
Native or Exotic: A Bibliographical Review of the Debate on Ecological Science Methodologies: Valuable Lessons for Urban Green Space Design




{kind=link}
{kind=link}
{kind=link}
Abstract
:1. Introduction
1.1. Context
1.2. The Contribution of Ecological Sciences in UGS Design and Current Trends
- -
- What ways forward do ecological sciences propose in order to maximize the vegetation’s ecological potential when designing Urban Green Spaces?
- -
- What are the arguments presented to support exotic species as the answer to climate change-adaptive and -resilient ecosystems? To what extent can these arguments justify the disturbance of the native biodiversity and the loss of landscape identity?
- -
- Can native communities adapt to climate change and safeguard ecosystem function through time? Are they more resilient to climate change and more efficient at mitigating the subsequent effects?
2. Materials and Methods
3. Results and Discussion
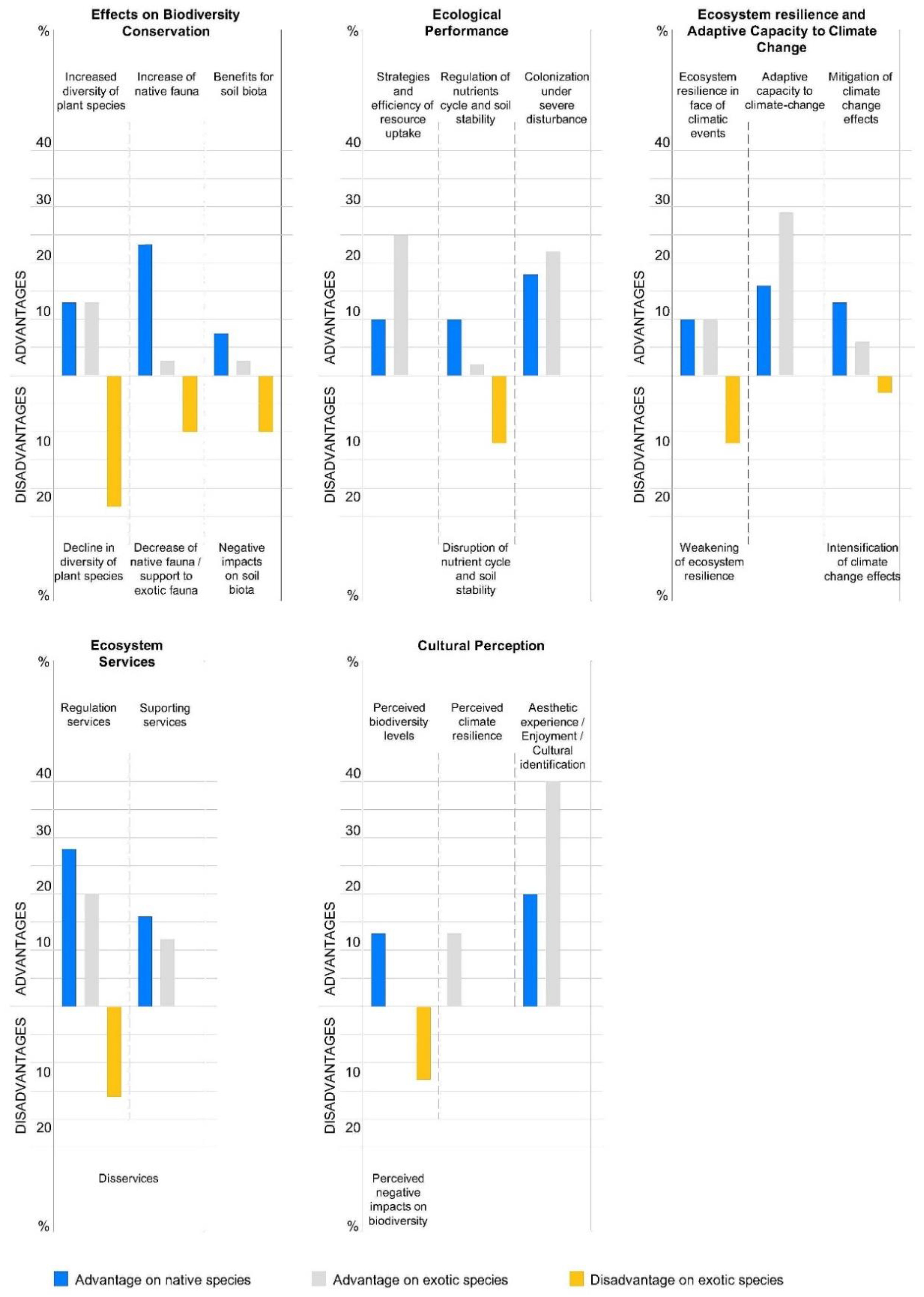
3.1. Effects on Biodiversity Conservation
3.2. Ecological Performance
3.3. Ecosystem Resilience and Adaptive Capacity to Climate Change
3.4. Ecosystem Services
3.5. Cultural Perception
4. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Grose, M.J. Gaps and futures in working between ecology and design for constructed ecologies. Landsc. Urban Plan. 2014, 132, 69–78. [Google Scholar] [CrossRef]
- Reimer, M.H. Unsettling eco-scapes: Aesthetic performances for sustainable futures. J. Landsc. Archit. 2010, 5, 24–37. [Google Scholar] [CrossRef]
- Pickett, S.T.A.; Cadenasso, M.L.; Grove, J.M.; Boone, C.G.; Groffman, P.M.; Irwin, E.; Kaushal, S.S.; Marshall, V.; McGrath, B.P.; Nilon, C.H.; et al. Urban ecological systems: Scientific foundations and a decade of progress. J. Environ. Manag. 2011, 92, 331–362. [Google Scholar] [CrossRef] [PubMed]
- Bryant, M.M. Urban landscape conservation and the role of ecological greenways at local and metropolitan scales. Landsc. Urban Plan. 2006, 76, 23–44. [Google Scholar] [CrossRef]
- Klaus, V.H.; Kiehl, K. A conceptual framework for urban ecological restoration and rehabilitation. Basic Appl. Ecol. 2021, 52, 82–94. [Google Scholar] [CrossRef]
- EEA. Building a coherent Trans-European Nature Network; Briefing no. 05/2020; European Environment Agency: Copenhagen, Denmark, 2020; p. 18. [Google Scholar]
- Yu, D.; Xun, B.; Shi, P.; Shao, H.; Liu, Y. Ecological restoration planning based on connectivity in an urban area. Ecol. Eng. 2012, 46, 24–33. [Google Scholar] [CrossRef]
- Yapp, G.; Walker, J.; Thackway, R. Linking vegetation type and condition to ecosystem goods and services. Ecol. Complex. 2010, 7, 292–301. [Google Scholar] [CrossRef]
- Carter, J.G.; Cavan, G.; Connelly, A.; Guy, S.; Handley, J.; Kazmierczak, A. Climate change and the city: Building capacity for urban adaptation. Prog. Plan. 2015, 95, 1–66. [Google Scholar] [CrossRef]
- Bark, R.H.; Osgood, D.E.; Colby, B.G.; Katz, G.; Stromberg, J. Habitat preservation and restoration: Do homebuyers have preferences for quality habitat? Ecol. Econ. 2009, 68, 1465–1475. [Google Scholar] [CrossRef]
- Evers, C.R.; Wardropper, C.B.; Branoff, B.; Granek, E.F.; Hirsch, S.L.; Link, T.E.; Olivero-Lora, S.; Wilson, C. The ecosystem services and biodiversity of novel ecosystems: A literature review. Glob. Ecol. Conserv. 2018, 13, e00362. [Google Scholar] [CrossRef]
- Higgs, E. Novel and designed ecosystems. Restor. Ecol. 2016, 25, 8–13. [Google Scholar] [CrossRef]
- Simonson, W.D.; Miller, E.; Jones, A.; García-Rangel, S.; Thornton, H.; McOwen, C. Enhancing climate change resilience of ecological restoration—A framework for action. Perspect. Ecol. Conserv. 2021, 19, 300–310. [Google Scholar] [CrossRef]
- Aquilué, N.; Messier, C.; Martins, K.T.; Dumais-Lalonde, V.; Mina, M. A simple-to-use management approach to boost adaptive capacity of forests to global uncertainty. For. Ecol. Manag. 2021, 481, 118692. [Google Scholar] [CrossRef]
- Esperon-Rodriguez, M.; Rymer, P.D.; Power, S.A.; Challis, A.; Prokopavicius, R.M.; Tjoelker, M.G. Functional adaptations and trait plasticity of urban trees along a climatic gradient. Urban For. Urban Green. 2020, 54, 126771. [Google Scholar] [CrossRef]
- Wood, S.L.R.; Dupras, J. Increasing functional diversity of the urban canopy for climate resilience: Potential tradeoffs with ecosystem services? Urban For. Urban Green. 2021, 58, 126972. [Google Scholar] [CrossRef]
- Ossola, A.; Lin, B.B. Making nature-based solutions climate-ready for the 50 °C world. Environ. Sci. Policy 2021, 123, 151–159. [Google Scholar] [CrossRef]
- Cardou, F.; Aubin, I.; Lapointe, M.; Shipley, B. Multifunctionality in practice: Measuring differences in urban woodland ecosystem properties via functional traits. Urban For. Urban Green. 2022, 68, 127453. [Google Scholar] [CrossRef]
- Berthon, K.; Thomas, F.; Bekessy, S. The role of ‘nativeness’ in urban greening to support animal biodiversity. Landsc. Urban Plan. 2021, 205, 103959. [Google Scholar] [CrossRef]
- Castro-Díez, P.; Alonso, Á.; Saldaña-López, A.; Granda, E. Effects of widespread non-native trees on regulating ecosystem services. Sci. Total Environ. 2021, 778, 146141. [Google Scholar] [CrossRef]
- Riley, C.B.; Herms, D.A.; Gardiner, M.M. Exotic trees contribute to urban forest diversity and ecosystem services in inner-city Cleveland, OH. Urban For. Urban Green. 2018, 29, 367–376. [Google Scholar] [CrossRef]
- Milanović, M.; Knapp, S.; Pyšek, P.; Kühn, I. Linking traits of invasive plants with ecosystem services and disservices. Ecosyst. Serv. 2020, 42, 101072. [Google Scholar] [CrossRef]
- Kumar Rai, P.; Singh, J.S. Invasive alien plant species: Their impact on environment, ecosystem services and human health. Ecol. Indic. 2020, 111, 106020. [Google Scholar] [CrossRef] [PubMed]
- Jarić, I.; Heger, T.; Castro Monzon, F.; Jeschke, J.M.; Kowarik, I.; McConkey, K.R.; Pyšek, P.; Sagouis, A.; Essl, F. Crypticity in Biological Invasions. Trends Ecol. Evol. 2019, 34, 291–302. [Google Scholar] [CrossRef] [PubMed]
- Turbelin, A.; Catford, J.A. Chapter 25—Invasive plants and climate change. In Climate Change, 3rd ed.; Letcher, T.M., Ed.; Elsevier: Amsterdam, The Netherlands, 2021; pp. 515–539. ISBN 978-0-12-821575-3. [Google Scholar]
- Simmons, M.; Venhaus, H.C.; Windhager, S. Exploiting the attributes of regional ecosystems for landscape design: The role of ecological restoration in ecological engineering. Ecol. Eng. 2007, 30, 201–205. [Google Scholar] [CrossRef]
- Olden, J.D.; LeRoy Poff, N.; Douglas, M.R.; Douglas, M.E.; Fausch, K.D. Ecological and evolutionary consequences of biotic homogenization. Trends Ecol. Evol. 2004, 19, 18–24. [Google Scholar] [CrossRef] [PubMed]
- Moher, D.; Liberati, A.; Tetzlaff, J.; Altman, D.G. Preferred reporting items for systematic reviews and meta-analyses: The PRISMA statement. BMJ 2009, 339, b2535. [Google Scholar] [CrossRef] [Green Version]
- Sun, F.; Zeng, L.; Cai, M.; Chauvat, M.; Forey, E.; Tariq, A.; Graciano, C.; Zhang, Z.; Gu, Y.; Zeng, F.; et al. An invasive and native plant differ in their effects on the soil food-web and plant-soil phosphorus cycle. Geoderma 2022, 410, 115672. [Google Scholar] [CrossRef]
- Raymundo, M.; Pastore, A.; Hille Ris Lambers, J.; Mayfield, M.M. Annual rainfall variation and dispersal limitation combine to alter invaded plant community diversity, dominance hierarchies and seeding phenology. Clim. Chang. Ecol. 2021, 2, 100024. [Google Scholar] [CrossRef]
- Barros, V.; Melo, A.; Santos, M.; Nogueira, L.; Frosi, G.; Santos, M.G. Different resource-use strategies of invasive and native woody species from a seasonally dry tropical forest under drought stress and recovery. Plant Physiol. Biochem. 2020, 147, 181–190. [Google Scholar] [CrossRef]
- Esquivel, J.; Echeverría, C.; Saldaña, A.; Fuentes, R. High functional diversity of forest ecosystems is linked to high provision of water flow regulation ecosystem service. Ecol. Indic. 2020, 115, 106433. [Google Scholar] [CrossRef]
- Shouman, S.; Mason, N.; Heberling, J.M.; Kichey, T.; Closset-Kopp, D.; Kobeissi, A.; Decocq, G. Leaf functional traits at home and abroad: A community perspective of sycamore maple invasion. For. Ecol. Manag. 2020, 464, 118061. [Google Scholar] [CrossRef]
- Díaz-Barradas, M.C.; Gallego-Fernández, J.B.; Zunzunegui, M. Plant response to water stress of native and non-native Oenothera drummondii populations. Plant Physiol. Biochem. 2020, 154, 219–228. [Google Scholar] [CrossRef] [PubMed]
- Zhang, Y.; Yu, C.; Xie, J.; Du, S.; Feng, J.; Guan, D. Comparison of fine root biomass and soil organic carbon stock between exotic and native mangrove. Catena 2021, 204, 105423. [Google Scholar] [CrossRef]
- Meira Castro, A.C.; Nunes, A.; Sousa, A.; Lourenço, L. Mapping the Causes of Forest Fires in Portugal by Clustering Analysis. Geosciences 2020, 10, 53. [Google Scholar] [CrossRef] [Green Version]
- Fogliata, P.; Cislaghi, A.; Sala, P.; Giupponi, L. An ecological analysis of the riparian vegetation for improving the riverine ecosystem management: The case of Lombardy region (North Italy). Landsc. Ecol. Eng. 2021, 17, 375–386. [Google Scholar] [CrossRef]
- Fang, J.; Deng, Y.; Che, R.; Han, C.; Zhong, W. Bacterial community composition in soils covered by different vegetation types in the Yancheng tidal marsh. Environ. Sci. Pollut. Res. 2020, 27, 21517–21532. [Google Scholar] [CrossRef] [PubMed]
- Tong, C.; Morris, J.T.; Huang, J.; Xu, H.; Wan, S. Changes in pore-water chemistry and methane emission following the invasion of Spartina alterniflora into an oliogohaline marsh. Limnol. Oceanogr. 2018, 63, 384–396. [Google Scholar] [CrossRef]
- Kotzen, B.; Branquinho, C.; Prasse, R. Does the exotic equal pollution? Landscape methods for solving the dilemma of using native versus non-native plant species in drylands. Land Degrad. Dev. 2020, 31, 2925–2935. [Google Scholar] [CrossRef]
- Gastauer, M.; Ramos, S.J.; Caldeira, C.F.; Siqueira, J.O. Reintroduction of native plants indicates the return of ecosystem services after iron mining at the Urucum Massif. Ecosphere 2021, 12, e03762. [Google Scholar] [CrossRef]
- Xia, S.; Wang, W.; Song, Z.; Kuzyakov, Y.; Guo, L.; Van Zwieten, L.; Li, Q.; Hartley, I.P.; Yang, Y.; Wang, Y.; et al. Spartina alterniflora invasion controls organic carbon stocks in coastal marsh and mangrove soils across tropics and subtropics. Glob. Chang. Biol. 2021, 27, 1627–1644. [Google Scholar] [CrossRef] [PubMed]
- Gaggini, L.; Rusterholz, H.-P.; Baur, B. The invasive plant Impatiens glandulifera affects soil fungal diversity and the bacterial community in forests. Appl. Soil Ecol. 2018, 124, 335–343. [Google Scholar] [CrossRef]
- Dairel, M.; Fidelis, A. The presence of invasive grasses affects the soil seed bank composition and dynamics of both invaded and non-invaded areas of open savannas. J. Environ. Manag. 2020, 276, 111291. [Google Scholar] [CrossRef]
- Tan, P.Y.; Wong, N.H.; Tan, C.L.; Jusuf, S.K.; Schmiele, K.; Chiam, Z.Q. Transpiration and cooling potential of tropical urban trees from different native habitats. Sci. Total Environ. 2020, 705, 135764. [Google Scholar] [CrossRef]
- Pinho, P.; Casanelles-Abella, J.; Luz, A.C.; Kubicka, A.M.; Branquinho, C.; Laanisto, L.; Neuenkamp, L.; Alós Ortí, M.; Obrist, M.K.; Deguines, N.; et al. Research agenda on biodiversity and ecosystem functions and services in European cities. Basic Appl. Ecol. 2021, 53, 124–133. [Google Scholar] [CrossRef]
- Quijas, S.; Romero-Duque, L.P.; Trilleras, J.M.; Conti, G.; Kolb, M.; Brignone, E.; Dellafiore, C. Linking biodiversity, ecosystem services, and beneficiaries of tropical dry forests of Latin America: Review and new perspectives. Ecosyst. Serv. 2019, 36, 100909. [Google Scholar] [CrossRef]
- Van der Biest, K.; Meire, P.; Schellekens, T.; D’hondt, B.; Bonte, D.; Vanagt, T.; Ysebaert, T. Aligning biodiversity conservation and ecosystem services in spatial planning: Focus on ecosystem processes. Sci. Total Environ. 2020, 712, 136350. [Google Scholar] [CrossRef]
- Pires, A.; Amaral, A.G.; Padgurschi, M.; Joly, C.; Scarano, F. Biodiversity research still falls short of creating links with ecosystem services and human well-being in a global hotspot. Ecosyst. Serv. 2018, 34, 68–73. [Google Scholar] [CrossRef]
- Pandey, S.; Gurr, G.M. Conservation biological control using Australian native plants in a brassica crop system: Seeking complementary ecosystem services. Agric. Ecosyst. Environ. 2019, 280, 77–84. [Google Scholar] [CrossRef]
- Barrico, L.; Castro, H.; Coutinho, A.; Gonçalves, M.T.; Freitas, H.; Castro, P. Plant and microbial biodiversity in urban forests and public gardens: Insights for cities’ sustainable development. Urban For. Urban Green. 2018, 29, 19–27. [Google Scholar] [CrossRef]
- Prendergast, K.S.; Tomlinson, S.; Dixon, K.W.; Bateman, P.W.; Menz, M.H.M. Urban native vegetation remnants support more diverse native bee communities than residential gardens in Australia’s southwest biodiversity hotspot. Biol. Conserv. 2022, 265, 109408. [Google Scholar] [CrossRef]
- Thaweepworadej, P.; Evans, K.L. Species richness and ecosystem services of tree assemblages along an urbanisation gradient in a tropical mega-city: Consequences for urban design. Urban For. Urban Green. 2022, 70, 127527. [Google Scholar] [CrossRef]
- Campbell, J.W.; Grodsky, S.M.; Monroe, A.P.; Martin, J.A. Bee (Apoidea) community response to perennial grass treatments managed for livestock production and conservation. Agric. Ecosyst. Environ. 2021, 313, 107391. [Google Scholar] [CrossRef]
- Olupot, W. The potential of natural succession to restore degraded areas of a Ugandan rainforest dominated by the exotic paper mulberry Broussonetia papyrifera (L.) L’Hér. ex Vent. For. Ecol. Manag. 2022, 504, 119861. [Google Scholar] [CrossRef]
- Gallinat, A.S.; Primack, R.B.; Lloyd-Evans, T.L. Can invasive species replace native species as a resource for birds under climate change? A case study on bird-fruit interactions. Biol. Conserv. 2020, 241, 108268. [Google Scholar] [CrossRef]
- Machar, I.; Šimek, P.; Schlossárek, M.; Pechanec, V.; Petrovič, F.; Brus, J.; Špinlerová, Z.; Seják, J. Comparison of bird diversity between temperate floodplain forests and urban parks. Urban For. Urban Green. 2022, 67, 127427. [Google Scholar] [CrossRef]
- Jimenez, M.F.; Pejchar, L.; Reed, S.E.; McHale, M.R. The efficacy of urban habitat enhancement programs for conserving native plants and human-sensitive animals. Landsc. Urban Plan. 2022, 220, 104356. [Google Scholar] [CrossRef]
- Monberg, R.J.; Howe, A.G.; Kepfer-Rojas, S.; Ravn, H.P.; Jensen, M.B. Vegetation development in a stormwater management system designed to enhance ecological qualities. Urban For. Urban Green. 2019, 46, 126463. [Google Scholar] [CrossRef]
- Scotton, M.; Andreatta, D. Anti-erosion rehabilitation: Effects of revegetation method and site traits on introduced and native plant cover and richness. Sci. Total Environ. 2021, 776, 145915. [Google Scholar] [CrossRef]
- Curado, G.; Gallego-Tévar, B.; Figueroa, E.; Castillo, J.M. Effects of removal of alien Spartina densiflora and restoration of native S. maritima on succession and zonation in European salt marshes. Estuar. Coast. Shelf Sci. 2020, 244, 105815. [Google Scholar] [CrossRef]
- Phillips, M.L.; Aronson, E.L.; Maltz, M.R.; Allen, E.B. Native and invasive inoculation sources modify fungal community assembly and biomass production of a chaparral shrub. Appl. Soil. Ecol. 2020, 147, 103370. [Google Scholar] [CrossRef]
- Champness, B.S.; Palmer, G.C.; Fitzsimons, J.A. Bringing the city to the country: Relationships between streetscape vegetation type and bird assemblages in a major regional centre. J. Urban Ecol. 2019, 5, juz018. [Google Scholar] [CrossRef]
- Mody, K.; Lerch, D.; Müller, A.-K.; Simons, N.K.; Blüthgen, N.; Harnisch, M. Flower power in the city: Replacing roadside shrubs by wildflower meadows increases insect numbers and reduces maintenance costs. PLoS ONE 2020, 15, e0234327. [Google Scholar] [CrossRef] [PubMed]
- Gallego-Tévar, B.; Grewell, B.J.; Figueroa, E.; Castillo, J.M. The role of exotic and native hybrids during ecological succession in salt marshes. J. Exp. Mar. Biol. Ecol. 2020, 523, 151282. [Google Scholar] [CrossRef]
- Zunzunegui, M.; Ruiz-Valdepeñas, E.; Sert, M.A.; Díaz-Barradas, M.C.; Gallego-Fernández, J.B. Field comparison of ecophysiological traits between an invader and a native species in a Mediterranean coastal dune. Plant Physiol. Biochem. 2020, 146, 278–286. [Google Scholar] [CrossRef] [PubMed]
- de Vos, D.; Nel, R.; Schoeman, D.; Harris, L.R.; du Preez, D. Effect of introduced Casuarina trees on the vulnerability of sea turtle nesting beaches to erosion. Estuar. Coast. Shelf Sci. 2019, 223, 147–158. [Google Scholar] [CrossRef]
- González-Montelongo, C.; Pérez-Vargas, I. Is an invasive alien tree able to sustain a similar lichen diversity as the native forest? The case of the sweet chestnut (Castanea sativa Mill.) and the laurel forest in Macaronesia. For. Ecol. Manag. 2021, 488, 119009. [Google Scholar] [CrossRef]
- Oduor, A.M.O.; Long, H.; Fandohan, A.B.; Liu, J.; Yu, X. An invasive plant provides refuge to native plant species in an intensely grazed ecosystem. Biol. Invasions 2018, 20, 2745–2751. [Google Scholar] [CrossRef]
- Martins, P.L.S.S.; Furtado, S.G.; Menini Neto, L. Could epiphytes be xenophobic? Evaluating the use of native versus exotic phorophytes by the vascular epiphytic community in an urban environment. Community Ecol. 2020, 21, 91–101. [Google Scholar] [CrossRef]
- Randriambanona, H.; Randriamalala, J.R.; Carrière, S.M. Native forest regeneration and vegetation dynamics in non-native Pinus patula tree plantations in Madagascar. For. Ecol. Manag. 2019, 446, 20–28. [Google Scholar] [CrossRef]
- Gonçalves, P.; Vierikko, K.; Elands, B.; Haase, D.; Catarina Luz, A.; Santos-Reis, M. Biocultural diversity in an urban context: An indicator-based decision support tool to guide the planning and management of green infrastructure. Environ. Sustain. Indic. 2021, 11, 100131. [Google Scholar] [CrossRef]
- Christensen, B.; Cashmore, P.; Crump, S.; Hobbs, J. Fire disturbance favours exotic species at Kaituna Wetland, Bay of Plenty. N. Z. J. Ecol. 2019, 43, 3369. [Google Scholar] [CrossRef]
- Cáceres, N.; Robbiati, F.O.; Hick, E.C.; Suárez, M.; Matoff, E.; Galetto, L.; Imhof, L. Analysis of biodiversity attributes for extensive vegetated roofs in a semiarid region of central Argentina. Ecol. Eng. 2022, 178, 106602. [Google Scholar] [CrossRef]
- Rey, F.; Bifulco, C.; Bischetti, G.B.; Bourrier, F.; De Cesare, G.; Florineth, F.; Graf, F.; Marden, M.; Mickovski, S.B.; Phillips, C.; et al. Soil and water bioengineering: Practice and research needs for reconciling natural hazard control and ecological restoration. Sci. Total Environ. 2019, 648, 1210–1218. [Google Scholar] [CrossRef]
- Pérez-Corona, M.E.; Pérez-Hernández, M.d.C.; Medina-Villar, S.; Andivia, E.; Bermúdez de Castro, F. Canopy species composition drives seasonal soil characteristics in a Mediterranean riparian forest. Eur. J. For. Res. 2021, 140, 1081–1093. [Google Scholar] [CrossRef]
- Vasquez-Valderrama, M.; González-M, R.; López-Camacho, R.; Baptiste, M.P.; Salgado-Negret, B. Impact of invasive species on soil hydraulic properties: Importance of functional traits. Biol. Invasions 2020, 22, 1849–1863. [Google Scholar] [CrossRef]
- Eastburn, D.J.; Roche, L.M.; Doran, M.P.; Blake, P.R.; Bouril, C.S.; Gamble, G.; Gornish, E.S. Seeding plants for long-term multiple ecosystem service goals. J. Environ. Manag. 2018, 211, 191–197. [Google Scholar] [CrossRef] [PubMed]
- Gao, X.; Zhao, X.; Li, H.; Guo, L.; Lv, T.; Wu, P. Exotic shrub species (Caragana korshinskii) is more resistant to extreme natural drought than native species (Artemisia gmelinii) in a semiarid revegetated ecosystem. Agric. For. Meteorol. 2018, 263, 207–216. [Google Scholar] [CrossRef]
- Dagenais, D.; Brisson, J.; Fletcher, T.D. The role of plants in bioretention systems; does the science underpin current guidance? Ecol. Eng. 2018, 120, 532–545. [Google Scholar] [CrossRef]
- Wang, J.; Fu, B.; Lu, N.; Wang, S.; Zhang, L. Water use characteristics of native and exotic shrub species in the semi-arid Loess Plateau using an isotope technique. Agric. Ecosyst. Environ. 2019, 276, 55–63. [Google Scholar] [CrossRef]
- Miller, D.L.; Alonzo, M.; Roberts, D.A.; Tague, C.L.; McFadden, J.P. Drought response of urban trees and turfgrass using airborne imaging spectroscopy. Remote Sens. Environ. 2020, 240, 111646. [Google Scholar] [CrossRef]
- Robbiati, F.O.; Cáceres, N.; Hick, E.C.; Suarez, M.; Soto, S.; Barea, G.; Matoff, E.; Galetto, L.; Imhof, L. Vegetative and thermal performance of an extensive vegetated roof located in the urban heat island of a semiarid region. Build. Environ. 2022, 212, 108791. [Google Scholar] [CrossRef]
- Esfahani, R.E.; Paço, T.A.; Martins, D.; Arsénio, P. Increasing the resistance of Mediterranean extensive green roofs by using native plants from old roofs and walls. Ecol. Eng. 2022, 178, 106576. [Google Scholar] [CrossRef]
- Sueltenfuss, J.P.; Cooper, D.J. A new approach for hydrologic performance standards in wetland mitigation. J. Environ. Manag. 2019, 231, 1154–1163. [Google Scholar] [CrossRef] [PubMed]
- Galleguillos, M.; Gimeno, F.; Puelma, C.; Zambrano-Bigiarini, M.; Lara, A.; Rojas, M. Disentangling the effect of future land use strategies and climate change on streamflow in a Mediterranean catchment dominated by tree plantations. J. Hydrol. 2021, 595, 126047. [Google Scholar] [CrossRef]
- Pang, C.-C.; Lo, W.-F.; Yan, R.W.-M.; Hau, B.C.-H. Plant community composition on landfill sites after multiple years of ecological restoration. Landsc. Res. 2020, 45, 458–469. [Google Scholar] [CrossRef]
- Guillen-Cruz, G.; Rodríguez-Sánchez, A.L.; Fernández-Luqueño, F.; Flores-Rentería, D. Influence of vegetation type on the ecosystem services provided by urban green areas in an arid zone of northern Mexico. Urban For. Urban Green. 2021, 62, 127135. [Google Scholar] [CrossRef]
- Singh, A.N.; Kumar, A. Ecological performances of exotic and native woody species on coal mine spoil in Indian dry tropical region. Ecol. Eng. 2022, 174, 106470. [Google Scholar] [CrossRef]
- Mughal, M.O.; Kubilay, A.; Fatichi, S.; Meili, N.; Carmeliet, J.; Edwards, P.; Burlando, P. Detailed investigation of vegetation effects on microclimate by means of computational fluid dynamics (CFD) in a tropical urban environment. Urban Clim. 2021, 39, 100939. [Google Scholar] [CrossRef]
- Ennos, R.; Cottrell, J.; Hall, J.; O’Brien, D. Is the introduction of novel exotic forest tree species a rational response to rapid environmental change?—A British perspective. For. Ecol. Manag. 2019, 432, 718–728. [Google Scholar] [CrossRef]
- Dudley, M.P.; Freeman, M.; Wenger, S.; Jackson, C.R.; Pringle, C.M. Rethinking foundation species in a changing world: The case for Rhododendron maximum as an emerging foundation species in shifting ecosystems of the southern Appalachians. For. Ecol. Manag. 2020, 472, 118240. [Google Scholar] [CrossRef]
- Li, S.-H.; Ge, Z.-M.; Xie, L.-N.; Chen, W.; Yuan, L.; Wang, D.-Q.; Li, X.-Z.; Zhang, L.-Q. Ecophysiological response of native and exotic salt marsh vegetation to waterlogging and salinity: Implications for the effects of sea-level rise. Sci. Rep. 2018, 8, 2441. [Google Scholar] [CrossRef] [PubMed]
- Guerrero, F.; Hernández, C.; Toledo, M.; Espinoza, L.; Carrasco, Y.; Arriagada, A.; Muñoz, A.; Taborga, L.; Bergmann, J.; Carmona, C. Leaf Thermal and Chemical Properties as Natural Drivers of Plant Flammability of Native and Exotic Tree Species of the Valparaíso Region, Chile. Int. J. Environ. Res. Public Health 2021, 18, 7191. [Google Scholar] [CrossRef] [PubMed]
- Blackhall, M.; Raffaele, E. Flammability of Patagonian invaders and natives: When exotic plant species affect live fine fuel ignitability in wildland-urban interfaces. Landsc. Urban Plan. 2019, 189, 1–10. [Google Scholar] [CrossRef]
- Abbas, S.; Nichol, J.E.; Fischer, G.A.; Wong, M.S.; Irteza, S.M. Impact assessment of a super-typhoon on Hong Kong’s secondary vegetation and recommendations for restoration of resilience in the forest succession. Agric. For. Meteorol. 2020, 280, 107784. [Google Scholar] [CrossRef]
- Wilsey, B.J.; Martin, L.M.; Kaul, A.D. Phenology differences between native and novel exotic-dominated grasslands rival the effects of climate change. J. Appl. Ecol. 2018, 55, 863–873. [Google Scholar] [CrossRef] [Green Version]
- Matesanz, S.; Valladares, F. Ecological and evolutionary responses of Mediterranean plants to global change. Environ. Exp. Bot. 2014, 103, 53–67. [Google Scholar] [CrossRef] [Green Version]
- Rice, K.J.; Emery, N.C. Managing microevolution: Restoration in the face of global change. Front. Ecol. Environ. 2003, 1, 469–478. [Google Scholar] [CrossRef]
- Azeñas, V.; Janner, I.; Medrano, H.; Gulías, J. Performance evaluation of five Mediterranean species to optimize ecosystem services of green roofs under water-limited conditions. J. Environ. Manag. 2018, 212, 236–247. [Google Scholar] [CrossRef]
- Volaire, F. Plant traits and functional types to characterise drought survival of pluri-specific perennial herbaceous swards in Mediterranean areas. Eur. J. Agron. 2008, 2–3, 116–124. [Google Scholar] [CrossRef]
- Garnier, E.; Vile, D.; Roumet, C.; Lavorel, S.; Grigulis, K.; Navas, M.-L.; Lloret, F. Inter- and intra-specific trait shifts among sites differing in drought conditions at the north western edge of the Mediterranean Region. Flora 2019, 254, 147–160. [Google Scholar] [CrossRef]
- Ferrier, S.; Harwood, T.D.; Ware, C.; Hoskins, A.J. A globally applicable indicator of the capacity of terrestrial ecosystems to retain biological diversity under climate change: The bioclimatic ecosystem resilience index. Ecol. Indic. 2020, 117, 106554. [Google Scholar] [CrossRef]
- Safford, H.D.; Vallejo, V.R. Chapter 12—Ecosystem management and ecological restoration in the Anthropocene: Integrating global change, soils, and disturbance in boreal and Mediterranean forests. In Developments in Soil Science; Busse, M., Giardina, C.P., Morris, D.M., Page-Dumroese, D.S., Eds.; Global Change and Forest Soils; Elsevier: Amsterdam, The Netherlands, 2019; Volume 36, pp. 259–308. [Google Scholar]
- Maxwell, S.L.; Reside, A.; Trezise, J.; McAlpine, C.A.; Watson, J.E. Retention and restoration priorities for climate adaptation in a multi-use landscape. Glob. Ecol. Conserv. 2019, 18, e00649. [Google Scholar] [CrossRef]
- Jensen, D.A.; Rao, M.; Zhang, J.; Grøn, M.; Tian, S.; Ma, K.; Svenning, J.-C. The potential for using rare, native species in reforestation– A case study of yews (Taxaceae) in China. For. Ecol. Manag. 2021, 482, 118816. [Google Scholar] [CrossRef]
- Bussotti, F.; Pollastrini, M.; Holland, V.; Brüggemann, W. Functional traits and adaptive capacity of European forests to climate change. Environ. Exp. Bot. 2015, 111, 91–113. [Google Scholar] [CrossRef]
- Hamberg, L.J.; Fraser, R.A.; Robinson, D.T.; Trant, A.J.; Murphy, S.D. Surface temperature as an indicator of plant species diversity and restoration in oak woodland. Ecol. Indic. 2020, 113, 106249. [Google Scholar] [CrossRef]
- Vivian, L.M.; Greet, J.; Jones, C.S. Responses of grasses to experimental submergence in summer: Implications for the management of unseasonal flows in regulated rivers. Aquat. Ecol. 2020, 54, 985–999. [Google Scholar] [CrossRef]
- Filgueiras, B.K.C.; Peres, C.A.; Melo, F.P.L.; Leal, I.R.; Tabarelli, M. Winner–Loser Species Replacements in Human-Modified Landscapes. Trends Ecol. Evol. 2021, 36, 545–555. [Google Scholar] [CrossRef]
- Teixeira, H.; Lillebø, A.I.; Culhane, F.; Robinson, L.; Trauner, D.; Borgwardt, F.; Kuemmerlen, M.; Barbosa, A.; McDonald, H.; Funk, A.; et al. Linking biodiversity to ecosystem services supply: Patterns across aquatic ecosystems. Sci. Total Environ. 2019, 657, 517–534. [Google Scholar] [CrossRef]
- Nsenga Kumwimba, M.; Dzakpasu, M.; Li, X. Potential of invasive watermilfoil (Myriophyllum spp.) to remediate eutrophic waterbodies with organic and inorganic pollutants. J. Environ. Manag. 2020, 270, 110919. [Google Scholar] [CrossRef]
- Kuras, E.R.; Warren, P.S.; Zinda, J.A.; Aronson, M.F.J.; Cilliers, S.; Goddard, M.A.; Nilon, C.H.; Winkler, R. Urban socioeconomic inequality and biodiversity often converge, but not always: A global meta-analysis. Landsc. Urban Plan. 2020, 198, 103799. [Google Scholar] [CrossRef]
- Vaz, A.S.; Castro-Díez, P.; Godoy, O.; Alonso, Á.; Vilà, M.; Saldaña, A.; Marchante, H.; Bayón, Á.; Silva, J.S.; Vicente, J.R.; et al. An indicator-based approach to analyse the effects of non-native tree species on multiple cultural ecosystem services. Ecol. Indic. 2018, 85, 48–56. [Google Scholar] [CrossRef]
- Scopelliti, M.; Carrus, G.; Adinolfi, C.; Suarez, G.; Colangelo, G.; Lafortezza, R.; Panno, A.; Sanesi, G. Staying in touch with nature and well-being in different income groups: The experience of urban parks in Bogotá. Landsc. Urban Plan. 2016, 148, 139–148. [Google Scholar] [CrossRef]
- Ward, S.G.; Amatangelo, K.L. Suburban gardening in Rochester, New York: Exotic plant preference and risk of invasion. Landsc. Urban Plan. 2018, 180, 161–165. [Google Scholar] [CrossRef]
- Hoyle, H.E. Climate-adapted, traditional or cottage-garden planting? Public perceptions, values and socio-cultural drivers in a designed garden setting. Urban For. Urban Green. 2021, 65, 127362. [Google Scholar] [CrossRef]
- Hoyle, H.; Hitchmough, J.; Jorgensen, A. Attractive, climate-adapted and sustainable? Public perception of non-native planting in the designed urban landscape. Landsc. Urban Plan. 2017, 164, 49–63. [Google Scholar] [CrossRef]
- Shackleton, S.E.; Shackleton, R.T. Local knowledge regarding ecosystem services and disservices from invasive alien plants in the arid Kalahari, South Africa. J. Arid Environ. 2018, 159, 22–33. [Google Scholar] [CrossRef]
- Alvarez, S.; Soto, J.R.; Escobedo, F.J.; Lai, J.; Kibria, A.S.M.G.; Adams, D.C. Heterogeneous preferences and economic values for urban forest structural and functional attributes. Landsc. Urban Plan. 2021, 215, 104234. [Google Scholar] [CrossRef]
- Vaz, A.S.; Gonçalves, J.F.; Pereira, P.; Santarém, F.; Vicente, J.R.; Honrado, J.P. Earth observation and social media: Evaluating the spatiotemporal contribution of non-native trees to cultural ecosystem services. Remote Sens. Environ. 2019, 230, 111193. [Google Scholar] [CrossRef]
- Lampinen, J.; Tuomi, M.; Fischer, L.K.; Neuenkamp, L.; Alday, J.G.; Bucharova, A.; Cancellieri, L.; Casado-Arzuaga, I.; Čeplová, N.; Cerveró, L.; et al. Acceptance of near-natural greenspace management relates to ecological and socio-cultural assigned values among European urbanites. Basic Appl. Ecol. 2021, 50, 119–131. [Google Scholar] [CrossRef]
- Threlfall, C.G.; Kendal, D. The distinct ecological and social roles that wild spaces play in urban ecosystems. Urban For. Urban Green. 2018, 29, 348–356. [Google Scholar] [CrossRef]
- Young, C.; Hofmann, M.; Frey, D.; Moretti, M.; Bauer, N. Psychological restoration in urban gardens related to garden type, biodiversity and garden-related stress. Landsc. Urban Plan. 2020, 198, 103777. [Google Scholar] [CrossRef]
- Fischer, L.K.; Honold, J.; Cvejić, R.; Delshammar, T.; Hilbert, S.; Lafortezza, R.; Nastran, M.; Nielsen, A.B.; Pintar, M.; van der Jagt, A.P.N.; et al. Beyond green: Broad support for biodiversity in multicultural European cities. Glob. Environ. Chang. 2018, 49, 35–45. [Google Scholar] [CrossRef]
- Shaw, A.; Miller, K.K.; Wescott, G. Australian native gardens: Is there scope for a community shift? Landsc. Urban Plan. 2017, 157, 322–330. [Google Scholar] [CrossRef]
- Stępniewska, M. The capacity of urban parks for providing regulating and cultural ecosystem services versus their social perception. Land Use Policy 2021, 111, 105778. [Google Scholar] [CrossRef]



Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
de Carvalho, C.A.; Raposo, M.; Pinto-Gomes, C.; Matos, R. Native or Exotic: A Bibliographical Review of the Debate on Ecological Science Methodologies: Valuable Lessons for Urban Green Space Design. Land 2022, 11, 1201. https://doi.org/10.3390/land11081201
de Carvalho CA, Raposo M, Pinto-Gomes C, Matos R. Native or Exotic: A Bibliographical Review of the Debate on Ecological Science Methodologies: Valuable Lessons for Urban Green Space Design. Land. 2022; 11(8):1201. https://doi.org/10.3390/land11081201
Chicago/Turabian Stylede Carvalho, Catarina Archer, Mauro Raposo, Carlos Pinto-Gomes, and Rute Matos. 2022. "Native or Exotic: A Bibliographical Review of the Debate on Ecological Science Methodologies: Valuable Lessons for Urban Green Space Design" Land 11, no. 8: 1201. https://doi.org/10.3390/land11081201
APA Stylede Carvalho, C. A., Raposo, M., Pinto-Gomes, C., & Matos, R. (2022). Native or Exotic: A Bibliographical Review of the Debate on Ecological Science Methodologies: Valuable Lessons for Urban Green Space Design. Land, 11(8), 1201. https://doi.org/10.3390/land11081201