
Which Traits Influence Bird Survival in the City? A Review


Abstract
1. Introduction
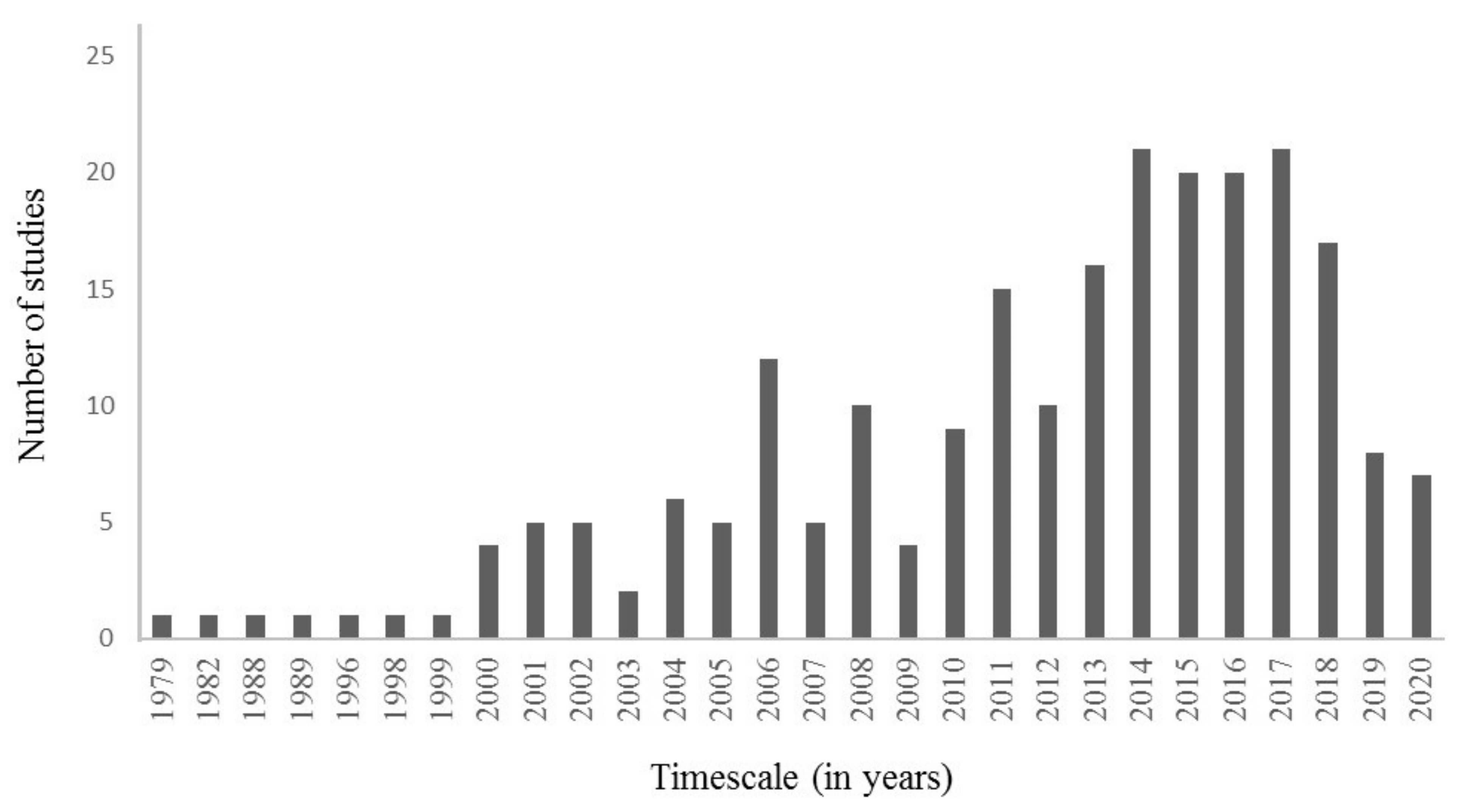
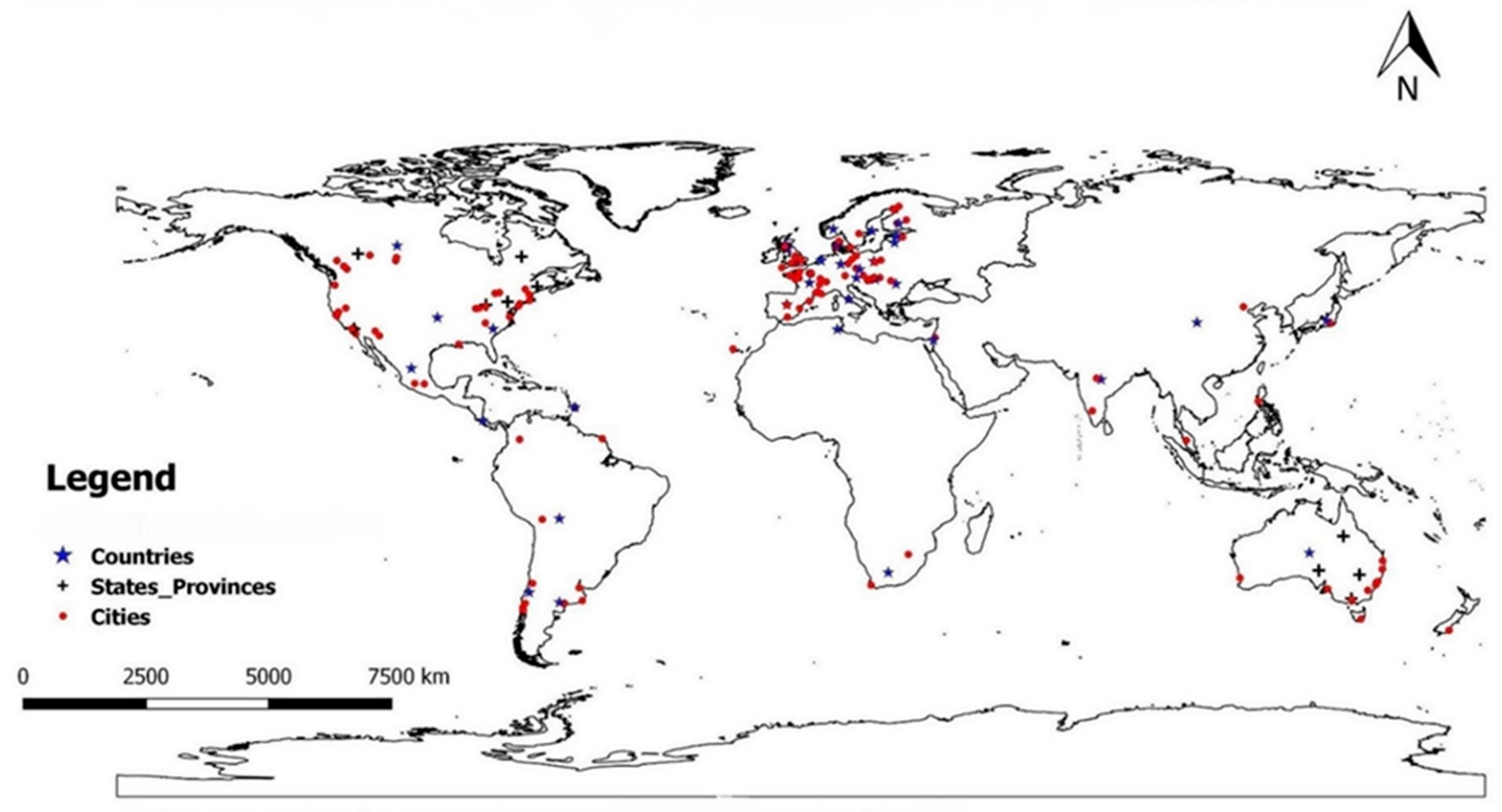
2. Materials and Methods
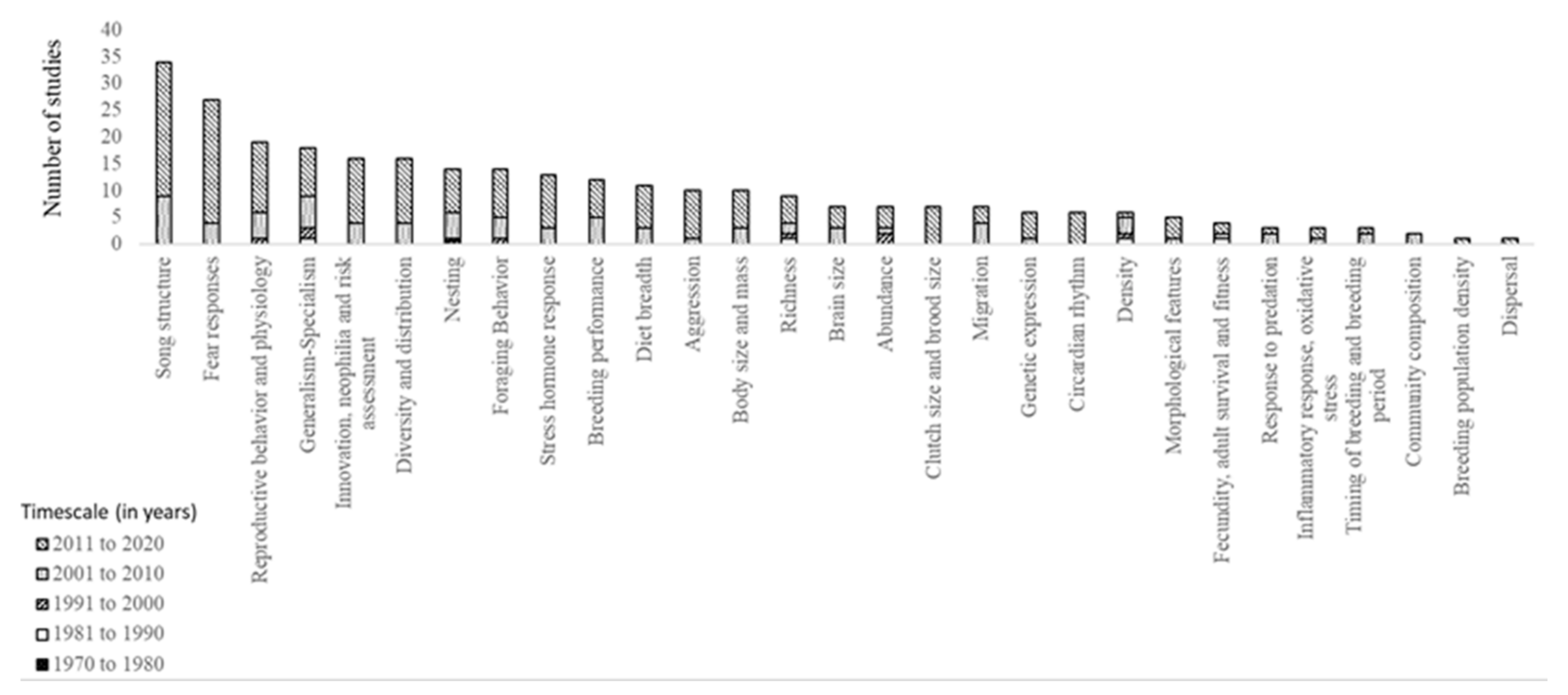
3. Results
3.1. Ecological Traits
3.1.1. Bird Diversity and Abundance
3.1.2. Generalism—Specialism
3.1.3. Diet Breadth
3.1.4. Nesting Sites
3.2. Life History Traits
3.3. Physiological Traits
3.3.1. Body Mass, Size, and Plumage Coloration
3.3.2. Brain Size
3.3.3. Stress Response Physiology
3.3.4. Reproductive Physiology
3.3.5. Immune and Inflammatory Responses, Oxidative Stress Response
3.4. Behavioral Traits
3.4.1. Song Structure
3.4.2. Boldness and Tolerance to Human Presence
3.4.3. Neophobic and Innovative Behavior
3.4.4. Aggression
3.5. Genetic Traits
4. Discussion
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
References
- McKinney, M.L. Urbanization, biodiversity and conservation. BioScience 2002, 52, 883–890. [Google Scholar] [CrossRef]
- Kowarik, I. Novel urban ecosystems, biodiversity, and conservation. Environ. Pollut 2011, 159, 1974–1983. [Google Scholar] [CrossRef] [PubMed]
- UNDESA. 2018 The Sustainable Development Goals Report; United Nations: New York, NY, USA, 2018; Available online: https://www.un.org/development/desa/publications/the-sustainable-development-goals-report-2018.html (accessed on 20 June 2020).
- Lepczyk, C.A.; Aronson, M.F.; Evans, K.L.; Goddard, M.A.; Lerman, S.B.; MacIvor, J.S. Biodiversity in the city: Fundamental questions for understanding the ecology of urban green spaces for biodiversity conservation. BioScience 2017, 67, 799–807. [Google Scholar] [CrossRef]
- Sol, D.; Duncan, R.P.; Blackburn, T.M.; Cassey, P.; Lefebvre, L. Big brains, enhanced cognition, and response of birds to novel environments. Proc. Natl. Acad. Sci. USA 2005, 102, 5460–5465. [Google Scholar] [CrossRef] [PubMed]
- Liu, Z.; He, C.; Wu, J. The relationship between habitat loss and fragmentation during urbanization: An empirical evaluation from 16 world cities. PLoS ONE 2016, 11, e0154613. [Google Scholar] [CrossRef] [PubMed]
- Belaire, J.A.; Westphal, L.M.; Whelan, C.J.; Minor, E.S. Urban residents’ perceptions of birds in the neighborhood: Biodiversity, cultural ecosystem services, and disservices. Condor 2015, 117, 192–202. [Google Scholar] [CrossRef]
- Aronson, M.F.; la Sorte, F.A.; Nilon, C.H.; Katti, M.; Goddard, M.A.; Lepczyk, C.A.; Warren, P.S.; Williams, N.S.; Cilliers, S.; Clarkson, B.; et al. A global analysis of the impacts of urbanization on bird and plant diversity reveals key anthropogenic drivers. Proc. R. Soc. B Biol. Sci. 2014, 281, 20133330. [Google Scholar] [CrossRef]
- Hensley, C.B.; Trisos, C.H.; Warren, P.S.; MacFarland, J.; Blumenshine, S.; Reece, J.; Katti, M. Effects of urbanization on native bird species in three southwestern US Cities. Front. Ecol. Evol. 2019, 7, 71. [Google Scholar] [CrossRef]
- Whelan, C.J.; Wenny, D.G.; Marquis, R.J. Ecosystem services provided by birds. Ann. N. Y. Acad. Sci. 2008, 1134, 25–60. [Google Scholar] [CrossRef]
- Sandström, U.G.; Angelstam, P.; Mikusiński, G. Ecological diversity of birds in relation to the structure of urban green space. Landsc. Urban Plan 2006, 77, 39–53. [Google Scholar] [CrossRef]
- Goodness, J.; Andersson, E.; Anderson, P.M.; Elmqvist, T. Exploring the links between functional traits and cultural ecosystem services to enhance urban ecosystem management. Ecol. Indic. 2016, 70, 597–605. [Google Scholar] [CrossRef]
- Heyman, E.; Gunnarsson, B.; Dovydavicius, L. Management of urban nature and its impact on bird ecosystem services. In Ecology and Conservation of Birds in Urban Environments; Murgui, E., Hedblom, M., Eds.; Springer: Cham, Switzerland, 2017; pp. 465–488. [Google Scholar]
- Gil, D.; Brumm, H. Avian Urban Ecology, 1st ed.; Oxford University Press: Oxford, UK, 2014; pp. xiii–xv. [Google Scholar]
- Kumar, N.; Gupta, U.; Malhotra, H.; Jhala, Y.V.; Qureshi, Q.; Gosler, A.G.; Sergio, F. The population density of an urban raptor is inextricably tied to human cultural practices. Proc. R. Soc. B Biol. Sci. 2019, 286, 20182932. [Google Scholar] [CrossRef] [PubMed]
- Chace, J.F.; Walsh, J.J. Urban effects on native avifauna: A review. Landsc. Urban Plan 2006, 74, 46–69. [Google Scholar] [CrossRef]
- Palacio, F.X. Urban exploiters have broader dietary niches than urban avoiders. Ibis 2019, 162, 42–49. [Google Scholar] [CrossRef]
- Leveau, L.M. Bird traits in urban–rural gradients: How many functional groups are there? J. Ornithol. 2013, 154, 655–662. [Google Scholar] [CrossRef]
- Lizée, M.H.; Mauffrey, J.F.; Tatoni, T.; Deschamps-Cottin, M. Monitoring urban environments on the basis of biological traits. Ecol. Indic. 2011, 11, 353–361. [Google Scholar] [CrossRef]
- Rooney, T.P.; Wiegmann, S.M.; Rogers, D.A.; Waller, D.M. Biotic impoverishment and homogenization in unfragmented forest understory communities. Conserv. Biol. 2004, 18, 787–798. [Google Scholar] [CrossRef]
- Shochat, E.; Lerman, S.; Fernández-Juricic, E. Birds in urban ecosystems: Population dynamics, community structure, biodiversity, and conservation. In Urban Ecosystem Ecology, 1st ed.; Aitkenhead-Peterson, J.A., Volder, A., Eds.; American Agronomy Society: Madison, WI, USA, 2010; pp. 75–86. [Google Scholar]
- Sol, D.; González-Lagos, C.; Moreira, D.; Maspons, J.; Lapiedra, O. Urbanization tolerance and the loss of avian diversity. Ecol. Lett. 2014, 17, 942–950. [Google Scholar] [CrossRef] [PubMed]
- Blumstein, D.T. Attention, habituation, and antipredator behavior: Implications for urban birds. In Avian Urban Ecology; Diego, G., Henrik, B., Eds.; Oxford University Press: Oxford, UK, 2014; pp. 41–53. [Google Scholar]
- Bonier, F. Hormones in the city: Endocrine ecology of urban birds. Horm. Behav. 2012, 61, 763–772. [Google Scholar] [CrossRef]
- Rayner, L.; Ikin, K.; Evans, M.J.; Gibbons, P.; Lindenmayer, D.B.; Manning, A.D. Avifauna and urban encroachment in time and space. Divers. Distrib. 2015, 21, 428–440. [Google Scholar] [CrossRef]
- Leveau, L.M.; Leveau, C.M.; Villegas, M.; Cursach, J.A.; Suazo, C.G. Bird communities along urbanization gradients: A comparative analysis among three neo-tropical cities. Ornitol. Neotrop. 2017, 28, 77–87. [Google Scholar]
- la Sorte, F.A.; Lepczyk, C.A.; Aronson, M.F.J.; Goddard, M.A.; Hedblom, M.; Katti, M.; MacGregor-Fors, I. The phylogenetic and functional diversity of regional breeding bird assemblages is reduced and constricted through urbanization. Divers. Distrib. 2018, 24, 928–938. [Google Scholar] [CrossRef]
- Saari, S.; Richter, S.; Higgins, M.; Oberhofer, M.; Jennings, A.; Faeth, S.H. Urbanization is not associated with increased abundance or decreased richness of terrestrial animals-dissecting the literature through meta-analysis. Urban Ecosyst. 2016, 19, 1251–1264. [Google Scholar] [CrossRef]
- Ibáñez-Álamo, J.D.; Rubio, E.; Benedetti, Y.; Morelli, F. Global loss of avian evolutionary uniqueness in urban areas. Glob. Chang. Biol. 2017, 23, 2990–2998. [Google Scholar] [CrossRef] [PubMed]
- Batáry, P.; Kurucz, K.; Suarez-Rubio, M.; Chamberlain, D.E. Non-linearities in bird responses across urbanization gradients: A meta-analysis. Glob. Chang. Biol. 2018, 24, 1046–1054. [Google Scholar] [CrossRef]
- Morelli, F.; Benedetti, Y.; Ibáñez-Álamo, J.D.; Jokimäki, J.; Mänd, R.; Tryjanowski, P.; Møller, A.P. Evidence of evolutionary homogenization of bird communities in urban environments across Europe. Glob. Ecol. Biogeogr. 2016, 25, 1284–1293. [Google Scholar] [CrossRef]
- Hagen, O.E.; Hagen, O.; Ibáñez-Álamo, J.D.; Petchey, O.L.; Evans, K.L. Impacts of urban areas and their characteristics on avian functional diversity. Front. Ecol. Evol. 2017, 5, 84. [Google Scholar] [CrossRef]
- Shochat, E.; Warren, P.S.; Faeth, S.H.; McIntyre, N.E.; Hope, D. From patterns to emerging processes in mechanistic urban ecology. Trends Ecol. Evol. 2006, 21, 186–191. [Google Scholar] [CrossRef]
- MacGregor-Fors, I.; Morales-Pérez, L.; Schondube, J.E. Migrating to the city: Responses of neo-tropical migrant bird communities to urbanization. Condor 2010, 112, 711–717. [Google Scholar] [CrossRef]
- Leveau, L.M. Urbanization, environmental stabilization and temporal persistence of bird species: A view from Latin America. PeerJ 2018, 6, e6056. [Google Scholar] [CrossRef]
- Faeth, S.H.; Bang, C.; Saari, S. Urban biodiversity: Patterns and mechanisms. Ann. N. Y. Acad. Sci. 2011, 1223, 69–81. [Google Scholar] [CrossRef] [PubMed]
- Paton, G.D.; Shoffner, A.V.; Wilson, A.M.; Gagne, S.A. The traits that predict the magnitude and spatial scale of forest bird responses to urbanization intensity. PLoS ONE 2019, 14, e0220120. [Google Scholar] [CrossRef] [PubMed]
- Julliard, R.; Jiguet, F.; Couvet, D. Common birds facing global changes: What makes a species at risk? Glob. Chang. Biol. 2004, 10, 148–154. [Google Scholar] [CrossRef]
- Callaghan, C.T.; Major, R.E.; Wilshire, J.H.; Martin, J.M.; Kingsford, R.T.; Cornwell, W.K. Generalists are the most urban-tolerant of birds: A phylogenetically controlled analysis of ecological and life history traits using a novel continuous measure of bird responses to urbanization. Oikos 2019, 128, 845–858. [Google Scholar] [CrossRef]
- Callaghan, C.T.; Benedetti, Y.; Wilshire, J.H.; Morelli, F. Avian trait specialization is negatively associated with urban tolerance. Oikos 2020, 129, 1541–1551. [Google Scholar] [CrossRef]
- Devictor, V.; Julliard, R.; Couvet, D.; Lee, A.; Jiguet, F. Functional homogenization effect of urbanization on bird communities. Conserv. Biol. 2007, 21, 41–751. [Google Scholar] [CrossRef]
- Futuyama, D.J.; Moreno, G. The evolution of ecological specialization. Annu. Rev. Ecol. Evol. S 1988, 19, 207–233. [Google Scholar] [CrossRef]
- Kitahara, M.; Sei, K.; Fujii, K. Patterns in the structure of grassland butterfly communities along a gradient of human disturbance: Further analysis based on the generalist/specialist concept. Popul. Ecol. 2000, 42, 135–144. [Google Scholar] [CrossRef]
- Kark, S.; Iwaniuk, A.; Schalimtzek, A.; Banker, E. Living in the city: Can anyone become an ‘urban exploiter? J. Biogeogr. 2007, 34, 638–651. [Google Scholar] [CrossRef]
- Evans, K.L.; Chamberlain, D.E.; Hatchwell, B.J.; Gregory, R.D.; Gaston, K.J. What makes an urban bird? Glob. Chang. Biol. 2011, 17, 1365–2486. [Google Scholar] [CrossRef]
- Morelli, F.; Benedetti, Y.; Su, T.; Zhou, B.; Moravec, D.; Šímová, P.; Liang, W. Taxonomic diversity, functional diversity and evolutionary uniqueness in bird communities of Beijing’s urban parks: Effects of land use and vegetation structure. Urban Urban Green. 2017, 23, 84–92. [Google Scholar] [CrossRef]
- Gutiérrez-Tapia, P.; Azócar, M.I.; Castro, S.A. A citizen-based platform reveals the distribution of functional groups inside a large city from the Southern Hemisphere: E-Bird and the urban birds of Santiago (Central Chile). Rev. Chil. Hist. Nat. 2018, 91, 3. [Google Scholar] [CrossRef]
- Lim, H.C.; Sodhi, N.S. Responses of avian guilds to urbanization in a tropical city. Landsc. Urban Plan 2004, 66, 199–215. [Google Scholar] [CrossRef]
- Boal, C.W. Urban raptor communities: Why some raptors and not others occupy urban environments. In Urban Raptors; Boal, C.W., Dykstra, C.R., Eds.; Island Press: Washington, DC, USA, 2018; pp. 36–50. [Google Scholar]
- Hindmarch, S.; Elliott, J.E. A specialist in the city: The diet of barn owls along a rural to urban gradient. Urban Ecosyst. 2015, 18, 477–488. [Google Scholar] [CrossRef]
- Rullman, S.; Marzluff, J.M. Raptor presence along an urban–wildland gradient: Influences of prey abundance and land cover. J. Raptor Res. 2014, 48, 257–272. [Google Scholar] [CrossRef]
- Thomson, V.K.; Stevens, T.; Jones, D.; Huijbers, C. Carrion preference in Australian coastal raptors: Effects of Urbanisation on scavenging. Sunbird 2016, 46, 16. [Google Scholar]
- Jones, D.N.; Reynolds, J.S. Feeding birds in our towns and cities: A global research opportunity. J. Avian Biol. 2008, 39, 265–271. [Google Scholar] [CrossRef]
- Hodgson, P.; French, K.; Major, R.E. Comparison of foraging behavior of small, urban-sensitive insectivores in continuous woodland and woodland remnants in a suburban landscape. Wildl. Res. 2006, 33, 591–603. [Google Scholar] [CrossRef]
- Marzluff, J.M.; Bowman, R.; Donnelly, R. (Eds.) A historical perspective on urban bird research: Trends, terms, and approaches. In Avian Ecology and Conservation in an Urbanizing; World Springer: Boston, MA, USA, 2001; pp. 1–17. [Google Scholar]
- Ikin, K.; Knight, E.; Lindenmayer, D.B.; Fischer, J.; Manning, A.D. Linking bird species traits to vegetation characteristics in a future urban development zone: Implications for urban planning. Urban Ecosyst. 2012, 15, 961–977. [Google Scholar] [CrossRef]
- Croci, S.; Butet, A.; Clergeau, P. Does urbanization filter birds on the basis of their biological traits. Condor 2008, 110, 223–240. [Google Scholar] [CrossRef]
- Reale, J.A.; Blair, R.B. Nesting success and life-history attributes of bird communities along an urbanization gradient. Urban Habitats 2005, 3, 1–24. [Google Scholar]
- Máthé, O.; Batáry, P. Insectivorous and open-cup nester bird species suffer the most from urbanization. Bird Study 2015, 62, 78–86. [Google Scholar] [CrossRef]
- Dykstra, C.R. City Lifestyles: Behavioral Ecology of Urban Raptors. In Urban Raptors; Boal, C.W., Dykstra, C.R., Eds.; Island Press: Washington, DC, USA, 2018; pp. 18–35. [Google Scholar]
- Gahbauer, M.A.; Bird, D.M.; Clark, K.E.; French, T.; Brauning, D.W.; Mcmorris, F.A. Productivity, mortality, and management of urban peregrine falcons in northeastern North America. J. Wildl. Manag. 2015, 79, 10–19. [Google Scholar] [CrossRef]
- Mason, C.F. Avian species richness and numbers in the built environment: Can new housing developments be good for birds? In Human Exploitation and Biodiversity Conservation; Hawksworth, D.L., Bull, A.T., Eds.; Springer: Dordrecht, The Netherlands, 2006; pp. 2365–2378. [Google Scholar]
- Thornton, M.; Todd, I.; Roos, S. Breeding success and productivity of urban and rural Eurasian Sparrowhawks Accipiter nisus in Scotland. Écoscience 2017, 24, 115–126. [Google Scholar] [CrossRef]
- Partecke, J.; Van’t-Hof, T.; Gwinner, E. Differences in the timing of reproduction between urban and forest European blackbirds (Turdus merula): Result of phenotypic flexibility or genetic differences? Proc. R. Soc. B Biol. Sci. 2004, 271, 1995–2001. [Google Scholar] [CrossRef]
- Atwell, J.W.; Cardoso, G.; Whittaker, D.J.; Price, T.D.; Ketterson, E.D. Hormonal, behavioral, and life-history traits exhibit correlated shifts in relation to population establishment in a novel environment. Am. Nat. 2014, 184, E147–E160. [Google Scholar] [CrossRef]
- Zhang, S.; Chen, X.; Zhang, J.; Li, H. Differences in the reproductive hormone rhythm of Tree sparrows (Passer montanus) from urban and rural sites in Beijing: The effect of anthropogenic light sources. Gen. Comp. Endocrinol. 2014, 206, 24–29. [Google Scholar] [CrossRef]
- Davies, S.; Behbahaninia, H.; Giraudeau, M.; Meddle, S.L.; Waites, K.; Deviche, P. Advanced seasonal reproductive development in a male urban bird is reflected in earlier plasma luteinizing hormone rise but not energetic status. Gen. Comp. Endocrinol. 2015, 224, 1–10. [Google Scholar] [CrossRef]
- Kettel, E.F.; Gentle, L.K.; Quinn, J.L.; Yarnell, R.W. The breeding performance of raptors in urban landscapes: A review and meta-analysis. J. Ornithol. 2018, 159, 1–18. [Google Scholar] [CrossRef]
- Chamberlain, D.E.; Cannon, A.R.; Toms, M.P.; Leech, D.I.; Hatchwell, B.J.; Gaston, K.J. Avian productivity in urban landscapes: A review and meta-analysis. Ibis 2009, 151, 1–18. [Google Scholar] [CrossRef]
- Marini, K.L.D.; Otter, K.A.; LaZerte, S.E.; Reudink, M.W. Urban environments are associated with earlier clutches and faster nestling feather growth compared to natural habitats. Urban Ecosyst. 2017, 20, 1291–1300. [Google Scholar] [CrossRef]
- Antonov, A.; Atanasova, D. Small-scale differences in the breeding ecology of urban and rural magpies (Pica pica). Ornis Fennica 2003, 80, 21–30. [Google Scholar]
- Shochat, E.; Lerman, S.B.; Katti, M.; Lewis, D.B. Linking optimal foraging behavior to bird community structure in an urban-desert landscape: Field experiments with artificial food patches. Am. Nat. 2004, 164, 232–243. [Google Scholar] [CrossRef] [PubMed]
- Caizergues, A.E.; Grégoire, A.; Charmantier, A. Urban versus forest ecotypes are not explained by divergent reproductive selection. Proc. R. Soc. B Biol. Sci. 2018, 285, 20180261. [Google Scholar] [CrossRef]
- Mennechez, G.; Clergeau, P. Effect of urbanization on habitat generalists: Starlings not so flexible? Acta Oecol. 2006, 30, 182–191. [Google Scholar] [CrossRef]
- Liker, A.; Papp, Z.; Bókony, V.; Lendvai, A.Z. Lean birds in the city: Body size and condition of house sparrows along the urbanization gradient. J. Anim. Ecol. 2008, 77, 789–795. [Google Scholar] [CrossRef]
- Auman, H.J.; Meathrel, C.E.; Richardson, A. Supersize me: Does anthropogenic food change the body condition of Silver Gulls? A comparison between urbanized and remote, non-urbanized areas. Waterbirds 2008, 31, 122–126. [Google Scholar] [CrossRef]
- Bókony, V.; Seress, G.; Nagy, S.; Lendvai, A.Z.; Liker, A. Multiple indices of body condition reveal no negative effect of urbanization in adult house sparrows. Landsc. Urban Plan 2012, 104, 75–84. [Google Scholar] [CrossRef]
- Leveau, L.M. Urbanization induces bird color homogenization. Landsc. Urban Plan 2019, 192, 103645. [Google Scholar] [CrossRef]
- Sol, D.; Bacher, S.; Reader, S.M.; Lefebvre, L. Brain size predicts the success of mammal species introduced into novel environments. Am. Nat. 2008, 172, 63–71. [Google Scholar] [CrossRef]
- Maklakov, A.A.; Immler, S.; Gonzalez-Voyer, A.; Rönn, J.; Kolm, N. Brains and the city: Big-brained passerine birds succeed in urban environments. Biol. Lett. 2011, 7, 730–732. [Google Scholar] [CrossRef] [PubMed]
- Carrete, M.; Tella, J.L. Inter-individual variability in fear of humans and relative brain size of the species are related to contemporary urban invasion in birds. PLoS ONE 2011, 6, e18859. [Google Scholar] [CrossRef] [PubMed]
- Møller, A.P. Behavioural and ecological predictors of urbanization. In Avian Urban Ecology; Gil, D., Brumm, H., Eds.; Oxford University Press: Oxford, UK, 2014; pp. 54–68. [Google Scholar]
- Møller, A.P.; Erritzøe, J. Brain size and urbanization in birds. Avian Res. 2015, 6, 8. [Google Scholar] [CrossRef]
- Grunst, M.L.; Rotenberry, J.T.; Grunst, A.S. Variation in adrenocortical stress physiology and condition metrics within a heterogeneous urban environment in the Song sparrow (Melospiza melodia). J. Avian Biol. 2014, 45, 574–583. [Google Scholar] [CrossRef]
- Davies, S.; Haddad, N.; Ouyang, J.Q. Stressful city sounds: Glucocorticoid responses to experimental traffic noise are environmentally dependent. Biol. Lett. 2017, 13, 20170276. [Google Scholar] [CrossRef] [PubMed]
- Russ, A.; Reitemeier, S.; Weissmann, A.; Gottschalk, J.; Einspanier, A.; Klenke, R. Seasonal and urban effects on the endocrinology of a wild passerine. Ecol. Evol. 2015, 5, 5698–5710. [Google Scholar] [CrossRef]
- Dominoni, D.M. The effects of light pollution on biological rhythms of birds: An integrated, mechanistic perspective. J. Ornithol. 2015, 156, 409–418. [Google Scholar] [CrossRef]
- Partecke, J.; Schwabl, I.; Gwinner, E. Stress and the city: Urbanization and its effects on the stress physiology in European blackbirds. Ecology 2006, 87, 1945–1952. [Google Scholar] [CrossRef]
- Fokidis, H.B.; Deviche, P. Plasma corticosterone of city and desert Curve-billed Thrashers, (Toxostoma curvirostre), in response to stress-related peptide administration. Comp. Biochem. Physiol. A Mol. Integr. Physiol. 2011, 159, 32–38. [Google Scholar] [CrossRef]
- Zhang, S.; Lei, F.; Liu, S.; Li, D.; Chen, C.; Wang, P. Variation in baseline corticosterone levels of Tree sparrow (Passer montanus) populations along an urban gradient in Beijing, China. J. Ornithol. 2011, 152, 801–806. [Google Scholar] [CrossRef]
- Davies, S.; Rodriguez, N.S.; Sweazea, K.L.; Deviche, P. The effect of acute stress and long-term corticosteroid administration on plasma metabolites in an urban and Desert songbird. Physiol. Biochem. Zool. 2013, 86, 47–60. [Google Scholar] [CrossRef] [PubMed]
- Fokidis, H.B.; Deviche, P. Brain arginine vasotocin immunoreactivity differs between urban and desert curve-billed thrashers, (Toxostoma curvirostre): Relationships with territoriality and stress physiology. Brain Behav. Evol. 2012, 79, 84–97. [Google Scholar] [CrossRef] [PubMed]
- Yeh, P.J.; Price, T.D. Adaptive phenotypic plasticity and the successful colonization of a novel environment. Am. Nat. 2004, 164, 531–542. [Google Scholar] [CrossRef] [PubMed]
- Rodewald, A.D.; Shustack, D.P. Urban flight: Understanding individual and population-level responses of Nearctic–Neotropical migratory birds to urbanization. J. Anim. Ecol. 2008, 77, 83–91. [Google Scholar] [CrossRef] [PubMed]
- Schoech, S.J.; Bowman, R.; Hahn, T.P.; Goymann, W.; Schwabl, I.; Bridge, E.S. The effects of low levels of light at night upon the endocrine physiology of Western scrub-jays (Aphelocoma californica). J. Exp. Zool. A Ecol. Integr. Physiol. 2013, 319, 527–538. [Google Scholar] [CrossRef] [PubMed]
- Brown, L.M.; Graham, C.H. Demography, traits and vulnerability to urbanization: Can we make generalizations? J. Appl. Ecol. 2015, 52, 1455–1464. [Google Scholar] [CrossRef]
- Iglesias-Carrasco, M.; Duchêne, D.A.; Head, M.L.; Møller, A.P.; Cain, K. Sex in the city: Sexual selection and urban colonization in passerines. Biol. Lett. 2019, 15, 20190257. [Google Scholar] [CrossRef]
- Dominoni, D.M.; Quetting, M.; Partecke, J. Long-term effects of chronic light pollution on seasonal functions of European blackbirds (Turdus merula). PLoS ONE 2013, 8, e85069. [Google Scholar] [CrossRef]
- Dominoni, D.M.; Quetting, M.; Partecke, J. Artificial light at night advances avian reproductive physiology. Proc. R. Soc. B Biol. Sci. 2013, 280, 20123017. [Google Scholar] [CrossRef]
- Fudickar, A.M.; Greives, T.J.; Abolins-Abols, M.; Atwell, J.W.; Meddle, S.L.; Friis, G.; Stricker, C.A.; Ketterson, E.D. Mechanisms associated with an advance in the timing of seasonal reproduction in an urban songbird. Front. Ecol. Evol. 2017, 5, 85. [Google Scholar] [CrossRef]
- Partecke, J.; Gwinner, E. Increased sedentariness in European blackbirds following urbanization: A consequence of local adaptation? Ecology 2007, 88, 882–890. [Google Scholar] [CrossRef] [PubMed]
- Longcore, T. Sensory ecology: Night lights alter reproductive behavior of Blue tits. Curr. Biol. 2010, 20, R893–R895. [Google Scholar] [CrossRef] [PubMed]
- Dominoni, D.M.; Goymann, W.; Helm, B.; Partecke, J. Urban-like night illumination reduces melatonin release in European blackbirds (Turdus merula): Implications of city life for biological time-keeping of songbirds. Front. Zool. 2013, 10, 60. [Google Scholar] [CrossRef] [PubMed]
- Reynolds, S.J.; Ibáñez-Álamo, J.D.; Sumasgutner, P.; Mainwaring, M.C. Urbanization and nest building in birds: A review of threats and opportunities. J. Ornithol. 2019, 160, 841–860. [Google Scholar] [CrossRef]
- Wawrzyniak, J.; Kaliński, A.; Glądalski, M.; Bańbura, M.; Markowski, M.; Skwarska, J.; Kaliński, P.; Cyżewska, I.; Bańbura, J. Long-term variation in laying date and clutch size of the Great tit (Parus major) in central Poland: A comparison between urban parkland and deciduous forest. Ardeola 2015, 62, 311–323. [Google Scholar] [CrossRef]
- Møller, A.P. Successful city dwellers: A comparative study of the ecological characteristics of urban birds in the Western Palearctic. Oecologia 2009, 159, 849–858. [Google Scholar] [CrossRef] [PubMed]
- Isaksson, C. Urbanization, oxidative stress and inflammation: A question of evolving, acclimatizing or coping with urban environmental stress. Funct. Ecol. 2015, 29, 913–923. [Google Scholar] [CrossRef]
- Watson, H.; Videvall, E.; Andersson, M.N.; Isaksson, C. Transcriptome analysis of a wild bird reveals physiological responses to the urban environment. Sci. Rep. 2017, 7, 44180. [Google Scholar] [CrossRef]
- Costantini, D.; Greives, T.J.; Hau, M.; Partecke, J. Does urban life change blood oxidative status in birds? J. Exp. Biol. 2014, 217, 2994–2997. [Google Scholar] [CrossRef]
- Fuller, R.A.; Warren, P.H.; Gaston, K.J. Daytime noise predicts nocturnal singing in urban robins. Biol. Lett. 2007, 3, 368–370. [Google Scholar] [CrossRef]
- Nemeth, E.; Brumm, H. Birds and anthropogenic noise: Are urban songs adaptive? Am. Nat. 2010, 176, 465–475. [Google Scholar] [CrossRef] [PubMed]
- Slabbekoorn, H.; Ripmeester, E.A.P. Birdsong and anthropogenic noise: Implications and applications for conservation. Mol. Ecol. 2008, 17, 72–83. [Google Scholar] [CrossRef] [PubMed]
- Slabbekoorn, H.; Boer-Visser, A. Cities change the songs of birds. Curr. Biol. 2006, 16, 2326–2331. [Google Scholar] [CrossRef] [PubMed]
- Laiolo, P. The Rufous-Collared sparrow (Zonotrichia capensis) utters higher frequency songs in urban habitats. Rev. Catalana d’Ornitologia 2011, 27, 25–30. [Google Scholar]
- Luther, D.A.; Derryberry, E.P. Birdsongs keep pace with city life: Changes in song over time in an urban songbird affects communication. Anim. Behav. 2012, 83, 1059–1066. [Google Scholar] [CrossRef]
- Job, J.R.; Kohler, S.L.; Gill, S.A. Song adjustments by an open habitat bird to anthropogenic noise, urban structure, and vegetation. Behav. Ecol. 2016, 27, 1734–1744. [Google Scholar] [CrossRef]
- LaZerte, S.E.; Otter, K.A.; Slabbekoorn, H. Mountain chickadees adjust songs, calls and chorus composition with increasing ambient and experimental anthropogenic noise. Urban Ecosyst. 2017, 20, 989–1000. [Google Scholar] [CrossRef]
- Cardoso, G.C.; Hu, Y.; Francis, C.D. The comparative evidence for urban species sorting by anthropogenic noise. R Soc. Open Sci. 2018, 5, 172059. [Google Scholar] [CrossRef]
- Moiron, M.; González-Lagos, C.; Slabbekoorn, H.; Sol, D. Singing in the city: High song frequencies are no guarantee for urban success in birds. Behav. Ecol. 2015, 26, 843–850. [Google Scholar] [CrossRef]
- Luther, D.A.; Phillips, J.; Derryberry, E.P. Not so sexy in the city: Urban birds adjust songs to noise but compromise vocal performance. Behav. Ecol. 2016, 27, 332–340. [Google Scholar] [CrossRef]
- Narango, D.L.; Rodewald, A.D. Urban-associated drivers of song variation along a rural–urban gradient. Behav. Ecol. 2016, 27, 608–616. [Google Scholar] [CrossRef]
- Phillips, J.N.; Derryberry, E.P. Urban sparrows respond to a sexually selected trait with increased aggression in noise. Sci. Rep. 2018, 8, 7505. [Google Scholar] [CrossRef] [PubMed]
- Ríos-Chelén, A.A.; Quirós-Guerrero, E.; Gil, D.; Macías-Garcia, C. Dealing with urban noise: Vermilion flycatchers sing longer songs in noisier territories. Behav. Ecol. Sociobiol. 2013, 67, 145–152. [Google Scholar] [CrossRef]
- Lowry, H.; Lill, A.; Wong, B.B.M. How noisy Does a noisy miner have to be? Amplitude adjustments of alarm calls in an avian urban ‘adapter’. PLoS ONE 2011, 7, e29960. [Google Scholar] [CrossRef] [PubMed]
- Potvin, D.A.; Mulder, R.A. Immediate, independent adjustment of call pitch and amplitude in response to varying background noise by Silvereyes (Zosterops lateralis). Behav. Ecol. 2013, 24, 1363–1368. [Google Scholar] [CrossRef]
- Knight, C.R.; Swaddle, J.P. Eastern Bluebirds alter their song in response to anthropogenic changes in the acoustic environment. Integr. Comp. Biol. 2015, 55, 418–431. [Google Scholar] [CrossRef]
- Díaz, M.; Parra, A.; Gallardo, C. Serins respond to anthropogenic noise by increasing vocal activity. Behav. Ecol. 2011, 22, 332–336. [Google Scholar] [CrossRef]
- Redondo, P.; Barrantes, G.; Sandoval, L. Urban noise influences vocalization structure in the House wren (Troglodytes aedon). Ibis 2013, 155, 621–625. [Google Scholar] [CrossRef]
- Moseley, D.L.; Phillips, J.N.; Derryberry, E.P.; Luther, D.A. Evidence for differing trajectories of songs in urban and rural populations. Behav. Ecol. 2019, 30, 1734–1742. [Google Scholar] [CrossRef]
- Potvin, D.A.; Mulder, R.A.; Parris, K.M. Silvereyes decrease acoustic frequency but increase efficacy of alarm calls in urban noise. Anim. Behav. 2014, 98, 27–33. [Google Scholar] [CrossRef]
- Potvin, D.A.; Parris, K.M.; Mulder, R.A. Geographically pervasive effects of urban noise on frequency and syllable rate of songs and calls in Silvereyes (Zosterops lateralis). Proc. R. Soc. B Biol. Sci. 2011, 278, 2464–2469. [Google Scholar] [CrossRef] [PubMed]
- Seger-Fullam, K.D.; Rodewald, A.D.; Soha, J.A. Urban noise predicts song frequency in Northern cardinals and American robins. Bioacoustics 2011, 20, 267–276. [Google Scholar] [CrossRef]
- Shannon, G.; McKenna, M.F.; Angeloni, L.M.; Crooks, K.R.; Fristrup, K.M.; Brown, E.; Warner, K.A.; Nelson, M.D.; White, C.; Briggs, J.; et al. A synthesis of two decades of research documenting the effects of noise on wildlife. Biol. Rev. 2016, 91, 982–1005. [Google Scholar] [CrossRef] [PubMed]
- Fernández-Juricic, E.; Poston, R.; Collibus, K.D.; Morgan, T.; Bastain, B.; Martin, C.; Jones, K.; Treminio, R. Microhabitat selection and singing behavior patterns of male house finches (Haemorhous mexicanus) in urban parks in a heavily urbanized landscape in the western U.S. Urban Habitats 2005, 3, 49–69. [Google Scholar]
- Badyaev, A.V.; Young, R.L.; Oh, K.P.; Addison, C. Evolution on a local scale: Developmental, functional, and genetic bases of divergence in bill form and associated changes in song structure between adjacent habitats. Evolution 2008, 62, 1951–1964. [Google Scholar] [CrossRef] [PubMed]
- Nordt, A.; Klenke, R. Sleepless in town—Drivers of the temporal shift in dawn song in urban European blackbirds. PLoS ONE 2013, 8, e71476. [Google Scholar] [CrossRef]
- Dorado-Correa, A.M.; Rodríguez-Rocha, M.; Brumm, H. Anthropogenic noise, but not artificial light levels predict song behavior in an equatorial bird. R Soc. Open Sci. 2016, 3, 160231. [Google Scholar] [CrossRef]
- Kempenaers, B.; Borgström, P.; Loës, P.; Schlicht, E.; Valcu, M. Artificial night lighting affects dawn song, extra-pair siring success, and lay date in songbirds. Curr. Biol. 2010, 20, 1735–1739. [Google Scholar] [CrossRef]
- Randler, C. Disturbances by dog barking increase vigilance in coots Fulica atra. Eur. J. Wildl. Res. 2006, 52, 265–270. [Google Scholar] [CrossRef]
- Bonnington, C.; Gaston, K.J.; Evans, K.L. Fearing the feline: Domestic cats reduce avian fecundity through trait-mediated indirect effects that increase nest predation by other species. J. Appl. Ecol. 2013, 50, 15–24. [Google Scholar] [CrossRef]
- Rebolo-Ifrán, N.; Carrete, M.; Sanz-Aguilar, A.; Rodríguez-Martínez, S.; Cabezas, S.; Marchant, T.A.; Bortolotti, G.R.; Tella, J.L. Links between fear of humans, stress and survival support a non-random distribution of birds among urban and rural habitats. Sci. Rep. 2015, 5, 13723. [Google Scholar] [CrossRef] [PubMed]
- Samia, D.S.M.; Nakagawa, S.; Nomura, F.; Rangel, T.F.; Blumstein, D.T. Increased tolerance to humans among disturbed wildlife. Nat. Commun. 2015, 6, 8877. [Google Scholar] [CrossRef] [PubMed]
- Eötvösa, C.B.; Magura, T.; Lövei, G.L. A meta-analysis indicates reduced predation pressure with increasing urbanization. Landsc. Urban Plan 2018, 180, 54–59. [Google Scholar] [CrossRef]
- Weaver, M.; Ligon, R.A.; Mousel, M.; McGraw, K.J. Avian anthrophobia? Behavioral and physiological responses of house finches (Haemorhous mexicanus) to human and predator threats across an urban gradient. Landsc. Urban Plan 2018, 179, 46–54. [Google Scholar] [CrossRef]
- McGiffin, A.; Lill, A.; Beckman, J.; Johnstone, C.P. Tolerance of human approaches by Common mynas along an urban-rural gradient. EMU Austral Ornithol. 2013, 113, 154–160. [Google Scholar] [CrossRef]
- Møller, A.P.; Grim, T.; Ibáñez-Álamo, J.D.; Markó, G.; Tryjanowski, P. Change in flight initiation distance between urban and rural habitats following a cold winter. Behav. Ecol. 2013, 24, 1211–1217. [Google Scholar] [CrossRef]
- Møller, A.P.; Díaz, M.; Flensted-Jensen, E.; Grim, T.; Ibáñez-Álamo, J.D.; Jokimäki, J.; Mänd, R.; Markó, G.; Tryjanowski, P. Urbanized birds have superior establishment success in novel environments. Oecologia 2015, 178, 943–950. [Google Scholar] [CrossRef]
- Vines, A.; Lill, A. Boldness and urban dwelling in little ravens. Wildl. Res. 2014, 42, 590–597. [Google Scholar] [CrossRef]
- Gravolin, I.; Key, M.; Lill, A. Boldness of urban Australian magpies and local traffic volume. Avian Biol. Res. 2014, 7, 244–250. [Google Scholar] [CrossRef]
- Samia, D.S.M.; Blumstein, D.T.; Díaz, M.; Grim, T.; Ibáñez-Álamo, J.D.; Jokimäki, J.; Tätte, K.; Markó, G.; Tryjanowski, P.; Møller, A.P. Rural-urban differences in escape behavior of European birds across a latitudinal gradient. Front. Ecol. Evol. 2017, 5, 66. [Google Scholar] [CrossRef]
- Morelli, F.; Mikula, P.; Benedetti, Y.; Bussière, R.; Jerzak, L.; Tryjanowski, P. Escape behavior of birds in urban parks and cemeteries across Europe: Evidence of behavioral adaptation to human activity. Sci. Total Environ. 2018, 631, 803–810. [Google Scholar] [CrossRef] [PubMed]
- Biondi, L.M.; Fuentes, G.M.; Córdoba, R.S.; Bó, M.S.; Cavalli, M.; Paterlini, C.A.; Castano, M.V.; García, G.O. Variation in boldness and novelty response between rural and urban predatory birds: The Chimango Caracara (Milvago chimango) as study case. Behav. Process 2020, 173, 104064. [Google Scholar] [CrossRef] [PubMed]
- Fleming, P.A.; Bateman, P.W. Scavenging opportunities modulate escape responses over a small geographic scale. Ethology 2017, 123, 205–212. [Google Scholar] [CrossRef]
- Feng, C.; Liang, W. Behavioral responses of black-headed gulls (Chroicocephalus ridibundus) to artificial provisioning in China. Glob. Ecol. Conserv. 2020, 21, e00873. [Google Scholar] [CrossRef]
- Pavlova, O.; Wronski, T. City gulls and their rural neighbours: Changes in escape and agonistic behaviour along a rural-to-urban gradient. In Advances in Environmental Research; Nova Science Publishers, Inc.: Hauppauge, NY, USA, 2020. [Google Scholar]
- Møller, A.P. Interspecific variation in fear responses predicts urbanization in birds. Behav. Ecol. 2010, 21, 365–371. [Google Scholar] [CrossRef]
- Kitchen, K.; Lill, A.; Price, M. Tolerance of human disturbance by urban magpie-larks. Aust. Field Ornithol. 2010, 27, 1. [Google Scholar]
- Gendall, J.; Lill, A.; Beckman, J. Tolerance of disturbance by humans in long-time resident and recent colonist urban doves. Avian Res. 2015, 6, 7. [Google Scholar] [CrossRef]
- Sol, D.; Maspons, J.; Gonzalez-Voyer, A.; Morales-Castilla, I.; Garamszegi, L.Z.; Møller, A.P. Risk-taking behavior, urbanization and the pace of life in birds. Behav. Ecol. Sociobiol. 2018, 72, 59–66. [Google Scholar] [CrossRef]
- Sol, D.; Griffin, A.S.; Bartomeus, I.; Boyce, H. Exploring or avoiding novel food resources? The novelty conflict in an invasive bird. PLoS ONE 2011, 6, e19535. [Google Scholar] [CrossRef]
- Shanahan, D.F.; Strohbach, M.W.; Warren, P.S.; Fuller, R.A. The challenges of urban living. In Avian Urban Ecology; Diego, G., Henrik, B., Eds.; Oxford University Press: Oxford, UK, 2014; pp. 3–20. [Google Scholar]
- Griffin, A.S.; Netto, K.; Peneaux, C. Neophilia, innovation and learning in an urbanized world: A critical evaluation of mixed findings. Curr. Opin. Behav. Sci. 2017, 17, 15–22. [Google Scholar] [CrossRef]
- Seress, G.; Bókony, V.; Heszberger, J.; Liker, A. Response to predation risk in urban and rural house sparrows. Ethology 2011, 117, 896–907. [Google Scholar] [CrossRef]
- Møller, A.P.; Ibáñez-Álamo, J.D. Escape behavior of birds provides evidence of predation being involved in urbanization. Anim. Behav. 2012, 84, 341–348. [Google Scholar] [CrossRef]
- Tryjanowski, P.; Møller, A.P.; Morelli, F.; Biaduń, W.; Brauze, T.; Ciach, M.; Czechowski, P.; Czyż, S.; Dulisz, B.; Goławski, A.; et al. Urbanization affects neophilia and risk-taking at bird-feeders. Sci. Rep. 2016, 6, 28575. [Google Scholar] [CrossRef] [PubMed]
- Audet, J.N.; Ducatez, S.; Lefebvre, L. The town bird and the country bird: Problem solving and immune-competence vary with urbanization. Behav. Ecol. 2016, 27, 637–644. [Google Scholar] [CrossRef]
- Papp, S.; Vincze, E.; Preiszner, B.; Liker, A.; Bókony, V. A comparison of problem-solving success between urban and rural house sparrows. Behav. Ecol. Sociobiol. 2015, 9, 471–480. [Google Scholar] [CrossRef]
- Ducatez, S.; Audet, J.N.; Ros, J.; Kayello, R.L.; Lefebvre, L. Innovativeness and the effects of urbanization on risk-taking behaviors in wild Barbados birds. Anim. Cogn. 2017, 20, 33–42. [Google Scholar] [CrossRef]
- Spelt, A.; Soutar, O.; Williamson, C.; Memmott, J.; Shamoun-Baranes, J.; Rock, P.; Windsor, S. Urban gulls adapt foraging schedule to human-activity patterns. Ibis 2020, 163, 274–282. [Google Scholar] [CrossRef]
- Yosef, R.; Zduniak, P.; Poliakov, Y.; Fingerman, A. Behavioural and reproductive flexibility of an invasive bird in an arid zone: A case of the Indian House Crow (Corvus splendens). J. Arid Environ. 2019, 168, 56–58. [Google Scholar] [CrossRef]
- Bókony, V.; Kulcsár, A.; Tóth, Z.; Liker, A. Personality traits and behavioral syndromes in differently urbanized populations of house sparrows (Passer domesticus). PLoS ONE 2012, 7, e36639. [Google Scholar] [CrossRef]
- Kozlovsky, D.Y.; Weissgerber, E.A.; Pravosudov, V.V. What makes specialized food-caching mountain chickadees successful city slickers? Proc. R. Soc. B Biol. Sci. 2017, 284, 20162613. [Google Scholar] [CrossRef]
- Greggor, A.L.; Clayton, N.S.; Fulford, A.J.C.; Thornton, A. Street smart: Faster approach towards litter in urban areas by highly neophobic corvids and less fearful birds. Anim. Behav. 2016, 117, 123–133. [Google Scholar] [CrossRef]
- Lill, A.; Hales, E. Behavioral and ecological keys to urban colonization by Little Ravens (Corvus mellori). Open Ornithol. J. 2015, 8, 22–31. [Google Scholar] [CrossRef]
- Campbell, M. An animal geography of avian feeding habits in Peterborough, Ontario. Area 2008, 40, 472–480. [Google Scholar] [CrossRef]
- Evans, J.; Boudreau, K.; Hyman, J. Behavioral syndromes in urban and rural populations of Song sparrows. Ethology 2010, 116, 588–595. [Google Scholar]
- Scales, J.; Hyman, J.; Hughes, M. Behavioral syndromes break down in urban Song sparrow populations. Ethology 2011, 117, 887–895. [Google Scholar] [CrossRef]
- Hasegawa, M.; Ligon, R.A.; Giraudeau, M.; Watanabe, M.; McGraw, K.J. Urban and colorful male house finches are less aggressive. Behav. Ecol. 2014, 25, 641–649. [Google Scholar] [CrossRef]
- Hardman, S.I.; Dalesman, S. Repeatability and degree of territorial aggression differs among urban and rural Great tits (Parus major). Sci. Rep. 2018, 8, 5042. [Google Scholar] [CrossRef]
- Galbreath, D.M.; Ichinose, T.; Furutani, T.; Yan, W.; Higuchi, H. Urbanization and its implications for avian aggression: A case study of urban black kites (Milvus migrans) along Sagami Bay in Japan. Landsc. Ecol. 2014, 29, 169–178. [Google Scholar] [CrossRef]
- McClelland, S.C.; Ribeiro, R.D.; Mielke, H.W.; Finkelstein, M.E.; Gonzales, C.R.; Jones, J.A.; Komdeur, J.; Derryberry, E.; Saltzberg, E.B.; Karubian, J. Sub-lethal exposure to lead is associated with heightened aggression in an urban songbird. Sci. Total Environ. 2019, 654, 593–603. [Google Scholar] [CrossRef]
- Kunca, T.; Yosef, R. Differential nest-defense to perceived danger in urban and rural areas by female Eurasian sparrowhawk (Accipiter nisus). PeerJ 2016, 4, e2070. [Google Scholar] [CrossRef]
- Senar, J.C.; Garamszegi, L.Z.; Tilgar, V.; Biard, C.; Moreno-Rueda, G.; Salmón, P.; Rivas, J.; Sprau, P.; Dingemanse, N.J.; Charmantier, A.; et al. Urban Great tits (Parus major) show higher distress calling and pecking rates than rural birds across Europe. Front. Ecol. Evol. 2017, 5, 163. [Google Scholar] [CrossRef]
- Minias, P.; Jedlikowski, J.; Włodarczyk, R. Development of urban behavior is associated with time since urbanization in a reed-nesting waterbird. Urban Ecosyst. 2018, 21, 1021–1028. [Google Scholar] [CrossRef]
- Alberti, M.; Marzluff, J.; Hunt, V.M. Urban driven phenotypic changes: Empirical observations and theoretical implications for eco-evolutionary feedback. Philos. Trans. R. Soc. B 2017, 372, 20160029. [Google Scholar] [CrossRef]
- Capilla-Lasheras, P.; Dominoni, D.M.; Babayan, S.A.; O’Shaughnessy, P.J.; Mladenova, M.; Woodford, L.; Pollock, C.J.; Barr, T.; Baldini, F.; Helm, B. Elevated immune gene expression is associated with poor reproductive success of urban Blue tits. Front. Ecol. Evol. 2017, 5, 64. [Google Scholar] [CrossRef]
- Mueller, J.C.; Partecke, J.; Hatchwell, B.J.; Gaston, K.J.; Evans, K.L. Candidate gene polymorphisms for behavioral adaptations during urbanization in blackbirds. Mol. Ecol. 2013, 22, 3629–3637. [Google Scholar] [CrossRef]
- Partecke, J.; Gwinner, E.; Bensch, S. Is urbanization of European blackbirds (Turdus merula) associated with genetic differentiation? J. Ornithol. 2006, 147, 549–552. [Google Scholar] [CrossRef]
- Richards, D.R.; Passy, P.; Oh, R.R. Impacts of population density and wealth on the quantity and structure of urban green space in tropical Southeast Asia. Landsc. Urban Plan 2017, 157, 553–560. [Google Scholar] [CrossRef]
- Harper, G.A.; Bunbury, N. Invasive rats on tropical islands: Their population biology and impacts on native species. Glob. Ecol. Conserv. 2015, 3, 607–627. [Google Scholar] [CrossRef]
- Ferenc, M.; Sedláček, O.; Fuchs, R. How to improve urban greenspace for woodland birds: Site and local-scale determinants of bird species richness. Urban Ecosyst. 2014, 17, 625–640. [Google Scholar] [CrossRef]
- Moreno-Mateos, D.; Rey Benayas, J.M.; Pérez-Camacho, L.; Montaña, E.D.L.; Rebollo, S.; Cayuela, L. Effects of land use on nocturnal birds in a Mediterranean agricultural landscape. Acta Ornithol. 2011, 46, 173–182. [Google Scholar] [CrossRef]





{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Trait Studied | Number of Studies (<4 Species Studied) | Number of Studies (>4 Species Studied) | Total |
|---|---|---|---|
| Ecological | |||
| Diversity | 1 | 14 | 15 |
| Nesting spaces | 7 | 9 | 16 |
| Abundance | 8 | 8 | |
| Richness | 10 | 10 | |
| Diet breadth | 1 | 9 | 10 |
| Generalist-specialist | 5 | 7 | 12 |
| Distribution pattern | 1 | 3 | 4 |
| Density | 3 | 3 | |
| Migration | 4 | 3 | 7 |
| Habitat dependence | 1 | 4 | 5 |
| Dispersal | 1 | 1 | |
| Sociality and sedentariness | 1 | 1 | |
| Community composition | 1 | 1 | |
| Habitat preference | 2 | 2 | |
| Life history | |||
| Reproduction cycle | 3 | 2 | 5 |
| Clutch size | 5 | 4 | 9 |
| Nesting success | 4 | 2 | 6 |
| Breeding timing and/or performance | 2 | 1 | 3 |
| Brood size | 2 | 2 | 4 |
| Fecundity and adult survival | 2 | 1 | 3 |
| Activity time range | 1 | 1 | |
| Migration | 1 | 1 | |
| Fitness | 1 | 1 | |
| Physiological | |||
| Reproductive physiology, circadian rhythm and migration | 12 | 12 | 24 |
| Brain size | 1 | 5 | 6 |
| Body size and/or mass | 5 | 4 | 9 |
| Inflammatory response, oxidative stress | 1 | 2 | 3 |
| Stress physiology | 10 | 1 | 11 |
| Endocrine traits | 1 | 1 | |
| Bill length | 2 | 2 | |
| Plumage coloration | 1 | 1 | 2 |
| Behavioral | |||
| Fear and stress responses | 17 | 10 | 27 |
| Foraging behavior innovation | 6 | 6 | 12 |
| Innovation, learning and problem-solving ability, Neophobia and risk assessment | 10 | 5 | 15 |
| Entire song structure | 7 | 3 | 10 |
| Song frequency and/or amplitude | 16 | 1 | 17 |
| Aggression | 11 | 11 | |
| Dawn chorus | 1 | 1 | |
| Call frequency, amplitude | 2 | 2 | |
| Song frequency, bandwidth | 2 | 2 | |
| Song syllable | 1 | 1 | |
| Song timing | 1 | 1 | |
| Genetic | |||
| Gene expression | 2 | 2 | |
| Genetic changes | 1 | 1 | 2 |
| Genetic divergence | 1 | 1 | 2 |
| Total | 148 | 139 |
| Bird Family | Common Name | Ecological | Life History | Physiological | Behavioral | Genetic | Total |
|---|---|---|---|---|---|---|---|
| Bird community (>2 families) | 58 | 7 | 15 | 29 | 2 | 111 | |
| Turdidae | Thrushes | 1 | 1 | 8 | 8 | 2 | 20 |
| Passerellidae | New world sparrows | 1 | 7 | 11 | 19 | ||
| Paridae | Tits | 3 | 11 | 2 | 16 | ||
| Passeridae | Old world sparrows | 3 | 5 | 8 | |||
| Fringillidae | Finches | 6 | 6 | ||||
| Cardinalidae | Cardinals | 4 | 4 | ||||
| Corvidae | Crows | 1 | 1 | 1 | 2 | 5 | |
| Strigidae | True owls | 3 | 1 | 2 | 6 | ||
| Zosteropidae | White-eyes | 4 | 4 | ||||
| Accipitridae | Accipiters | 2 | 4 | 1 | 4 | 11 | |
| Mimidae | Mimids | 2 | 1 | 3 | |||
| Rallidae | Rails | 3 | 3 | ||||
| Emberizidae | American sparrows | 2 | 2 | ||||
| Muscicapidae | Old world flycatchers | 2 | 2 | ||||
| Sturnidae | Starlings | 2 | 2 | ||||
| Troglodytidae | Wrens | 1 | 1 | 2 | |||
| Tyrannidae | Tyrants flycatchers | 1 | 1 | 2 | |||
| Tytonidae | Barn owls | 2 | 1 | 3 | |||
| Artamidae | Crows | 1 | 1 | ||||
| Columbidae | Pigeons and doves | 1 | 1 | ||||
| Corcoracidae | Australian mudnesters | 1 | 1 | ||||
| Falconidae | Falcons and caracaras | 2 | 3 | 1 | 6 | ||
| Icteridae | Songbirds | 1 | 1 | ||||
| Laridae | Gulls | 1 | 5 | 6 | |||
| Meliphagidae | Honeyeaters | 1 | 1 | ||||
| Monarchidae | Monarch flycatchers | 1 | 1 | ||||
| Nectariniidae | Sunbirds | 1 | 1 | ||||
| Thraupidae | Tanagers | 1 | 1 | ||||
| Pandionidae | Ospreys | 1 | 1 | ||||
| Haematopodidae | Oystercatchers | 1 | 1 | ||||
| Total | 71 | 23 | 42 | 109 | 6 | 251 |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Patankar, S.; Jambhekar, R.; Suryawanshi, K.R.; Nagendra, H. Which Traits Influence Bird Survival in the City? A Review. Land 2021, 10, 92. https://doi.org/10.3390/land10020092
Patankar S, Jambhekar R, Suryawanshi KR, Nagendra H. Which Traits Influence Bird Survival in the City? A Review. Land. 2021; 10(2):92. https://doi.org/10.3390/land10020092
Chicago/Turabian StylePatankar, Swaroop, Ravi Jambhekar, Kulbhushansingh Ramesh Suryawanshi, and Harini Nagendra. 2021. "Which Traits Influence Bird Survival in the City? A Review" Land 10, no. 2: 92. https://doi.org/10.3390/land10020092
APA StylePatankar, S., Jambhekar, R., Suryawanshi, K. R., & Nagendra, H. (2021). Which Traits Influence Bird Survival in the City? A Review. Land, 10(2), 92. https://doi.org/10.3390/land10020092