
Responses of Typical Riparian Vegetation to Annual Variation of River Flow in a Semi-Arid Climate Region: Case Study of China’s Xiliao River


 ,
,
Abstract
1. Introduction
2. Materials and Methods
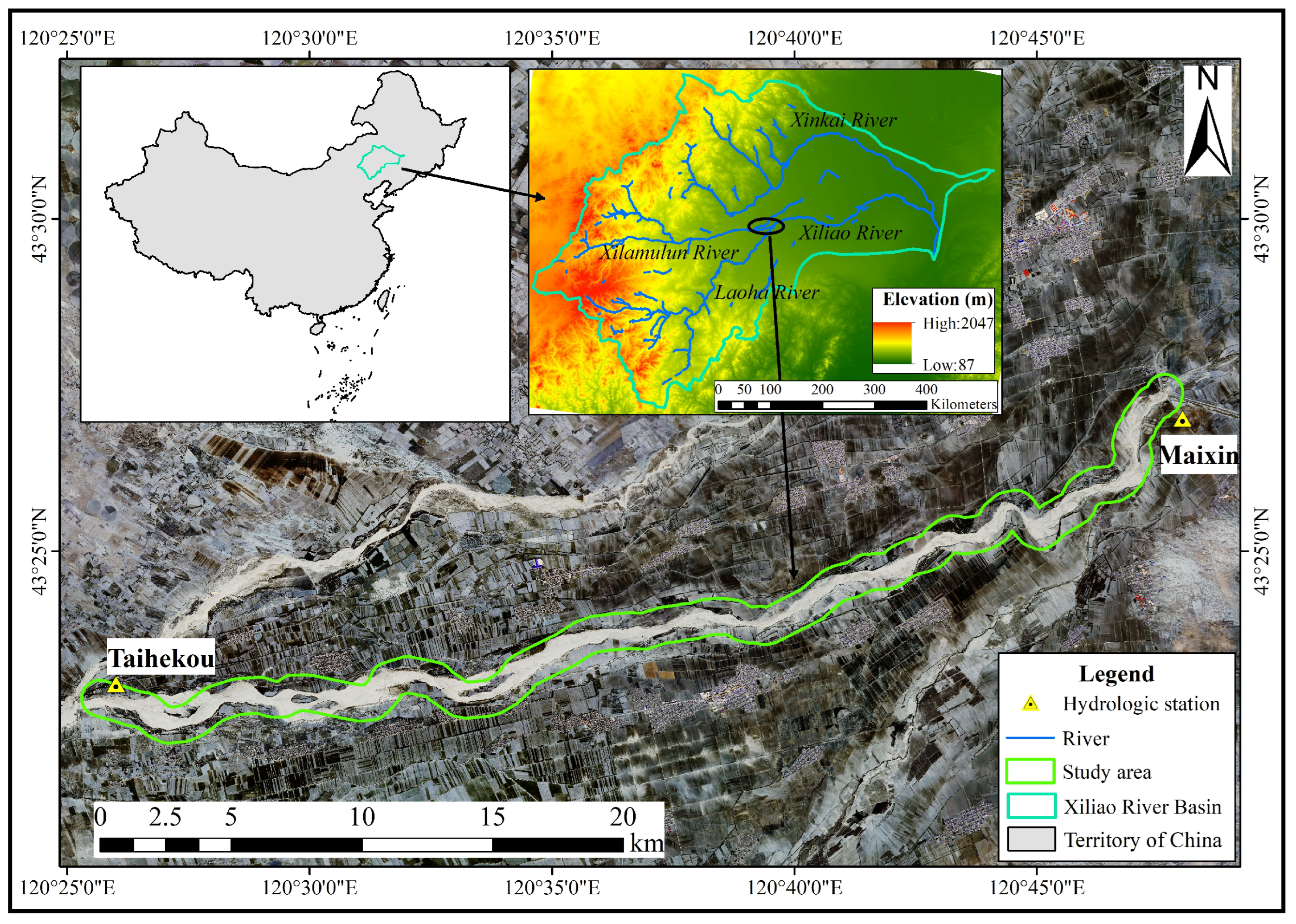
2.1. Study Area
2.2. Data Sources
2.3. Selection of Hydrological, Riparian Vegetation, and Meteorological Indicators
2.4. Trend Tests and Relationship Simulation
2.4.1. Analysis of Key Driving Factors for Riparian Vegetation
2.4.2. Fitting of the Vegetation Indicators and Key Driving Factors
3. Results
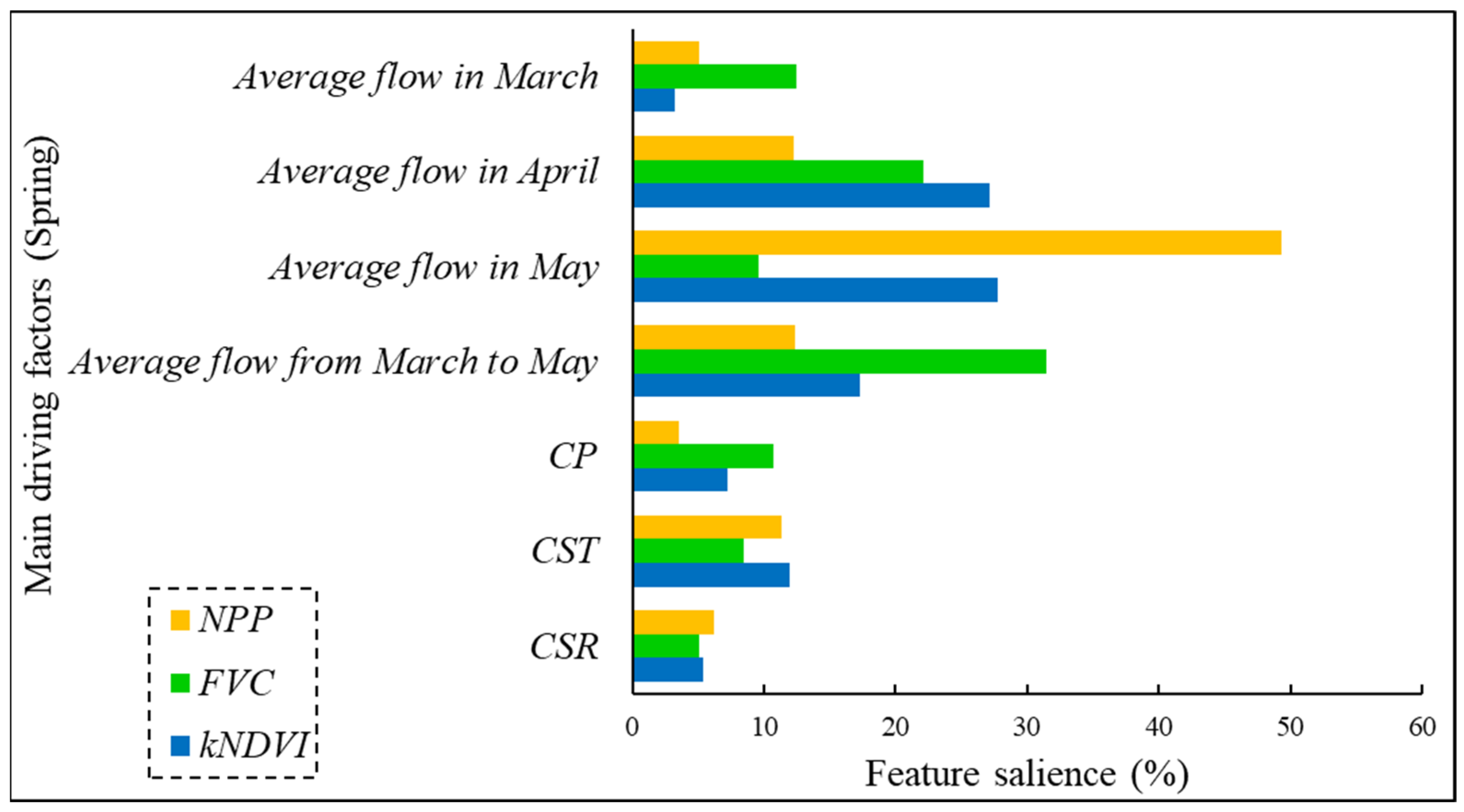
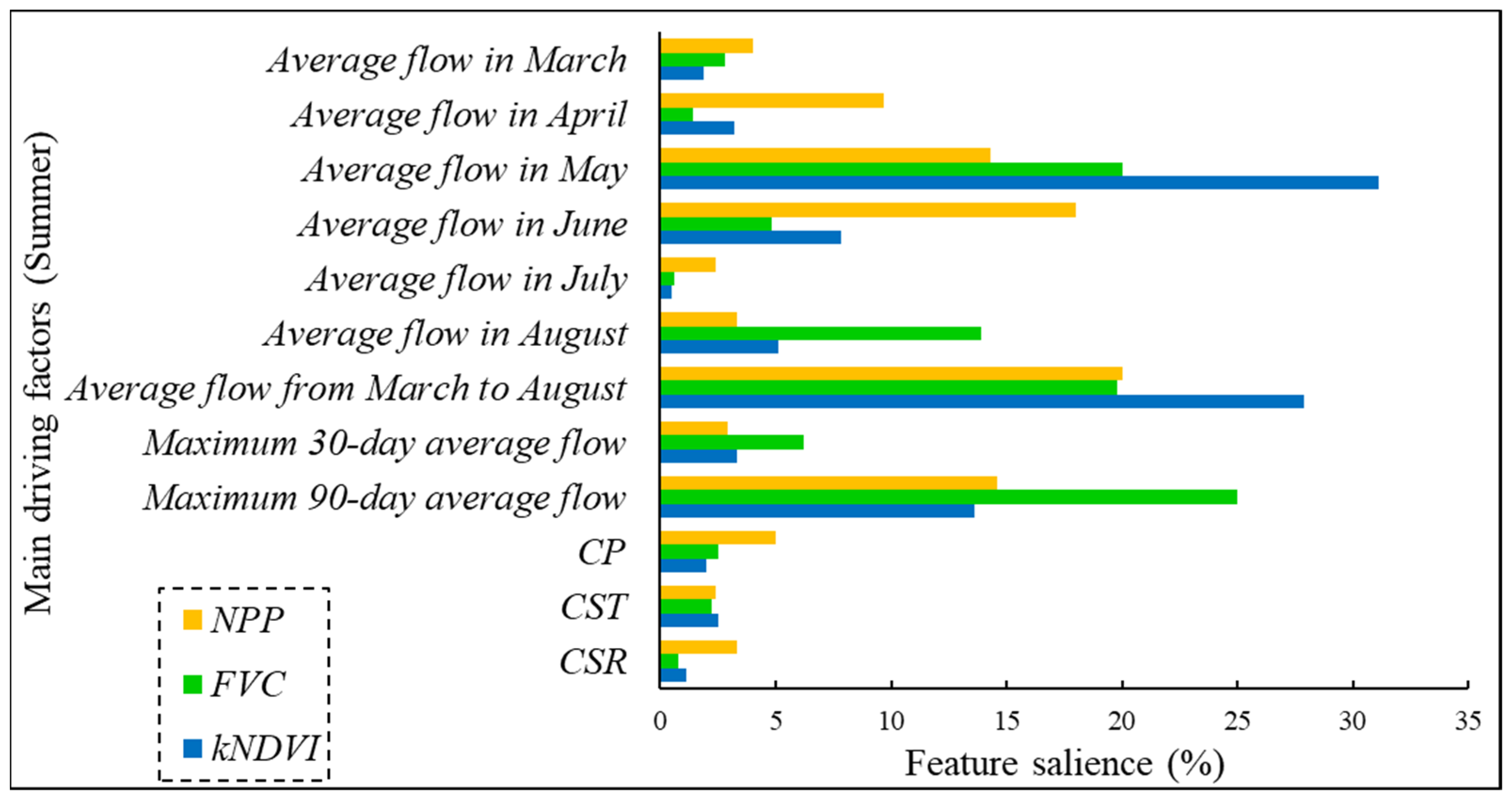
3.1. Analysis of Key Driving Factors for Riparian Vegetation
3.1.1. Analysis of Key Driving Factors for Riparian Vegetation in May
3.1.2. Analysis of Key Driving Factors for Riparian Vegetation in August
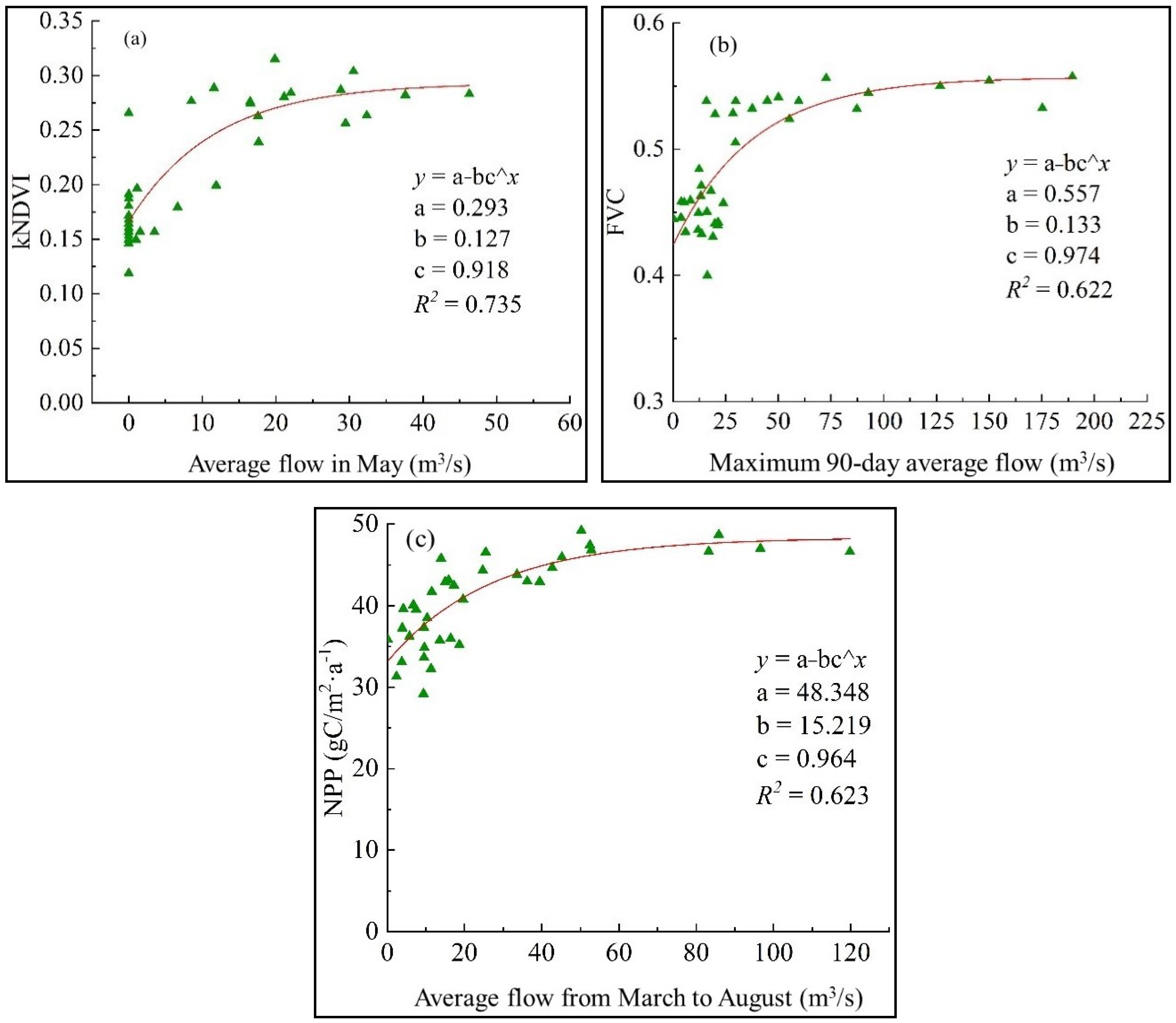
3.2. Simulation of Relationships Between Riparian Vegetation and Its Key Driving Factors
3.2.1. Relationships Between Riparian Vegetation and Its Key Driving Factors in May
3.2.2. Relationships Between Riparian Vegetation and Its Key Driving Factors in August
4. Discussion
4.1. Flow Factors Had More Effect on Riparian Vegetation than Climate Factors
4.2. Riparian Vegetation Needs More Appropriate Pulse Flows and Base Flows
5. Conclusions
Author Contributions
Funding
Data Availability Statement
Conflicts of Interest
References
- Perry, L.G.; Andersen, D.C.; Reynolds, L.V.; Nelson, S.M.; Shafroth, P.B. Vulnerability of riparian ecosystems to elevated CO2 and climate change in arid and semiarid western North America. Glob. Change Biol. 2012, 18, 821–842. [Google Scholar] [CrossRef]
- Garssen, A.G.; Verhoeven, J.T.A.; Soons, M.B. Effects of climate-induced increases in summer drought on riparian plant species: A meta-analysis. Freshw. Biol. 2014, 59, 1052–1063. [Google Scholar] [CrossRef]
- Poff, N.L.; Allan, J.D.; Bain, M.B.; Karr, J.R.; Prestegaard, K.L.; Richter, B.D.; Sparks, R.E.; Stromberg, J.C. The natural flow regime. Bioscience 1997, 47, 769–784. [Google Scholar] [CrossRef]
- Broich, M.; Tulbure, M.G.; Verbesselt, J.; Xin, Q.C.; Wearne, J. Quantifying Australia’s dryland vegetation response to flooding and drought at sub-continental scale. Remote Sens. Environ. 2018, 212, 60–78. [Google Scholar] [CrossRef]
- Stella, J.C.; Rodríguez-González, P.M.; Dufour, S.; Bendix, J. Riparian vegetation research in Mediterranean-climate regions: Common patterns, ecological processes, and considerations for management. Hydrobiologia 2013, 719, 291–315. [Google Scholar] [CrossRef]
- Diehl, R.M.; Merritt, D.M.; Wilcox, A.C.; Scott, M.L. Applying Functional Traits to Ecogeomorphic Processes in Riparian Ecosystems. Bioscience 2017, 67, 729–743. [Google Scholar] [CrossRef]
- Lan, D.; Yu, R.H. New grassland riparian zone delineation method for calculating ecological water demand to guide management goals. River Res. Appl. 2020, 36, 1838–1851. [Google Scholar] [CrossRef]
- Sims, N.C.; Colloff, M.J. Remote sensing of vegetation responses to flooding of a semi-arid floodplain: Implications for monitoring ecological effects of environmental flows. Ecol. Indic. 2012, 18, 387–391. [Google Scholar] [CrossRef]
- Murray-Hudson, M.; Wolski, P.; Ringrose, S. Scenarios of the impact of local and upstream changes in climate and water use on hydro-ecology in the Okavango Delta, Botswana. J. Hydrol. 2006, 331, 73–84. [Google Scholar] [CrossRef]
- Zhu, K.; Chiariello, N.R.; Tobeck, T.; Fukami, T.; Field, C.B. Nonlinear, interacting responses to climate limit grassland production under global change. Proc. Natl. Acad. Sci. USA 2016, 113, 10589–10594. [Google Scholar] [CrossRef]
- Beauchamp, V.B.; Shafroth, P.B. Floristic composition, beta diversity, and nestedness of reference sites for restoration of xeroriparian areas. Ecol. Appl. 2011, 21, 465–476. [Google Scholar] [CrossRef] [PubMed]
- McShane, R.R.; Auerbach, D.A.; Friedman, J.M.; Auble, G.T.; Shafroth, P.B.; Merigliano, M.F.; Scott, M.L.; Poff, N.L. Distribution of invasive and native riparian woody plants across the western USA in relation to climate, river flow, floodplain geometry and patterns of introduction. Ecography 2015, 38, 1254–1265. [Google Scholar] [CrossRef]
- Woodhouse, C.A.; Pederson, G.T.; Morino, K.; McAfee, S.A.; McCabe, G.J. Increasing influence of air temperature on upper Colorado River streamflow. Geophys. Res. Lett. 2016, 43, 2174–2181. [Google Scholar] [CrossRef]
- Milly, P.C.D.; Dunne, K.A. Colorado River flow dwindles as warming-driven loss of reflective snow energizes evaporation. Science 2020, 367, 1252–1255. [Google Scholar] [CrossRef] [PubMed]
- Zema, D.A.; Bombino, G.; Denisi, P.; Lucas-Borja, M.E.; Zimbone, S.M. Evaluating the effects of check dams on channel geometry, bed sediment size and riparian vegetation in Mediterranean mountain torrents. Sci. Total Environ. 2018, 642, 327–340. [Google Scholar] [CrossRef]
- Diehl, R.M.; Wilcox, A.C.; Stella, J.C. Evaluation of the integrated riparian ecosystem response to future flow regimes on semiarid rivers in Colorado, USA. J. Environ. Manag. 2020, 271, 111037. [Google Scholar] [CrossRef]
- Newman, B.D.; Wilcox, B.P.; Archer, S.R.; Breshears, D.D.; Dahm, C.N.; Duffy, C.J.; McDowell, N.G.; Phillips, F.M.; Scanlon, B.R.; Vivoni, E.R. Ecohydrology of water-limited environments: A scientific vision. Water Resour. Res. 2006, 42, W06302. [Google Scholar] [CrossRef]
- Merritt, D.M.; Scott, M.L.; Poff, N.L.; Auble, G.T.; Lytle, D.A. Theory, methods and tools for determining environmental flows for riparian vegetation: Riparian vegetation-flow response guilds. Freshw. Biol. 2010, 55, 206–225. [Google Scholar] [CrossRef]
- Diehl, R.M.; Wilcox, A.C.; Merritt, D.M.; Perkins, D.W.; Scott, J.A. Development of an eco-geomorphic modeling framework to evaluate riparian ecosystem response to flow-regime changes. Ecol. Eng. 2018, 123, 112–126. [Google Scholar] [CrossRef]
- Nagler, P.L.; Glenn, E.P.; Hinojosa-Huerta, O. Synthesis of ground and remote sensing data for monitoring ecosystem functions in the Colorado River Delta, Mexico. Remote Sens. Environ. 2009, 113, 1473–1485. [Google Scholar] [CrossRef]
- Mukherjee, K.; Pal, S. Hydrological and landscape dynamics of floodplain wetlands of the Diara region, Eastern India. Ecol. Indic. 2021, 121, 106961. [Google Scholar] [CrossRef]
- Powell, S.J.; Jakeman, A.; Croke, B. Can NDVI response indicate the effective flood extent in macrophyte dominated floodplain wetlands? Ecol. Indic. 2014, 45, 486–493. [Google Scholar] [CrossRef]
- Dzubáková, K.; Molnar, P.; Schindler, K.; Trizna, M. Monitoring of riparian vegetation response to flood disturbances using terrestrial photography. Hydrol. Earth Syst. Sci. 2015, 19, 195–208. [Google Scholar] [CrossRef]
- Gurnell, A.M.; Bertoldi, W.; Corenblit, D. Changing river channels: The roles of hydrological processes, plants and pioneer fluvial landforms in humid temperate, mixed load, gravel bed rivers. Earth-Sci. Rev. 2012, 111, 129–141. [Google Scholar] [CrossRef]
- Corenblit, D.; Baas, A.; Balke, T.; Bouma, T.; Fromard, F.; Garófano-Gómez, V.; González, E.; Gurnell, A.M.; Hortobágyi, B.; Julien, F.; et al. Engineer pioneer plants respond to and affect geomorphic constraints similarly along water-terrestrial interfaces world-wide. Global Ecol. Biogeogr. 2015, 24, 1363–1376. [Google Scholar] [CrossRef]
- Gonsamo, A.; Chen, J.M.; Lombardozzi, D. Global vegetation productivity response to climatic oscillations during the satellite era. Glob. Change Biol. 2016, 22, 3414–3426. [Google Scholar] [CrossRef] [PubMed]
- Gu, Z.J.; Duan, X.W.; Shi, Y.D.; Li, Y.; Pan, X. Spatiotemporal variation in vegetation coverage and its response to climatic factors in the Red River Basin, China. Ecol. Indic. 2018, 93, 54–64. [Google Scholar] [CrossRef]
- Cai, D.W.; Ge, Q.S.; Wang, X.M.; Liu, B.L.; Goudie, A.S.; Hu, S. Contributions of ecological programs to vegetation restoration in arid and semiarid China. Environ. Res. Lett. 2020, 15, 114046. [Google Scholar] [CrossRef]
- Piao, S.L.; Cui, M.D.; Chen, A.P.; Wang, X.H.; Ciais, P.; Liu, J.; Tang, Y.H. Altitude and temperature dependence of change in the spring vegetation green-up date from 1982 to 2006 in the Qinghai-Xizang Plateau. Agric. For. Meteorol. 2011, 151, 1599–1608. [Google Scholar] [CrossRef]
- Xu, L.; Myneni, R.B.; Chapin, F.S.; Callaghan, T.V.; Pinzon, J.E.; Tucker, C.J.; Zhu, Z.; Bi, J.; Ciais, P.; Tommervik, H.; et al. Temperature and vegetation seasonality diminishment over northern lands. Nat. Clim. Change 2013, 3, 581–586. [Google Scholar] [CrossRef]
- Ukkola, A.M.; Prentice, I.C.; Keenan, T.F.; van Dijk, A.I.J.M.; Viney, N.R.; Myneni, R.B.; Bi, J. Reduced streamflow in water-stressed climates consistent with CO effects on vegetation. Nat. Clim. Change 2016, 6, 75–78. [Google Scholar] [CrossRef]
- Shao, R.; Zhang, B.Q.; Su, T.X.; Biao, L.; Cheng, L.Y.; Xue, Y.Y.; Yang, W.J. Estimating the Increase in Regional Evaporative Water Consumption as a Result of Vegetation Restoration Over the Loess Plateau, China. J. Geophys. Res.-Atmos. 2019, 124, 11783–11802. [Google Scholar] [CrossRef]
- Kou, P.L.; Xu, Q.; Jin, Z.; Yunus, A.P.; Luo, X.B.; Liu, M.H. Complex anthropogenic interaction on vegetation greening in the Chinese Loess Plateau. Sci. Total Environ. 2021, 778, 146065. [Google Scholar] [CrossRef] [PubMed]
- Nemani, R.R.; Keeling, C.D.; Hashimoto, H.; Jolly, W.M.; Piper, S.C.; Tucker, C.J.; Myneni, R.B.; Running, S.W. Climate-driven increases in global terrestrial net primary production from 1982 to 1999. Science 2003, 300, 1560–1563. [Google Scholar] [CrossRef] [PubMed]
- Reyer, C.P.O.; Leuzinger, S.; Rammig, A.; Wolf, A.; Bartholomeus, R.P.; Bonfante, A.; de Lorenzi, F.; Dury, M.; Gloning, P.; Abou Jaoudé, R.; et al. A plant’s perspective of extremes: Terrestrial plant responses to changing climatic variability. Glob. Change Biol. 2013, 19, 75–89. [Google Scholar] [CrossRef]
- Gao, J.b.; Jiao, K.W.; Wu, S.H.; Ma, D.Y.; Zhao, D.S.; Yin, Y.H.; Dai, E.F. Past and future effects of climate change on spatially heterogeneous vegetation activity in China. Earths Future 2017, 5, 679–692. [Google Scholar] [CrossRef]
- Hou, W.J.; Gao, J.B.; Wu, S.H.; Dai, E.F. Interannual Variations in Growing-Season NDVI and Its Correlation with Climate Variables in the Southwestern Karst Region of China. Remote Sens. 2015, 7, 11105–11124. [Google Scholar] [CrossRef]
- Wu, D.H.; Zhao, X.; Liang, S.L.; Zhou, T.; Huang, K.C.; Tang, B.J.; Zhao, W.Q. Time-lag effects of global vegetation responses to climate change. Glob. Change Biol. 2015, 21, 3520–3531. [Google Scholar] [CrossRef] [PubMed]
- Xu, H.J.; Wang, X.P.; Yang, T.B. Trend shifts in satellite-derived vegetation growth in Central Eurasia, 1982–2013. Sci. Total Environ. 2017, 579, 1658–1674. [Google Scholar] [CrossRef]
- Chu, H.S.; Venevsky, S.; Wu, C.; Wang, M.H. NDVI-based vegetation dynamics and its response to climate changes at Amur-Heilongjiang River Basin from 1982 to 2015. Sci. Total Environ. 2019, 650, 2051–2062. [Google Scholar] [CrossRef] [PubMed]
- Merritt, D.M.; Wohl, E.E. Processes governing hydrochory along rivers: Hydraulics, hydrology, and dispersal phenology. Ecol. Appl. 2002, 12, 1071–1087. [Google Scholar] [CrossRef]
- Karrenberg, S.; Edwards, P.J.; Kollmann, J. The life history of Salicaceae living in the active zone of floodplains. Freshw. Biol. 2002, 47, 733–748. [Google Scholar] [CrossRef]
- Farmer, R.E.; Bonner, F.T. Germination and Initial Growth of Eastern Cottonwood as Influenced by Moisture Stress Temperature and Storage. Bot. Gaz. 1967, 128, 211–215. [Google Scholar] [CrossRef]
- Greet, J.; Webb, J.A.; Cousens, R.D. The importance of seasonal flow timing for riparian vegetation dynamics: A systematic review using causal criteria analysis. Freshw. Biol. 2011, 56, 1231–1247. [Google Scholar] [CrossRef]
- Lin, N.; Jiang, R.Z.; Liu, Q.; Yang, H.; Liu, H.L.; Yang, Q. Quantifying the Spatiotemporal Variation of Evapotranspiration of Different Land Cover Types and the Contribution of Its Associated Factors in the Xiliao River Plain. Remote Sens. 2022, 14, 252. [Google Scholar] [CrossRef]
- Chen, P.F. Monthly net primary productivity of China’s terrestrial ecosystems north of 18°N latitude: 1 km raster dataset (1985–2015). J. Glob. Change Data Discov. 2019, 3, 34–41. [Google Scholar]
- De Girolamo, A.M.; Barca, E.; Pappagallo, G.; Lo Porto, A. Simulating ecologically relevant hydrological indicators in a temporary river system. Agric. Water Manag. 2017, 180, 194–204. [Google Scholar] [CrossRef]
- Hao, Z.; Rallings, A.M.; Espinoza, V.; Luo, P.P.; Duan, W.L.; Peng, Q.D.; Gao, Y.; Viers, J.H. Flowing from East to West: A bibliometric analysis of recent advances in environmental flow science in China. Ecol. Indic. 2021, 125, 107358. [Google Scholar] [CrossRef]
- Richter, B.D.; Baumgartner, J.V.; Powell, J.; Braun, D.P. A method for assessing hydrologic alteration within ecosystems. Conserv. Biol. 1996, 10, 1163–1174. [Google Scholar] [CrossRef]
- Richter, B.D.; Mathews, R.; Wigington, R. Ecologically sustainable water management: Managing river flows for ecological integrity. Ecol. Appl. 2003, 13, 206–224. [Google Scholar] [CrossRef]
- Arthington, A.H.; Bunn, S.E.; Poff, N.L.; Naiman, R.J. The challenge of providing environmental flow rules to sustain river ecosystems. Ecol. Appl. 2006, 16, 1311–1318. [Google Scholar] [CrossRef] [PubMed]
- Poff, N.L.; Zimmerman, J.K.H. Ecological responses to altered flow regimes: A literature review to inform the science and management of environmental flows. Freshw. Biol. 2010, 55, 194–205. [Google Scholar] [CrossRef]
- Hua, X.B.; Sirguey, P.; Ohlemüller, R. Recent trends in the timing of the growing season in New Zealand’s natural and semi-natural grasslands. Gisci Remote Sens. 2021, 58, 1090–1111. [Google Scholar] [CrossRef]
- Chen, B.J.; Jiapaer, G.; Yu, T.; Zhang, L.C.; Tu, H.Y.; Liang, H.W.; Lina, K.X.; Ju, T.W.; Ling, Q. The role of climatic factor timing on grassland net primary productivity in Altay, Xinjiang. Ecol. Indic. 2023, 157, 111243. [Google Scholar] [CrossRef]
- Xu, L.L.; Zhang, X.Z.; Wang, Y.L.; Fu, Y.; Yan, H.; Qian, S.; Cheng, L. Drivers of phenology shifts and their effect on productivity in northern grassland of China during 1984–2017-evidence from long-term observational data. Int. J. Biometeorol. 2021, 65, 527–539. [Google Scholar] [CrossRef] [PubMed]
- Bradford, J.B.; Lauenroth, W.K.; Burke, I.C.; Paruelo, J.M. The influence of climate, soils, weather, and land use on primary production and biomass seasonality in the US Great Plains. Ecosystems 2006, 9, 934–950. [Google Scholar] [CrossRef]
- Dong, Y.; Yin, D.Q.; Li, X.; Huang, J.X.; Su, W.; Li, X.C.; Wang, H.S. Spatial-Temporal Evolution of Vegetation NDVI in Association with Climatic, Environmental and Anthropogenic Factors in the Loess Plateau, China during 2000–2015: Quantitative Analysis Based on Geographical Detector Model. Remote Sens. 2021, 13, 4380. [Google Scholar] [CrossRef]
- Feng, X.J.; Tian, J.; Wang, Y.X.; Wu, J.J.; Liu, J.; Ya, Q.; Li, Z.S. Spatio-Temporal Variation and Climatic Driving Factors of Vegetation Coverage in the Yellow River Basin from 2001 to 2020 Based on. Forests 2023, 14, 620. [Google Scholar] [CrossRef]
- Camps-Valls, G.; Campos-Taberner, M.; Moreno-Martínez, A.; Walther, S.; Duveiller, G.; Cescatti, A.; Mahecha, M.D.; Muñoz-Marí, J.; García-Haro, F.J.; Guanter, L.; et al. A unified vegetation index for quantifying the terrestrial biosphere. Sci. Adv. 2021, 7, eabc7447. [Google Scholar] [CrossRef]
- Wu, D.H.; Wu, H.; Zhao, X.; Zhou, T.; Tang, B.J.; Zhao, W.Q.; Jia, K. Evaluation of Spatiotemporal Variations of Global Fractional Vegetation Cover Based on GIMMS NDVI Data from 1982 to 2011. Remote Sens. 2014, 6, 4217–4239. [Google Scholar] [CrossRef]
- Tian, F.; Fensholt, R.; Verbesselt, J.; Grogan, K.; Horion, S.; Wang, Y.J. Evaluating temporal consistency of long-term global NDVI datasets for trend analysis. Remote Sens. Environ. 2015, 163, 326–340. [Google Scholar] [CrossRef]
- Yan, E.P.; Wang, G.X.; Lin, H.; Xia, C.Z.; Sun, H. Phenology-based classification of vegetation cover types in Northeast China using MODIS NDVI and EVI time series. Int. J. Remote Sens. 2015, 36, 489–512. [Google Scholar] [CrossRef]
- Gang, C.C.; Zhao, W.; Zhao, T.; Zhang, Y.; Gao, X.R.; Wen, Z.M. The impacts of land conversion and management measures on the grassland net primary productivity over the Loess Plateau, Northern China. Sci. Total Environ. 2018, 645, 827–836. [Google Scholar] [CrossRef]
- Bai, Z.G.; Dent, D.L.; Olsson, L.; Schaepman, M.E. Proxy global assessment of land degradation. Soil. Use Manag. 2008, 24, 223–234. [Google Scholar] [CrossRef]
- Liao, S.M.; Xue, L.Q.; Dong, Z.C.; Zhu, B.L.; Zhang, K.; Wei, Q.; Fu, F.B.; Wei, G.H. Cumulative ecohydrological response to hydrological processes in arid basins. Ecol. Indic. 2020, 111, 106005. [Google Scholar] [CrossRef]
- Zhang, Y.L.; Song, C.H.; Band, L.E.; Sun, G.; Li, J.X. Reanalysis of global terrestrial vegetation trends from MODIS products: Browning or greening? Remote Sens. Environ. 2017, 191, 145–155. [Google Scholar] [CrossRef]
- Dou, X.; Ma, X.F.; Huo, T.C.; Zhu, J.T.; Zhao, C.Y. Assessment of the environmental effects of ecological water conveyance over 31 years for a terminal lake in Central Asia. Catena 2022, 208, 105725. [Google Scholar] [CrossRef]
- Karabadji, N.E.; Korba, A.A.; Assi, A.; Seridi, H.; Aridhi, S.; Dhifli, W. Accuracy and diversity-aware multi-objective approach for random forest construction. Expert. Syst. Appl. 2023, 225, 120138. [Google Scholar] [CrossRef]
- Schonlau, M.; Zou, R.Y. The random forest algorithm for statistical learning. Stata J. 2020, 20, 3–29. [Google Scholar] [CrossRef]
- Breiman, L. Random forests. Mach. Learn. 2001, 45, 5–32. [Google Scholar] [CrossRef]
- Phillips, J.D. Evolutionary geomorphology: Thresholds and nonlinearity in landform response to environmental change. Hydrol. Earth Syst. Sci. 2006, 10, 731–742. [Google Scholar] [CrossRef]
- Liu, Y.L.; Lei, H.M. Responses of Natural Vegetation Dynamics to Climate Drivers in China from 1982 to 2011. Remote Sens. 2015, 7, 10243–10268. [Google Scholar] [CrossRef]
- Wen, L.; Mason, T.J.; Ryan, S.; Ling, J.E.; Saintilan, N.; Rodriguez, J. Monitoring long-term vegetation condition dynamics in persistent semi-arid wetland communities using time series of Landsat data. Sci. Total Environ. 2023, 905, 167212. [Google Scholar] [CrossRef]
- Guo, L.; Cheng, J.M.; Luedeling, E.; Koerner, S.E.; He, J.S.; Xu, J.C.; Gang, C.C.; Li, W.; Luo, R.M.; Peng, C.H. Critical climate periods for grassland productivity on China’s Loess Plateau. Agric. For. Meteorol. 2017, 233, 101–109. [Google Scholar] [CrossRef]
- Soons, M.B.; de Groot, G.A.; Ramirez, M.T.C.; Fraaije, R.G.A.; Verhoeven, J.T.A.; de Jager, M. Directed dispersal by an abiotic vector: Wetland plants disperse their seeds selectively to suitable sites along the hydrological gradient via water. Funct. Ecol. 2017, 31, 499–508. [Google Scholar] [CrossRef]
- Bewley, J.D. Seed germination and dormancy. Plant Cell 1997, 9, 1055–1066. [Google Scholar] [CrossRef] [PubMed]
- Jansson, R.; Zinko, U.; Merritt, D.M.; Nilsson, C. Hydrochory increases riparian plant species richness: A comparison between a free-flowing and a regulated river. J. Ecol. 2005, 93, 1094–1103. [Google Scholar] [CrossRef]
- Jäkäläniemi, A.; Tuomi, J.; Siikamäki, P.; Kilpiä, A. Colonization-extinction and patch dynamics of the perennial riparian plant. J. Ecol. 2005, 93, 670–680. [Google Scholar] [CrossRef]
- Zhang, S.H.; Ye, Z.X.; Chen, Y.N.; Xu, Y.F. Vegetation responses to an ecological water conveyance project in the lower reaches of the Heihe River basin. Ecohydrology 2017, 10, e1866. [Google Scholar] [CrossRef]
- Jia, L.; Shang, H.; Hu, G.; Menenti, M. Phenological response of vegetation to upstream river flow in the Heihe Rive basin by time series analysis of MODIS data. Hydrol. Earth Syst. Sci. 2011, 15, 1047–1064. [Google Scholar] [CrossRef]
- Ramírez-Hernández, J.; Rodríguez-Burgueño, J.E.; Zamora-Arroyo, F.; Carreón-Diazconti, C.; Pérez-González, D. lMimic pulse-base flows and groundwater in a regulated river in semiarid land: Riparian restoration issues. Ecol. Eng. 2015, 83, 239–248. [Google Scholar] [CrossRef]
- Nagler, P.L.; Hinojosa-Huerta, O.; Glenn, E.P.; Garcia-Hernandez, J.; Romo, R.; Curtis, C.; Huete, A.R.; Nelson, S.G. Regeneration of native trees in the presence of invasive saltcedar in the Colorado River delta, Mexico. Conserv. Biol. 2005, 19, 1842–1852. [Google Scholar] [CrossRef]
- Robertson, A.I.; Bacon, P.; Heagney, G. The responses of floodplain primary production to flood frequency and timing. J. Appl. Ecol. 2001, 38, 126–136. [Google Scholar] [CrossRef]
- Richardson, A.D.; Black, T.A.; Ciais, P.; Delbart, N.; Friedl, M.A.; Gobron, N.; Hollinger, D.Y.; Kutsch, W.L.; Longdoz, B.; Luyssaert, S.; et al. Influence of spring and autumn phenological transitions on forest ecosystem productivity. Philos. Trans. R. Soc. B Biol. Sci. 2010, 365, 3227–3246. [Google Scholar] [CrossRef]
- Nilsson, C.; Brown, R.L.; Jansson, R.; Merritt, D.M. The role of hydrochory in structuring riparian and wetland vegetation. Biol. Rev. 2010, 85, 837–858. [Google Scholar] [CrossRef] [PubMed]
- Fraaije, R.G.A.; ter Braak, C.J.F.; Verduyn, B.; Breeman, L.B.S.; Verhoeven, J.T.A.; Soons, M.B. Early plant recruitment stages set the template for the development of vegetation patterns along a hydrological gradient. Funct. Ecol. 2015, 29, 971–980. [Google Scholar] [CrossRef]
- Januschke, K.; Brunzel, S.; Haase, P.; Hering, D. Effects of stream restorations on riparian mesohabitats, vegetation and carabid beetles. Biodivers. Conserv. 2011, 20, 3147–3164. [Google Scholar] [CrossRef]
- Lorenz, A.W.; Korte, T.; Sundermann, A.; Januschke, K.; Haase, P. Macrophytes respond to reach-scale river restorations. J. Appl. Ecol. 2012, 49, 202–212. [Google Scholar] [CrossRef]
- Bino, G.; Sisson, S.A.; Kingsford, R.T.; Thomas, R.F.; Bowen, S. Developing state and transition models of floodplain vegetation dynamics as a tool for conservation decision-making: A case study of the Macquarie Marshes Ramsar wetland. J. Appl. Ecol. 2015, 52, 654–664. [Google Scholar] [CrossRef]
- Wu, H.W.; Zhao, G.Q.; Li, X.Y.; Wang, Y.; He, B.; Jiang, Z.Y.; Zhang, S.Y.; Sun, W. Identifying water sources used by alpine riparian plants in a restoration zone on the Qinghai-Tibet Plateau: Evidence from stable isotopes. Sci. Total Environ. 2019, 697, 134092. [Google Scholar] [CrossRef]
- Liu, X.; Zhuang, Q.L.; Lai, L.M.; Zhou, J.H.; Sun, Q.L.; Yi, S.G.; Liu, B.B.; Zheng, Y.R. Soil water use sources and patterns in shrub encroachment in semiarid grasslands of Inner Mongolia. Agric. For. Meteorol. 2021, 308, 108579. [Google Scholar] [CrossRef]
- Jarchow, C.J.; Nagler, P.L.; Glenn, E.P. Greenup and evapotranspiration following the Minute 319 pulse flow to Mexico: An analysis using Landsat 8 Normalized Difference Vegetation Index (NDVI) data. Ecol. Eng. 2017, 106, 776–783. [Google Scholar] [CrossRef]
- Scott, R.L.; Shuttleworth, W.J.; Goodrich, D.C.; Maddock, T. The water use of two dominant vegetation communities in a semiarid riparian ecosystem. Agric. For. Meteorol. 2000, 105, 241–256. [Google Scholar] [CrossRef]
- Puijalon, S.; Bouma, T.J.; Douady, C.J.; van Groenendael, J.; Anten, N.P.R.; Martel, E.; Bornette, G. Plant resistance to mechanical stress: Evidence of an avoidance-tolerance trade-off. New Phytol. 2011, 191, 1141–1149. [Google Scholar] [CrossRef] [PubMed]
- Grygoruk, M.; Kochanek, K.; Miroslaw-Swiatek, D. Analysis of long-term changes in inundation characteristics of near-natural temperate riparian habitats in the Lower Basin of the Biebrza Valley, Poland. J. Hydrol.-Reg. Stud. 2021, 36, 100844. [Google Scholar] [CrossRef]
- Main, A.C.; Greet, J.; Vivian, L.M.; Jones, C.S. Warmer water temperatures exacerbate the negative impacts of inundation on herbaceous riparian plants. Freshw. Biol. 2022, 67, 1162–1173. [Google Scholar] [CrossRef]
- Capon, S.J.; Brock, M.A. Flooding, soil seed bank dynamics and vegetation resilience of a hydrologically variable desert floodplain. Freshw. Biol. 2006, 51, 206–223. [Google Scholar] [CrossRef]
- Tiegs, S.D.; O’Leary, J.F.; Pohl, M.M.; Munill, C.L. Flood disturbance and riparian species diversity on the Colorado River Delta. Biodivers. Conserv. 2005, 14, 1175–1194. [Google Scholar] [CrossRef]
- Ivory, S.J.; McGlue, M.M.; Spera, S.; Silva, A.; Bergier, I. Vegetation, rainfall, and pulsing hydrology in the Pantanal, the world’s largest tropical wetland. Environ. Res. Lett. 2019, 14, 124017. [Google Scholar] [CrossRef]


{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Indicator Type | Indicator Name | Units | Ecological Significance | Extraction Method |
|---|---|---|---|---|
| River flow indicators | Average flow in each month from March to August Average flow for the whole period from March to May Average flow for the whole period from March to August Maximum 30-day average flow Maximum 90-day average flow | m3/s | Numerical statistics | |
| Climate indicators | Cumulative precipitation from March to May and from March to August | mm | ENVI data analyzed in ArcGIS (www.esri.com) | |
| Cumulative solar radiation from March to May and from March to August | W/m2 |
| ||
| Cumulative surface temperature from March to May and from March to August | K | |||
| Vegetation indicators | kNDVI in May and August | [59] | ||
| FVC in May and August |
| [61,62] | ||
| NPP in May and August | gC/(m2·year) | ArcGIS and ENVI statistics |
| Riparian Vegetation Indicators | Accuracy of Calibration | Accuracy of Validation | ||||
|---|---|---|---|---|---|---|
| R2 | RMSE | MAE | R2 | RMSE | MAE | |
| kNDVI | 0.869 | 0.003 | 0.003 | 0.837 | 0.003 | 0.003 |
| FVC | 0.851 | 0.012 | 0.010 | 0.803 | 0.016 | 0.013 |
| NPP | 0.874 | 1.018 | 0.811 | 0.886 | 0.841 | 0.743 |
| Riparian Vegetation Indicators | Accuracy of Calibration | Accuracy of Validation | ||||
|---|---|---|---|---|---|---|
| R2 | RMSE | MAE | R2 | RMSE | MAE | |
| kNDVI | 0.921 | 0.017 | 0.014 | 0.898 | 0.018 | 0.014 |
| FVC | 0.915 | 0.015 | 0.012 | 0.858 | 0.016 | 0.011 |
| NPP | 0.899 | 1.715 | 1.374 | 0.851 | 2.015 | 1.666 |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2025 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Yan, X.; Yang, W.; Pu, Z.; Zhang, Q.; Chen, Y.; Chen, J.; Xiang, W.; Chen, H.; Cheng, Y.; Zhao, Y. Responses of Typical Riparian Vegetation to Annual Variation of River Flow in a Semi-Arid Climate Region: Case Study of China’s Xiliao River. Land 2025, 14, 198. https://doi.org/10.3390/land14010198
Yan X, Yang W, Pu Z, Zhang Q, Chen Y, Chen J, Xiang W, Chen H, Cheng Y, Zhao Y. Responses of Typical Riparian Vegetation to Annual Variation of River Flow in a Semi-Arid Climate Region: Case Study of China’s Xiliao River. Land. 2025; 14(1):198. https://doi.org/10.3390/land14010198
Chicago/Turabian StyleYan, Xiangzhao, Wei Yang, Zaohong Pu, Qilong Zhang, Yutong Chen, Jiaqi Chen, Weiqi Xiang, Hongyu Chen, Yuyang Cheng, and Yanwei Zhao. 2025. "Responses of Typical Riparian Vegetation to Annual Variation of River Flow in a Semi-Arid Climate Region: Case Study of China’s Xiliao River" Land 14, no. 1: 198. https://doi.org/10.3390/land14010198
APA StyleYan, X., Yang, W., Pu, Z., Zhang, Q., Chen, Y., Chen, J., Xiang, W., Chen, H., Cheng, Y., & Zhao, Y. (2025). Responses of Typical Riparian Vegetation to Annual Variation of River Flow in a Semi-Arid Climate Region: Case Study of China’s Xiliao River. Land, 14(1), 198. https://doi.org/10.3390/land14010198