
Toward a Global Classification of Coastal Anthromes


Abstract
:1. Introduction
2. Coastal Biomes Are Anthromes
2.1. Natural Ecosystems Embedded in Human Systems
2.2. Coastal Anthromes Are Mosaics
3. Toward Classifying Coastal Anthromes
3.1. Integrating Existing Classifications and Data Sets
3.1.1. Terrestrial Anthromes
3.1.2. Inshore and Nearshore Coastal Anthromes
3.2. Identifying and Distinguishing Coastal Anthromes
- that “anthropogenic biomes will differ substantially in terms of basic ecosystem processes…and biodiversity…when measured across each biome in the field, and that these differences will be at least as great as those between the conventional biomes when observed using equivalent methods at the same spatial scale”;
- that “these differences will be driven by differences in population density and land use between the biomes…”; and
- that “the degree to which anthropogenic biomes explain global patterns of ecosystem processes and biodiversity will increase over time, in tandem with anticipated future increases in human influence on ecosystems” ([18], p. 445).
4. Challenges of Historical Legacy in Coastal Novel Ecosystems
4.1. Of Footprint and Function
4.2. Three Anthropogenic Pathways to Novel Ecosystems
4.3. Coupled Human–Natural Systems
4.3.1. The “Wild” Monoculture of Inshore American Lobster
4.3.2. Urbanised Deltas
4.3.3. Beach Nourishment and Developed Coastlines
4.4. Geoengineered Coasts
4.5. Challenges Make Opportunities
5. Conclusions
Acknowledgments
Conflicts of Interest
References
- Gillis, J.R. The Human Shore: Seacoasts in History; University of Chicago Press: Chicago, IL, USA, 2012. [Google Scholar]
- Vitousek, P.M.; Mooney, H.A.; Lubchenco, J.; Melillo, J.M. Human domination of Earth’s ecosystems. Science 1997, 277, 494–499. [Google Scholar] [CrossRef]
- Halpern, B.S.; Walbridge, S.; Selkoe, K.A.; Kappel, C.V.; Micheli, F.; D’Agrosa, C.; Bruno, J.F.; Casey, K.S.; Ebert, C.; Fox, H.E.; et al. A global map of human impact on marine ecosystems. Science 2008, 319, 948–952. [Google Scholar] [CrossRef] [PubMed]
- Halpern, B.S.; Frazier, M.; Potapenko, J.; Casey, K.S.; Koenig, K.; Longo, C.; Lowndes, J.S.; Rockwood, R.C.; Selig, E.R.; Selkoe, K.A.; et al. Spatial and temporal changes in cumulative human impacts on the world’s ocean. Nature Commun. 2015, 6, 7615. [Google Scholar] [CrossRef] [PubMed]
- Wong, P.P.; Losada, I.J.; Gattuso, J.-P.; Hinkel, J.; Khattabi, A.; McInnes, K.L.; Saito, Y.; Sallenger, A. Coastal systems and low-lying areas. In Climate Change 2014: Impacts, Adaptation, and Vulnerability. Part A: Global and Sectoral Aspects. Contribution of Working Group II to the Fifth Assessment Report of the Intergovernmental Panel on Climate Change; Field, C.B., Barros, V.R., Dokken, D.J., Mach, K.J., Mastrandrea, M.D., Bilir, T.E., Chatterjee, M., Ebi, K.L., Estrada, Y.O., Genova, R.C., et al., Eds.; Cambridge University Press: Cambridge, UK; New York, NY, USA, 2014; pp. 361–409. [Google Scholar]
- McCauley, D.J.; Pinsky, M.L.; Palumbi, S.R.; Estes, J.A.; Joyce, F.H.; Warner, R.R. Marine defaunation: Animal loss in the global ocean. Science 2015, 347, 247. [Google Scholar] [CrossRef] [PubMed]
- WWF International. The Living Blue Planet Report 2015. Available online: http://www.worldwildlife.org/publications/living-blue-planet-report-2015 (accessed on 1 October 2016).
- Erlandson, J.M. The archaeology of aquatic adaptations: Paradigms for a new millennium. J. Archaeol. Res. 2001, 9, 287–350. [Google Scholar] [CrossRef]
- Erlandson, J.M.; Rick, T.C. Archeology meets marine historical ecology: the antiquity of maritime cultures and human impacts on marine fisheries and ecosystems. Annu. Rev. Mar. Sci. 2010, 2, 231–251. [Google Scholar] [CrossRef] [PubMed]
- Marean, C.W.; Bar-Matthews, M.; Bernatchez, J.; Fisher, E.; Goldberg, P.; Herries, A.I.; Jacobs, Z.; Jerardino, A.; Karkanas, P.; Minichillo, T.; et al. Early human use of marine resources and pigment in South Africa during the Middle Pleistocene. Nature 2007, 449, 905–908. [Google Scholar] [CrossRef] [PubMed]
- Macaulay, V.; Hill, C.; Achilli, A.; Rengo, C.; Clarke, D.; Meehan, W.; Blackburn, J.; Semino, O.; Scozzari, R.; Cruciani, F.; et al. Single, rapid coastal settlement of Asia revealed by analysis of complete mitochondrial genomes. Science 2005, 308, 1034–1036. [Google Scholar] [CrossRef] [PubMed]
- O’Connor, S.; Ono, R.; Clarkson, C. Pelagic fishing at 42,000 years before the present and the maritime skills of modern humans. Science 2011, 334, 1117–1121. [Google Scholar] [CrossRef] [PubMed]
- Fujita, M.; Yamasaki, S.; Katagiri, C.; Oshiro, I.; Sano, K.; Kurozumi, T.; Sugawara, H.; Kunikita, D.; Matsuzaki, H.; Kano, A.; et al. Advanced maritime adaptation in the western Pacific coastal region extends back to 35,000–30,000 years before present. Proc. Natl. Acad. Sci. USA 2016, 113, 11184–11189. [Google Scholar] [CrossRef] [PubMed]
- Pedersen, M.W.; Ruter, A.; Schweger, C.; Friebe, H.; Staff, R.A.; Kjeldsen, K.K.; Mendoza, M.L.; Beaudoin, A.B.; Zutter, C.; Larsen, N.K.; et al. Postglacial viability and colonization in North America’s ice-free corridor. Nature 2016, 537, 45–49. [Google Scholar] [CrossRef] [PubMed]
- Fagan, B.M. The Long Summer: How Climate Changed Civilization; Basic Books: New York, NY, USA, 2004. [Google Scholar]
- Ziegler, M.; Simon, M.H.; Hall, I.R.; Barker, S.; Stringer, C.; Zahn, R. Development of Middle Stone Age innovation linked to rapid climate change. Nature Commun. 2013, 4, 1905. [Google Scholar] [CrossRef] [PubMed]
- O’Connell, J.F.; Allen, J. The process, biotic impact, and global implications of the human colonization of Sahul about 47,000 years ago. J. Archaeol. Sci. 2015, 56, 73–84. [Google Scholar] [CrossRef]
- Ellis, E.C.; Ramankutty, N. Putting people in the map: Anthropogenic biomes of the world. Front. Ecol. Environ. 2008, 6, 439–447. [Google Scholar] [CrossRef]
- Ellis, E.C. Anthropogenic transformation of the terrestrial biosphere. Philos. T. Roy. Soc. A 2011, 369, 1010–1035. [Google Scholar] [CrossRef] [PubMed]
- Hooke, R.L. On the efficacy of humans as geomorphic agents. GSA Today 1994, 4, 217, 224–225. [Google Scholar]
- Crutzen, P.J. Geology of mankind. Nature 2002, 415, 23. [Google Scholar] [CrossRef] [PubMed]
- Steffen, W.; Grinevald, J.; Crutzen, P.; McNeill, J. The Anthropocene: Conceptual and historical perspectives. Philos. T. Roy. Soc. A 2011, 369, 842–867. [Google Scholar] [CrossRef] [PubMed]
- Halpern, B.S.; Selkoe, K.A.; Micheli, F.; Kappel, C.V. Evaluating and ranking the vulnerability of global marine ecosystems to anthropogenic threats. Conserv. Biol. 2007, 21, 1301–1315. [Google Scholar] [CrossRef] [PubMed]
- Nordstrom, K.F. Beaches and Dunes of Developed Coasts; Cambridge University Press: Cambridge, UK; New York, NY, USA, 2000. [Google Scholar]
- Syvitski, J.P.; Vörösmarty, C.J.; Kettner, A.J.; Green, P. Impact of humans on the flux of terrestrial sediment to the global coastal ocean. Science 2005, 308, 376–380. [Google Scholar] [CrossRef] [PubMed]
- Halpern, B.S.; Ebert, C.M.; Kappel, C.V.; Madin, E.M.P.; Micheli, F.; Perry, M.; Selkoe, K.A.; Walbridge, S. Global priority areas for incorporating land–sea connections in marine conservation. Conserv. Lett. 2009, 2, 189–196. [Google Scholar] [CrossRef]
- Lazarus, E.D.; Ellis, M.A.; Murray, A.B.; Hall, D.M. An evolving research agenda for human–coastal systems. Geomorphology 2016, 256, 81–90. [Google Scholar] [CrossRef]
- Jackson, J.B.C.; Kirby, M.X.; Berger, W.H.; Bjorndal, K.A.; Botsford, L.W.; Bourque, B.J.; Bradbury, R.H.; Cooke, R.; Erlandson, J.; Estes, J.A.; et al. Historical overfishing and the recent collapse of coastal ecosystems. Science 2001, 293, 629–637. [Google Scholar] [CrossRef] [PubMed]
- Pauly, D.; Christensen, V.; Guénette, S.; Pitcher, T.J.; Sumaila, U.R.; Walters, C.J.; Watson, R.; Zeller, D. Towards sustainability in world fisheries. Nature 2002, 418, 689–695. [Google Scholar] [CrossRef] [PubMed]
- Pusceddu, A.; Bianchelli, S.; Martín, J.; Puig, P.; Palanques, A.; Masqué, P.; Danovaro, R. Chronic and intensive bottom trawling impairs deep-sea biodiversity and ecosystem functioning. Proc. Natl. Acad. Sci. USA 2014, 111, 8861–8866. [Google Scholar] [CrossRef] [PubMed]
- Adger, W.N. Social and ecological resilience: are they related? Prog. Hum. Geog. 2000, 24, 347–364. [Google Scholar] [CrossRef]
- Steinberg, P.E. Navigating to multiple horizons: Toward a geography of ocean-space. Prof. Geogr. 1999, 51, 366–375. [Google Scholar] [CrossRef]
- Steinberg, P.E. The Social Construction of the Ocean; Cambridge University Press: Cambridge, UK, 2001. [Google Scholar]
- Halpern, B.S.; Fujita, R. Assumptions, challenges, and future directions in cumulative impact analysis. Ecosphere 2013, 4, 1–11. [Google Scholar] [CrossRef]
- Haff, P.K. Hillslopes, rivers, plows, and trucks: Mass transport on Earth’s surface by natural and technological processes. Earth Surf. Proc. Land. 2010, 35, 1157–1166. [Google Scholar] [CrossRef]
- Jackson, J.B.C. Ecological extinction and evolution in the brave new ocean. Proc. Natl. Acad. Sci. USA 2008, 105 (Supp. 1), 11458–11465. [Google Scholar] [CrossRef] [PubMed]
- Jackson, J.B.C. The future of the oceans past. Philos. T. Roy. Soc. B 2010, 365, 3765–3778. [Google Scholar] [CrossRef] [PubMed]
- Doney, S.C. The growing human footprint on coastal and open-ocean biogeochemistry. Science 2010, 328, 1512–1516. [Google Scholar] [CrossRef] [PubMed]
- Vörösmarty, C.J.; McIntyre, P.B.; Gessner, M.O.; Dudgeon, D.; Prusevich, A.; Green, P.; Glidden, S.; Bunn, S.E.; Sullivan, C.A.; Liermann, C.R.; et al. Global threats to human water security and river biodiversity. Nature 2010, 467, 555–561. [Google Scholar] [CrossRef] [PubMed]
- Diaz, R.J.; Rosenberg, R. Spreading dead zones and consequences for marine ecosystems. Science 2008, 321, 926–929. [Google Scholar] [CrossRef] [PubMed]
- Jenny, J.-P.; Francus, P.; Normandeau, A.; Lapointe, F.; Perga, M.-E.; Ojala, A.; Schimmeimann, A.; Zolitschka, B. Global spread of hypoxia in freshwater ecosystems during the last three centuries is caused by rising local human pressure. Glob. Change Biol. 2016, 22, 1481–1489. [Google Scholar] [CrossRef] [PubMed]
- Rabalais, N.N.; Cai, W.-J.; Carstensen, J.; Conley, D.J.; Fry, B.; Hu, X.; Quinones-Rivera, Z.; Rosenberg, R.; Slomp, C.P.; Turner, R.E.; et al. Eutrophication-driven deoxygenation in the coastal ocean. Oceanography 2014, 27, 172–183. [Google Scholar] [CrossRef]
- Sawyer, A.H.; David, C.H.; Famiglietti, J.S. Continental patterns of submarine groundwater discharge reveal coastal vulnerabilities. Science 2016, 353, 705–707. [Google Scholar] [CrossRef] [PubMed]
- Pauly, D.; Christensen, V. Primary production required to sustain global fisheries. Nature 1995, 374, 255–257. [Google Scholar] [CrossRef]
- Watling, L.; Norse, E.A. Disturbance of the seabed by mobile fishing gear: A comparison to forest clearcutting. Conserv. Biol. 1998, 12, 1180–1197. [Google Scholar] [CrossRef]
- Pelling, M.; Blackburn, S. Megacities and the Coast: Risk, Resilience and Transformation; Routledge: London, UK; New York, NY, USA, 2014. [Google Scholar]
- Valiela, I.; Bowen, J.L.; York, J.K. Mangrove forests: One of the world’s threatened major tropical environments. BioScience 2001, 51, 807–815. [Google Scholar] [CrossRef]
- Polidoro, B.A.; Carpenter, K.E.; Collins, L.; Duke, N.C.; Ellison, A.M.; Ellison, J.C.; Farnsworth, E.J.; Fernando, E.S.; Kathiresan, K.; Koedam, N.E.; et al. The loss of species: Mangrove extinction risk and geographic areas of global concern. PLoS ONE 2010, 5, e10095. [Google Scholar] [CrossRef] [PubMed]
- Giri, C.; Ochieng, E.; Tieszen, L.L.; Zhu, Z.; Singh, A.; Loveland, T.; Masek, J.; Duke, N. Status and distribution of mangrove forests of the world using earth observation satellite data. Global Ecol. Biogeogr. 2011, 20, 154–159. [Google Scholar] [CrossRef]
- Orth, R.J.; Carruthers, T.J.; Dennison, W.C.; Duarte, C.M.; Fourqurean, J.W.; Heck, K.L.; Hughes, A.R.; Kendrick, G.A.; Kenworthy, W.J.; Olyarnik, S.; et al. A global crisis for seagrass ecosystems. BioScience 2006, 56, 987–996. [Google Scholar] [CrossRef]
- Waycott, M.; Duarte, C.M.; Carruthers, T.J.; Orth, R.J.; Dennison, W.C.; Olyarnik, S.; Calladine, A.; Fourqurean, J.W.; Heck, K.L.; Hughes, A.R.; et al. Accelerating loss of seagrasses across the globe threatens coastal ecosystems. Proc. Natl. Acad. Sci. USA 2009, 106, 12377–12381. [Google Scholar] [CrossRef] [PubMed]
- Gedan, K.B.; Silliman, B.R.; Bertness, M.D. Centuries of human-driven change in salt marsh ecosystems. Annu. Rev. Mar. Sci. 2009, 1, 117–141. [Google Scholar] [CrossRef] [PubMed]
- Kirwan, M.L.; Megonigal, J.P. Tidal wetland stability in the face of human impacts and sea-level rise. Nature 2013, 504, 53–60. [Google Scholar] [CrossRef] [PubMed]
- Lotze, H.K.; Lenihan, H.S.; Bourque, B.J.; Bradbury, R.H.; Cooke, R.G.; Kay, M.C.; Kidwell, S.M.; Kirby, M.X.; Peterson, C.H.; Jackson, J.B.C. Depletion, degradation, and recovery potential of estuaries and coastal seas. Science 2006, 312, 1806–1809. [Google Scholar] [CrossRef] [PubMed]
- Kirby, M.X. Fishing down the coast: historical expansion and collapse of oyster fisheries along continental margins. Proc. Natl. Acad. Sci. USA 2004, 101, 13096–13099. [Google Scholar] [CrossRef] [PubMed]
- Pandolfi, J.M.; Bradbury, R.H.; Sala, E.; Hughes, T.P.; Bjorndal, K.A.; Cooke, R.G.; McArdle, D.; McClenachan, L.; Newman, M.J.; Paredes, G.; et al. Global trajectories of the long-term decline of coral reef ecosystems. Science 2003, 301, 955–958. [Google Scholar] [CrossRef] [PubMed]
- Bellwood, D.R.; Hughes, T.P.; Folke, C.; Nyström, M. Confronting the coral reef crisis. Nature 2004, 429, 827–833. [Google Scholar] [CrossRef] [PubMed]
- Steneck, R.S.; Graham, M.H.; Bourque, B.J.; Corbett, D.; Erlandson, J.M.; Estes, J.A.; Tegner, M.J. Kelp forest ecosystems: Biodiversity, stability, resilience and future. Environ. Conserv. 2002, 29, 436–459. [Google Scholar] [CrossRef]
- Nordstrom, K.F. Beaches and dunes of human-altered coasts. Prog. Phys. Geog. 1994, 18, 497–516. [Google Scholar] [CrossRef]
- Jackson, J.B.C. What was natural in the coastal oceans? Proc. Natl. Acad. Sci. USA 2001, 98, 5411–5418. [Google Scholar] [CrossRef] [PubMed]
- Goodall, D.W. (Ed.) Ecosystems of the World; Elsevier: Amsterdam, The Netherlands, 1974–; Volume 38.
- Olson, D.M.; Dinerstein, E.; Wikramanayake, E.D.; Burgess, N.D.; Powell, G.V.; Underwood, E.C.; D’amico, J.A.; Itoua, I.; Strand, H.E.; Morrison, J.C.; et al. Terrestrial ecoregions of the world: A new map of life on Earth. BioScience 2001, 51, 933–938. [Google Scholar] [CrossRef]
- Smith, H.D. The development and management of the world ocean. Ocean Coast. Manage. 1994, 24, 3–16. [Google Scholar] [CrossRef]
- Smith, H.D. The industrialisation of the world ocean. Ocean Coast. Manage. 2000, 43, 11–28. [Google Scholar] [CrossRef]
- Douvere, F. The importance of marine spatial planning in advancing ecosystem-based sea use management. Mar. Policy 2008, 32, 762–771. [Google Scholar] [CrossRef]
- Martin, K.S.; Hall-Arber, M. The missing layer: Geo-technologies, communities, and implications for marine spatial planning. Mar. Policy 2008, 32, 779–786. [Google Scholar] [CrossRef]
- Foley, M.M.; Halpern, B.S.; Micheli, F.; Armsby, M.H.; Caldwell, M.R.; Crain, C.M.; Prahler, E.; Rohr, N.; Sivas, D.; Beck, M.W.; et al. Guiding ecological principles for marine spatial planning. Mar. Policy 2010, 34, 955–966. [Google Scholar] [CrossRef]
- Paine, L. The Sea and Civilization: A Maritime History of the World; Atlantic Books Ltd.: London, UK, 2014. [Google Scholar]
- McGranahan, G.; Balk, D.; Anderson, B. The rising tide: Assessing the risks of climate change and human settlements in low elevation coastal zones. Environ. Urban. 2007, 19, 17–37. [Google Scholar] [CrossRef]
- Olson, D.M.; Dinerstein, E. The Global 200: A representation approach to conserving the Earth’s most biologically valuable ecoregions. Conserv. Biol. 1998, 12, 502–515. [Google Scholar] [CrossRef]
- Doody, J.P. Coastal Conservation and Management; Springer: London, UK, 2001. [Google Scholar]
- Bakker, J.P. The impact of grazing on plant communities, plant populations and soil conditions on salt marshes. Vegetatio 1985, 62, 391–398. [Google Scholar] [CrossRef]
- Kiehl, K.; Eischeid, I.; Gettner, S.; Walter, J. Impact of different sheep grazing intensities on salt marsh vegetation in northern Germany. J. Veg. Sci. 1996, 7, 99–106. [Google Scholar] [CrossRef]
- Blanco-Libreros, J.F.; Estrada-Urrea, E.A. Mangroves on the edge: Anthrome-dependent fragmentation influences ecological condition (Turbo, Colombia, Southern Caribbean). Diversity 2015, 7, 206–228. [Google Scholar] [CrossRef]
- Vafeidis, A.T.; Nicholls, R.J.; McFadden, L.; Tol, R.S.; Hinkel, J.; Spencer, T.; Grashoff, P.S.; Boot, G.; Klein, R.J. A new global coastal database for impact and vulnerability analysis to sea-level rise. J. Coastal Res. 2008, 24, 917–924. [Google Scholar] [CrossRef]
- Folke, C.; Kautsky, N. Aquaculture with its environment: Prospects for sustainability. Ocean Coast. Manage. 1992, 17, 5–24. [Google Scholar] [CrossRef]
- Newkirk, G. Sustainable coastal production systems: A model for integrating aquaculture and fisheries under community management. Ocean Coast. Manage. 1996, 32, 69–83. [Google Scholar] [CrossRef]
- Brzeski, V.; Newkirk, G. Integrated coastal food production systems—A review of current literature. Ocean Coast. Manage. 1997, 34, 55–71. [Google Scholar] [CrossRef]
- Troell, M.; Kautsky, N.; Folke, C. Applicability of integrated coastal aquaculture systems. Ocean Coast. Manage. 1999, 42, 63–70. [Google Scholar]
- Chopin, T.; Buschmann, A.H.; Halling, C.; Troell, M.; Kautsky, N.; Neori, A.; Kraemer, G.P.; Zertuche-González, J.A.; Yarish, C.; Neefus, C. Integrating seaweeds into marine aquaculture systems: A key toward sustainability. J. Phycol. 2001, 37, 975–986. [Google Scholar] [CrossRef]
- Warnock, N.; Page, G.W.; Ruhlen, T.D.; Nur, N.; Takekawa, J.Y.; Hanson, J.T. Management and conservation of San Francisco Bay salt ponds: Effects of pond salinity, area, tide, and season on Pacific Flyway waterbirds. Waterbirds 2002, 25, 79–92. [Google Scholar]
- Brown, B.; Wilson, W.H. The role of commercial digging of mudflats as an agent for change of infaunal intertidal populations. J. Exp. Mar. Biol. Ecol. 1997, 218, 49–61. [Google Scholar] [CrossRef]
- Mouritsen, O.G.; Mouritsen, J.D.; Johansen, M. Seaweeds: Edible, Available, and Sustainable; University of Chicago Press: Chicago, IL, USA, 2013. [Google Scholar]
- Dalton, R. Aquaculture: Fishing for trouble. Nature 2004, 431, 502–504. [Google Scholar] [CrossRef] [PubMed]
- Stergiou, K.I.; Tsikliras, A.C.; Pauly, D. Farming up Mediterranean food webs. Conserv. Biol. 2009, 23, 230–232. [Google Scholar] [CrossRef] [PubMed]
- Steneck, R.S.; Hughes, T.P.; Cinner, J.E.; Adger, W.N.; Arnold, S.N.; Berkes, F.; Boudreau, S.A.; Brown, K.; Folke, C.; Gunderson, L.; et al. Creation of a gilded trap by the high economic value of the Maine lobster fishery. Conserv. Biol. 2011, 25, 904–912. [Google Scholar] [CrossRef] [PubMed]
- Puig, P.; Canals, M.; Company, J.B.; Martín, J.; Amblas, D.; Lastras, G.; Palanques, A.; Calafat, A.M. Ploughing the deep sea floor. Nature 2012, 489, 286–289. [Google Scholar] [CrossRef] [PubMed]
- Petersen, J.K.; Malm, T. Offshore windmill farms: Threats to or possibilities for the marine environment. Ambio 2006, 35, 75–80. [Google Scholar] [CrossRef]
- Quigel, J.C.; Thornton, W.L. Rigs to reefs—A case history. B. Mar. Sci. 1989, 44, 799–806. [Google Scholar]
- Spalding, M.D.; Fox, H.E.; Allen, G.R.; Davidson, N.; Ferdaña, Z.A.; Finlayson, M.A.X.; Halpern, B.S.; Jorge, M.A.; Lombana, A.L.; Lourie, S.A.; et al. Marine ecoregions of the world: A bioregionalization of coastal and shelf areas. BioScience 2007, 57, 573–583. [Google Scholar] [CrossRef]
- Halpern, B.S.; McLeod, K.L.; Rosenberg, A.A.; Crowder, L.B. Managing for cumulative impacts in ecosystem-based management through ocean zoning. Ocean Coast. Manage. 2008, 51, 203–211. [Google Scholar] [CrossRef]
- Ellis, E.C.; Klein Goldewijk, K.; Siebert, S.; Lightman, D.; Ramankutty, N. Anthropogenic transformation of the biomes, 1700 to 2000. Glob. Ecol. Biogeogr. 2010, 19, 589–606. [Google Scholar] [CrossRef]
- Ellis, E.C.; Kaplan, J.O.; Fuller, D.Q.; Vavrus, S.; Goldewijk, K.K.; Verburg, P.H. Used planet: A global history. Proc. Natl. Acad. Sci. USA 2013, 110, 7978–7985. [Google Scholar] [CrossRef] [PubMed]
- International Comprehensive Ocean-Atmosphere Data Set (ICOADS). US Maury collection (ship’s names and voyages, 1784–1863). Available online: http://icoads.noaa.gov/maury.html (accessed on 1 October 2016).
- Ames, E.P. Atlantic cod stock structure in the Gulf of Maine. Fisheries 2004, 29, 10–28. [Google Scholar] [CrossRef]
- Smith, T.M.; Reynolds, R.W.; Peterson, T.C.; Lawrimore, J. Improvements to NOAA’s historical merged land-ocean surface temperature analysis (1880–2006). J. Climate 2008, 21, 2283–2296. [Google Scholar] [CrossRef]
- Pinsky, M.L.; Jensen, O.P.; Ricard, D.; Palumbi, S.R. Unexpected patterns of fisheries collapse in the world’s oceans. Proc. Natl. Acad. Sci. USA 2011, 108, 8317–8322. [Google Scholar] [CrossRef] [PubMed]
- Airoldi, L.; Beck, M.W. Loss, status and trends for coastal marine habitats of Europe. Oceanogr. Mar. Biol. 2007, 45, 345–405. [Google Scholar]
- McLean, J.H. Sublittoral ecology of kelp beds of the open coast area near Carmel, California. Biol. Bull. 1962, 122, 95–114. [Google Scholar] [CrossRef]
- Dayton, P.K.; Tegner, M.J.; Edwards, P.B.; Riser, K.L. Sliding baselines, ghosts, and reduced expectations in kelp forest communities. Ecol. Appl. 1998, 8, 309–322. [Google Scholar] [CrossRef]
- Lebel, L.; Anderies, J.M.; Campbell, B.; Folke, C.; Hatfield-Dodds, S.; Hughes, T.P.; Wilson, J. Governance and the capacity to manage resilience in regional social-ecological systems. Ecol. Soc. 2006, 11, 19. [Google Scholar] [CrossRef]
- Foster, D.; Aber, J.; Cogbill, C.; Hart, C.; Colburn, E.; D’Amato, A.; Donahue, B.; Driscoll, C.; Ellison, A.; Fahey, T.; et al. Wildlands and Woodlands: A Vision for the New England Landscape; Harvard University Press: Cambridge, MA, USA, 2010; Available online: http://harvardforest.fas.harvard.edu/other-tags/wildlands-woodlands (accessed on 1 October 2016).
- Kirwan, M.L.; Murray, A.B.; Donnelly, J.P.; Corbett, D.R. Rapid wetland expansion during European settlement and its implication for marsh survival under modern sediment delivery rates. Geology 2011, 39, 507–510. [Google Scholar] [CrossRef]
- Barbier, E.B.; Hacker, S.D.; Kennedy, C.; Koch, E.W.; Stier, A.C.; Silliman, B.R. The value of estuarine and coastal ecosystem services. Ecol. Monogr. 2011, 81, 169–193. [Google Scholar] [CrossRef]
- Loarie, S.R.; Duffy, P.B.; Hamilton, H.; Asner, G.P.; Field, C.B.; Ackerly, D.D. The velocity of climate change. Nature 2009, 462, 1052–1055. [Google Scholar] [CrossRef] [PubMed]
- Neilson, R.P. Transient ecotone response to climatic change: some conceptual and modeling approaches. Ecol. Appl. 1993, 3, 385–395. [Google Scholar] [CrossRef] [PubMed]
- Lloyd, K.M.; McQueen, A.A.M.; Lee, B.J.; Wilson, R.C.B.; Walker, S.; Wilson, J.B. Evidence on ecotone concepts from switch, environmental and anthropogenic ecotones. J. Veg. Sci. 2000, 11, 903–910. [Google Scholar] [CrossRef]
- Pinsky, M.L.; Worm, B.; Fogarty, M.J.; Sarmiento, J.L.; Levin, S.A. Marine taxa track local climate velocities. Science 2013, 341, 1239–1242. [Google Scholar] [CrossRef] [PubMed]
- Lazarus, E.D.; McGill, B.J. Pushing the pace of tree species migration. PLoS ONE 2014, 9, e105380. [Google Scholar] [CrossRef] [PubMed]
- Pinsky, M.L.; Fogarty, M. Lagged social-ecological responses to climate and range shifts in fisheries. Climatic Change 2012, 115, 883–891. [Google Scholar] [CrossRef]
- Myers, N. Environmental refugees: A growing phenomenon of the 21st century. Philos. T. Roy. Soc. B 2002, 357, 609–613. [Google Scholar] [CrossRef] [PubMed]
- Hundley, N. Water and the West: the Colorado River Compact and the Politics of Water in the American West; University of California Press: Berkeley, CA, USA, 2009. [Google Scholar]
- Lazarus, E.D. Threshold effects of hazard mitigation in coastal human-environmental systems. Earth Surf. Dynam. 2014, 2, 35–45. [Google Scholar] [CrossRef]
- Holland, J. Emergence: From Chaos to Order; Basic Books: New York, NY, USA, 1998. [Google Scholar]
- Werner, B.T.; Mcnamara, D.E. Dynamics of coupled human-landscape systems. Geomorphology 2007, 91, 393–407. [Google Scholar] [CrossRef]
- Marsh, G.P. Man and Nature, or Physical Geography as Modified by Human Action; Scribner: New York, NY, USA, 1869. [Google Scholar]
- Marsh, G.P. The Earth as Modified by Human Action; Scribner: New York, NY, USA, 1882. [Google Scholar]
- Turner, B.L.; Matson, P.A.; McCarthy, J.J.; Corell, R.W.; Christensen, L.; Eckley, N.; Hovelsrud-Broda, G.K.; Kasperson, J.X.; Kasperson, R.E.; Luers, A.; et al. Illustrating the coupled human–environment system for vulnerability analysis: three case studies. Proc. Natl. Acad. Sci. USA 2003, 100, 8080–8085. [Google Scholar] [CrossRef] [PubMed]
- Liu, J.; Dietz, T.; Carpenter, S.R.; Alberti, M.; Folke, C.; Moran, E.; Pell, A.N.; Deadman, P.; Kratz, T.; Lubchenco, J.; et al. Complexity of coupled human and natural systems. Science 2007, 317, 1513–1516. [Google Scholar] [CrossRef] [PubMed]
- Scheffer, M. Critical Transitions in Nature and Society; Princeton University Press: Princeton, NJ, USA, 2009. [Google Scholar]
- Ostrom, E. A general framework for analyzing sustainability of social-ecological systems. Science 2009, 325, 419–422. [Google Scholar] [CrossRef] [PubMed]
- Di Baldassarre, G.; Viglione, A.; Carr, G.; Kuil, L.; Salinas, J.L.; Blöschl, G. Socio-hydrology: conceptualising human-flood interactions. Hydrol. Earth Syst. Sci. 2013, 17, 3295–3303. [Google Scholar] [CrossRef]
- Wilson, J.; Yan, L.; Wilson, C. The precursors of governance in the Maine lobster fishery. Proc. Natl. Acad. Sci. USA 2007, 104, 15212–15217. [Google Scholar] [CrossRef] [PubMed]
- Jury, S.H.; Howell, H.; O’Grady, D.F.; Watson, W.H., III. Lobster trap video: in situ video surveillance of the behaviour of Homarus americanus in and around traps. Mar. Freshwater Res. 2001, 52, 1125–1132. [Google Scholar] [CrossRef]
- Hutchings, J.A. Collapse and recovery of marine fishes. Nature 2000, 406, 882–885. [Google Scholar] [CrossRef] [PubMed]
- Dugmore, A.J.; McGovern, T.H.; Vésteinsson, O.; Arneborg, J.; Streeter, R.; Keller, C. Cultural adaptation, compounding vulnerabilities and conjunctures in Norse Greenland. Proc. Natl. Acad. Sci. USA 2012, 109, 3658–3663. [Google Scholar] [CrossRef] [PubMed]
- Young, N.E.; Schweinsberg, A.D.; Briner, J.P.; Schaefer, J.M. Glacier maxima in Baffin Bay during the Medieval Warm Period coeval with Norse settlement. Sci. Adv. 2015, 1, e1500806. [Google Scholar] [CrossRef] [PubMed]
- Boonstra, W.J.; de Boer, F.W. The historical dynamics of social–ecological traps. Ambio 2014, 43, 260–274. [Google Scholar] [CrossRef] [PubMed]
- Criss, R.E.; Shock, E.L. Flood enhancement through flood control. Geology 2001, 29, 875–878. [Google Scholar] [CrossRef]
- Syvitski, J.P.; Kettner, A.J.; Overeem, I.; Hutton, E.W.; Hannon, M.T.; Brakenridge, G.R.; Day, J.; Vörösmarty, C.; Saito, Y.; Giosan, L.; et al. Sinking deltas due to human activities. Nature Geosci. 2009, 2, 681–686. [Google Scholar] [CrossRef]
- Syvitski, J.P. Deltas at risk. Sustain. Sci. 2008, 3, 23–32. [Google Scholar] [CrossRef]
- McNamara, D.E.; Keeler, A. A coupled physical and economic model of the response of coastal real estate to climate risk. Nat. Clim. Change 2013, 3, 559–562. [Google Scholar] [CrossRef]
- McNamara, D.E.; Murray, A.B.; Smith, M.D. Coastal sustainability depends on how economic and coastline responses to climate change affect each other. Geophys. Res. Lett. 2011, 38, L07401. [Google Scholar] [CrossRef]
- Ells, K.; Murray, A.B. Long-term, non-local coastline responses to local shoreline stabilization. Geophys. Res. Lett. 2012, 39, L19401. [Google Scholar] [CrossRef]
- Rogers, L.J.; Moore, L.J.; Goldstein, E.B.; Hein, C.J.; Lorenzo-Trueba, J.; Ashton, A.D. Anthropogenic controls on overwash deposition: Evidence and consequences. J. Geophys. Res. Earth 2015, 120, 2609–2624. [Google Scholar] [CrossRef]
- National Research Council (NRC). Reducing Coastal Risk on the East and Gulf Coasts; National Academies Press: Washington, DC, USA, 2014. [Google Scholar]
- Hanson, H.; Brampton, A.; Capobianco, M.; Dette, H.H.; Hamm, L.; Laustrup, C.; Lechuga, A.; Spanhoff, R. Beach nourishment projects, practices, and objectives—A European overview. Coast. Eng. 2002, 47, 81–111. [Google Scholar] [CrossRef]
- NRC. Beach Nourishment and Protection; National Academies Press: Washington, DC, USA, 1995. [Google Scholar]
- Lazarus, E.D.; McNamara, D.E.; Smith, M.D.; Gopalakrishnan, S.; Murray, A.B. Emergent behavior in a coupled economic and coastline model for beach nourishment. Nonlin. Process. Geophys. 2011, 18, 989–999. [Google Scholar] [CrossRef]
- Nordstrom, K. Beach nourishment and coastal habitats: Research needs to improve compatibility. Restor. Ecol. 2005, 13, 215–222. [Google Scholar] [CrossRef]
- Peterson, C.H.; Bishop, M.J. Assessing the environmental impacts of beach nourishment. BioScience 2005, 55, 887–896. [Google Scholar] [CrossRef]
- Peterson, C.H.; Bishop, M.J.; Johnson, G.A.; D’Anna, L.M.; Manning, L.M. Exploiting beach filling as an unaffordable experiment: benthic intertidal impacts propagating upwards to shorebirds. J. Exp. Mar. Biol. Ecol. 2006, 338, 205–221. [Google Scholar] [CrossRef]
- Speybroeck, J.; Bonte, D.; Courtens, W.; Gheskiere, T.; Grootaert, P.; Maelfait, J.P.; Mathys, M.; Provoost, S.; Sabbe, K.; Stienen, E.W.; et al. Beach nourishment: an ecologically sound coastal defence alternative? A review. Aquat. Conserv. 2006, 16, 419–435. [Google Scholar] [CrossRef]
- Grain, D.A.; Bolten, A.B.; Bjorndal, K.A. Effects of beach nourishment on sea turtles: review and research initiatives. Restor. Ecol. 1995, 3, 95–104. [Google Scholar] [CrossRef]
- Hays, G.C.; Ashworth, J.S.; Barnsley, M.J.; Broderick, A.C.; Emery, D.R.; Godley, B.J.; Henwood, A.; Jones, E.L. The importance of sand albedo for the thermal conditions on sea turtle nesting beaches. Oikos 2001, 93, 87–94. [Google Scholar] [CrossRef]
- Steinitz, M.J.; Salmon, M.; Wyneken, J. Beach renourishment and loggerhead turtle reproduction: A seven year study at Jupiter Island, Florida. J. Coastal Res. 1998, 14, 1000–1013. [Google Scholar]
- Grootjans, A.P.; Geelen, H.W.T.; Jansen, A.J.M.; Lammerts, E.J. Restoration of coastal dune slacks in the Netherlands. Hydrobiologia 2002, 478, 181–203. [Google Scholar] [CrossRef]
- Nordstrom, K.F.; Jackson, N.L.; Bruno, M.S.; de Butts, H.A. Municipal initiatives for managing dunes in coastal residential areas: a case study of Avalon, New Jersey, USA. Geomorphology 2002, 47, 137–152. [Google Scholar] [CrossRef]
- Goldstein, E.B.; Moore, L.J. Stability and bistability in a one-dimensional model of coastal foredune height. J. Geophys. Res. Earth Surf. 2016, 121, 964–977. [Google Scholar] [CrossRef]
- Walters, D.C.; Kirwan, M.L. Optimal hurricane overwash thickness for maximizing marsh resilience to sea level rise. Ecol. Evol. 2016, 6, 2948–2956. [Google Scholar]
- Schupp, C.A.; Winn, N.T.; Pearl, T.L.; Kumer, J.P.; Carruthers, T.J.; Zimmerman, C.S. Restoration of overwash processes creates piping plover (Charadrius melodus) habitat on a barrier island (Assateague Island, Maryland). Estuar. Coast. Shelf S. 2013, 116, 11–20. [Google Scholar] [CrossRef]
- Smith, M.D.; Slott, J.M.; McNamara, D.; Murray, A.B. Beach nourishment as a dynamic capital accumulation problem. J. Environ. Econ. Manag. 2009, 58, 58–71. [Google Scholar] [CrossRef]
- Gopalakrishnan, S.; Smith, M.D.; Slott, J.M.; Murray, A.B. The value of disappearing beaches: a hedonic pricing model with endogenous beach width. J. Environ. Econ. Manag. 2011, 61, 297–310. [Google Scholar] [CrossRef]
- Mileti, D. Disasters by Design: A Reassessment of Natural Hazards in the United States; Joseph Henry Press: Washington, DC, USA, 1999. [Google Scholar]
- Armstrong, S.B.; Lazarus, E.D.; Limber, P.W.; Goldstein, E.B.; Thorpe, C.; Ballinger, R.C. Indications of a positive feedback between coastal development and beach nourishment. Earth’s Future 2016, 4, 626–635. [Google Scholar] [CrossRef]
- Krippendorf, J. Ecological approach to tourism marketing. Tourism Manage. 1987, 8, 174–176. [Google Scholar] [CrossRef]
- McCool, S.F.; Moisey, R.N. (Eds.) Tourism, Recreation and Sustainability: Linking Culture and the Environment, 2nd ed.; CAB International: Oxford, UK; New York, NY, USA, 2008.
- Tao, T.C.; Wall, G. Tourism as a sustainable livelihood strategy. Tourism Manage. 2009, 30, 90–98. [Google Scholar] [CrossRef]
- Cooper, J.A.G.; McKenna, J. Boom and bust: the influence of macroscale economics on the world’s coasts. J. Coastal Res. 2009, 253, 533–538. [Google Scholar] [CrossRef]
- Adger, W.N.; Hughes, T.P.; Folke, C.; Carpenter, S.R.; Rockström, J. Social-ecological resilience to coastal disasters. Science 2005, 309, 1036–1039. [Google Scholar] [CrossRef] [PubMed]
- Watkins, D. What China has been building in the South China Sea. New York Times. 27 October 2015. Available online: http://www.nytimes.com/interactive/2015/07/30/world/asia/what-china-has-been-building-in-the-south-china-sea.html?_r=0 (accessed on 1 October 2016).
- United Nations Environment Programme (UNEP). Sand, rarer than one thinks. Available online: http://www.unep.org/pdf/UNEP_GEAS_March_2014.pdf (accessed on 1 October 2016).
- Dutch Delta Commission. Available online: http://www.deltacommissie.com/ (accessed on 1 October 2016).
- Kabat, P.; van Vierssen, W.; Veraart, J.; Vellinga, P.; Aerts, J. Climate proofing the Netherlands. Nature 2005, 438, 283–284. [Google Scholar] [CrossRef] [PubMed]
- Kabat, P.; Fresco, L.O.; Stive, M.J.F.; Veerman, C.P.; van Alphen, J.S.V.L.; Parmet, B.W.A.H.; Hazeleger, W.; Katsman, C.A. Dutch coasts in transition. Nature Geosci. 2009, 2, 450–452. [Google Scholar] [CrossRef]
- Temmerman, S.; Meire, P.; Bouma, T.J.; Herman, P.M.; Ysebaert, T.; de Vriend, H.J. Ecosystem-based coastal defence in the face of global change. Nature 2013, 504, 79–83. [Google Scholar] [CrossRef] [PubMed]
- Cheong, S.M.; Silliman, B.; Wong, P.P.; van Wesenbeeck, B.; Kim, C.K.; Guannel, G. Coastal adaptation with ecological engineering. Nature Clim. Change 2013, 3, 787–791. [Google Scholar] [CrossRef]
- Van Slobbe, E.; de Vriend, H.J.; Aarninkhof, S.; Lulofs, K.; de Vries, M.; Dircke, P. Building with nature: In search of resilient storm surge protection strategies. Nature Hazards 2013, 65, 947–966. [Google Scholar] [CrossRef]
- French, P.W. Managed realignment—the developing story of a comparatively new approach to soft engineering. Estuar. Coast. Shelf S. 2006, 67, 409–423. [Google Scholar] [CrossRef]
- Louisiana Coastal Protection and Restoration Authority. Louisiana’s Comprehensive Master Plan for a Stainable Coast; CPRA: Baton Rouge, LA, USA, 2012. Available online: http://coastal.la.gov/a-common-vision/2012-coastal-master-plan/ (accessed on 1 October 2016).
- Paola, C.; Twilley, R.R.; Edmonds, D.A.; Kim, W.; Mohrig, D.; Parker, G.; Viparelli, E.; Voller, V.R. Natural processes in delta restoration: application to the Mississippi Delta. Ann. Rev. Mar. Sci. 2011, 3, 67–91. [Google Scholar] [CrossRef] [PubMed]
- Giosan, L.; Syvitski, J.; Constantinescu, S.; Day, J. Climate change: Protect the world’s deltas. Nature 2014, 516, 31–33. [Google Scholar] [CrossRef] [PubMed]
- Stive, M.J.; de Schipper, M.A.; Luijendijk, A.P.; Aarninkhof, S.G.; van Gelder-Maas, C.; van Thiel de Vries, J.S.; de Vries, S.; Henriquez, M.; Marx, S.; Ranasinghe, R. A new alternative to saving our beaches from sea-level rise: The sand engine. J. Coastal Res. 2013, 29, 1001–1008. [Google Scholar] [CrossRef]
- Brown, J.M.; Phelps, J.J.; Barkwith, A.; Hurst, M.D.; Ellis, M.A.; Plater, A.J. The effectiveness of beach mega-nourishment, assessed over three management epochs. J. Environ. Manage. 2016, 184, 400–408. [Google Scholar] [CrossRef] [PubMed]
- Tollefson, J. Hurricane sweeps into climate-adaptation debate. Nature 2012, 491, 167–168. [Google Scholar] [CrossRef] [PubMed]
- Feuer, A. Protecting New York City, before next time. New York Times. 3 November 2012. Available online: http://www.nytimes.com/2012/11/04/nyregion/protecting-new-york-city-before-next-time.html (accessed on 1 October 2016).
- Arkema, K.K.; Guannel, G.; Verutes, G.; Wood, S.A.; Guerry, A.; Ruckelshaus, M.; Kareiva, P.; Lacayo, M.; Silver, J.M. Coastal habitats shield people and property from sea-level rise and storms. Nature Clim. Change 2013, 3, 913–918. [Google Scholar] [CrossRef]
- Brandon, C.M.; Woodruff, J.D.; Orton, P.M.; Donnelly, J.P. Evidence for elevated coastal vulnerability following large-scale historical oyster bed harvesting. Earth Surf. Proc. Land. 2016, 41, 1136–1143. [Google Scholar] [CrossRef]
- Haff, P.K. Neogeomorphology, prediction, and the Anthropic landscape. In Prediction in Geomorphology; Wilcock, P.R., Iverson, R.M., Eds.; Geophysical Monograph Series; American Geophysical Union: Washington, DC, USA, 2003; Volume 135, pp. 15–26. [Google Scholar]
- Ma, Z.; Melville, D.S.; Liu, J.; Chen, Y.; Yang, H.; Ren, W.; Zhang, Z.; Piersma, T.; Li, B. Rethinking China’s new great wall. Science 2014, 346, 912–914. [Google Scholar] [CrossRef] [PubMed]
- Marriner, N.; Morhange, C.; Doumet-Serhal, C.; Carbonel, P. Geoscience rediscovers Phoenicia’s buried harbors. Geology 2006, 34, 1–4. [Google Scholar] [CrossRef]
- Qingzhou, W. The protection of China’s ancient cities from flood damage. Disasters 1989, 13, 193–227. [Google Scholar] [CrossRef] [PubMed]
- Valverde, H.R.; Trembanis, A.C.; Pilkey, O.H. Summary of beach nourishment episodes on the US east coast barrier islands. J. Coast. Res. 1999, 15, 1100–1118. [Google Scholar]
- Hapke, C.J.; Kratzmann, M.G.; Himmelstoss, E.A. Geomorphic and human influence on large-scale coastal change. Geomorphology 2013, 199, 160–170. [Google Scholar] [CrossRef]
- Willis, C.M.; Griggs, G.B. Reductions in fluvial sediment discharge by coastal dams in California and implications for beach sustainability. J. Geol. 2003, 111, 167–182. [Google Scholar] [CrossRef]
- Hinkel, J.; Lincke, D.; Vafeidis, A.T.; Perrette, M.; Nicholls, R.J.; Tol, R.S.; Marzeion, B.; Fettweis, X.; Ionescu, C.; Levermann, A. Coastal flood damage and adaptation costs under 21st century sea-level rise. Proc. Natl. Acad. Sci. USA 2014, 111, 3292–3297. [Google Scholar] [CrossRef] [PubMed]
- Dearing, J.A.; Acma, B.; Bub, S.; Chambers, F.M.; Chen, X.; Cooper, J.; Crook, D.; Dong, X.H.; Dotterweich, M.; Edwards, M.E.; et al. Social-ecological systems in the Anthropocene: the need for integrating social and biophysical records at regional scales. Anthropocene Rev. 2015, 2, 220–246. [Google Scholar] [CrossRef]
- Walker, B.; Meyers, J.A. Thresholds in ecological and socialecological systems: A developing database. Ecol. Soc. 2004, 3, 1–16. [Google Scholar]
- McNamara, D.E.; Werner, B.T. Coupled barrier island–resort model: 2. Tests and predictions along Ocean City and Assateague Island National Seashore, Maryland. J. Geophys. Res. Earth Surf. 2008, 113, F01017. [Google Scholar] [CrossRef]
- Haff, P.K. Technology and human purpose: The problem of solids transport on the Earth’s surface. Earth Syst. Dynam. 2012, 3, 149–156. [Google Scholar] [CrossRef]
- Haff, P.K. Purpose in the Anthropocene: Dynamical role and physical basis. Anthropocene 2016. [Google Scholar] [CrossRef]
- Ellis, E.C.; Antill, E.C.; Kreft, H. All is not loss: plant biodiversity in the Anthropocene. PLoS ONE 2012, 7, e30535. [Google Scholar] [CrossRef] [PubMed]
- Liu, J.; Hull, V.; Batistella, M.; DeFries, R.; Dietz, T.; Fu, F.; Hertel, T.W.; Izaurralde, R.C.; Lambin, E.F.; Li, S.; et al. Framing sustainability in a telecoupled world. Ecol. Soc. 2013, 18, 26. [Google Scholar] [CrossRef]
- Rindfuss, R.R.; Walsh, S.J.; Turner, B.L.; Fox, J.; Mishra, V. Developing a science of land change: challenges and methodological issues. Proc. Natl. Acad. Sci. USA 2004, 101, 13976–13981. [Google Scholar] [CrossRef] [PubMed]
- Turner, B.L.; Lambin, E.F.; Reenberg, A. The emergence of land change science for global environmental change and sustainability. Proc. Natl. Acad. Sci. USA 2007, 104, 20666–20671. [Google Scholar] [CrossRef] [PubMed]
- Parker, D.C.; Hessl, A.; Davis, S.C. Complexity, land-use modeling, and the human dimension: dundamental challenges for mapping unknown outcome spaces. Geoforum 2008, 39, 789–804. [Google Scholar] [CrossRef]
- Van Asselen, S.; Verburg, P.H. Land cover change or land-use intensification: simulating land system change with a global-scale land change model. Glob. Change Biol. 2013, 19, 3648–3667. [Google Scholar] [CrossRef] [PubMed]



{kind=link}
{kind=link}
{kind=link}
{kind=link}
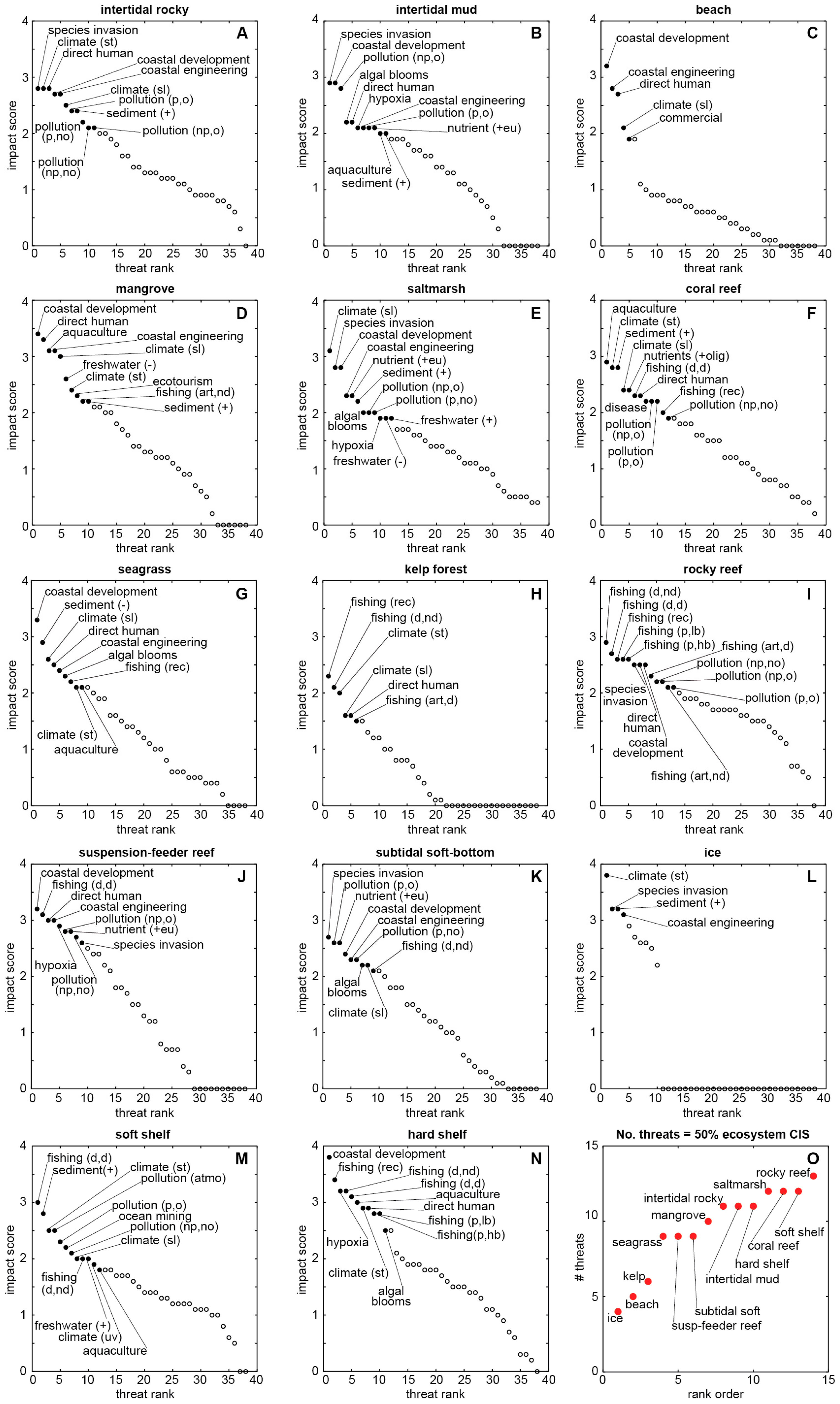
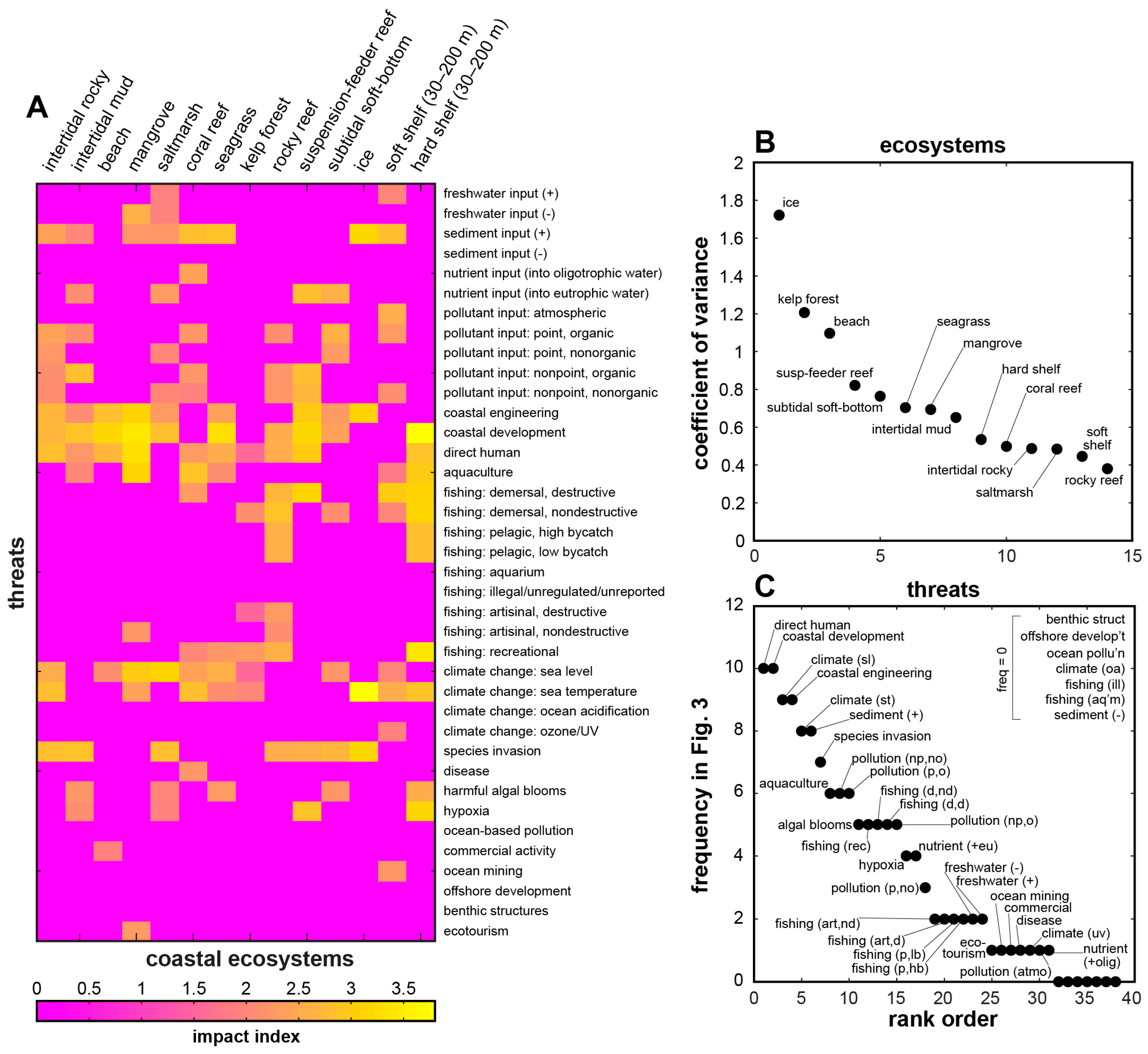
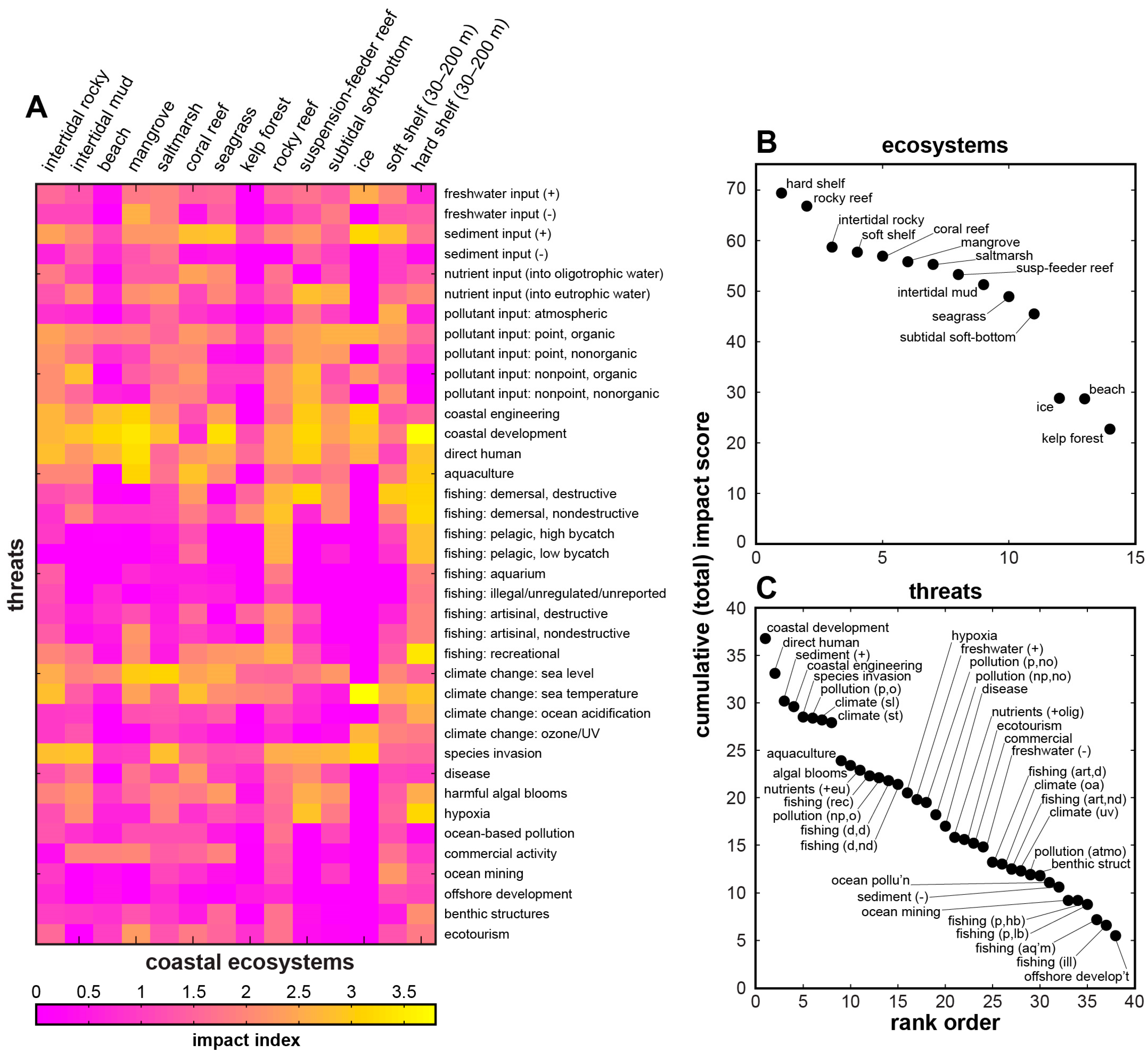
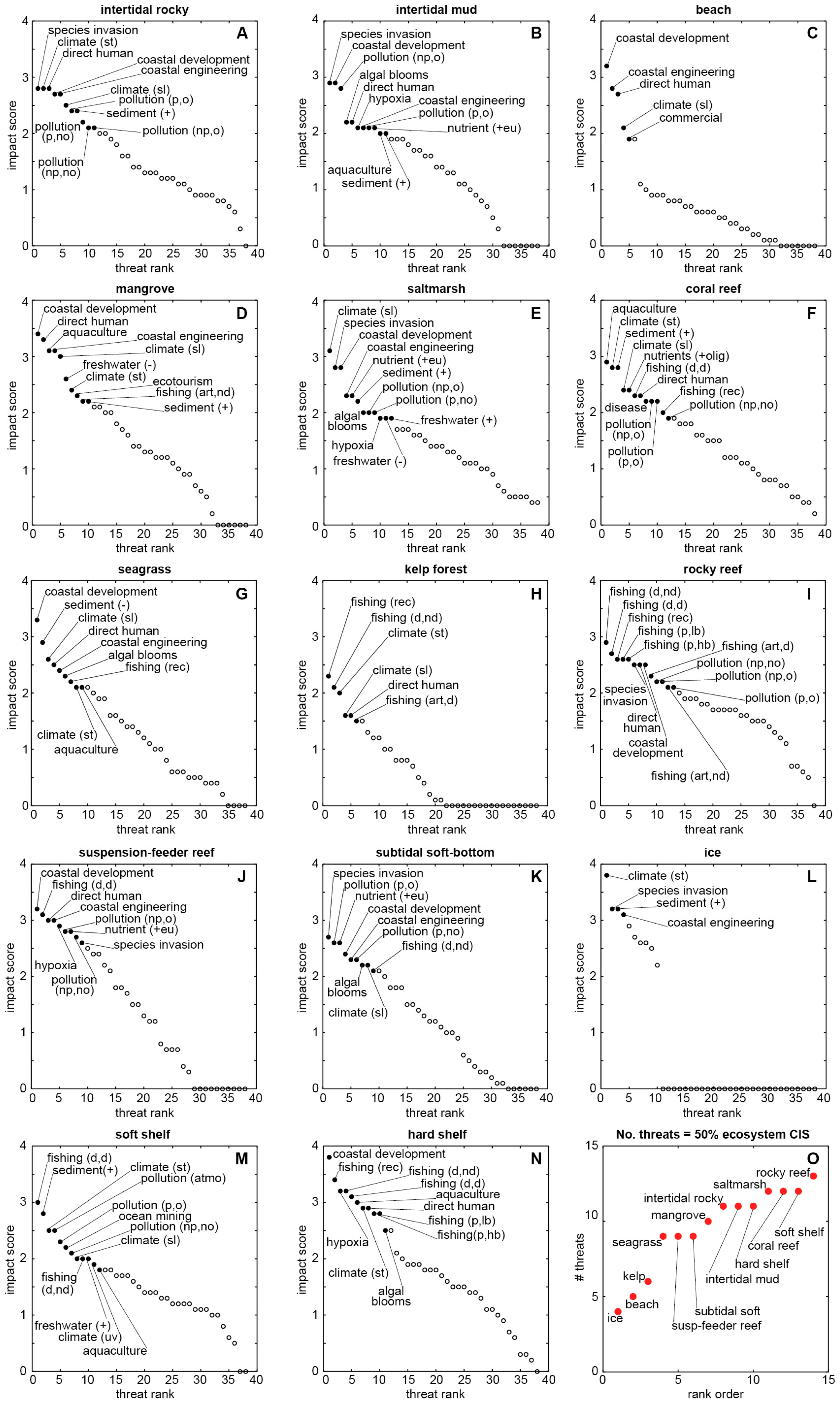
| Biomes & Coastal Natural Systems | Anthromes & Coastal Human Systems | Coastal Impacts & Threats | |
|---|---|---|---|
| Goodall [61] | Ellis & Ramankutty [18] | Halpern et al. [3] | Halpern et al. [23] |
| natural terrestrial coastal wet coastal dry aquatic (marine) intertidal & littoral coral reefs estuaries & enclosed seas continental shelves managed aquatic ecosystems | exurban population densities (persons per km2) dense (>100) residential (10–100) populated (1–10) remote (<1) groups dense settlements villages croplands rangelands forested wild | impacts nutrients (fertilizer) organic pollutants (pesticides) inorganic pollutants (impervious surfaces) direct human (population density) fishing: pelagic, low bycatch fishing: pelagic, high bycatch fishing: demersal, destructive fishing: demersal, non-destructive, low bycatch fishing: demersal, non-destructive, high bycatch fishing: artisanal oil rigs (benthic structures) invasive species ocean pollution shipping ocean sea-surface temperature UV ocean acidification "missing" impacts hypoxic zones coastal engineering (habitat alteration) non-shipping cargo (ferries, cruise ships) aquaculture disease fishing: illegal, unreported, unregulated fishing: recreational sedimentation change freshwater input change tourism [historical data] | threats (see Figure 2, Figure 3 and Figure 4) freshwater input increase freshwater input decrease sediment input increase sediment input decrease nutrient input into oligotrophic system nutrient input into eutrophic system pollutant input: atmospheric pollutant input: point source, organic pollutant input: point source, nonorganic pollutant input: nonpoint source, organic pollutant input: nonpoint source, nonorganic coastal engineering coastal development direct human aquaculture fishing: demersal, destructive fishing: demersal, nondestructive fishing: pelagic, high bycatch fishing: pelagic, low bycatch fishing: aquarium fishing: illegal, unreported, unregulated fishing: artisanal, destructive fishing: artisanal, nondestructive fishing: recreational climate change: sea level climate change: sea temperature climate change: ocean acidification climate change: UV species invasion disease harmful algal blooms hypoxia ocean-based pollution commercial activity ocean mining offshore development benthic structures ecotourism |
| Olson/WWF/Global 200 [62] | IPCC [5] | ||
| terrestrial mangroves freshwater large river deltas temperate coastal rivers tropical/subtropical coastal rivers oceanic islands marine polar temperate shelves & sea temperate upwelling tropical upwelling coral | human systems human settlements industry, infrastructure, transport, & network industries fisheries, aquaculture, & agriculture coastal tourism & recreation health | ||
| IPCC [5] | |||
| natural low-lying coastal systems beaches, barriers, dunes rocky coasts wetlands, seagrass coral reefs coastal aquifers estuaries, lagoons deltas | |||
| Halpern et al. [23] | |||
| coastal ecosystems (see Figure 2, Figure 3 and Figure 4) intertidal rocky intertidal mud beach mangrove saltmarsh coral reef seagrass kelp rocky reef suspension-feeder reef subtidal soft bottom ice soft shelf (30–200 m) hard shelf (30–200 m) | |||
| Terrestrial Anthromes [18] | Coastal Anthromes (Proposed Here) | Marine Anthromes (Speculative) |
|---|---|---|
| < ~10 m elevation | ||
| dense settlements | dense settlements | |
| urban | urban | |
| dense settlements | dense settlements | |
| villages | villages | |
| rice villages | rice villages | |
| irrigated villages | irrigated villages | |
| cropped & pastoral villages | cropped & pastoral villages | |
| pastoral villages | pastoral villages | |
| rainfed villages | rainfed villages | |
| rainfed mosaic villages | rainfed mosaic villages | |
| croplands | croplands | |
| residential irrigated cropland | residential irrigated cropland | |
| residential rainfed mosaic | residential rainfed mosaic | |
| populated irrigated cropland | populated irrigated cropland | |
| populated rainfed cropland | populated rainfed cropland | |
| remote croplands | remote croplands | |
| rangelands | rangelands | |
| residential rangelands | residential rangelands | |
| populated rangelands | populated rangelands | |
| remote rangelands | remote rangelands | |
| forested | forested | |
| populated forest | populated forest | |
| remote forest | remote forest | |
| wildlands | wildlands | |
| wild forest | wild forest | |
| sparse trees | sparse trees | |
| barren | Barren | |
| < 200 m depth OR < 200 nm offshore (Limit of EEZ) | > 200 m depth OR > 200 nm offshore (Limit of EEZ) | |
| direct human | direct human | |
| industrial complex | maritime use mosaic | |
| coastal engineering | offshore fisheries | |
| tourism & recreation mosaic | intensive pelagic fishing | |
| inshore/nearshore maritime use mosaic | deep-sea demersal fishing | |
| terrestrial input zones | low-intensity offshore fishing | |
| inshore sediment/nutrient/pollutant inputs | benthic infrastructure | |
| nearshore sediment/nutrient/pollutant | seabed mining | |
| mixing | oil rigs | |
| aquaculture | wind farms | |
| high-pollution inshore aquaculture | ocean wildlands | |
| low-pollution inshore aquaculture | ||
| onshore aquaculture | ||
| nearshore aquaculture | ||
| fisheries | ||
| intensive inshore demersal fishing | ||
| intensive inshore non-demersal fishing | ||
| low-intensity inshore fishing | ||
| intensive nearshore demersal fishing | ||
| intensive nearshore non-demersal fishing | ||
| low-intensity nearshore fishing | ||
| coastal wildlands |
© 2017 by the author. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license ( http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Lazarus, E.D. Toward a Global Classification of Coastal Anthromes. Land 2017, 6, 13. https://doi.org/10.3390/land6010013
Lazarus ED. Toward a Global Classification of Coastal Anthromes. Land. 2017; 6(1):13. https://doi.org/10.3390/land6010013
Chicago/Turabian StyleLazarus, Eli D. 2017. "Toward a Global Classification of Coastal Anthromes" Land 6, no. 1: 13. https://doi.org/10.3390/land6010013
APA StyleLazarus, E. D. (2017). Toward a Global Classification of Coastal Anthromes. Land, 6(1), 13. https://doi.org/10.3390/land6010013