Abstract
Determining the availability and supply capacity of soil inorganic nitrogen (N) can effectively guide the appropriate application of N fertilizers during crop cultivation. However, the mechanism underlying soil inorganic N production remains unknown for cash crops in karst regions. In this study, the rates of organic N mineralization to ammonium (NH4+) and NH4+ nitrification to nitrate (NO3−) were determined using a 15N tracing technique to evaluate the supply capacity of inorganic N in soils from woodland and pitaya plantations with different cultivation years (3, 9, and 15 years) in the subtropical karst region of China. The conversion of woodland to pitaya plantations significantly decreased the content of soil organic carbon (SOC), total N, calcium (Ca), and magnesium (Mg), along with the soil pH and cation-exchange capacity (CEC), but significantly increased the content of available potassium, available phosphorus, iron, and aluminum, in a more pronounced fashion with the increasing length of pitaya cultivation. The conversion of woodland to pitaya plantations has not significantly changed soil NH4+ and NO3− content, but this land use has resulted in divergent effects on mineralization and nitrification rates. Compared to woodland (5.49 mg N kg−1 d−1), pitaya cultivation significantly reduced the mineralization rate to 0.62–2.38 mg N kg−1 d−1. Conversely, the nitrification rate significantly increased from 4.71 mg N kg−1 d−1 in soil under woodland to 9.32 mg N kg−1 d−1 in soil under 3-year pitaya cultivation, but this rate decreased to 1.74 mg N kg−1 d−1 under 15-year cultivation. Furthermore, the mean residence time of inorganic N was significantly higher in long-term than in short-term pitaya plantations, indicating the decline in inorganic N turnover with the increasing length of pitaya cultivation. Taken together, long-term pitaya cultivation could significantly decrease the supply capacity and turnover of inorganic N in soil. The Ca, Mg, SOC, and total N content, as well as CEC, were significantly and positively related to the mineralization rate, but negatively related to the mean residence time of NH4+ and NO3−, suggesting that the incorporation of organic matter can accelerate the soil inorganic N supply and turnover for long-term pitaya plantation in subtropical regions.
1. Introduction
Karst landforms developed from carbonate rock are widely distributed in China and occupy an area of approximately 3.44 million km2 [1]. Due to their geological specificity, limited arable land resources exist in karst regions. Thus, sloped woodland or shrubland is often deforested to cultivate common crops (e.g., beans, corn). However, in addition to low economic income, this cropping pattern inevitably induces some negative environmental and ecological effects, such as large-scale “rocky desertification” in karst regions [1,2]. Thus, the cultivation of high-yield and economically sustainable crops is urgently needed in order to promote local farmers’ income. Due to inherent ecological benefits (e.g., barren resistance, easy growth, soil, and water conservation) and high economic costs, pitaya (Hylocereus undatus) has been introduced and planted in the subtropical karst regions of southwest China [3]. Statistical data show that the pitaya cultivation area in China has increased from approximately 3 × 103 ha in 2011 to 4 × 104 ha in 2017 [4]. Noticeably, the slow growth and low yield of pitaya frequently hinder the sustainable development of this crop [5,6]. In addition to climate factors and management practices, soil fertility is a critical factor affecting pitaya productivity.
As the key indicator of soil fertility, nitrogen (N) is an important nutrient element that limits crop growth and yield [7,8]. With the exception of small-molecule organic N, ammonium (NH4+) and nitrate (NO3−) are the main forms of N for crop uptake [9,10]. In previous studies, NH4+ and NO3− content have been widely measured to assess the availability of N, which can indicate whether the available N supply in soil satisfies the growing demand of crops [11,12]. However, this method cannot differentiate the mechanisms underlying inorganic N production [13,14]. In soil, inorganic N needs to be produced through the mineralization of organic N to NH4+, and the subsequent nitrification of NH4+ to NO3− [14,15]. Therefore, the supply process of inorganic N in soil can be clearly quantified by the determination of mineralization and nitrification, which is of particular importance for proposing appropriate fertilizer management for pitaya cultivation. However, the intensity of mineralization and nitrification, along with the affecting factors in subtropical karst regions, remains unknown.
In karst regions, the soils are characterized by high pH, calcium (Ca), and magnesium (Mg) [16,17]. In the undisturbed natural ecosystems (e.g., woodland) of karst regions, high Ca levels can stimulate the accumulation of soil organic matter, which may result in high mineralization and nitrification rates in the soil [18,19]. When woodland is converted to pitaya plantations, the mineralization and nitrification processes may be significantly altered due to the changes in management practices (e.g., tillage and fertilization) and soil properties. The conversion of woodlands to pitaya plantations can greatly decrease organic N content if organic fertilizer cannot be applied [20,21]. If this is the case, pitaya cultivation may decrease the mineralization in the soil due to the decrease in organic N. On the other hand, the intensive application of N fertilizer, combined with tillage, may stimulate the nitrification of NH4+ to NO3− by increasing the abundance or activity of ammonia-oxidizing archaea (AOA) and/or bacteria (AOB) [22,23,24]. Noticeably, such stimulatory effects may be inhibited with the increasing duration of pitaya cultivation, because long-term crop cultivation can significantly decrease soil organic matter, causing the formation of a heavy clay texture. This may inhibit mineralization and nitrification rates, reducing the supply capacity and turnover of inorganic N in soil [19,21], even when large amounts of mineral N fertilizer are applied to pitaya plantations. Thus, we hypothesized that long-term pitaya cultivation would decrease the supply capacity and turnover of inorganic N by reducing mineralization and nitrification rates in karst regions.
To verify this hypothesis, woodland and pitaya plantations with different cultivation durations were chosen from the subtropical karst region of southwest China, and the mineralization and nitrification rates in soils were determined using the 15N tracing technique. The objective of this study was to clarify the changes in inorganic N supply capacity and turnover in soils, along with the influencing factors.
2. Materials and Methods
2.1. Sites and Soil Sample Collections
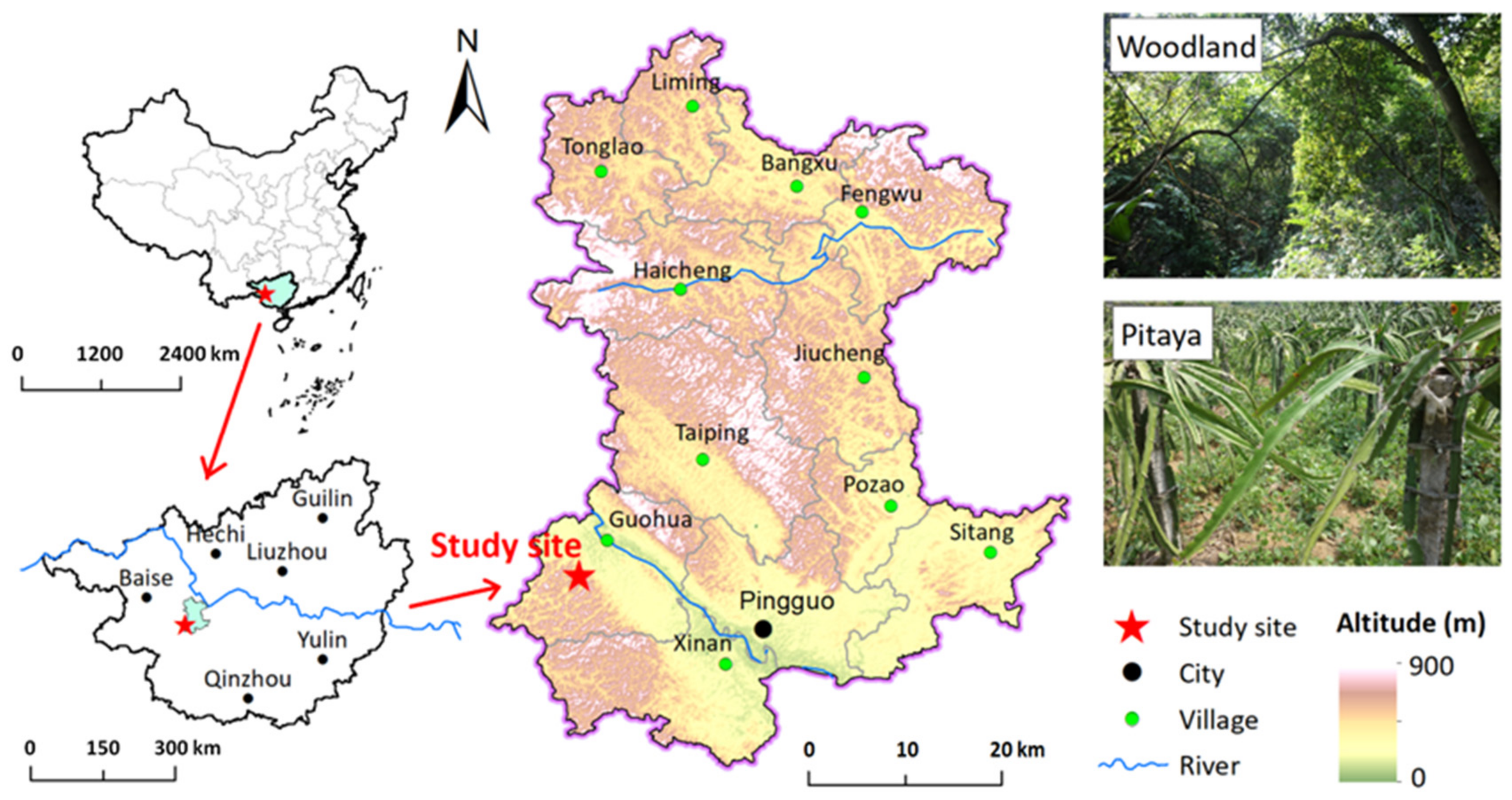
The study site was located in the Guohua karst experimental site (23°22′ N, 107°23′ E), Pingguo County, Guangxi Province, southwest China (Figure 1). The study region has a subtropical monsoon climate, with an annual average temperature of approximately 19–21.5 °C, annual average evaporation of 1572 mm, and relative humidity of above 80%. The soil type was classified as Alfisol developed from limestone according to the USDA soil taxonomy. Most of the peasants in this region live in mountainous karst regions with poor crop production and living conditions. Local people can only grow plants on slope lands that have been deforested, resulting in rocky desertification problems. To promote the restoration of barren hill vegetation, reduce water and soil loss, and increase farmers’ income, pitaya was introduced in this area in 2003.
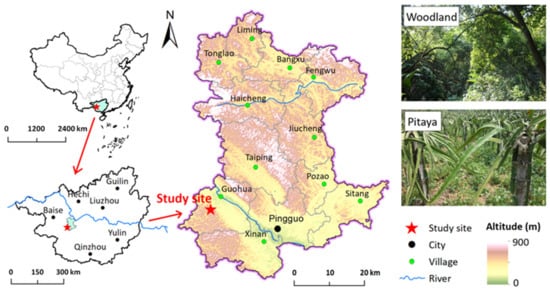
Figure 1.
Schematic diagram of the study site.
Three pitaya plantations with different cultivation times (3, 9, and 15 years) were chosen, all of which had been converted from woodland, and the adjacent woodland was also chosen as a control. Both woodland and pitaya plantations were on land steeper than 10° at an elevation of approximately 410 m asl. The main tree species at woodland sites include Zenia insignis, Melia azedarach, Apodytes dimidiate, and Choerospondias axillaris, accompanied by shrubs including Alchornea trewioides, Cipadessa cinerascens, and Vitex negundo. The ground cover is occupied by grass communities dominated by Nephrolepis auriculata, Eupatorium odoratum, Microstegium gratum, and Cymbopogon caesius. Four pitaya per pillar were arranged, maintaining 3 m × 3 m spacing. Approximately 165, 136, 145, and 12,210 kg ha−1 y−1 of N, P2O5, K2O, and sheep manure, respectively, were applied to the pitaya fields. The N, potassium (K), and phosphorus (P) content of the sheep manure were 6.5, 5.0, and 3.0 g kg−1, respectively.
Three representative sites (50 m × 50 m) were randomly established as the spatial replication for the woodland and each pitaya plantation, and the distance between each site was over 50 m. At each site, five plots with 20 m intervals between them were staked out. After removing the litter of topsoil, two soil cores that were 5 cm in diameter were set up from the 0–15 cm layer in each plot and, subsequently, all of the soil cores from the five plots were composited to form one sample per site. Consequently, there were 12 soil samples in total to be studied. Stones, plant roots, and residuals were removed from the sampled soils, which were then passed through a 2 mm sieve. Part of soil the samples was air-dried for the determination of the basic soil properties, and other fresh soils were used to determine the abundances of AOA and AOB, as well as the rates of mineralization and nitrification.
2.2. 15N Labelling Experiment
A 15N labelling experiment in the laboratory was performed according to a previously published method [18,21]. There were two 15N treatments (15NH4NO3 and NH415NO3), with three replicates for each treatment and two incubation times. A series of fresh soil (30 g of oven-dried basis) was weighed in 250 mL Erlenmeyer flasks and pre-incubated for 24 h in darkness at 25 °C. After the pre-incubation, the 15NH4NO3 or NH415NO3 (10 atom% excess) solutions were evenly added to the soil surface of each flask, and the application rate of NH4NO3 reached 50 mg NH4+–N (kg dry soil)−1 and 50 mg NO3−–N (kg dry soil)−1. Subsequently, distilled water was applied to adjust the soil moisture to 60% water-holding capacity (WHC). All flasks were sealed with plastic film containing small holes, and then incubated in darkness again at 25 °C. The soils in the flasks were extracted with 150 mL of 2 M KCl solution at 0.5 h and 24 h after the application of NH4NO3. The KCl extracts were used to determine the content and 15N atom% excess of NH4+ and NO3− with a continuous-flow analyzer (Skalar, Breda, The Netherlands) and a Sercon Integra2 isotope ratio mass spectrometer (Sercon Ltd., Crewe, UK), respectively. The detailed information on the preparation of 15N samples of NH4+ and NO3− pools was presented by Xie et al. (2018) [14] and Garousi et al. (2021) [19].
2.3. Analyses of Soil Properties
The soil organic carbon (SOC) and total N were determined using a Sercon Integra2 elemental analyzer, when inorganic C in soil was removed with a diluted phosphoric acid solution. Soil pH was determined via extraction with a 1:2.5 (w:v) soil:water ratio, and subsequently analyzed using SevenExcellence pH/mV and Conductivity Meters. After removing organic material and carbonates using hydrogen peroxide and hydrochloric acid, respectively, the soil texture—including sand (50–2000 µm), silt (2–50 µm), and clay (<2 µm)—was analyzed using a laser particle characterization analyzer (Beckman Coulter LS-230, Brea, CA, USA) [19]. The cation-exchange capacity (CEC) was determined via the ammonium acetate method [25]. The content of total K, P, Ca, Mg, iron (Fe), and aluminum (Al) in the soil were analyzed using total X-ray fluorescence (XRF) spectroscopy. Available P and available K were determined via the NaHCO3 extraction method [26] and the NH4OAc extraction method [27], respectively. After the extraction with the citrate bicarbonate dithionite system, free Fe and Al oxides were measured using ICP-AES [28].
2.4. Determination of Archaeal amoA and Bacterial amoA Abundances
A FastDNA® Spin Kit for Soil (MP Biomedicals, Cleveland, OH, USA) was used to extract the soil DNA, which was subsequently quantified and purified using a NanoDrop ND-2000 Spectrophotometer (NanoDrop Technologies, Wilmington, DE, USA). The quantitative PCR method was carried out to determine the genes of archaeal amoA (AOA) and bacterial amoA (AOB) using a CFX96 Optical Real-Time Detection System (Bio-Rad, Laboratories Inc., Hercules, CA, USA). The Arch-amoAF/Arch-amoAR [29] and amoA1F/amoA2R [30] were the primers of the AOA and AOB genes, respectively. The detailed information of DNA extraction, along with the amplification and quantitation of PCR, was obtained from the works of Garousi et al. (2021) [19] and Wang et al. (2015) [31].
2.5. Statistical Data Analyses
According to the changes in content and atom% of NH4+ in the 15NH4NO3 treatment or NO3− in the NH415NO3 treatment between times t1 (0.5 h) and t2 (24 h), the mineralization and nitrification rates [32], respectively, were calculated as follows:
where MNorg is the mineralization rate (mg N kg−1 d−1), while [NH4+]t1, [NH4+]t2, APEt1, and APEt2 are the content (mg N kg−1) and 15N atom% excess of NH4+ at time 0.5 h and 24 h, respectively.
where ONH4 is the nitrification rate (mg N kg−1 d−1), while [NO3−]t1, [NO3−]t2, APEt1, and APEt2 are the content (mg N kg−1) and 15N atom% excess of NO3− at time 0.5 h and 24 h, respectively.
The mean residence times (d) of NH4+ (MRT NH4+) and NO3− (MRT NO3−) were calculated using the equation developed by Corre et al. [33], which can indicate the turnover of inorganic N.
where c(NH4+) and c(NO3−) are the initial content of NH4+ and NO3− (mg N kg−1) in the soil, respectively.
Differences in soil properties, AOA and AOB abundances, and N-cycling rates between pitaya plantations and woodlands were estimated by analysis of variance (ANOVA) at the p < 0.05 significance level. Redundancy analysis (RDA) was used to determine the relationships between soil properties, AOA and AOB abundances, and N-cycling rates, using Canoco 5.
3. Results
3.1. Soil Properties and AOA/AOB Abundances
The soil properties were different between the woodlands and pitaya plantations (Table 1). Compared to woodlands (72.5 mg C kg−1 and 6.83 mg N kg−1), pitaya cultivation significantly decreased the content of SOC and TN to 12.8–22.3 mg C kg−1 and 1.47–2.87 mg N kg−1, respectively (p < 0.05). The soil C/N ratio, WHC, and CEC also decreased significantly after the conversion of woodlands to pitaya plantations (p < 0.05). Compared to woodlands (6.52), the soil pH did not significantly change in short-term pitaya plantations (3 years) (p > 0.05), but significantly decreased to 5.57 in long-term pitaya plantations (15 years) (p < 0.05). The content of Ca and Mg in soils under woodlands was about 6.05–17.7 and 4.83–6.48 times higher than in soils under pitaya cultivation, respectively. Conversely, pitaya cultivation greatly increased the available K, available P, total K, P, Fe, Al, free Fe, and Al oxides content as well as the silt composition, with significant differences between woodlands and the 15-year pitaya plantation (p < 0.05). No significant differences in clay and/or clay compositions were found in any of the studied soils (p > 0.05). In comparison to woodlands (8.42 and 16.0 mg N kg−1, respectively), pitaya cultivation increased NH4+ and NO3− content to 12.2–22.5 mg N kg−1 and 21.4–27.2 mg N kg−1, respectively, but the difference was not significant (p > 0.05). The content of both Ca and Mg was significantly and positively correlated with SOC and TN content, WHC, and CEC, suggesting the importance of Ca and Mg in maintaining soil conditions.

Table 1.
Soil properties under woodland and three pitaya plantations (n = 12).
In all of the studied soils, AOA abundance was significantly higher than AOB abundance (p < 0.05, Table 1). Compared to woodland (6.70 × 107 and 2.54 × 106 copies g−1, respectively), the abundance of AOA and AOB in soil under 3-year pitaya cultivation significantly increased to 4.19 × 108 and 1.36 × 107 copies g−1, respectively. However, the abundance of AOA and AOB significantly decreased with prolonged pitaya cultivation, with the lowest values found in soil under the 15-year pitaya cultivation.
3.2. N-Cycling Rates
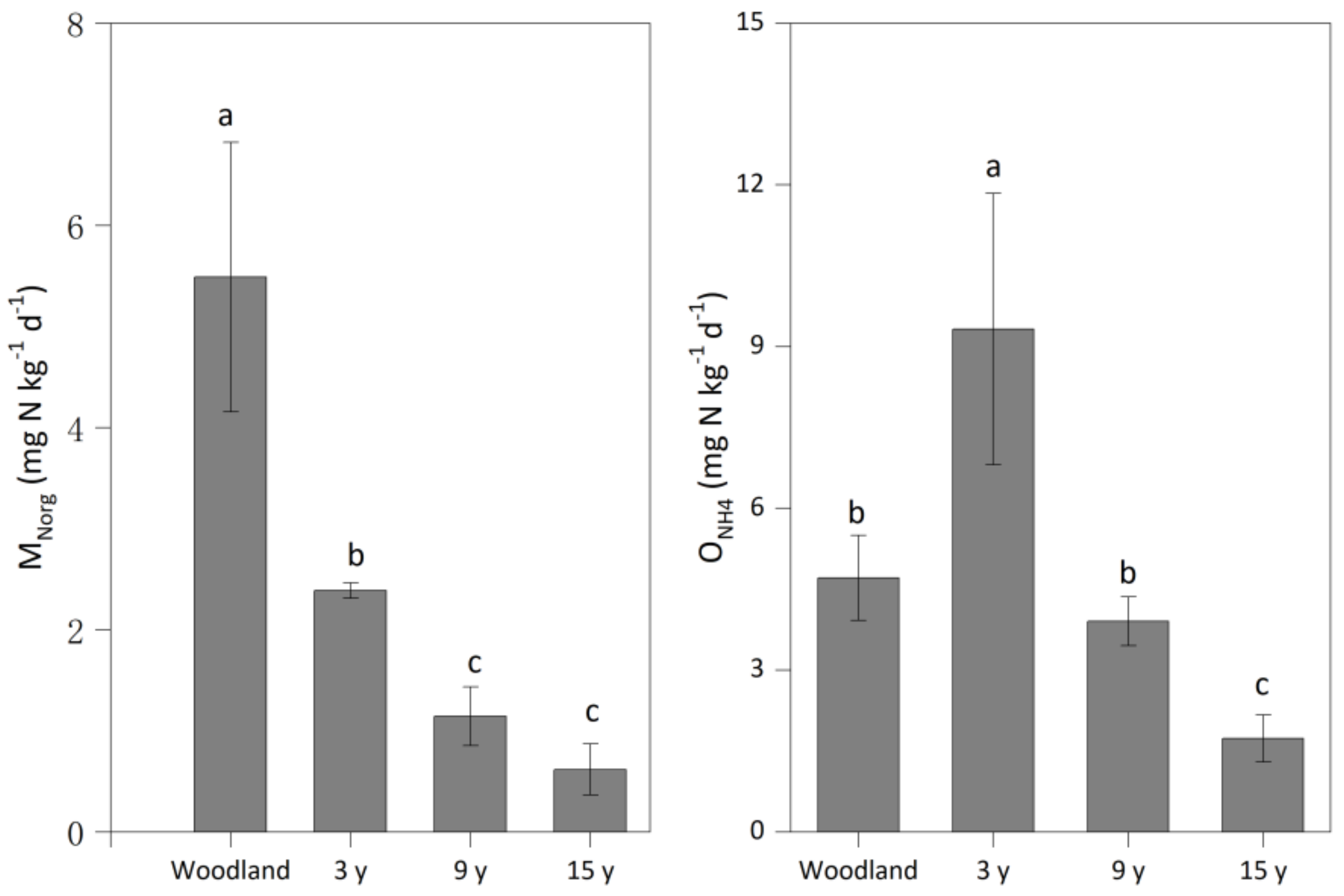
Compared to woodland (5.49 mg N kg−1 d−1), pitaya cultivation significantly decreased the MNorg rate to 2.38–0.62 mg N kg−1 d−1—a change that was more obvious with the increase in the duration of pitaya cultivation. A higher ONH4 rate (9.32 mg N kg−1 d−1) was observed in soil under the 3-year pitaya cultivation than in woodland (4.71 mg N kg−1 d−1), while this rate significantly decreased to 3.91 mg N kg−1 d−1 in soil under the 9-year pitaya cultivation and 1.74 mg N kg−1 d−1 in soil under the 15-year pitaya cultivation (Figure 2).
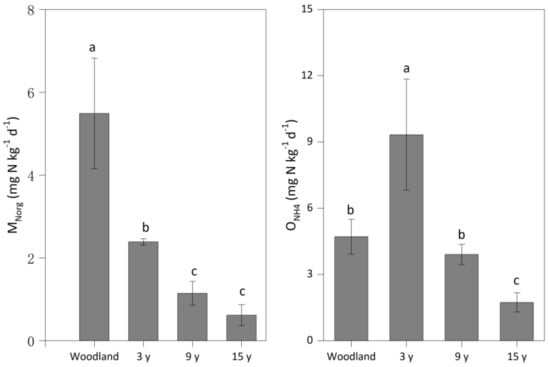
Figure 2.
The mineralization (MNorg) and nitrification (ONH4) rates in soils under woodland and three pitaya plantations. Error bars depict standard deviations. Identical letters for MNorg or ONH4 indicate no significant differences between woodland and pitaya plantations at p > 0.05.
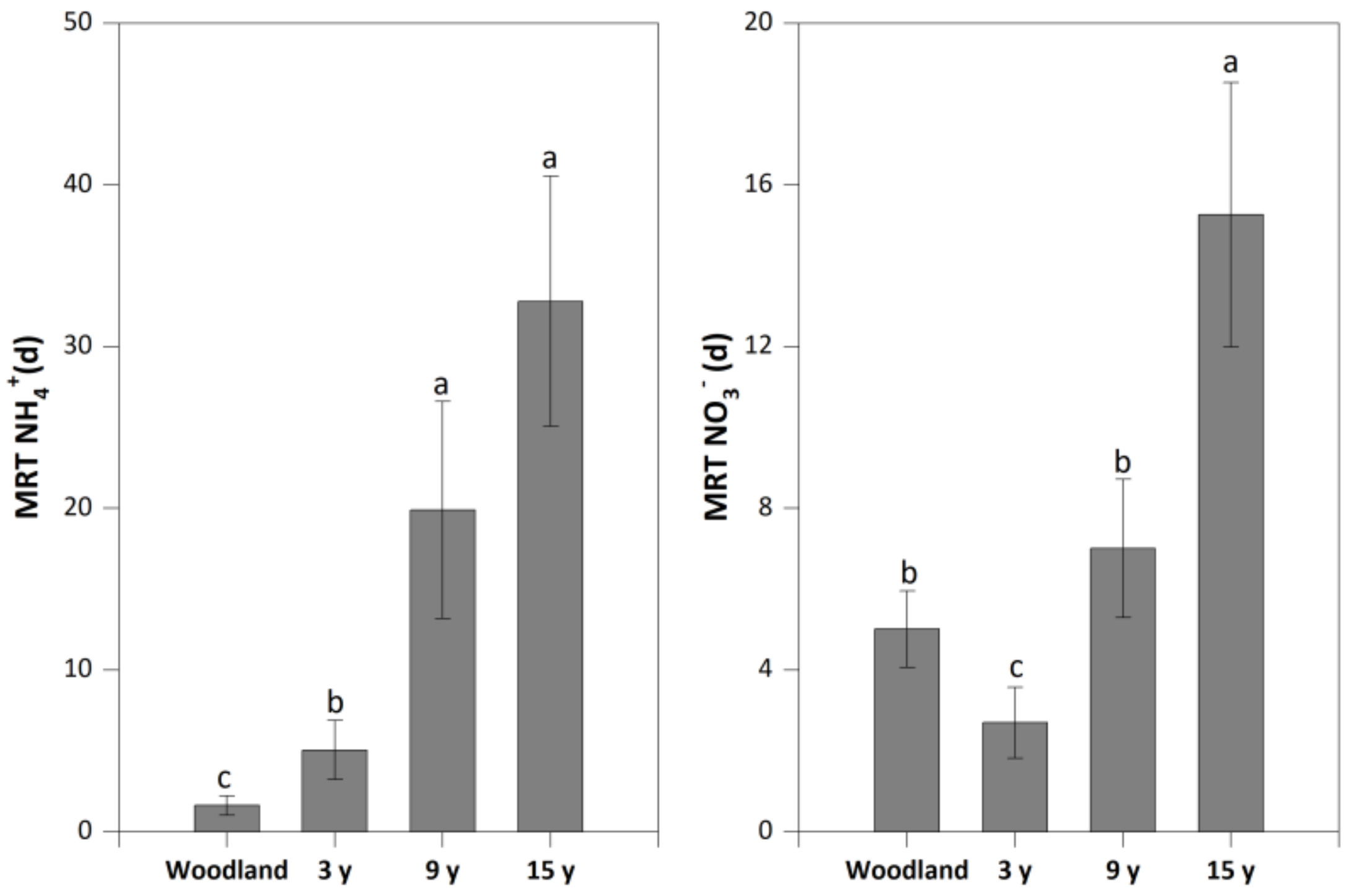
Compared to woodland (1.63 d), pitaya cultivation significantly increased the mean residence time of NH4+ to 5.06–32.8 d, with the highest value in soil under the 15-year pitaya cultivation. Conversely, a lower mean residence time of NO3− in soil under the 3-year pitaya cultivation (2.70 d) was found than that in woodland (5.01 d), while this rate significantly increased to 15.3 d in soil under the 15-year pitaya cultivation (Figure 3).
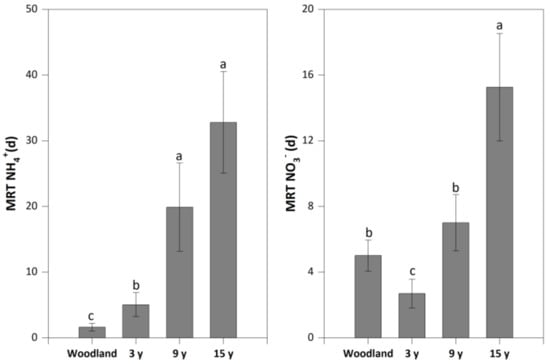
Figure 3.
The mean residence time of NH4+ (MRT NH4+) and NO3− (MRT NO3−) in soils under woodland and three pitaya plantations. Error bars depict standard deviations. Identical letters for MRT NH4+ and MRT NO3− indicate no significant differences between woodland and the three pitaya plantations at p > 0.05.
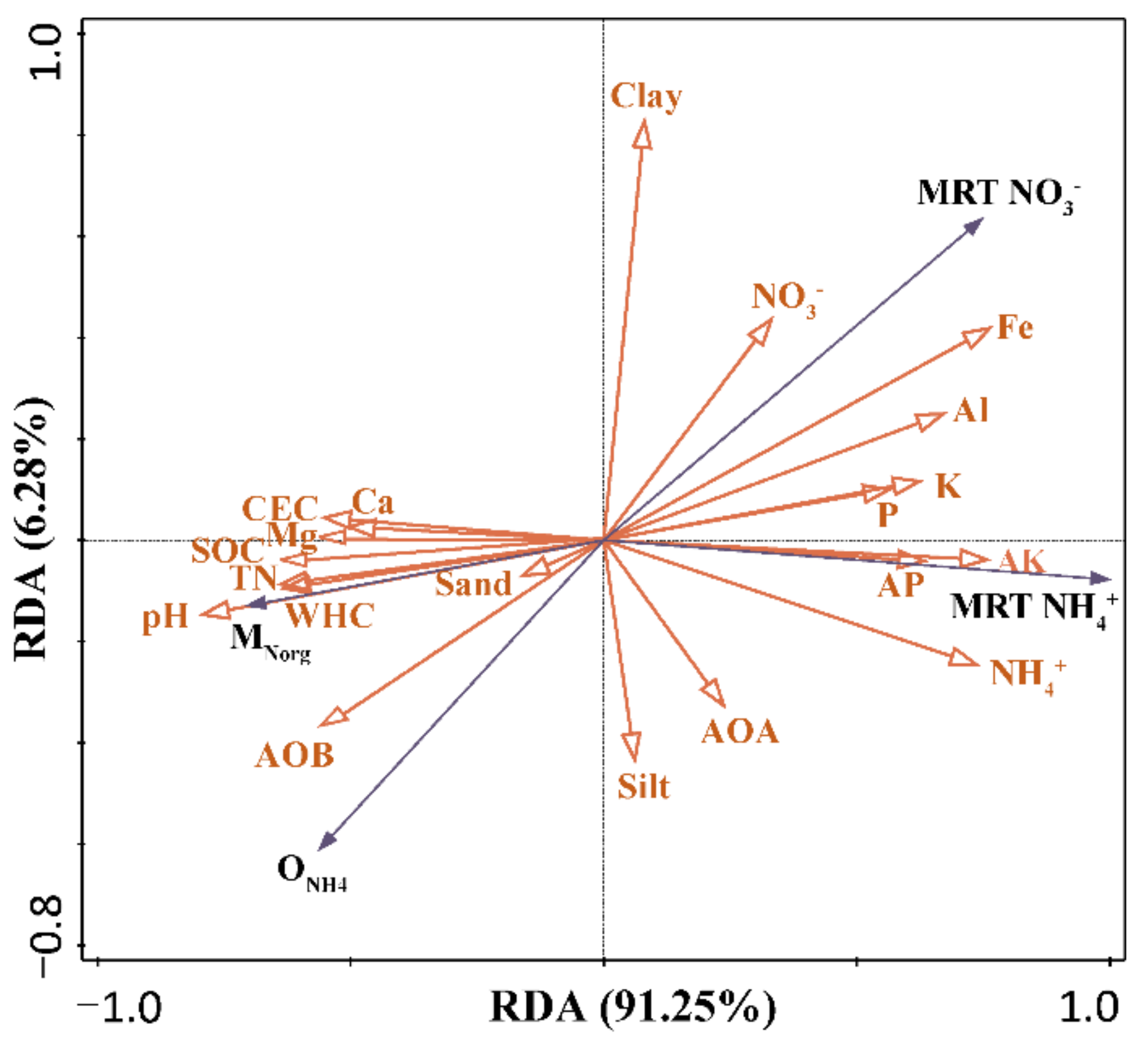
The correlation analyses showed that SOC, total N, Ca, and Mg content, along with CEC, were significantly and positively correlated with the mineralization rate, but negatively correlated with the mean residence time of NH4+ and NO3− (Figure 4), indicating that the decline in soil quality negatively affected the turnover of inorganic N.
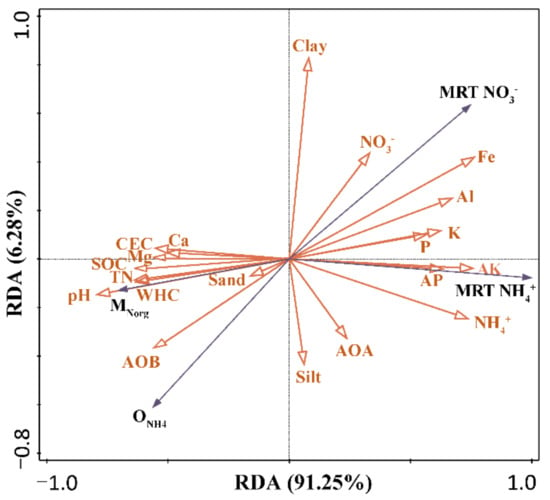
Figure 4.
Redundancy analysis (RDA) of soil N-cycling rates, AOA and AOB abundance, and soil properties under woodland and pitaya plantations.
4. Discussion
In this study, pitaya cultivation greatly increased the content of inorganic N, total P, total K, available P, and available K. According to the classification standard of soil fertility from the Soil Survey Office of China (Table S1), both P and K content in soils under pitaya cultivation were at high levels, which could be attributed to the application of large amounts of mineral fertilizer. However, pitaya cultivation significantly decreased the SOC and TN content compared to woodland. The present study showed that the SOC and TN content decreased approximately five-fold in the 15-year pitaya plantation compared to woodland, which is consistent with the findings of previous studies conducted in other long-term crop plantations (e.g., bananas, citrus, rubber) in subtropical karst regions [21,34]. Due to the reduction in soil organic matter, soil may become heavily clayed in long-term pitaya plantations with high clay composition (>50%). These results suggest the possible decline in soil quality under long-term crop cultivation in karst regions, which may adversely affect soil biotic factors (e.g., microbial abundance and activity), thereby altering soil N cycling.
In this study, the mineralization rate in soils under woodland in the subtropical karst region reached 5.49 mg N kg−1 d−1, which was significantly higher than those in highly weathered red soils in non-karst regions of the same latitude (1.01–4.63; average 2.83 mg N kg−1 d−1) [18,35,36]. Furthermore, a higher nitrification rate was also found in our studied sites than in the red soil of woodlands in subtropical non-karst regions (0.06–1.45; average 0.76 mg N kg−1 d−1) [18,35,36]. This difference could be attributed to the characteristics of calcareous soils in karst regions. Calcareous soil is characterized by Ca enrichment, which is the important factor driving the changes in pH, SOC, and TN content, along with other properties (soil moisture, CEC, etc.). In undisturbed ecosystems in karst regions, high Ca levels can promote the accumulation of soil organic matter and the formation of macroaggregates [37,38,39], all of which can improve the soil quality and soil structure. In addition, soil pH is also positively correlated with Ca content. Consequently, the increase in the substrate (i.e., organic N) facilitates the occurrence of mineralization to produce more NH4+. Previous studies have found a significant and positive correlation between pH and nitrification [22,35]. Under high pH conditions, NH4+ is easily oxidized to NO3− by the nitrifying microorganisms [35,40]. Due to high mineralization and nitrification rates, inorganic N content was relatively high, and was mainly dominated by NO3− in the studied soils under woodlands in the karst region (Table 1).
Compared to woodland, pitaya cultivation significantly reduced the soil mineralization rate, and this effect was more obvious with the increase in the duration of pitaya cultivation. When pitaya had been planted for more than 9 years, the mineralization rate was lower than 1 mg N kg−1 d−1. Similar results have also been found for other crops (e.g., citrus, rubber) cultivated for more than 7 years in karst regions, with mineralization rates of 0.61–1.34 mg N kg−1 d−1 [19,21]. These results suggest that long-term crop cultivation can greatly decrease the mineralization rate in soil in karst regions, indicating a decline in soil inorganic N supply, which supports our hypothesis. The decreased organic matter with the increasing duration of pitaya cultivation may explain the low mineralization rates in karst regions, because significant and positive relationships between mineralization rate, SOC, and total N content were found in this study (Figure 4). When woodland is converted to pitaya plantations, the changes in management practices—including tillage, fertilization, and the reduction in coverage due to the removal of understory vegetation—can lead to the rapid loss of organic matter, thereby reducing the substrate for mineralization [24,41]. In addition, the soil is heavily clayed after long-term crop cultivation in karst regions, which may decrease the microbial abundance and activity involved in the mineralization of organic N [19,21].
Generally, the application of N fertilizers can increase nitrification rates by stimulating the abundance and/or activity of soil-nitrifying microorganisms, especially under high pH conditions [22,42]. Thus, an increased nitrification rate can be expected in soils amended with mineral N fertilizer. Indeed, a high nitrification rate (9.32 mg N kg−1 d−1) was observed in the soil under 3-year pitaya cultivation. However, this was not the case for the 15-year pitaya plantation, which had an extremely low nitrification rate (only 1.74 mg N kg−1 d−1), even though a large amount of N fertilizer was applied (Table 1). Similar results were also found in other crop plantations (e.g., rubber, citrus) with long-term cultivation [19,21]. An important mechanism underlying the reduced nitrification rate may be that low soil organic matter content in the long-term pitaya plantations can destroy the soil structure, causing the soil to become clayey [19,21]. This could restrict the diffusion of oxygen and, in turn, inhibit the stimulatory effect of N fertilizer on nitrification in soil [43]. In addition, with the increasing duration of pitaya cultivation, the increase in soil iron and aluminum oxide content could also have an adverse impact on nitrifying microorganisms, resulting in a decline in the nitrification rate [14,44]. In this study, long-term pitaya cultivation significantly decreased the abundances of AOA and AOB compared to short-term pitaya cultivation, and the abundance of AOB was greatly positively correlated with the nitrification rate (Figure 4), further supporting the above speculation. All of these results suggest that long-term pitaya cultivation can significantly deteriorate soil quality (e.g., the decline in SOC and TN content, clayed texture, and the increase in Fe and Al oxides) and, subsequently, decrease the supply capacity of inorganic N by decreasing mineralization and nitrification rates. Noticeably, 15-year pitaya cultivation decreased soil pH by one unit compared to woodlands. Considering that nitrification is an important process in the release of hydrogen (H+) [45], the continuous occurrence of nitrification caused by N fertilizers, combined with the direct loss of Ca, could be responsible for the significant decline in pH during pitaya cultivation.
Noticeably, long-term pitaya cultivation significantly increased the mean residence time of inorganic N in soil, indicating a decline in inorganic N turnover. In this case, large amounts of N fertilizer in the form of NH4+ or NO3− may not be effectively recycled, and may subsequently be lost in long-term pitaya plantations. Therefore, some appropriate management practices are especially imperative to regulate soil mineralization and nitrification processes, and to stimulate the turnover of inorganic N. Based on the positive relationships between mineralization rate, the mean residence time of inorganic N, and the content of organic C and total N, the application of organic fertilizers may accelerate inorganic N turnover in soil. Further work is needed to study whether organic matter can improve the deteriorated soil environment to stimulate N cycling.
5. Conclusions
The conversion of woodland to pitaya cultivation significantly changed the basic soil properties, causing declines in the SOC, TN, Ca, and Mg content as well as in the pH levels in the pitaya plantations. In addition, pitaya cultivation greatly decreased the mineralization and NH4+ turnover rates, decreasing the inorganic N supply capacity, and this effect was more pronounced with the increasing duration of pitaya cultivation. Conversely, short-term pitaya cultivation significantly stimulated nitrification and NO3− turnover rates, while long-term pitaya cultivation inhibited both rates. Overall, long-term pitaya cultivation significantly reduced the supply capacity and turnover of soil inorganic N. Thus, appropriate management practices must be developed in the future in order to increase soil inorganic N turnover in long-term pitaya plantations.
Supplementary Materials
The following supporting information can be downloaded at: https://www.mdpi.com/article/10.3390/land11060781/s1, Table S1: Classification of soil fertility from Soil Survey Office of China (1992).
Author Contributions
Conceptualization, L.Y. and X.Z.; investigation, L.Y., X.Z., J.L. and D.W.; formal analysis, L.Y., J.L. and D.W.; writing—review and editing, L.Y., L.M. and T.Z. All authors have read and agreed to the published version of the manuscript.
Funding
This work was supported by grants from the National Natural Science Foundation of China (41877348) and the Guangxi Science and Technology Program (GUIKE AD21196001).
Institutional Review Board Statement
Not applicable.
Informed Consent Statement
Not applicable.
Data Availability Statement
Not applicable.
Conflicts of Interest
The authors declare no conflict of interest.
References
- Jiang, Z.C.; Lian, Y.Q.; Qin, X.Q. Rocky desertification in Southwest China: Impacts, causes, and restoration. Earth Sci. Rev. 2014, 132, 1–12. [Google Scholar] [CrossRef]
- Wang, S.J.; Liu, Q.M.; Zhang, D.F. Karst rocky desertification in Southwestern China: Geomorphology, landuse, impact and rehabilitation. Land Degrad. Dev. 2004, 15, 115–121. [Google Scholar] [CrossRef]
- Cheng, F.; Lu, H.F.; Ren, H.; Zhou, L.; Zhang, L.H.; Li, J.; Lu, X.J.; Huang, D.W.; Zhao, D. Integrated emergy and economic evaluation of three typical rocky desertification control modes in karst areas of Guizhou Province, China. J. Clean Prod. 2017, 161, 1104–1128. [Google Scholar] [CrossRef]
- Fresh Plaza. 2018. Available online: https://www.freshplaza.cn/article/8812800/ (accessed on 1 February 2022).
- Luo, J.; Xu, M.; Qi, Z.; Xiong, R.; Cheng, Y.; Liu, C.L.; Wei, S.S.; Tang, H. Differential responses of the soil microbial community in two pitaya orchards with different mulch types. Sci Rep. 2019, 9, 10413. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Luu, T.T.H.; Le, T.L.; Huynh, N.; Quintela-Alonso, P. Dragon fruit: A review of health benefits and nutrients and its sustainable development under climate changes in Vietnam. Czech J. Food Sci. 2021, 39, 71–94. [Google Scholar] [CrossRef]
- Norouzi, M.; Ayoubi, S.; Jalalian, A.; Khademi, H.; Dehghani, A.A. Predicting rainfed wheat quality and quantity by artificial neural network using terrain and soil characteristics. Acta Agr. Scand B 2010, 60, 341–352. [Google Scholar] [CrossRef]
- Schiattone, M.I.; Viggiani, R.; Venere, D.D.; Sergio, L.; Cantore, V.; Todorovic, M.; Perniola, M.; Candido, V. Impact of irrigation regime and nitrogen rate on yield, quality and water use efficiency of wild rocket under greenhouse conditions. Sci. Hortic-Amst. 2018, 229, 182–192. [Google Scholar] [CrossRef]
- Schimel, J.P.; Bennett, J. Nitrogen mineralization: Challenges of a changing paradigm. Ecology 2004, 85, 591–602. [Google Scholar] [CrossRef]
- Hobbie, E.A.; Högberg, P. Nitrogen isotopes link mycorrhizal fungi and plants to nitrogen dynamics. New Phytol. 2012, 196, 367–382. [Google Scholar] [CrossRef]
- Zhang, W.; Zhao, J.; Pan, F.J.; Li, D.J.; Chen, H.S.; Wang, K.L. Changes in nitrogen and phosphorus limitation during secondary succession in a karst region in Southwest China. Plant Soil. 2015, 391, 77–91. [Google Scholar] [CrossRef]
- Liu, X.; Zhang, W.; Wu, M.; Ye, Y.Y.; Wang, K.L.; Li, D.J. Changes in soil nitrogen stocks following vegetation restoration in a typical karst catchment. Land Degrad. Dev. 2019, 30, 60–72. [Google Scholar] [CrossRef] [Green Version]
- Müller, C.; Rütting, T.; Kattge, J.; Laughlin, R.J.; Stevens, R.J. Estimation of parameters in complex 15N tracing models by Monte Carlo sampling. Soil Biol. Biochem. 2007, 39, 715–726. [Google Scholar] [CrossRef]
- Xie, Y.; Yang, L.; Zhu, T.B.; Yang, H.; Zhang, J.B.; Yang, J.L.; Cao, J.H.; Bai, B.; Jiang, Z.C.; Liang, Y.M.; et al. Rapid recovery of nitrogen retention capacity in a subtropical acidic soil following afforestation. Soil Biol. Biochem. 2018, 120, 171–180. [Google Scholar] [CrossRef]
- Rütting, T.; Clough, T.J.; Müller, C.; Lieffering, M.; Newton, P.C.D. Ten years of elevated atmospheric carbon dioxide alters soil nitrogen transformations in a sheep–grazed pasture. Glob. Chang. Biol. 2010, 16, 2530–2542. [Google Scholar] [CrossRef]
- Bárcenas-Moreno, G.; Rousk, J.; Bååth, E. Fungal and bacterial recolonisation of acid and alkaline forest soils following artificial heat treatments. Soil Biol. Biochem. 2011, 43, 1023–1033. [Google Scholar] [CrossRef]
- Chen, H.; Li, D.J.; Xiao, K.C.; Wang, K.L. Soil microbial processes and resource limitation in karst and non-karst forests. Funct. Ecol. 2018, 32, 1400–1409. [Google Scholar] [CrossRef]
- Li, D.J.; Yang, Y.; Chen, H.; Xiao, K.C.; Song, T.Q.; Wang, K.L. Soil gross nitrogen transformations in typical Karst and nonkarst forests, Southwest China. J. Geophys. Res Biogeo. 2017, 122, 2831–2840. [Google Scholar] [CrossRef]
- Garousi, F.; Shan, Z.J.; Ni, K.; Yang, H.; Shan, J.; Cao, J.H.; Jiang, Z.C.; Yang, J.L.; Zhu, T.B.; Müller, C. Decreased inorganic N supply capacity and turnover in calcareous soil under degraded rubber plantation in the tropical karst region. Geoderma 2021, 381, 114754. [Google Scholar] [CrossRef]
- Zhang, J.B.; Zhu, T.B.; Meng, T.Z.; Zhang, Y.C.; Yang, J.J.; Yang, W.Y.; Müller, C.; Cai, Z.C. Agricultural land use affects nitrate production and conservation in humid subtropical soils in China. Soil Biol. Biochem. 2013, 62, 107–114. [Google Scholar] [CrossRef]
- Shan, Z.J.; Yin, Z.; Yang, H.; Zuo, C.Q.; Zhu, T.B. Long-term cultivation of fruit plantations decreases mineralization and nitrification rates in calcareous soil in the karst region in Southwestern China. Forests 2020, 11, 1282. [Google Scholar] [CrossRef]
- Chu, H.Y.; Fujii, T.; Morimoto, S.; Lin, X.G.; Yagi, K. Population size and specific nitrification potential of soil ammonia–oxidizing bacteria under long–term fertilizer management. Soil Biol. Biochem. 2008, 40, 1960–1963. [Google Scholar] [CrossRef]
- Zhang, J.B.; Cai, Z.C.; Yang, W.Y.; Zhu, T.B.; Yu, Y.J.; Yan, X.Y.; Jia, Z.J. Long-term field fertilization affects soil nitrogen transformations in a rice-wheat-rotation cropping system. Plant Nutr. Soil Sci. 2012, 175, 939–946. [Google Scholar] [CrossRef]
- Yang, H.; Garousi, F.; Wang, J.; Cao, J.H.; Xu, X.L.; Zhu, T.B.; Müller, C. Land use effects on gross soil nitrogen transformations in karst desertification area. Plant Soil. 2021, 17. [Google Scholar] [CrossRef]
- Ross, D.S.; Ketterings, Q. Recommended methods for determining soil cation exchange capacity. Recomm. Soil Test. Proced. Northeast. USA 1995, 2, 62–70. [Google Scholar]
- Olsen, S.R. Estimation of Available Phosphorus in Soils by Extraction with Sodium Bicarbonate; US Department of Agriculture: Washington, DC, USA, 1954.
- Sparks, D.L. Methods of Soil Analysis, Part 3. Chemical Methods. Madison, W.I., Ed.; Soil Science Society of America and American Society of Agronomy: Madison, WI, USA, 1996; pp. 555–574. [Google Scholar]
- Aguilera, N.H.; Jackson, M.L. Iron oxide removal from soils and clays. Soil Sci. Soc. Am. J. 1953, 17, 359–364. [Google Scholar] [CrossRef]
- Francis, C.A.; Roberts, K.J.; Beman, J.M.; Santoro, A.E.; Oakley, B.B. Ubiquity and diversity of ammonia-oxidizing archaea in water columns and sediments of the ocean. Proc. Natl. Acad. Sci. USA 2005, 102, 14683–14688. [Google Scholar] [CrossRef] [Green Version]
- Rotthauwe, J.H.; Witzel, K.P.; Liesack, W. The ammonia monooxygenase structural gene amoA as a functional marker: Molecular fine-scale analysis of natural ammonia-oxidizing populations. Appl. Environ. Microb. 1997, 63, 4704–4712. [Google Scholar] [CrossRef] [Green Version]
- Wang, X.D.; Le, T.B.K.; Lajoie, B.R.; Dekker, J.; Laub, M.T.; Rudner, D.Z. Condensin promotes the juxtaposition of DNA flanking its loading site in Bacillus subtilis. Genes. Dev. 2015, 29, 1661–1675. [Google Scholar] [CrossRef] [Green Version]
- Kirkham, D.; Bartholomew, W.V. Equations for following nutrient transformations in soil utilizing tracer data. Soil Sci. Soc. Am. Proc. 1954, 18, 33–34. [Google Scholar] [CrossRef]
- Corre, M.D.; Brumme, R.; Veldkamp, E.; Beese, F.O. Changes in nitrogen cycling and retention processes in soils under spruce forests along a nitrogen enrichment gradient in Germany. Global Change Biol. 2007, 13, 1509–1527. [Google Scholar] [CrossRef]
- Qin, X.H.; Yang, C.; Yang, L.; Ma, E.; Meng, L.; Zhu, T.B. Response of Gross Mineralization and Nitrification Rates to Banana Cultivation Sites Converted from Natural Forest in Subtropical China. Land 2021, 10, 376. [Google Scholar] [CrossRef]
- Zhang, J.B.; Zhu, T.B.; Cai, Z.C.; Müller, C. Nitrogen cycling in forest soils across climate gradients in Eastern China. Plant Soil. 2011, 342, 419–432. [Google Scholar] [CrossRef]
- Zhang, J.B.; Cai, Z.C.; Zhu, T.B.; Yang, W.Y.; Müller, C. Mechanisms for the retention of inorganic N in acidic forest soils of southern China. Sci. Rep. 2013, 3, 2342. [Google Scholar] [CrossRef] [PubMed]
- Lützow, M.V.; Kögel-Knabner, I.; Ekschmitt, K.; Matzner, E.; Guggenberger, G.; Marschner, B.; Flessa, H. Stabilization of organic matter in temperate soils: Mechanisms and their relevance under different soil conditions-a review. Eur. J. Soil Sci. 2006, 57, 426–445. [Google Scholar] [CrossRef]
- Kaiser, M.; Walter, K.; Ellerbrock, R.H.; Sommer, M. Effects of land use and mineral characteristics on the organic carbon content, and the amount and composition of Na-pyrophosphate-soluble organic matter, in subsurface soils. Eur. J. Soil Sci. 2011, 62, 226–236. [Google Scholar] [CrossRef]
- Xiao, K.C.; He, T.G.; Chen, H.; Peng, W.X.; Song, T.Q.; Wang, K.L.; Li, D.J. Impacts of vegetation restoration strategies on soil organic carbon and nitrogen dynamics in a karst area, Southwest China. Ecol. Eng. 2017, 101, 247–254. [Google Scholar] [CrossRef]
- Wan, Y.J.; Ju, X.T.; Ingwersen, J.; Schwarz, U.; Stange, C.F.; Zhang, F.S.; Streck, T. Gross nitrogen transformations and related nitrous oxide emissions in an intensively used calcareous soil. Soil Sci. Soc. Am. J. 2009, 73, 102–112. [Google Scholar] [CrossRef]
- Xiao, S.S.; Ye, Y.Y.; Xiao, D.; Chen, W.R.; Wang, K.L. Effects of tillage on soil N availability, aggregate size, and microbial biomass in a subtropical karst region. Soil Till. Res. 2019, 192, 187–195. [Google Scholar] [CrossRef]
- Norton, J.; Ouyang, Y. Controls and adaptive management of nitrification in agricultural soils. Front. Microbiol. 2019, 10, 1931. [Google Scholar] [CrossRef] [Green Version]
- Nawaz, M.F.; Bourrié, G.; Trolard, F. Soil compaction impact and modelling. A review. Agron. Sustain. Dev. 2013, 33, 291–309. [Google Scholar] [CrossRef] [Green Version]
- Jiang, X.J.; Xin, X.P.; Li, S.W.; Zhou, J.C.; Zhu, T.B.; Müller, C.; Cai, Z.C.; Wright, A.L. Effects of Fe oxide on N transformations in subtropical acid soils. Sci. Rep. 2015, 5, 8615. [Google Scholar] [CrossRef]
- Zhao, W.; Cai, Z.C.; Xu, Z.H. Does ammonium-based N addition influence nitrification and acidification in humid subtropical soils of China? Plant Soil 2007, 297, 213–221. [Google Scholar] [CrossRef]
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).