T-cells stimulated by autologous and allogeneic B and T lymphoid cells. Cell Immunol. 1988, 116, 263–273. [Google Scholar] [CrossRef]
- Gilliland, L.K.; Norris, N.A.; Grosmaire, L.S.; Ferrone, S.; Gladstone, P.; Ledbetter, J.A. Signal transduction in lymphocyte activation through crosslinking of HLA class I molecules. Hum. Immunol. 1989, 25, 269–289. [Google Scholar] [CrossRef]
- Turco, M.C.; De Felice, M.; Alfinito, F.; Lamberti, A.; Costanzo, F.; Giordano, M.; Martinelli, V.; Rotoli, B.; Ferrone, S.; Venuta, S. Proliferative pathways in CD1- CD3+ CD4+ CD8+ T-prolymphocytic leukemic cells: Analysis with monoclonal antibodies and cytokines. Blood 1989, 74, 1651–1657. [Google Scholar] [CrossRef] [PubMed]
- Taylor, D.S.; Nowell, P.C.; Kornbluth, J. Anti-HLA class I antibodies inhibit the T cell-independent proliferation of human B lymphocytes. J. Immunol. 1987, 139, 1792–1796. [Google Scholar] [PubMed]
- De Felice, M.; Turco, M.C.; Giarrusso, P.C.; Corbo, L.; Pizzano, R.; Martinelli, V.; Ferrone, S.; Venuta, S. Differential regulatory role of monomorphic and polymorphic determinants of histocompatibility leukocyte antigen class I antigens in monoclonal antibody OKT3-induced T cell proliferation. J. Immunol. 1987, 139, 2683–2689. [Google Scholar] [PubMed]
- De Felice, M.; Turco, M.C.; Corbo, L.; Carandente Giarrusso, P.; Lamberti, A.; Valerio, G.; Temponi, M.; Costanzo, F.; Ferrone, S.; Venuta, S. Lack of a role of monocytes in the inhibition by monoclonal antibodies to monomorphic and polymorphic determinants of HLA class I antigens of PHA-P-induced peripheral blood mononuclear cell proliferation. Cell Immunol. 1989, 122, 164–177. [Google Scholar] [CrossRef]
- Huet, S.; Boumsell, L.; Dausset, J.; Degos, L.; Bernard, A. The required interaction between monocytes and peripheral blood T lymphocytes (T-PBL) upon activation via CD2 or CD3. Role of HLA class I molecules from accessory cells and the differential response of T-PBL subsets. Eur. J. Immunol. 1988, 18, 1187–1194. [Google Scholar] [CrossRef]
- Ravindranath, M.H.; Terasaki, P.I.; Pham, T.; Jucaud, V.; Kawakita, S. Suppression of blastogenesis and proliferation of activated CD4(+) T-cells: Intravenous immunoglobulin (IVIg) versus novel anti-human leucocyte antigen (HLA)-E mAbs mimicking HLA-I reactivity of IVIg. Clin. Exp. Immunol. 2014, 178, 154–177. [Google Scholar] [CrossRef]
- Mustelin, T.; Vang, T.; Bottini, T.N. Protein tyrosine phosphatases and the immune response. Nat. Rev. Immunol. 2005, 5, 43–57. [Google Scholar] [CrossRef]
- Modai, D.; Berman, S.; Sheleg, Y.; Cohn, M.; Weissgarten, J.; Averbukh, Z. Interleukin-2 receptor is similarly expressed by activated lymphocytes from patients on chronic hemodialysis and healthy subjects. Clin. Immunol. Immunopathol. 1990, 55, 237–241. [Google Scholar] [CrossRef]
- Stentz, F.B.; Kitabchi, A.E. Transcriptome and proteome expression in activated human CD4 and CD8 T-lymphocytes. Biochem. Biophys. Res. Commun. 2004, 324, 692–696. [Google Scholar] [CrossRef]
- Le Gall, S.; Erdtmann, L.; Benichou, S.; Berlioz-Torrent, C.; Liu, L.; Benarous, R.; Heard, J.M.; Schwartz, O. Nef interacts with the mu subunit of clathrin adaptor complexes and reveals a cryptic sorting signal in MHC I molecules. Immunity 1998, 8, 483–495. [Google Scholar] [CrossRef]
- Cohen, G.B.; Gandhi, R.T.; Davis, D.M.; Mandelboim, O.; Chen, B.K.; Strominger, J.L.; Baltimore, D. The selective downregulation of class I major histocompatibility complex proteins by HIV-1 protects HIV-infected cells from NK cells. Immunity 1999, 10, 661–671. [Google Scholar] [CrossRef]
- Santos, S.G.; Antoniou, A.N.; Sampaio, P.; Powis, S.J.; Arosa, F.A. Lack of tyrosine 320 impairs spontaneous endocytosis and enhances release of HLA-B27 molecules. J. Immunol. 2006, 176, 2942–2949. [Google Scholar] [CrossRef]
- Guild, B.; Strominger, J.L. Human and murine class I MHC antigens share conserved serine 335, the site of HLA phosphorylation in vivo. J. Biol. Chem. 1984, 259, 9235–9240. [Google Scholar] [CrossRef]
- Arosa, F.A.; de Jesus, O.; Porto, G.; Carmo, A.M.; de Sousa, M. Calreticulin is expressed on the cell-surface of activated human peripheral blood T lymphocytes in association with major histocompatibility complex class I molecules. J. Biol. Chem. 1999, 274, 16917–16922. [Google Scholar] [CrossRef]
- Allen, R.L.; Trowsdale, J. Recognition of classical and heavy chain forms of HLA-B27 by leukocyte receptors. Curr. Mol. Med. 2004, 4, 59–65. [Google Scholar] [CrossRef]
- Jones, D.C.; Kosmoliaptsis, V.; Apps, R.; Lapaque, N.; Smith, I.; Kono, A.; Chang, C.; Boyle, L.H.; Taylor, C.J.; Trowsdale, J.; et al. HLA class I allelic sequence and conformation regulate leukocyte Ig-like receptor binding. J. Immunol. 2011, 186, 2990–2997. [Google Scholar] [CrossRef]
- Tamir, I.; Cambier, J.C. Antigen receptor signaling: Integration of protein tyrosine kinase functions. Oncogene 1988, 17, 135364. [Google Scholar] [CrossRef]
- EL Hilali, F.; Jucaud, V.; EL Hilali, H.; Bhuiyan, M.H.; Mancuso, A.; LiuSullivan, N.; Elidrissi, A.; Mazouz, H. Characterization of the Anti-HLA Class I and II IgG Antibodies in Moroccan IVIg Using Regular Beads and Ibeads in Luminex Multiplex Single Antigen Immunoassay. Int. J. Immunol. 2017, 5, 53–65. [Google Scholar] [CrossRef][Green Version]
- Hara, M.; Kingsley, C.I.; Niimi, M.; Read, S.; Turvey, S.E.; Bushell, A.R.; Morris, P.J.; Powrie, F.; Wood, K.J. IL-10 is required for regulatory T-cells to mediate tolerance to alloantigens in vivo. J. Immunol. 2001, 166, 3789–3796. [Google Scholar] [CrossRef]
- Levings, M.; Sangregorio, R.; Roncarolo, M.G. Human CD25+CD4+ T regulatory cells suppress naive and memory T-cell proliferation and can be expanded in vitro without loss of function. J. Exp. Med. 2001, 193, 1295–1301. [Google Scholar] [CrossRef]
- Van Maurik, A.; Wood, K.J.; Jones, N. Cutting edge: CD4+CD25+ alloantigen-specific immunoregulatory cells that can prevent CD8+ T-cell-mediated graft rejection: Implications for anti-CD154 immunotherapy. J. Immunol. 2002, 169, 5401–5404. [Google Scholar] [CrossRef]
- Jia, S.; Li, C.; Wang, G.; Yang, J.; Zu, Y. The T helper type 17/regulatory T cell imbalance in patients with acute Kawasaki disease. Clin. Exp. Immunol. 2010, 162, 131–137. [Google Scholar] [CrossRef]
- Kasztalska, K.; Ciebiada, M.; Cebula-Obrzut, B.; Górski, P. Intravenous immunoglobulin replacement therapy in the treatment of patients with common variable immunodeficiency disease: An open-label prospective study. Clin. Drug Investig. 2011, 31, 299–307. [Google Scholar] [CrossRef]
- Levy, E.M.; Bianchini, M.; Von Euw, E.M.; Barrio, M.M.; Bravo, A.I.; Furman, D.; Domenichini, E.; Macagno, C.; Pinsky, V.; Zucchini, C.; et al. Human leukocyte antigen-E protein is overexpressed in primary human colorectal cancer. Int. J. Oncol. 2008, 32, 633–641. [Google Scholar] [CrossRef]
- Gonçalves, M.A.; Le Discorde, M.; Simões, R.T.; Rabreau, M.; Soares, E.G.; Donadi, E.A.; Carosella, E.D. Classical and non-classical HLA molecules and p16 (INK4a) expression in precursors’ lesions and invasive cervical cancer. Eur. J. Obstet. Gynecol. Reprod. Biol. 2008, 141, 70–74. [Google Scholar] [CrossRef] [PubMed]
- Huang, Z.; Hyodo, H.; Fujii, T.; Nagamatsu, T.; Matsumoto, J.; Kawana, K.; Yamashita, T.; Yasugi, T.; Kozuma, S.; Taketani, Y. Effect of progesterone on HLA-E gene expression in JEG-3 choriocarcinoma cell line. Am. J. Reprod. Immunol. 2009, 61, 221–226. [Google Scholar] [CrossRef] [PubMed]
- Chen, A.; Shen, Y.; Xia, M.; Xu, L.; Pan, N.; Yin, Y.; Miao, F.; Shen, C.; Xie, W.; Zhang, J. Expression of the nonclassical HLA class I and MICA/B molecules in human hepatocellular carcinoma. Neoplasma 2011, 58, 371–376. [Google Scholar] [CrossRef] [PubMed]
- Kren, L.; Slaby, O.; Muckova, K.; Lzicarova, E.; Sova, M.; Vybihal, V.; Svoboda, T.; Fadrus, P.; Lakomy, R.; Vanhara, P.; et al. Expression of immune-modulatory molecules HLA-G and HLA-E by tumor cells in glioblastomas: An unexpected prognostic significance? Neuropathology 2011, 31, 129–134. [Google Scholar] [CrossRef] [PubMed]
- Kren, L.; Fabian, P.; Slaby, O.; Janikova, A.; Soucek, O.; Sterba, J.; Krenova, Z.; Michalek, J.; Kral, Z. Multifunctional immune-modulatory protein HLA-E identified in classical Hodgkin lymphoma: Possible implications. Pathol. Res. Pract. 2012, 208, 45–49. [Google Scholar] [CrossRef]
- Tremante, E.; Ginebri, A.; Lo Monaco, E.; Benassi, B.; Frascione, P.; Grammatico, P.; Cappellacci, S.; Catricalà, C.; Arcelli, D.; Natali, P.G.; et al. A melanoma immune response signature including Human Leukocyte Antigen-E. Pigment. Cell Melanoma Res. 2014, 27, 103–112. [Google Scholar] [CrossRef]
- De Kruijf, E.M.; Sajet, A.; van Nes, J.G.; Natanov, R.; Putter, H.; Smit, V.T.; Liefers, G.J.; van den Elsen, P.J.; van de Velde, C.J.; Kuppen, P.J. HLA-E and HLA-G expression in classical HLA class I-negative tumors is of prognostic value for clinical outcome of early breast cancer patients. J. Immunol. 2010, 185, 7452–7459. [Google Scholar] [CrossRef]
- Bossard, C.; Bezieau, S.; Matysiak-Budnik, T.; Volteau, C.; Laboisse, C.L.; Jotereau, F.; Mosnier, J.-F. HLA-E/beta2 microglobulin overexpression in colorectal cancer is associated with recruitment of inhibitory immune cells and tumor progression. Int. J. Cancer 2012, 131, 855–863. [Google Scholar] [CrossRef]
- Dutta, N.; Majumder, D.; Gupta, A.; Mazumder, D.N.; Banerjee, S. Analysis of human lymphocyte antigen class I expression in gastric cancer by reverse transcriptase-polymerase chain reaction. Hum. Immunol. 2005, 66, 164–169. [Google Scholar] [CrossRef]
- Van Esch, E.M.; Tummers, B.; Baartmans, V.; Osse, E.M.; Ter Haar, N.; Trietsch, M.D.; Hellebrekers, B.W.; Holleboom, C.A.; Nagel, H.T.; Tan, L.T.; et al. Alterations in classical and nonclassical HLA expression in recurrent and progressive HPV-induced usual vulvar intraepithelial neoplasia and implications for immunotherapy. Int. J. Cancer 2014, 135, 830–842. [Google Scholar] [CrossRef]
- Bianchini, M.; Levy, E.; Zucchini, C.; Pinski, V.; Macagno, C.; De Sanctis, P.; Valvassori, L.; Carinci, P.; Mordoh, J. Comparative study of gene expression by cDNA microarray in human colorectal cancer tissues and normal mucosa. Int. J. Oncol. 2006, 29, 83–94. [Google Scholar] [CrossRef]
- Dutta, N.; Gupta, A.; Mazumder, D.N.; Banerjee, S. Down-regulation of locus-specific human lymphocyte antigen class I expression in Epstein-Barr virus-associated gastric cancer: Implication for viral-induced immune evasion. Cancer 2006, 106, 1685–1693. [Google Scholar] [CrossRef]
- Zhang, X.; Lin, A.; Zhang, J.G.; Bao, W.G.; Xu, D.P.; Ruan, Y.Y.; Yan, W.H. Alteration of HLA-F and HLA I antigen expression in the tumor is associated with survival in patients with esophageal squamous cell carcinoma. Int. J. Cancer 2013, 132, 82–89. [Google Scholar] [CrossRef]
- Ashrafi, G.H.; Haghshenas, M.R.; Marchetti, B.; O’Brien, P.M.; Campo, M.S. E5 protein of human papillomavirus type 16 selectively downregulates surface HLA class I. Int. J. Cancer 2005, 113, 276–283. [Google Scholar] [CrossRef]
- Näsman, A.; Andersson, E.; Nordfors, C.; Grün, N.; Johansson, H.; Munck-Wikland, E.; Massucci, G.; Dalianis, T.; Ramqvist, T. MHC class I expression in HPV positive and negative tonsillar squamous cell carcinoma in correlation to clinical outcome. Int. J. Cancer 2013, 132, 72–81. [Google Scholar] [CrossRef] [PubMed]
- Campoli, M.; Ferrone, S. HLA antigen changes in malignanT-cells: Epigenetic mechanisms and biologic significance. Oncogene 2008, 27, 5869–5885. [Google Scholar] [CrossRef] [PubMed]
- Petrie, E.J.; Clements, C.S.; Lin, J.; Sullivan, L.C.; Johnson, D.; Huyton, T.; Heroux, A.; Hoare, H.L.; Beddoe, T.; Reid, H.H.; et al. CD94-NKG2A recognition of human leukocyte antigen (HLA)-E bound to an HLA class I leader sequence. J. Exp. Med. 2008, 205, 725–735. [Google Scholar] [CrossRef] [PubMed]
- Schell, T.D.; Mylin, L.M.; Tevethia, S.S.; Joyce, S. The assembly of functional beta(2)-microglobulin-free MHC class I molecules that interact with peptides and CD8(+) T lymphocytes. Int. Immunol. 2002, 14, 775–782. [Google Scholar] [CrossRef]
- Ding, J.; Feng, Y.; Zheng, Z.H.; Li, X.Y.; Wu, Z.B.; Zhu, P. Increased expression of human leucocyte antigen class I free heavy chains on monocytes of patients with spondyloarthritis and cells transfected with HLA-B27. Int. J. Immunogenet. 2015, 42, 4–10. [Google Scholar] [CrossRef]
- Pröll, J.; Blaschitz, A.; Hutter, H.; Dohr, G. First trimester human endovascular trophoblasT-cells express both HLA-C and HLA-G. Am. J. Reprod. Immunol. 1999, 42, 30–36. [Google Scholar] [CrossRef] [PubMed]
- Blaschitz, A.; Hutter, H.; Dohr, G. HLA Class I protein expression in the human placenta. Early Pregnancy 2001, 5, 67–69. [Google Scholar] [PubMed]
- Siegel, R.J.; Bridges, L., Jr.; Ahmed, S. HLA-C: An Accomplice in Rheumatic Diseases. ACR Open Rheumatol. 2019, 1, 571–579. [Google Scholar] [CrossRef]
- Gambino, C.M.; Di Bona, D.; Aiello, A.; Carru, C.; Duro, G.; Guggino, G.; Ferrante, A.; Zinellu, A.; Caruso, C.; Candore, G.; et al. HLA-C1 ligands are associated with increased susceptibility to systemic lupus erythematosus. Hum. Immunol. 2018, 79, 172–177. [Google Scholar] [CrossRef]

















{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| HLA Class I Alleles | HLA Class II Alleles | |||||||
|---|---|---|---|---|---|---|---|---|
| α-Chain | α-Chain | β-Chain | ||||||
| Genes | Alleles | Proteins | Genes | Alleles | Proteins | Genes | Alleles | Proteins |
| A | 7452 | 4355 | DRA | 32 | 5 | DRB1 | 3196 | 2152 |
| B | 8849 | 5343 | DRB2 | 1 | 0 | |||
| C | 7393 | 4095 | DRB3 | 423 | 314 | |||
| E | 310 | 121 | DRB4 | 215 | 139 | |||
| F | 50 | 7 | DRB5 | 171 | 131 | |||
| G | 102 | 35 | DQA1 | 442 | 205 | DQB1 | 2230 | 1407 |
| DQA2 | 40 | 11 | ||||||
| DPA1 | 406 | 173 | DPB1 | 1958 | 1223 | |||
| HLA-I Isoforms and Domains | ||||||||||||||||||||||
|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|
| α1-Domain | α2-Domain | α3-Domain | ||||||||||||||||||||
| A | 25–114 | 115–200 | 201–298 | |||||||||||||||||||
| B | 25–114 | 115–200 | 201–298 | |||||||||||||||||||
| C | 25–114 | 115–200 | 201–298 | |||||||||||||||||||
| Amino Acid Sequences Shared by Almost All Known Alleles of Hla-A, -B, -C, -E, -F and -G, and Their Positions in CD8 Binding α3-Domain | ||||||||||||||||||||||
| 222 | E | A | T | L | R | C | W | A | L | G | F | Y | P | A | E | I | T | L | T | W | Q | 242 |
| 244 | D | G | E | G | H | T | Q | D | 251 | |||||||||||||
| 253 | E | L | V | E | T | R | P | A | G | D | G | T | F | Q | K | W | A | 269 | ||||
| HLA-II Isoforms and Domains | ||||||||||||||||||||||
| α1-Domain | α2-Domain | β1-Domain | β2-Domain | |||||||||||||||||||
| DRA | 26–109 | 110–203 | DRB | 30–124 | 125–227 | |||||||||||||||||
| DQA | 24–119 | 120–203 | DQB | 30–121 | 122–215 | |||||||||||||||||
| DPA | 29–115 | 116–209 | DPB | 33–126 | 127–229 | |||||||||||||||||
| Amino Acid Sequences Shared by Almost All Alleles of HLA-DR,-DQ and DP & Their Positions in CD4 Binding β2-Domain | ||||||||||||||||||||||
| DRB | 150 | N | G | D | W | T | F | Q | T | L | V | M | L | E | 162 | |||||||
| 185 | T | V | E | W | R | A | R | S | E | S | A | Q | S | K | 198 | |||||||
| DQB | 150 | N | G | D | W | T | F | Q | T | L | V | M | L | E | 162 | |||||||
| 185 | T | V | E | W | R | A | R | S | E | S | A | Q | S | K | 198 | |||||||
| DPB | 148 | N | G | D | W | T | F | Q | T | L | V | M | L | E | 160 | |||||||
| 183 | T | V | E | W | R | A | R | S | E | S | A | Q | S | K | 196 | |||||||
| HLA-E Peptide Sequences | HLA Alleles | Frequency of Occurrence in HLA by Rank | Prediction Scores | Immunogenicity Rank | |||||||
|---|---|---|---|---|---|---|---|---|---|---|---|
| Classical HLA-Ia | Non-Classical HLA-Ib | Method 1 | Method 2 | Method 3 | Method 4 | ||||||
| [Total Number of Amino Acids] | A | B | Cw | F | G | Beta-Turn | Antigenicity | Flexibility | Hydrophilicity | ||
| Chou & Fasman [7] | Kolaskar & Tangaonkar [8] | Karplus & Schulz [9] | Parker, et al. [10] | ||||||||
| 141AYDGKDY147 [11] | 491 | 831 | 271 | 21 | 30 | 1 | 1.204 | 0.989 | 1.061 | 4.243 | 1 |
| 150LNEDLRSWTA159 [12] | 239 | 219 | 261 | 21 | 30 | 2 | 1.046 | 0.983 | 1.039 | 2.443/2.329 | 2 |
| 161DTAAQI166 [6] | 0 | 824 | 248 | 0 | 30 | 3 | 0.813 | 1.065 | 0.978 | 1.957 | 3 |
| 187TCVEWL192 [6] | 282 | 206 | 200 | 0 | 30 | 4 | 0.841 | 1.115 | 0.929 | −0.914 | 4 |
| PtGF | KtF | PjGS | MrF | NrS1 | HrS2 | Ho | Rn | Th | Sk | Vd | Me | Cr | Ad | Ra | ||
|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|
| NCAlbumin | 25 | 190 | 120 | 36 | 943 | 503 | 55 | 537 | 91 | |||||||
| PC | 788 | 791 | 1977 | 1317 | 1915 | 1274 | 1418 | 909 | ||||||||
| Anti-HLA-A IgG | A*0101 | 811 | 856 | 1962 | 1372 | 3592 | 1326 | 880 | 1393 | 568 | 8337 | 2847 | ||||
| A*0201 | 588 | 595 | 782 | 770 | 2798 | 1006 | ||||||||||
| A*0203 | 725 | 976 | 765 | 3576 | 1293 | |||||||||||
| A*0206 | 620 | 617 | 1959 | 610 | ||||||||||||
| A*0301 | 2564 | 3091 | 882 | 502 | 3837 | 2258 | 558 | |||||||||
| A*1101 | 1386 | 1413 | ||||||||||||||
| A*1102 | 722 | 1003 | 3528 | 1005 | 2776 | 903 | 6486 | 2080 | ||||||||
| A*2301 | 556 | 5166 | 1956 | 610 | 2914 | 3253 | 1071 | |||||||||
| A*2402 | 588 | 5003 | 2331 | 3357 | 3602 | 978 | ||||||||||
| A*2403 | 513 | 809 | 3331 | 511 | 1984 | 1159 | 1465 | 5223 | 1429 | |||||||
| A*2501 | 1112 | 523 | 5187 | 3482 | 546 | 2767 | 2174 | |||||||||
| A*2601 | 775 | 3042 | 1074 | 954 | 4224 | 2621 | ||||||||||
| A*2901 | 657 | 553 | 1361 | 534 | ||||||||||||
| A*2902 | 506 | 593 | 968 | |||||||||||||
| A*3001 | 718 | 1103 | 605 | 1680 | 3314 | 1416 | ||||||||||
| A*3002 | 1021 | 525 | 590 | 2005 | 885 | |||||||||||
| A*3101 | 877 | 1743 | 586 | 3954 | 1632 | |||||||||||
| A*3201 | 2600 | 8422 | 1142 | 2692 | 577 | |||||||||||
| A*3301 | 864 | 2464 | 5584 | 914 | 511 | |||||||||||
| A*3303 | 3138 | 767 | ||||||||||||||
| A*3401 | 1385 | 2810 | 1455 | 853 | ||||||||||||
| A*3402 | 667 | 2179 | 1594 | 1386 | 535 | |||||||||||
| A*3601 | 508 | 785 | 5724 | 574 | 4266 | 1373 | ||||||||||
| A*4301 | 1489 | 506 | 1632 | 567 | ||||||||||||
| A*6601 | 2102 | 1618 | 2506 | 2088 | ||||||||||||
| A*6602 | 1021 | 810 | 671 | 2815 | 526 | |||||||||||
| A*6801 | 1426 | 3008 | 1374 | 1941 | 712 | |||||||||||
| A*6802 | 1372 | 587 | 585 | 2713 | 877 | |||||||||||
| A*6901 | 548 | 638 | 1579 | 2336 | 512 | 895 | 2050 | 6736 | 2171 | |||||||
| A*7401 | 917 | 2817 | 2176 | |||||||||||||
| A*8001 | 994 | 798 | 1733 | 1536 | 4700 | 1828 | 2457 | 683 | 609 | 1214 | 3009 | 1132 | ||||
| Anti-HLA-B IgG | B*0702 | 791 | 645 | 846 | ||||||||||||
| B*0801 | 605 | 1738 | 1449 | 2221 | 1068 | 503 | 6274 | 2380 | 1261 | |||||||
| B*1301 | 1143 | 663 | ||||||||||||||
| B*1302 | 849 | |||||||||||||||
| B*1401 | 670 | |||||||||||||||
| B*1402 | 1235 | 506 | 555 | |||||||||||||
| B*1501 | 1144 | 581 | ||||||||||||||
| B*1502 | 2104 | 4381 | ||||||||||||||
| B*1503 | 1177 | |||||||||||||||
| B*1510 | 597 | 1729 | 866 | 784 | 2226 | |||||||||||
| B*1511 | 1365 | 1075 | 3617 | 1153 | 627 | 604 | 1441 | 990 | 1507 | 545 | 667 | |||||
| B*1512 | 550 | 2487 | 768 | 1634 | 508 | 572 | 805 | 1149 | ||||||||
| B*1513 | 507 | 2356 | 643 | 1906 | ||||||||||||
| B*1516 | 3544 | |||||||||||||||
| B*1801 | 1342 | 807 | 568 | |||||||||||||
| B*3501 | 1244 | 6375 | ||||||||||||||
| B*3701 | 1137 | |||||||||||||||
| B*3801 | 1720 | |||||||||||||||
| B*3901 | 616 | |||||||||||||||
| B*4001 | 582 | |||||||||||||||
| B*4002 | 1095 | |||||||||||||||
| B*4006 | 671 | 818 | ||||||||||||||
| B*4101 | 774 | |||||||||||||||
| B*4201 | 531 | 915 | ||||||||||||||
| B*4402 | 994 | 590 | ||||||||||||||
| B*4501 | 1412 | |||||||||||||||
| B*4601 | 674 | 817 | 2203 | 1314 | 823 | 1104 | 503 | 1086 | 1982 | 740 | ||||||
| B*4701 | 669 | |||||||||||||||
| B*4801 | 842 | 545 | ||||||||||||||
| B*4901 | 747 | 4828 | 986 | |||||||||||||
| B*5001 | 710 | 1753 | 1091 | |||||||||||||
| B*5101 | 529 | 5536 | 678 | 1550 | ||||||||||||
| B*5102 | 556 | 5125 | 733 | 610 | 8066 | |||||||||||
| B*5201 | 1844 | 694 | ||||||||||||||
| B*5301 | 1013 | 676 | 2216 | 600 | 1218 | 702 | 2217 | |||||||||
| B*5401 | 688 | 553 | 1946 | 555 | 814 | 506 | 808 | |||||||||
| B*5501 | 853 | 514 | ||||||||||||||
| B*5601 | 578 | 1020 | 730 | |||||||||||||
| B*5701 | 2892 | 11,023 | 1346 | 4190 | ||||||||||||
| B*5703 | 1821 | 7141 | 1143 | 3874 | ||||||||||||
| B*5801 | 1864 | 10,634 | 500 | 2602 | ||||||||||||
| B*5901 | 1022 | 1152 | 5069 | 916 | 651 | 1525 | 845 | 6396 | 705 | |||||||
| B*6701 | 796 | 543 | 517 | 1089 | 990 | |||||||||||
| B*7301 | 1185 | 679 | 622 | |||||||||||||
| B*7801 | 3702 | 4296 | ||||||||||||||
| B*8101 | 827 | 789 | 526 | 553 | ||||||||||||
| B*8201 | 707 | 570 | 1627 | 1439 | 715 | 1358 | 798 | 591 | 1140 | |||||||
| Anti-HLA-C IgG | C*0102 | 1219 | 733 | 920 | 1503 | 1287 | 1079 | 717 | ||||||||
| C*0202 | 568 | 712 | 1160 | 1012 | 569 | 874 | 772 | 1088 | ||||||||
| C*0302 | 717 | 916 | 1205 | 1516 | 556 | 972 | 833 | |||||||||
| C*0303 | 1034 | 907 | 1248 | 1575 | 657 | 568 | 1414 | |||||||||
| C*0304 | 712 | 932 | 1080 | 1567 | 666 | 503 | 799 | |||||||||
| C*0401 | 1470 | 530 | 607 | 983 | 799 | 1049 | 689 | 844 | 1139 | |||||||
| C*0501 | 1146 | 778 | 551 | 803 | ||||||||||||
| C*0602 | 913 | 1001 | 1890 | 7846 | 1287 | 1458 | 1834 | 1137 | 866 | 1293 | 1553 | |||||
| C*0702 | 841 | 1344 | 1666 | 1531 | 1162 | 1226 | 1307 | 580 | 808 | 1325 | ||||||
| C*0801 | 762 | 733 | 526 | 681 | ||||||||||||
| C*1203 | 1107 | 2244 | 601 | 1409 | 1188 | 696 | 639 | |||||||||
| C*1402 | 545 | 794 | 900 | 784 | 1345 | 658 | 949 | 561 | 1347 | |||||||
| C*1502 | 578 | 1027 | 3112 | 712 | 848 | 1314 | 1210 | 1391 | 743 | |||||||
| C*1601 | 618 | 856 | 1856 | 539 | 535 | 801 | 608 | 900 | ||||||||
| C*1701 | 933 | 1202 | 1696 | 1568 | 1651 | 677 | 1304 | 589 | 2376 | 1684 | ||||||
| C*1802 | 601 | 531 | 1210 | 2987 | 1578 | 678 | 1204 | 601 |
| Mr | Ho | Rn | Th | Sk | Vd | Me | Cr | Ad | ||
|---|---|---|---|---|---|---|---|---|---|---|
| NC (Albumin) | 862 | 2101 | 940 | 1652 | 5052 | 1451 | 874 | 2752 | 1326 | |
| Anti-HLA-A IgG profile after affinity purification | A*0101 | 631 | 1697 | 747 | 1255 | 1352 | 505 | 646 | 1895 | 804 |
| A*0201 | 661 | 1563 | 797 | 1209 | 1668 | 648 | 2172 | 878 | ||
| A*0203 | 712 | 1641 | 756 | 1155 | 1727 | 604 | 2064 | 886 | ||
| A*0206 | 789 | 1804 | 911 | 1324 | 2127 | 518 | 730 | 2650 | 965 | |
| A*0301 | 833 | 2197 | 911 | 1587 | 1898 | 555 | 733 | 2408 | 1018 | |
| A*1101 | 970 | 2645 | 1066 | 1639 | 2053 | 730 | 808 | 2892 | 1132 | |
| A*1102 | 741 | 1684 | 1056 | 1277 | 1662 | 579 | 637 | 2044 | 949 | |
| A*2301 | 897 | 2980 | 1133 | 1857 | 2007 | 688 | 1074 | 2841 | 1193 | |
| A*2402 | 1196 | 2967 | 1299 | 2284 | 2576 | 776 | 1175 | 3558 | 1437 | |
| A*2403 | 1016 | 2446 | 1108 | 1931 | 2261 | 687 | 894 | 2519 | 1238 | |
| A*2501 | 702 | 1847 | 804 | 1207 | 1576 | 505 | 693 | 2322 | 942 | |
| A*2601 | 1104 | 3401 | 1136 | 1976 | 2125 | 797 | 988 | 2721 | 1272 | |
| A*2901 | 902 | 2911 | 1013 | 1720 | 1640 | 778 | 892 | 2870 | 1090 | |
| A*2902 | 927 | 4136 | 1044 | 1781 | 2150 | 784 | 869 | 2832 | 1145 | |
| A*3001 | 1063 | 2421 | 1080 | 1751 | 3524 | 702 | 891 | 2540 | 1227 | |
| A*3002 | 686 | 1780 | 797 | 1286 | 1631 | 519 | 636 | 2077 | 896 | |
| A*3101 | 656 | 2074 | 741 | 1164 | 1746 | 615 | 2351 | 801 | ||
| A*3201 | 779 | 3294 | 866 | 1523 | 1533 | 660 | 772 | 2249 | 966 | |
| A*3301 | 994 | 2807 | 958 | 1761 | 2475 | 669 | 835 | 2668 | 1111 | |
| A*3303 | 752 | 2229 | 806 | 1357 | 1804 | 646 | 2177 | 891 | ||
| A*3401 | 1141 | 3851 | 1162 | 2004 | 2741 | 858 | 1121 | 2746 | 1272 | |
| A*3402 | 688 | 1675 | 778 | 1258 | 1546 | 513 | 663 | 2195 | 861 | |
| A*3601 | 881 | 1977 | 924 | 1510 | 1848 | 652 | 750 | 2257 | 1010 | |
| A*4301 | 1114 | 3782 | 1180 | 2081 | 2452 | 709 | 963 | 2308 | 1342 | |
| A*6601 | 924 | 3247 | 935 | 1575 | 2620 | 615 | 785 | 2061 | 1066 | |
| A*6602 | 983 | 3189 | 972 | 1759 | 3099 | 643 | 826 | 2226 | 1069 | |
| A*6801 | 651 | 1427 | 705 | 1236 | 1587 | 673 | 1914 | 775 | ||
| A*6802 | 932 | 2216 | 1065 | 1555 | 2094 | 989 | 750 | 3338 | 983 | |
| A*6901 | 1262 | 2777 | 1314 | 2237 | 2614 | 835 | 1138 | 2835 | 1521 | |
| A*7401 | 875 | 2719 | 897 | 1519 | 1734 | 602 | 755 | 1802 | 1025 | |
| A*8001 | 931 | 2885 | 1033 | 1676 | 2282 | 730 | 876 | 2141 | 1127 | |
| Anti-HLA-B IgG profile after affinity purification | B*0702 | 699 | 1549 | 825 | 1193 | 1792 | 657 | 2036 | 839 | |
| B*0801 | 796 | 2020 | 890 | 1447 | 1650 | 556 | 703 | 2576 | 975 | |
| B*1301 | 1331 | 3178 | 1356 | 2231 | 2725 | 817 | 1088 | 3103 | 1461 | |
| B*1302 | 967 | 2500 | 1096 | 1639 | 2385 | 660 | 837 | 2697 | 1126 | |
| B*1401 | 1198 | 2702 | 1231 | 2022 | 2203 | 950 | 1162 | 2876 | 1368 | |
| B*1402 | 977 | 2439 | 1093 | 1797 | 2176 | 748 | 919 | 2324 | 1093 | |
| B*1501 | 747 | 1602 | 860 | 1329 | 2026 | 718 | 2134 | 945 | ||
| B*1502 | 942 | 2610 | 1070 | 1764 | 2453 | 637 | 928 | 2745 | 1233 | |
| B*1503 | 986 | 2277 | 1039 | 1724 | 2208 | 595 | 739 | 2588 | 1130 | |
| B*1510 | 965 | 2934 | 1047 | 1714 | 2138 | 742 | 887 | 2357 | 1175 | |
| B*1511 | 1443 | 3526 | 1473 | 2466 | 2989 | 1010 | 1169 | 3462 | 1567 | |
| B*1512 | 751 | 2163 | 829 | 1496 | 2721 | 2523 | 3030 | 1147 | ||
| B*1513 | 1232 | 2917 | 1321 | 2105 | 2597 | 839 | 1097 | 2868 | 1455 | |
| B*1516 | 1205 | 4241 | 1389 | 2398 | 2725 | 1360 | 1199 | 3158 | 1503 | |
| B*1801 | 752 | 2024 | 853 | 1400 | 1641 | 565 | 658 | 2196 | 912 | |
| B*2705 | 595 | 1378 | 681 | 1084 | 2021 | 589 | 1990 | 764 | ||
| B*2708 | 551 | 1514 | 651 | 932 | 1669 | 520 | 2107 | 711 | ||
| B*3501 | 940 | 2199 | 1003 | 1667 | 2216 | 608 | 864 | 3054 | 1195 | |
| B*3701 | 1518 | 4054 | 1967 | 2613 | 2897 | 949 | 1259 | 3369 | 1540 | |
| B*3801 | 1191 | 2110 | 1095 | 1706 | 2060 | 621 | 853 | 2778 | 1160 | |
| B*3901 | 575 | 1291 | 657 | 1051 | 1446 | 520 | 1800 | 757 | ||
| B*4001 | 781 | 1980 | 910 | 1448 | 1761 | 520 | 698 | 3228 | 980 | |
| B*4002 | 624 | 1669 | 713 | 1124 | 1313 | 575 | 2044 | 783 | ||
| B*4006 | 1354 | 3682 | 1403 | 2280 | 2773 | 1006 | 1109 | 3287 | 1511 | |
| B*4101 | 760 | 1823 | 848 | 1413 | 1659 | 528 | 688 | 2299 | 940 | |
| B*4201 | 694 | 1603 | 765 | 1160 | 2014 | 632 | 2121 | 857 | ||
| B*4402 | 1121 | 2455 | 1196 | 1885 | 2067 | 710 | 2587 | 2575 | 1230 | |
| B*4403 | 811 | 2028 | 1044 | 1382 | 1972 | 515 | 2353 | 2474 | 932 | |
| B*4501 | 756 | 1814 | 786 | 1325 | 1780 | 500 | 2709 | 2221 | 918 | |
| B*4601 | 1090 | 2941 | 1466 | 2220 | 2396 | 898 | 1163 | 3297 | 1470 | |
| B*4701 | 879 | 2687 | 994 | 1729 | 1867 | 670 | 823 | 2567 | 1064 | |
| B*4801 | 1015 | 2291 | 1088 | 1769 | 1908 | 770 | 858 | 3066 | 1173 | |
| B*4901 | 929 | 2200 | 978 | 1725 | 2046 | 580 | 820 | 2569 | 1168 | |
| B*5001 | 675 | 1898 | 784 | 1207 | 1563 | 500 | 607 | 2152 | 880 | |
| B*5101 | 1118 | 2580 | 1214 | 2079 | 2259 | 724 | 973 | 2494 | 1355 | |
| B*5102 | 1116 | 2754 | 1140 | 1997 | 2271 | 842 | 976 | 2609 | 1341 | |
| B*5201 | 1050 | 3355 | 1227 | 1934 | 2261 | 744 | 982 | 2391 | 1299 | |
| B*5301 | 1251 | 3024 | 1359 | 2264 | 2474 | 845 | 1100 | 2725 | 1469 | |
| B*5401 | 1011 | 2528 | 1101 | 1737 | 2264 | 748 | 859 | 2661 | 1192 | |
| B*5501 | 955 | 2327 | 1020 | 1629 | 2089 | 599 | 806 | 2427 | 1111 | |
| B*5601 | 907 | 2698 | 1025 | 1594 | 2244 | 553 | 843 | 2667 | 1186 | |
| B*5701 | 762 | 2155 | 937 | 1512 | 2081 | 538 | 723 | 2144 | 990 | |
| B*5703 | 995 | 2740 | 1171 | 1864 | 2345 | 746 | 953 | 2789 | 1218 | |
| B*5801 | 1338 | 3218 | 1424 | 2331 | 3213 | 924 | 1185 | 2930 | 1523 | |
| B*5901 | 1155 | 2730 | 1207 | 1960 | 2502 | 766 | 1002 | 2600 | 1377 | |
| B*6701 | 661 | 1601 | 827 | 1180 | 1895 | 709 | 700 | 2412 | 841 | |
| B*7301 | 842 | 2326 | 960 | 1590 | 2458 | 555 | 932 | 2132 | 1085 | |
| B*7801 | 1150 | 2527 | 1186 | 1964 | 2544 | 743 | 948 | 2538 | 1346 | |
| B*8101 | 910 | 2241 | 1117 | 1658 | 2130 | 649 | 790 | 3022 | 1100 | |
| B*8201 | 2293 | 2886 | 1288 | 2311 | 2168 | 864 | 1006 | 2819 | 1445 | |
| Anti-HLA-C IgG profile after affinity purification | C*0102 | 1740 | 4894 | 1735 | 2972 | 2873 | 1221 | 1397 | 3892 | 1684 |
| C*0202 | 1941 | 4568 | 1894 | 3222 | 3936 | 1410 | 1504 | 3505 | 2011 | |
| C*0302 | 1830 | 4391 | 1829 | 3077 | 3369 | 1169 | 1512 | 3621 | 1967 | |
| C*0303 | 1663 | 3828 | 1602 | 2839 | 2851 | 1182 | 1409 | 3076 | 1849 | |
| C*0304 | 1900 | 4334 | 1801 | 2974 | 3272 | 1234 | 1538 | 3437 | 1977 | |
| C*0401 | 2166 | 6468 | 2126 | 3870 | 3863 | 1587 | 1897 | 3951 | 2305 | |
| C*0501 | 1967 | 3806 | 1726 | 3015 | 3404 | 1216 | 1912 | 3560 | 1886 | |
| C*0602 | 2165 | 4446 | 2175 | 3835 | 3672 | 1627 | 1732 | 4236 | 2108 | |
| C*0702 | 2551 | 5861 | 2388 | 4231 | 4327 | 1721 | 2066 | 4513 | 2605 | |
| C*0801 | 1510 | 3273 | 1467 | 2541 | 2896 | 1004 | 1218 | 3105 | 1555 | |
| C*1203 | 1588 | 3638 | 1551 | 2763 | 3129 | 1279 | 1382 | 3905 | 1674 | |
| C*1402 | 2077 | 5509 | 2009 | 3508 | 3440 | 1425 | 1640 | 3862 | 2073 | |
| C*1502 | 1408 | 3095 | 1375 | 2598 | 2614 | 1086 | 1183 | 3487 | 1481 | |
| C*1601 | 1724 | 4233 | 1739 | 3118 | 3473 | 1363 | 1526 | 3727 | 1967 | |
| C*1701 | 1857 | 4062 | 2103 | 3552 | 3823 | 1368 | 1625 | 3703 | 1991 | |
| C*1802 | 2275 | 4848 | 2292 | 3959 | 3990 | 1632 | 2054 | 4296 | 2353 |
| Immunogen HLA-E Alleles | Groups | Reactivity of Monoclonal Antibodies with HLA-I Alleles | Number of mAbs | |||||
|---|---|---|---|---|---|---|---|---|
| HLA-Ia | HLA-Ib | |||||||
| HLA-A | HLA-B | HLA-C | HLA-E | HLA-F | HLA-G | |||
| HLA-ER/G107 | A | (–) | (–) | (–) | (+) | (–) | (–) | 25 |
| B | (–) | (–) | (–) | (+) | (+) | (+) | 8 | |
| C | A*1101+ | (+) | (+) | (+) | (–) | (–) | 83 | |
| D | (+) | (+) | (+) | (+) | (–) | (–) | 54 | |
| E | (+) | (+) | (+) | (+) | (–) | (+) | 18 | |
| F | (+) | (+) | (+) | (+) | (+) | (+) | 6 | |
| HLA-ER107 | A1 | (–) | B*4006+ | (–) | (+) | (–) | (–) | 1 |
| B2 | (–) | (+) | (+) | (+) | (–) | (–) | 30 | |
| C3 | A*1101+ | (+) | (+) | (+) | (+) | (–) | 1 | |
| D4 | A*1101+ | (+) | (+) | (+) | (+) | (+) | 1 | |
| HLA-EG107 | E5 | (–) | (–) | (–) | (+) | (+) | (–) | 1 |
| F6 | (–) | (–) | (–) | (+) | (–) | (+) | 1 | |
| G7 | (–) | (+) | (–) | (+) | (–) | (–) | 4 | |
| H8 | (+) | (+) | (+) | (+) | (+) | (–) | 10 | |
| Mean Fluorescent Intensity (MFI)* withTFL-006 | |||||||
|---|---|---|---|---|---|---|---|
| HLA-I | LABSCreen (LS) | LC | HLA-I | LABSCreen (LS) | LC | ||
| Lot # 11 | Lot # 10 | 3005613 | Lot # 11 | Lot # 10 | 3005613 | ||
| 20 mg/mL | 20 mg/mL | 20 mg/mL | 20 mg/mL | ||||
| A*01:01 | 852 | 933 | 0 | B*07:02 | 670 | 862 | 0 |
| A*02:01 | 542 | 339 | 0 | B*08:01 | 1204 | 1226 | 0 |
| A*02:03 | 156 | 1018 | 0 | B*13:01 | 675 | n/a | |
| A*02:06 | 616 | n/a | B*13:02 | 4923 | 2514 | 0 | |
| A*03:01 | 210 | 193 | 0 | B*14:01 | 1983 | 7805 | 0 |
| A*11:01 | 3900 | 4782 | 0 | B*14:02 | 197 | 1831 | 0 |
| A*11:02 | 321 | 537 | 0 | B*15:01 | 2541 | 335 | 0 |
| A*23:01 | 125 | 133 | 0 | B*15:02 | 809 | 1935 | 0 |
| A*24:02 | 580 | 716 | 0 | B*15:03 | 1146 | 1822 | 0 |
| A*24:03 | 1042 | 2516 | 0 | B*15:10 | 579 | n/a | |
| A*25:01 | 349 | 194 | 0 | B*15:11 | 1658 | n/a | |
| A*26:01 | 2273 | 2221 | 0 | B*15:12 | 1875 | 770 | 0 |
| A*29:01 | 1667 | 1017 | 0 | B*15:13 | 2437 | 3135 | 0 |
| A*29:02 | 715 | 778 | 0 | B*15:16 | 882 | 3076 | 0 |
| A*30:01 | 1243 | 1496 | 0 | B*18:01 | 2891 | 3096 | 0 |
| A*30:02 | 728 | n/a | B*27:05 | 3252 | 634 | 0 | |
| A*31:01 | 687 | 396 | 0 | B*27:08 | 2095 | 1659 | 0 |
| A*32:01 | 341 | 515 | 0 | B*35:01 | 517 | 6128 | 0 |
| A*33:01 | 574 | 1038 | 0 | B*37:01 | 4352 | 2650 | 0 |
| A*33:03 | 1039 | 554 | 0 | B*38:01 | 2425 | 2521 | 0 |
| A*34:01 | 3002 | n/a | B*39:01 | 4447 | 704 | 0 | |
| A*34:02 | 1165 | 1535 | 0 | B*40:01 | 330 | 3429 | 0 |
| A*36:01 | 941 | 1353 | 0 | B*40:02 | 3354 | 2697 | 0 |
| A*43:01 | 2768 | 2479 | 0 | B*40:06 | 1869 | n/a | |
| A*66:01 | 2145 | 1886 | 0 | B*41:01 | 1364 | 3739 | 0 |
| A*66:02 | 1327 | 1454 | 0 | B*42:01 | 957 | 347 | 0 |
| A*68:01 | 454 | 713 | 0 | B*44:02 | 892 | 3650 | 0 |
| A*68:02 | 578 | 1185 | 0 | B*44:03 | 2516 | 1829 | 0 |
| A*69:01 | 1786 | 3128 | 0 | B*45:01 | 2568 | 1736 | 0 |
| A*74:01 | 603 | 652 | 0 | B*46:01 | 1415 | 3572 | 0 |
| A*80:01 | 3758 | 3132 | 0 | B*47:01 | 1859 | 2152 | 0 |
| B*48:01 | 2418 | 3262 | 0 | ||||
| Cw*01:02 | 2820 | 4066 | 0 | B*49:01 | 3042 | 1554 | 0 |
| Cw*02:02 | 3403 | 7446 | 0 | B*50:01 | 4246 | 1799 | 0 |
| Cw*03:02 | 6614 | n/a | B*51:01 | 2413 | 2461 | 0 | |
| Cw*03:03 | 3071 | 2458 | 0 | B*51:02 | 696 | n/a | |
| Cw*03:04 | 8167 | 4504 | 0 | B*52:01 | 1745 | 2146 | 0 |
| Cw*04:01 | 4182 | 3337 | 0 | B*53:01 | 695 | 5442 | 0 |
| Cw*05:01 | 2567 | 9124 | 0 | B*54:01 | 2427 | 1662 | 0 |
| Cw*06:02 | 5794 | 5644 | 0 | B*55:01 | 4867 | 2519 | 0 |
| Cw*07:02 | 1689 | 8702 | 0 | B*56:01 | 3084 | 3662 | 0 |
| Cw*08:01 | 4479 | 6090 | 0 | B*57:01 | 141 | 2089 | 0 |
| Cw*12:03 | 6731 | n/a | B*57:03 | 1878 | n/a | ||
| Cw*14:02 | 3627 | 3937 | 0 | B*58:01 | 3190 | 5268 | 0 |
| Cw*15:02 | 4651 | 4465 | 0 | B*59:01 | 1278 | 3553 | 0 |
| Cw*16:01 | 2706 | 4648 | 0 | B*67:01 | 3520 | 406 | 0 |
| Cw*17:01 | 5760 | 8296 | 0 | B*73:01 | 3565 | 1423 | 0 |
| Cw*18:02 | 829 | n/a | 0 | B*78:01 | 5271 | 2996 | 0 |
| B*81:01 | 2446 | 1525 | 0 | ||||
| B*82:02 | 1959 | n/a | |||||
| Octagam-IVIg | CD4+ T-Lymphoblasts | CD8+ T-Lymphoblasts | |||||||||||||||
|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|
| Blastogenesis | Proliferation | Blastogenesis | Proliferation | ||||||||||||||
| M1/2 | M3-5 | M1/2 | M3-5 | ||||||||||||||
| Mean | SD | p2 | Mean | SD | Mean | SD | p2 | Mean | SD | p2 | Mean | SD | Mean | SD | p2 | ||
| no PHA | 46 | 8 | 73 | 17 | 1 | 1 | 53 | 13 | 80 | 20 | 4 | 7 | |||||
| with PHA only | 1685 | 89 | <0.0001 | 338 | 15 | 1486 | 95 | <0.0001 | 1951 | 171 | <0.0001 | 329 | 15 | 1751 | 193 | <0.0001 | |
| PHA + IVIg (1/10) | 945 | 87 | 0.0005 | 405 | 5 | 675 | 68 | 0.0003 | 1134 | 13 | 0.001 | 431 | 115 | 893 | 9 | 0.002 | |
| PHA + IVIg (1/20) | 1365 | 100 | 0.019 | 432 | 23 | 1105 | 71 | 0.005 | 1717 | 198 | NS | 380 | 17 | 1596 | 46 | 0.005 | |
| PHA + IVIg (1/40) | 1796 | 81 | NS | 464 | 31 | 1540 | 43 | NS | 2280 | 127 | NS | 419 | 61 | 2036 | 137 | NS | |
| TFL-006 (ascites) | Concentration | ||||||||||||||||
| No PHA | 39 | 6 | 53 | 13 | 1 | 1 | 70 | 3 | 53 | 13 | 1 | 1 | |||||
| With PHA | 1977 | 62 | <0.0001 | 320 | 3 | 1850 | 64 | <0.0001 | 403 | 31 | 0.003 | 320 | 3 | 1850 | 64 | <0.0001 | |
| (1/7) | 127.0 µg/mL | 934 | 69 | <0.0001 | 506 | 30 | 576 | 51 | <0.0001 | 176 | 75 | 0.02 | 506 | 30 | 576 | 51 | <0.0001 |
| (1/70) | 12.67 µg/mL | 1259 | 105 | <0.0001 | 470 | 22 | 954 | 118 | 0.0003 | 191 | 72 | 0.02 | 470 | 22 | 954 | 118 | 0.0003 |
| (1/140) | 6.336 µg/mL | 1437 | 30 | 0.0002 | 485 | 27 | 1162 | 57 | 0.0002 | 207 | 67 | 0.02 | 485 | 27 | 1162 | 57 | 0.0002 |
| (1/280) | 3.168 µg/mL | 1660 | 110 | 0.01 | 404 | 20 | 1450 | 120 | 0.007 | 248 | 16 | 0.005 | 404 | 20 | 1450 | 120 | 0.007 |
| (1/560) | 1.584 µg/mL | 1851 | 72 | NS. | 390 | 21 | 1669 | 75 | 0.03 | 477 | 52 | NS | 390 | 21 | 1669 | 75 | 0.03 |
| TFL-007 (ascites) | |||||||||||||||||
| No PHA | 34 | 5 | 40 | 6 | 3 | 3 | 34 | 5 | 40 | 6 | 3 | 3 | |||||
| With PHA | 2198 | 224 | <0.0001 | 333 | 15 | 2021 | 221 | <0.0001 | 2198 | 224 | <0.0001 | 333 | 15 | 2021 | 221 | <0.0001 | |
| (1/7) | 89.57 µg/mL | 1050 | 67 | 0.001 | 462 | 27 | 714 | 60 | 0.0006 | 1050 | 67 | 0.001 | 462 | 27 | 714 | 60 | 0.0006 |
| (1/70) | 8.957 µg/mL | 1372 | 106 | 0.0050 | 473 | 35 | 1072 | 38 | 0.0020 | 1372 | 106 | 0.0050 | 473 | 35 | 1072 | 38 | 0.0020 |
| (1/140) | 4.937 µg/mL | 1789 | 132 | 0.05 | 521 | 71 | 1471 | 142 | 0.02 | 1789 | 132 | 0.05 | 521 | 71 | 1471 | 142 | 0.02 |
| TFL-037 & TFL-033 | Failed to Suppres Blastogenesis or Proliferation | ||||||||||||||||
| HLA-I Reactivity of IVIg (Octagam, Mexico) | HLA-A | HLA-B | HLA-C | HLA-E | HLA-F | HLA-G | |||||||||||
| 30 | 47 | 16 | + | + | + | ||||||||||||
| TFL-006 | 31 | 48 | 16 | + | + | + | |||||||||||
| TFL-007 | 26 | 44 | 16 | + | + | + | |||||||||||
| TFL-037 | 11 | 37 | 14 | + | 0 | 0 | |||||||||||
| TFL-033 | 0 | 0 | 0 | + | 0 | 0 | |||||||||||
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Ravindranath, M.H.; El Hilali, F.; Amato-Menker, C.J.; El Hilali, H.; Selvan, S.R.; Filippone, E.J. Role of HLA-I Structural Variants and the Polyreactive Antibodies They Generate in Immune Homeostasis. Antibodies 2022, 11, 58. https://doi.org/10.3390/antib11030058
Ravindranath MH, El Hilali F, Amato-Menker CJ, El Hilali H, Selvan SR, Filippone EJ. Role of HLA-I Structural Variants and the Polyreactive Antibodies They Generate in Immune Homeostasis. Antibodies. 2022; 11(3):58. https://doi.org/10.3390/antib11030058
Chicago/Turabian StyleRavindranath, Mepur H., Fatiha El Hilali, Carly J. Amato-Menker, Hajar El Hilali, Senthamil R. Selvan, and Edward J. Filippone. 2022. "Role of HLA-I Structural Variants and the Polyreactive Antibodies They Generate in Immune Homeostasis" Antibodies 11, no. 3: 58. https://doi.org/10.3390/antib11030058
APA StyleRavindranath, M. H., El Hilali, F., Amato-Menker, C. J., El Hilali, H., Selvan, S. R., & Filippone, E. J. (2022). Role of HLA-I Structural Variants and the Polyreactive Antibodies They Generate in Immune Homeostasis. Antibodies, 11(3), 58. https://doi.org/10.3390/antib11030058