Trends in Levels of Allochthonous Dissolved Organic Carbon in Natural Water: A Review of Potential Mechanisms under a Changing Climate

{kind=link}
Abstract
:1. Introduction
2. Characterization of Natural Organic Matter in Water Systems
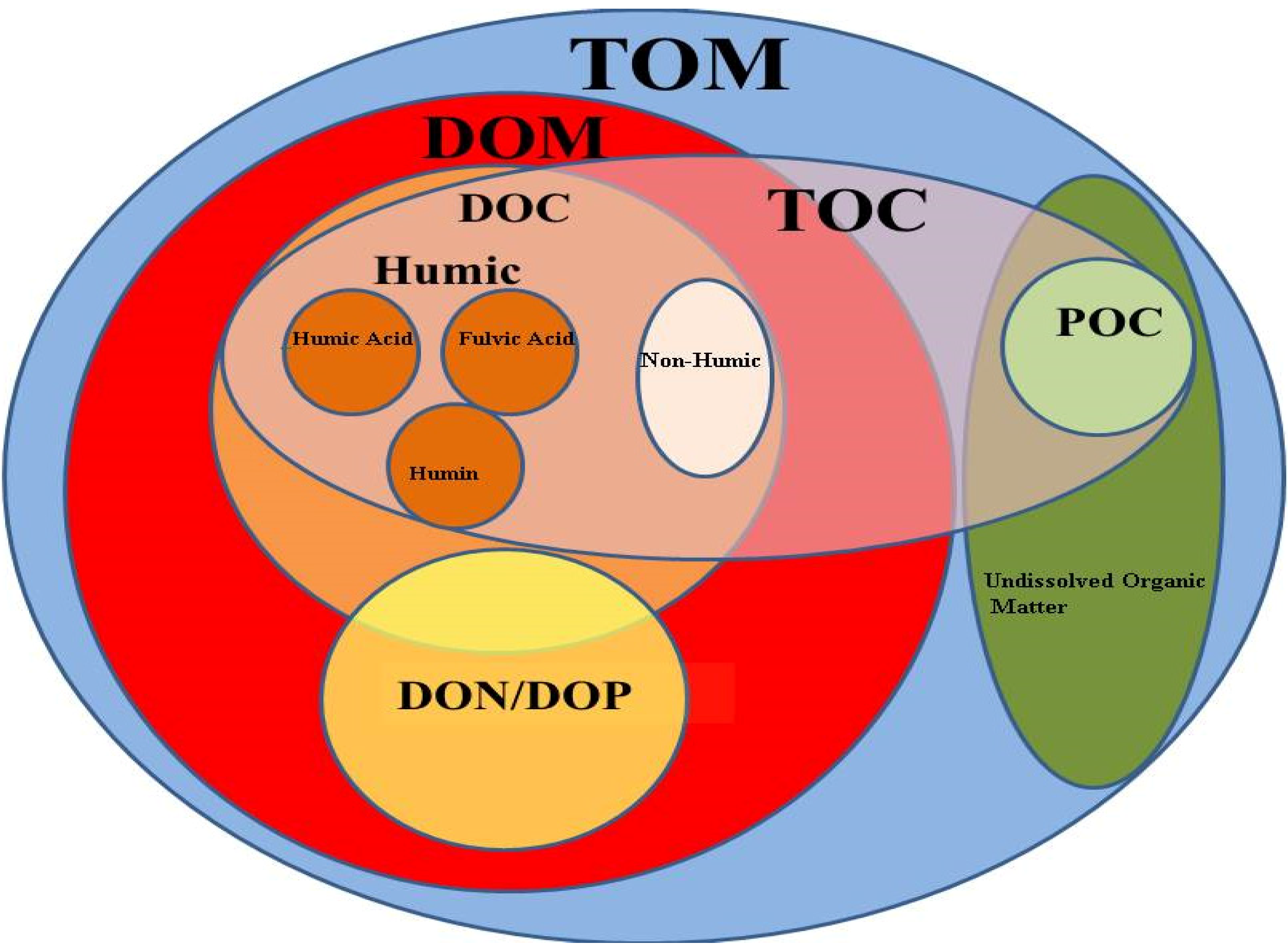
2.1. Natural Organic Matter Definition and Composition
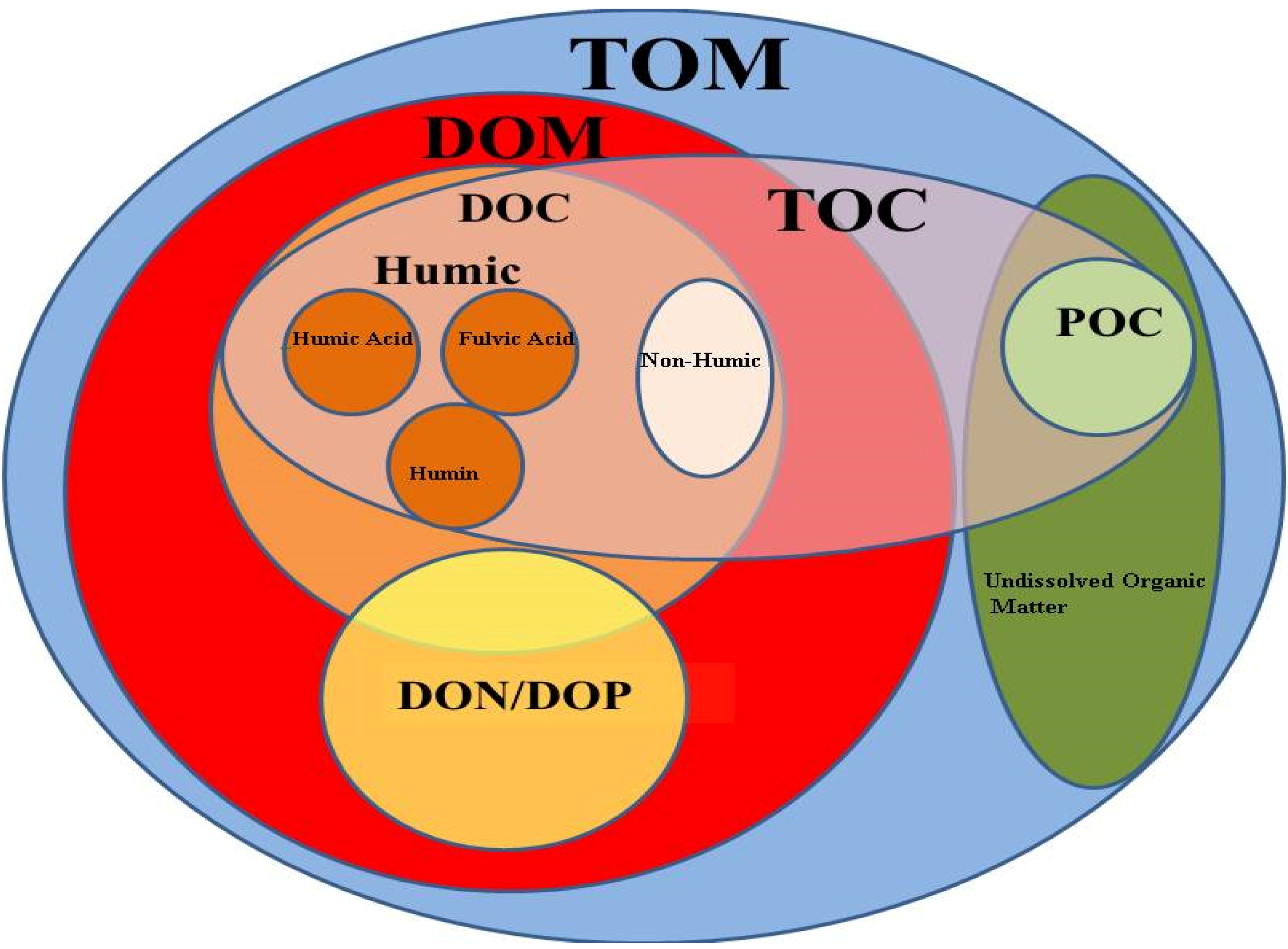
2.2. DOC Sources and Production
3. Changing DOC Concentrations in Natural Waters
4. Mechanisms for Changing DOC Levels
4.1. Atmospheric Chemical Deposition-Related Increases in DOC
4.2. Climate Change Driven Increases in DOC
4.2.1. Increasing Atmospheric CO2
4.2.2. Increasing Temperature
4.2.3. Combined Effect of Increased Atmospheric CO2 Concentration and Temperature
4.2.4. Changing Hydrology and Its Effect on DOC Concentrations
4.3. Land-Use and Its Effect on DOC Levels
5. The Impact of Increased DOC on Drinking Water Supply and Treatment
6. Conclusions
Acknowledgments
Conflicts of Interest
References
- Sinsabaugh, R.L.; Findlay, S. Dissolved organic matter: Out of the black box into the mainstream. In Aquatic Ecosystems: Interactivity of Dissolved Organic Matter; Elsevier Science: San Diego, CA, USA, 2003; pp. 479–496. [Google Scholar]
- Filella, M.; Rodriguez-Murillo, J.C. Long-Term trends of organic carbon concentrations in freshwaters: Strengths and weaknesses of existing evidence. Water 2014, 6, 1360–1418. [Google Scholar]
- Worrall, F.; Burt, T.P. Has the composition of fluvial DOC changed? Spatiotemporal patterns in the DOC-color relationship. Glob. Biogeochem. Cycl. 2010, 24. [Google Scholar] [CrossRef]
- Gallard, H.; von Gunten, U. Chlorination of natural organic matter: Kinetics of chlorination and of THM formation. Water Res. 2002, 36, 65–74. [Google Scholar] [PubMed]
- Sharp, E.L.; Parsons, S.A.; Jefferson, B. Seasonal variations in natural organic matter and its impact on coagulation in water treatment. Sci. Total Environ. 2006, 363, 183–194. [Google Scholar] [PubMed]
- Reynolds, B.; Chamberlain, P.M.; Poskitt, J.; Woods, C.; Scott, W.A.; Rowe, E.C.; Robinson, D.A.; Frogbrook, Z.L.; Keith, A.M.; Henrys, P.A.; et al. Countryside survey: National “Soil change” 1978–2007 for topsoils in great britain-acidity, carbon, and total nitrogen status. Vadose Zone J. 2013, 12. [Google Scholar] [CrossRef]
- Evans, C.D.; Monteith, D.T.; Fowler, D.; Cape, J.N.; Brayshaw, S. Hydrochloric acid: An overlooked driver of environmental change. Environ. Sci. Technol. 2011, 45, 1887–1894. [Google Scholar] [PubMed]
- Monteith, D.T.; Stoddard, J.L.; Evans, C.D.; de Wit, H.A.; Forsius, M.; Hogasen, T.; Wilander, A.; Skjelkvale, B.L.; Jeffries, D.S.; Vuorenmaa, J.; et al. Dissolved organic carbon trends resulting from changes in atmospheric deposition chemistry. Nature 2007, 450, 537–540. [Google Scholar] [PubMed]
- Evans, C.D.; Chapman, P.J.; Clark, J.M.; Monteith, D.T.; Cresser, M.S. Alternative explanations for rising dissolved organic carbon export from organic soils. Glob. Chang. Biol. 2006, 12, 2044–2053. [Google Scholar]
- Findlay, S.E.G. Increased carbon transport in the hudson river: Unexpected consequence of nitrogen deposition? Front. Ecol. Environ. 2005, 3, 133–137. [Google Scholar] [CrossRef]
- Freeman, C.; Evans, C.D.; Monteith, D.T.; Reynolds, B.; Fenner, N. Export of organic carbon from peat soils. Nature 2001, 412, 785–785. [Google Scholar] [PubMed]
- Freeman, C.; Fenner, N.; Ostle, N.J.; Kang, H.; Dowrick, D.J.; Reynolds, B.; Lock, M.A.; Sleep, D.; Hughes, S.; Hudson, J. Export of dissolved organic carbon from peatlands under elevated carbon dioxide levels. Nature 2004, 430, 195–198. [Google Scholar] [PubMed]
- Wilson, H.F.; Saiers, J.E.; Raymond, P.A.; Sobczak, W.V. Hydrologic drivers and seasonality of dissolved organic carbon concentration, nitrogen content, bioavailability, and export in a forested new england stream. Ecosystems 2013, 16, 604–616. [Google Scholar]
- Hongve, D.; Riise, G.; Kristiansen, J.F. Increased colour and organic acid concentrations in norwegian forest lakes and drinking water—A result of increased precipitation? Aquat. Sci. 2004, 66, 231–238. [Google Scholar]
- Anderson, N.; Dietz, R.; Engstrom, D. Land-Use change, not climate, controls organic carbon burial in lakes. Proc. R. Soc. B Biol. Sci. 2013, 280. [Google Scholar] [CrossRef]
- Cool, G.; Lebel, A.; Sadiq, R.; Rodriguez, M.J. Impact of catchment geophysical characteristics and climate on the regional variability of dissolved organic carbon (DOC) in surface water. Sci. Total Environ. 2014, 490, 947–956. [Google Scholar] [PubMed]
- McGuire, A.D.; Anderson, L.G.; Christensen, T.R.; Dallimore, S.; Guo, L.D.; Hayes, D.J.; Heimann, M.; Lorenson, T.D.; Macdonald, R.W.; Roulet, N. Sensitivity of the carbon cycle in the arctic to climate change. Ecol. Monogr. 2009, 79, 523–555. [Google Scholar]
- Detwiler, R.P.; Hall, C.A.S. Tropical forests and the global carbon-cycle. Science 1988, 239, 42–47. [Google Scholar] [PubMed]
- Brown, S.; Lugo, A.E. The storage and production of organic-matter in tropical forests and their role in the global carbon-cycle. Biotropica 1982, 14, 161–187. [Google Scholar]
- Spencer, R.G.M.; Hernes, P.J.; Ruf, R.; Baker, A.; Dyda, R.Y.; Stubbins, A.; Six, J. Temporal controls on dissolved organic matter and lignin biogeochemistry in a pristine tropical river, democratic republic of Congo. J. Geophys. Res. Biogeosci. 2010, 115. [Google Scholar] [CrossRef]
- Cramer, W.; Bondeau, A.; Schaphoff, S.; Lucht, W.; Smith, B.; Sitch, S. Tropical forests and the global carbon cycle: Impacts of atmospheric carbon dioxide, climate change and rate of deforestation. Philos. Trans. R. Soc. Lond. Ser. B Biol. Sci. 2004, 359, 331–343. [Google Scholar]
- Monteith, D.; Evans, C.; Henrys, P.; Simpson, G.; Malcolm, I. Trends in the hydrochemistry of acid-sensitive surface waters in the UK 1988–2008. Ecol. Indic. 2014, 37, 287–303. [Google Scholar]
- Evans, C.D.; Monteith, D.T.; Reynolds, B.; Clark, J.M. Buffering of recovery from acidification by organic acids. Sci. Total Environ. 2008, 404, 316–325. [Google Scholar] [PubMed]
- Battin, T.J.; Luyssaert, S.; Kaplan, L.A.; Aufdenkampe, A.K.; Richter, A.; Tranvik, L.J. The boundless carbon cycle. Nat. Geosci. 2009, 2, 598–600. [Google Scholar]
- Filella, M. Freshwaters: Which NOM matters? Environ. Chem. Lett. 2009, 7, 21–35. [Google Scholar] [CrossRef]
- Thurman, E.M. Organic Geochemistry of Natural Waters; Kluwer Academic: Dordrecht, the Netherlands, 1985. [Google Scholar]
- McDonald, S.; Bishop, A.G.; Prenzler, P.D.; Robards, K. Analytical chemistry of freshwater humic substances. Anal. Chim. Acta 2004, 527, 105–124. [Google Scholar]
- Leenheer, J.A.; Croue, J.P. Characterizing aquatic dissolved organic matter. Environ. Sci. Technol. 2003, 37, 18A–26A. [Google Scholar] [PubMed]
- Malcolm, R.L. The uniqueness of humic substances in each of soil, stream and marine environments. Anal. Chim. Acta 1990, 232, 19–30. [Google Scholar]
- Drever, J.I. The Geochemistry of Natural Waters: Surface and Groundwater Environments; Prentice Hall: Upper Saddle River, NJ, USA, 1997. [Google Scholar]
- Evans, C.D.; Monteith, D.T.; Cooper, D.M. Long-Term increases in surface water dissolved organic carbon: Observations, possible causes and environmental impacts. Environ. Pollut. 2005, 137, 55–71. [Google Scholar] [PubMed]
- Stevenson, I.L.; Schnitzer, M. Transmission electron-microscopy of extracted fulvic and humic acids. Soil Sci. 1982, 133, 179–185. [Google Scholar]
- Cory, R.M.; McKnight, D.M. Fluorescence spectroscopy reveals ubiquitous presence of oxidized and reduced quinones in dissolved organic matter. Environ. Sci. Technol. 2005, 39, 8142–8149. [Google Scholar] [PubMed]
- Parlanti, E.; Morin, B.; Vacher, L. Combined 3D-spectrofluorometry, high performance liquid chromatography and capillary electrophoresis for the characterization of dissolved organic matter in natural waters. Org. Geochem. 2002, 33, 221–236. [Google Scholar]
- Abdulla, H.A.N.; Sleighter, R.L.; Hatcher, P.G. Two dimensional correlation analysis of fourier transform ion cyclotron resonance mass spectra of dissolved organic matter: A new graphical analysis of trends. Anal. Chem. 2013, 85, 3895–3902. [Google Scholar] [PubMed]
- Koch, B.P.; Witt, M.R.; Engbrodt, R.; Dittmar, T.; Kattner, G. Molecular formulae of marine and terrigenous dissolved organic matter detected by electrospray ionization fourier transform ion cyclotron resonance mass spectrometry. Geochim. Cosmochim. Acta 2005, 69, 3299–3308. [Google Scholar]
- Gonsior, M.; Peake, B.M.; Cooper, W.T.; Podgorski, D.; D’Andrilli, J.; Cooper, W.J. Photochemically induced changes in dissolved organic matter identified by ultrahigh resolution fourier transform ion cyclotron resonance mass spectrometry. Environ. Sci. Technol. 2009, 43, 698–703. [Google Scholar] [PubMed]
- Brown, T.L.; Rice, J.A. Effect of experimental parameters on the ESI FT-ICR mass spectrum of fulvic acid. Anal. Chem. 2000, 72, 384–390. [Google Scholar] [PubMed]
- Kujawinski, E.B.; Hatcher, P.G.; Freitas, M.A. High-Resolution fourier transform ion cyclotron resonance mass spectrometry of humic and fulvic acids: Improvements and comparisons. Anal. Chem. 2002, 74, 413–419. [Google Scholar] [PubMed]
- Benner, R.; Kaiser, K. Biological and photochemical transformations of amino acids and lignin phenols in riverine dissolved organic matter. Biogeochemistry 2011, 102, 209–222. [Google Scholar]
- Mutabaruka, R.; Hairiah, K.; Cadisch, G. Microbial degradation of hydrolysable and condensed tannin polyphenol-protein complexes in soils from different land-use histories. Soil Biol. Biochem. 2007, 39, 1479–1492. [Google Scholar]
- Fenner, N.; Freeman, C.; Reynolds, B. Hydrological effects on the diversity of phenolic degrading bacteria in a peatland: Implications for carbon cycling. Soil Biol. Biochem. 2005, 37, 1277–1287. [Google Scholar]
- Orth, A.B.; Denny, M.; Tien, M. Overproduction of lignin-degrading enzymes by and isolate of phanerochaete-chrysosporium. Appl. Environ. Microbiol. 1991, 57, 2591–2596. [Google Scholar] [PubMed]
- McKnight, D.M.; Andrews, E.D.; Spaulding, S.A.; Aiken, G.R. Aquatic fulvic-acids in algal-rich antarctic ponds. Limnol. Oceanogr. 1994, 39, 1972–1979. [Google Scholar]
- McKnight, D.M.; Boyer, E.W.; Westerhoff, P.K.; Doran, P.T.; Kulbe, T.; Andersen, D.T. Spectrofluorometric characterization of dissolved organic matter for indication of precursor organic material and aromaticity. Limnol. Oceanogr. 2001, 46, 38–48. [Google Scholar]
- Jaffe, R.; McKnight, D.; Maie, N.; Cory, R.; McDowell, W.H.; Campbell, J.L. Spatial and temporal variations in DOM composition in ecosystems: The importance of long-term monitoring of optical properties. J. Geophys. Res. Biogeosci. 2008, 113. [Google Scholar] [CrossRef]
- SanClements, M.D.; Oelsner, G.P.; McKnight, D.M.; Stoddard, J.L.; Nelson, S.J. New insights into the source of decadal increases of dissolved organic matter in acid-sensitive lakes of the northeastern United States. Environ. Sci. Technol. 2012, 46, 3212–3219. [Google Scholar] [PubMed]
- Coble, P.G.; Green, S.A.; Blough, N.V.; Gagosian, R.B. Characterization of dissolved organic-matter in the black-sea by fluorescence spectroscopy. Nature 1990, 348, 432–435. [Google Scholar]
- Coble, P.G. Characterization of marine and terrestrial DOM in seawater using excitation emission matrix spectroscopy. Mar. Chem. 1996, 51, 325–346. [Google Scholar]
- Gone, D.L.; Seidel, J.; Batiot, C.; Bamory, K.; Ligban, R.; Biemi, J. Using fluoresence spectroscopy eem to evaluate the efficiency of organic matter removal during coagulation-flocculation of a tropical surface water (agbo reservoir). J. Hazard. Mater. 2009, 172, 693–699. [Google Scholar] [PubMed]
- Santin, C.; Yamashita, Y.; Otero, X.L.; Alvarez, M.A.; Jaffe, R. Characterizing humic substances from estuarine soils and sediments by excitation-emission matrix spectroscopy and parallel factor analysis. Biogeochemistry 2009, 96, 131–147. [Google Scholar]
- Walker, S.A.; Amon, R.M.W.; Stedmon, C.; Duan, S.; Louchouarn, P. The use of PARAFAC modeling to trace terrestrial dissolved organic matter and fingerprint water masses in coastal Canadian arctic surface waters. J. Geophys. Res. Biogeosci. 2009, 114. [Google Scholar] [CrossRef]
- Pagano, T.; Ross, A.D.; Chiarelli, J.; Kenny, J.E. Multidimensional fluorescence studies of the phenolic content of dissolved organic carbon in humic substances. J. Environ. Monit. 2012, 14, 937–943. [Google Scholar] [PubMed]
- Sobek, S.; Tranvik, L.J.; Prairie, Y.T.; Kortelainen, P.; Cole, J.J. Patterns and regulation of dissolved organic carbon: An analysis of 7,500 widely distributed lakes. Limnol. Oceanogr. 2007, 52, 1208–1219. [Google Scholar]
- Williams, C.J.; Yamashita, Y.; Wilson, H.F.; Jaffe, R.; Xenopoulos, M.A. Unraveling the role of land use and microbial activity in shaping dissolved organic matter characteristics in stream ecosystems. Limnol. Oceanogr. 2010, 55, 1159–1171. [Google Scholar]
- Algesten, G.; Sobek, S.; Bergstrom, A.K.; Jonsson, A.; Tranvik, L.J.; Jansson, M. Contribution of sediment respiration to summer CO2 emission from low productive boreal and subarctic lakes. Microb. Ecol. 2005, 50, 529–535. [Google Scholar] [PubMed]
- Mattsson, T.; Kortelain, P.; Laubel, A.; Evans, D.; Pujo-Pay, M.; Raike, A.; Conan, P. Export of dissolved organic matter in relation to land use along a european climatic gradient. Sci. Total Environ. 2009, 407, 1967–1976. [Google Scholar] [PubMed]
- Stanley, E.H.; Powers, S.M.; Lottig, N.R.; Buffam, I.; Crawford, J.T. Contemporary changes in dissolved organic carbon (DOC) in human-dominated rivers: Is there a role for DOC management? Freshw. Biol. 2012, 57, 26–42. [Google Scholar]
- Jonsson, A.; Meili, M.; Bergstrom, A.K.; Jansson, M. Whole-Lake mineralization of allochthonous and autochthonous organic carbon in a large humic lake (ortrasket, N. sweden). Limnol. Oceanogr. 2001, 46, 1691–1700. [Google Scholar]
- Royer, T.V.; David, M.B. Export of dissolved organic carbon from agricultural streams in Illinois, USA. Aquat. Sci. 2005, 67, 465–471. [Google Scholar]
- Bida, M.R. Quantity and Composition of Stream Dissolved Organic Matter in the Watershed of Conesus Lake, New York. Master’s Thesis, Rochester Institute of Technology, Rochester, NY, USA,, 15 July 2013. [Google Scholar]
- Tipping, E.; Woof, C. Humic substances in acid organic soils—Modeling their release to the soil solution in terms of humic charge. J. Soil Sci. 1990, 41, 573–586. [Google Scholar]
- Pind, A.; Freeman, C.; Lock, M.A. Enzymatic degradation of phenolic materials in peatlands—Measurement of phenol oxidase activity. Plant Soil 1994, 159, 227–231. [Google Scholar]
- Kalbitz, K.; Solinger, S.; Park, J.H.; Michalzik, B.; Matzner, E. Controls on the dynamics of dissolved organic matter in soils: A review. Soil Sci. 2000, 165, 277–304. [Google Scholar]
- Wershaw, R.L. Evaluation of Conceptual Models of Natural Organic Matter (humus) from a Consideration of the Chemical and Biochemical Processes of Humification; U.S. Geological Survey: Reston, VA, USA, 2004. [Google Scholar]
- Findlay, S.; Sinsabaugh, R.L. Aquatic Ecosystems: Interactivity of Dissolved Organic Matter; Academic Press: San Diego, CA, USA, 2003. [Google Scholar]
- Aitkenhead-Peterson, J.A.; McDowell, W.H.; Neff, J.C. Sources, production, and regulation of allochthonous dissolved organic matter inputs to surface waters. In Aquatic Ecosystems: Interactivity of Dissolved Organic Matter; Findlay, S.E.G., Sinsabaugh, R.L., Eds.; Elsevier Science: San Diego, CA, USA, 2003; pp. 25–70. [Google Scholar]
- Gorham, E. Northern peatlands—Role in the carbon-cycle and probable responses to climatic warming. Ecol. Appl. 1991, 1, 182–195. [Google Scholar]
- Fenner, N.; Ostle, N.J.; McNamara, N.; Sparks, T.; Harmens, H.; Reynolds, B.; Freeman, C. Elevated CO2 effects on peatland plant community carbon dynamics and DOC production. Ecosystems 2007, 10, 635–647. [Google Scholar]
- Holmes, R.M.; McClelland, J.W.; Peterson, B.J.; Tank, S.E.; Bulygina, E.; Eglinton, T.I.; Gordeev, V.V.; Gurtovaya, T.Y.; Raymond, P.A.; Repeta, D.J.; et al. Seasonal and annual fluxes of nutrients and organic matter from large rivers to the arctic ocean and surrounding seas. Estuaries Coasts 2012, 35, 369–382. [Google Scholar]
- Kang, H.; Freeman, C.; Jang, I. Global increases in dissolved organic carbon in rivers and their implications. Korean J. Limnol. 2010, 43, 453–458. [Google Scholar]
- Engelhaupt, E.; Bianchi, T.S. Sources and composition of high-molecular-weight dissolved organic carbon in a Southern Louisiana tidal stream (bayou trepagnier). Limnol. Oceanogr. 2001, 46, 917–926. [Google Scholar]
- Skjelkvale, B.L.; Mannio, J.; Wilander, A.; Andersen, T. Recovery from acidification of lakes in Finland, Norway and Sweden 1990–1999. Hydrol. Earth Syst. Sci. 2001, 5, 327–337. [Google Scholar]
- Dillon, P.J.; Skjelkvale, B.L.; Somers, K.M.; Torseth, K. Coherent responses of sulphate concentration in norwegian lakes: Relationships with sulphur deposition and climate indices. Hydrol. Earth Syst. Sci. 2003, 7, 596–608. [Google Scholar]
- Driscoll, C.T.; Driscoll, K.M.; Roy, K.M.; Mitchell, M.J. Chemical response of lakes in the adirondack region of new york to declines in acidic deposition. Environ. Sci. Technol. 2003, 37, 2036–2042. [Google Scholar] [PubMed]
- Hudson, J.J.; Dillon, P.J.; Somers, K.M. Long-Term patterns in dissolved organic carbon in boreal lakes: The role of incident radiation, precipitation, air temperature, southern oscillation and acid deposition. Hydrol. Earth Syst. Sci. 2003, 7, 390–398. [Google Scholar]
- Pastor, J.; Solin, J.; Bridgham, S.D.; Updegraff, K.; Harth, C.; Weishampel, P.; Dewey, B. Global warming and the export of dissolved organic carbon from boreal peatlands. Oikos 2003, 100, 380–386. [Google Scholar]
- Xenopoulos, M.A.; Lodge, D.M.; Frentress, J.; Kreps, T.A.; Bridgham, S.D.; Grossman, E.; Jackson, C.J. Regional comparisons of watershed determinants of dissolved organic carbon in temperate lakes from the upper great lakes region and selected regions globally. Limnol. Oceanogr. 2003, 48, 2321–2334. [Google Scholar]
- Worrall, F.; Burt, T.; Adamson, J. Can climate change explain increases in DOC flux from upland peat catchments? Sci. Total Environ. 2004, 326, 95–112. [Google Scholar] [CrossRef]
- Worrall, F.; Harriman, R.; Evans, C.D.; Watts, C.D.; Adamson, J.; Neal, C.; Tipping, E.; Burt, T.; Grieve, I.; Monteith, D. Trends in dissolved organic carbon in UK rivers and lakes. Biogeochemistry 2004, 70, 369–402. [Google Scholar]
- Fahey, T.J.; Siccama, T.G.; Driscoll, C.T.; Likens, G.E.; Campbell, J.; Johnson, C.E.; Battles, J.J.; Aber, J.D.; Cole, J.J.; Fisk, M.C.; et al. The biogeochemistry of carbon at hubbard brook. Biogeochemistry 2005, 75, 109–176. [Google Scholar]
- Harrison, J.A.; Caraco, N.; Seitzinger, S.P. Global patterns and sources of dissolved organic matter export to the coastal zone: Results from a spatially explicit, global model. Glob. Biogeochem. Cycl. 2005, 19. [Google Scholar] [CrossRef]
- Skjelkvale, B.L.; Borg, H.; Hindar, A.; Wilander, A. Large scale patterns of chemical recovery in lakes in Norway and Sweden: Importance of seasalt episodes and changes in dissolved organic carbon. In Proceedings of the 7th International Conference on Acid Deposition, Prague, Czech Republic, 12–17 June 2005; pp. 1174–1180.
- Skjelkvale, B.L.; Stoddard, J.L.; Jeffries, D.S.; Torseth, K.; Hogasen, T.; Bowman, J.; Mannio, J.; Monteith, D.T.; Mosello, R.; Rogora, M.; et al. Regional scale evidence for improvements in surface water chemistry 1990–2001. Environ. Pollut. 2005, 137, 165–176. [Google Scholar] [PubMed]
- Burns, D.A.; McHale, M.R.; Driscoll, C.T.; Roy, K.M. Response of surface water chemistry to reduced levels of acid precipitation: Comparison of trends in two regions of New York, USA. Hydrol. Process. 2006, 20, 1611–1627. [Google Scholar]
- Gueguen, C.; Guo, L.D.; Wang, D.; Tanaka, N.; Hung, C.C. Chemical characteristics and origin of dissolved organic matter in the Yukon river. Biogeochemistry 2006, 77, 139–155. [Google Scholar]
- Roulet, N.; Moore, T.R. Environmental chemistry—Browning the waters. Nature 2006, 444, 283–284. [Google Scholar] [PubMed]
- Vuorenmaa, J.; Forsius, M.; Mannio, J. Increasing trends of total organic carbon concentrations in small forest lakes in finland from 1987 to 2003. Sci. Total Environ. 2006, 365, 47–65. [Google Scholar] [PubMed]
- Fenner, N.; Freeman, C.; Lock, M.A.; Harmens, H.; Reynolds, B.; Sparks, T. Interactions between elevated CO2 and warming could amplify DOC exports from peatland catchments. Environ. Sci. Technol. 2007, 41, 3146–3152. [Google Scholar] [PubMed]
- Worrall, F.; Burt, T.P. Flux of dissolved organic carbon from UK rivers. Glob. Biogeochem. Cycl. 2007, 21. [Google Scholar] [CrossRef]
- Worrall, F.; Burt, T.P. Trends in DOC concentration in great britain. J. Hydrol. 2007, 346, 81–92. [Google Scholar]
- Worrall, F.; Burt, T.P. The effect of severe drought on the dissolved organic carbon (DOC) concentration and flux from British rivers. J. Hydrol. 2008, 361, 262–274. [Google Scholar]
- Worrall, F.; Guilbert, T.; Besien, T. The flux of carbon from rivers: The case for flux from england and wales. Biogeochemistry 2007, 86, 63–75. [Google Scholar]
- De Wit, H.A.; Mulder, J.; Hindar, A.; Hole, L. Long-Term increase in dissolved organic carbon in streamwaters in Norway is response to reduced acid deposition. Environ. Sci. Technol. 2007, 41, 7706–7713. [Google Scholar] [PubMed]
- Couture, S.; Houle, D.; Gagnon, C. Increases of dissolved organic carbon in temperate and boreal lakes in Quebec, Canada. Environ. Sci. Pollut. Res. 2012, 19, 361–371. [Google Scholar]
- Lawrence, G.B.; Dukett, J.E.; Houck, N.; Snyder, P.; Capone, S. Increases in dissolved organic carbon accelerate loss of toxic al in Adirondack lakes recovering from acidification. Environ. Sci. Technol. 2013, 47, 7095–7100. [Google Scholar] [PubMed]
- Schindler, D.W.; Curtis, P.J.; Bayley, S.E.; Parker, B.R.; Beaty, K.G.; Stainton, M.P. Climate-Induced changes in the dissolved organic carbon budgets of boreal lakes. Biogeochemistry 1997, 36, 9–28. [Google Scholar]
- Hejzlar, J.; Dubrovsky, M.; Buchtele, J.; Ruzicka, M. The apparent and potential effects of climate change on the inferred concentration of dissolved organic matter in a temperate stream (the Malse River, South Bohemia). In Proceedings of the Detecting Environmental Change—Science and Society Conference, London, England, 16–20 July 2001; pp. 143–152.
- Oulehle, F.; Chuman, T.; Majer, V.; Hruska, J. Chemical recovery of acidified bohemian lakes between 1984 and 2012: The role of acid deposition and bark beetle induced forest disturbance. Biogeochemistry 2013, 116, 83–101. [Google Scholar]
- Frey, K.E.; Smith, L.C. Amplified carbon release from vast west Siberian Peatlands by 2100. Geophys. Res. Lett. 2005, 32. [Google Scholar] [CrossRef]
- Prokushkin, A.S.; Kajimoto, T.; Prokushkin, S.G.; McDowell, W.H.; Abaimov, A.P.; Matsuura, Y. Climatic Factors influencing fluxes of dissolved organic carbon from the forest floor in a continuous-permafrost Siberian Watershed. In Proceedings of the 12th Annual Conference of the International-Boreal-Forest-Research-Association, Fairbanks, AK, USA, 3–6 May 2004; pp. 2130–2140.
- Gordeev, V.V.; Kravchishina, M.D. River flux of dissolved organic carbon (DOC) and particulate organic carbon (POC) to the Arctic Ocean: What are the consequences of the global changes? In Influence of Climate Change on the Changing Arctic and Sub-Arctic Conditions; Nihoul, J.C.J., Kostianoy, A.G., Eds.; Springer: Dordrecht, the Netherlands, 2009; pp. 145–160. [Google Scholar]
- Porcal, P.; Koprivnjak, J.F.; Molot, L.A.; Dillon, P.J. Humic substances-part 7: The biogeochemistry of dissolved organic carbon and its interactions with climate change. Environ. Sci. Pollut. Res. 2009, 16, 714–726. [Google Scholar]
- Tranvik, L.J.; Downing, J.A.; Cotner, J.B.; Loiselle, S.A.; Striegl, R.G.; Ballatore, T.J.; Dillon, P.; Finlay, K.; Fortino, K.; Knoll, L.B.; et al. Lakes and reservoirs as regulators of carbon cycling and climate. Limnol. Oceanogr. 2009, 54, 2298–2314. [Google Scholar]
- Ritson, J.P.; Graham, N.J.D.; Templeton, M.R.; Clark, J.M.; Gough, R.; Freeman, C. The impact of climate change on the treatability of dissolved organic matter (DOM) in upland water supplies: A UK perspective. Sci. Total Environ. 2014, 473, 714–730. [Google Scholar] [PubMed]
- Stoddard, J.L.; Jeffries, D.S.; Lukewille, A.; Clair, T.A.; Dillon, P.J.; Driscoll, C.T.; Forsius, M.; Johannessen, M.; Kahl, J.S.; Kellogg, J.H.; et al. Regional trends in aquatic recovery from acidification in North America and Europe. Nature 1999, 401, 575–578. [Google Scholar]
- Ulrich, K.U.; Paul, L.; Meybohm, A. Response of drinking-water reservoir ecosystems to decreased acidic atmospheric deposition in SE Germany: Trends of chemical reversal. Environ. Pollut. 2006, 141, 42–53. [Google Scholar] [PubMed]
- Rowe, E.C.; Tipping, E.; Posch, M.; Oulehle, F.; Cooper, D.M.; Jones, T.G.; Burden, A.; Hall, J.; Evans, C.D. Predicting nitrogen and acidity effects on long-term dynamics of dissolved organic matter. Environ. Pollut. 2014, 184, 271–282. [Google Scholar] [PubMed]
- Fowler, D.; Smith, R.I.; Muller, J.B.A.; Hayman, G.; Vincent, K.J. Changes in the atmospheric deposition of acidifying compounds in the UK between 1986 and 2001. Environ. Pollut. 2005, 137, 15–25. [Google Scholar] [PubMed]
- Holland, E.A.; Braswell, B.H.; Sulzman, J.; Lamarque, J.F. Nitrogen deposition onto the United States and Western Europe: Synthesis of observations and models. Ecol. Appl. 2005, 15, 38–57. [Google Scholar]
- Pregitzer, K.S.; Zak, D.R.; Burton, A.J.; Ashby, J.A.; MacDonald, N.W. Chronic nitrate additions dramatically increase the export of carbon and nitrogen from northern hardwood ecosystems. Biogeochemistry 2004, 68, 179–197. [Google Scholar]
- Yano, Y.; McDowell, W.H.; Aber, J.D. Biodegradable dissolved organic carbon in forest soil solution and effects of chronic nitrogen deposition. Soil Biol. Biochem. 2000, 32, 1743–1751. [Google Scholar]
- Currie, W.S.; Aber, J.D.; McDowell, W.H.; Boone, R.D.; Magill, A.H. Vertical transport of dissolved organic C and N under long-term N amendments in pine and hardwood forests. Biogeochemistry 1996, 35, 471–505. [Google Scholar]
- Bragazza, L.; Freeman, C.; Jones, T.; Rydin, H.; Limpens, J.; Fenner, N.; Ellis, T.; Gerdol, R.; Hajek, M.; Hajek, T.; et al. Atmospheric nitrogen deposition promotes carbon loss from peat bogs. Proc. Natl. Acad. Sci. USA 2006, 103, 19386–19389. [Google Scholar] [PubMed]
- Evans, C.D.; Reynolds, B.; Hinton, C.; Hughes, S.; Norris, D.; Grant, S.; Williams, B. Effects of decreasing acid deposition and climate change on acid extremes in an upland stream. Hydrol. Earth Syst. Sci. 2008, 12, 337–351. [Google Scholar]
- Krug, E.C.; Frink, C.R. Acid-Rain on acid soil—A new perspective. Science 1983, 221, 520–525. [Google Scholar] [PubMed]
- Evans, A.; Zelazny, L.W.; Zipper, C.E. Division s-7—Forest and range soils—Solution parameters influencing dissolved organic-carbon levels in 3 forest soils. Soil Sci. Soc. Am. J. 1988, 52, 1789–1792. [Google Scholar]
- Evans, C.D.; Jones, T.G.; Burden, A.; Ostle, N.; Zielinski, P.; Cooper, M.D.A.; Peacock, M.; Clark, J.M.; Oulehle, F.; Cooper, D.; et al. Acidity controls on dissolved organic carbon mobility in organic soils. Glob. Chang. Biol. 2012, 18, 3317–3331. [Google Scholar]
- Worrall, F.; Gibson, H.S.; Burt, T.P. Production vs. Solubility in controlling runoff of DOC from peat soils—The use of an event analysis. J. Hydrol. 2008, 358, 84–95. [Google Scholar]
- Solomon, S. Climate Change 2007: The Physical Science Basis: Part of the Working Group I Contribution to the Fourth Assessment Report of the Intergovernmental Panel on Climate Change; Cambridge University Press: Cambridge, MA, USA, 2007. [Google Scholar]
- Stocker, T.F.; Qin, D.; Plattner, G.-K.; Tignor, M.; Allen, S.K.; Boschung, J.; Nauels, A.; Xia, Y.; Bex, V.; Midgley, P.M. Climate change 2013: The physical science basis. In Intergovernmental Panel on Climate Change, Working Group I Contribution to the IPCC Fifth Assessment Report (AR5); Cambridge University Press: New York, NY, USA, 2013. [Google Scholar]
- Houghton, J.T. Climate Change 2001: The Scientific Basis: Contribution of Working Group I to the Third Assessment Report of the Intergovernmental Panel on Climate Change; Cambridge University Press: Cambridge, MA, USA, 2001. [Google Scholar]
- Harrison, A.F.; Taylor, K.; Scott, A.; Poskitt, J.; Benham, D.; Grace, J.; Chaplow, J.; Rowland, P. Potential effects of climate change on DOC release from three different soil types on the northern pennines UK: Examination using field manipulation experiments. Glob. Chang. Biol. 2008, 14, 687–702. [Google Scholar]
- Pace, M.L.; Cole, J.J. Synchronous variation of dissolved organic carbon and color in lakes. Limnol. Oceanogr. 2002, 47, 333–342. [Google Scholar]
- Tipping, E.; Smith, E.J.; Bryant, C.L.; Adamson, J.K. The organic carbon dynamics of a moorland catchment in NW England. Biogeochemistry 2007, 84, 171–189. [Google Scholar]
- Kominoski, J.S.; Moore, P.A.; Wetzel, R.G.; Tuchman, N.C. Elevated CO2 alters leaf-litter-derived dissolved organic carbon: Effects on stream periphyton and crayfish feeding preference. J. North Am. Benthol. Soc. 2007, 26, 663–672. [Google Scholar]
- Norby, R.J.; DeLucia, E.H.; Gielen, B.; Calfapietra, C.; Giardina, C.P.; King, J.S.; Ledford, J.; McCarthy, H.R.; Moore, D.J.P.; Ceulemans, R.; et al. Forest response to elevated CO2 is conserved across a broad range of productivity. Proc. Natl. Acad. Sci. USA 2005, 102, 18052–18056. [Google Scholar] [PubMed]
- Freeman, C.; Baxter, R.; Farrar, J.F.; Jones, S.E.; Plum, S.; Ashendon, T.W.; Stirling, C. Could competition between plants and microbes regulate plant nutrition and atmospheric CO2 concentrations? Sci. Total Environ. 1998, 220, 181–184. [Google Scholar] [CrossRef]
- Kane, E.S.; Mazzoleni, L.R.; Kratz, C.J.; Hribljan, J.A.; Johnson, C.P.; Pypker, T.G.; Chimner, R. Peat porewater dissolved organic carbon concentration and lability increase with warming: A field temperature manipulation experiment in a poor-fen. Biogeochemistry 2014, 119, 161–178. [Google Scholar]
- Clark, J.M.; Chapman, P.J.; Adamson, J.K.; Lane, S.N. Influence of drought-induced acidification on the mobility of dissolved organic carbon in peat soils. Glob. Chang. Biol. 2005, 11, 791–809. [Google Scholar]
- Worrall, F.; Burt, T.P.; Adamson, J.K. Linking pulses of atmospheric deposition to DOC release in an upland peat-covered catchment. Glob. Biogeochem. Cycl. 2008, 22, 15. [Google Scholar]
- Wittwer, S.; Robb, W. Carbon dioxide enrichment of greenhouse atmospheres for food crop production. Econ. Bot. 1964, 18, 34–56. [Google Scholar]
- Norby, R.J.; Zak, D.R. Ecological lessons from free-air CO2 enrichment (FACE) experiments. Annu. Rev. Ecol. Evol. Syst. 2011, 42, 181–203. [Google Scholar]
- Clair, T.A.; Arp, P.; Moore, T.R.; Dalva, M.; Meng, F.R. Gaseous carbon dioxide and methane, as well as dissolved organic carbon losses from a small temperate wetland under a changing climate. In Proceedings of the Advances in Terrestrial Ecosystem: Carbon Inventory Measurements and Monitoring Conference, Raleigh, NC, USA, 3–5 October 2000; pp. S143–S148.
- Mitsch, W.J.; Gosselink, J.G. Wetlands; Van Nostrand Reinhold: New York, NY, USA, 1993. [Google Scholar]
- Woodin, S.; Graham, B.; Killick, A.; Skiba, U.; Cresser, M. Nutrient limitation of the long-term response of heather [Calluna-vulgaris (L) Hull] to CO2 enrichment. New Phytol. 1992, 122, 635–642. [Google Scholar]
- Dawes, M.A.; Hagedorn, F.; Handa, I.T.; Streit, K.; Ekblad, A.; Rixen, C.; Korner, C.; Hattenschwiler, S. An alpine treeline in a carbon dioxide-rich world: Synthesis of a nine-year free-air carbon dioxide enrichment study. Oecologia 2013, 171, 623–637. [Google Scholar] [PubMed]
- Jiang, H.B.; Wang, Y.; Kanost, M.R. Pro-Phenol oxidase activating proteinase from an insect, manduca sexta: A bacteria-inducible protein similar to drosophila Easter. Proc. Natl. Acad. Sci. USA 1998, 95, 12220–12225. [Google Scholar] [PubMed]
- Wetzel, R.G. Gradient-Dominated osystems—Sources and regulatory functions of dissolved organic-matter in fresh-water ecosystems. In Proceedings of the Symposium on Dissolved Organic Matter in Lacustrine Ecosystems: Energy Source and System Regulator, Helsinki, Finland, 6–10 August 1990; pp. 181–198.
- Freeman, C.; Lock, M.A.; Marxsen, J.; Jones, S.E. Inhibitory effects of high-molecular-weight dissolved organic-matter upon metabolic processes in biofilms from contrasting rivers and streams. Freshw. Biol. 1990, 24, 159–166. [Google Scholar]
- Sinsabaugh, R.L.; Antibus, R.K.; Linkins, A.E. An enzymatic approach to the analysis of microbial activity during plant litter decomposition. In Proceedings of the Workshop on Modern Techniques in Soil Ecology, Atlanta, GA, USA, 11–15 September 1989; Elsevier Science Bv: Atlanta, GA, USA; pp. 43–54.
- Freeman, C.; Ostle, N.; Kang, H. An enzymic “latch” on a global carbon store—A shortage of oxygen locks up carbon in peatlands by restraining a single enzyme. Nature 2001, 409, 149. [Google Scholar] [PubMed]
- Fenner, N.; Freeman, C. Drought-Induced carbon loss in peatlands. Nat. Geosci. 2011, 4, 895–900. [Google Scholar]
- Nistor, C.; Rose, A.; Farre, M.; Stoica, L.; Wollenberger, U.; Ruzgas, T.; Pfeiffer, D.; Barcelo, D.; Gorton, L.; Emneus, J. In-Field monitoring of cleaning efficiency in waste water treatment plants using two phenol-sensitive biosensors. In Proceedings of the 8th Workshop on Biosensors for Environmental Monitoring, Lisbon, Portugal, 11–13 September 2000; pp. 3–17.
- Tranvik, L.J.; Jansson, M. Climate change—Terrestrial export of organic carbon. Nature 2002, 415, 861–862. [Google Scholar]
- Evans, C.D.; Freeman, C.; Monteith, D.T.; Reynolds, B.; Fenner, N. Climate change—Terrestrial export of organic carbon—Reply. Nature 2002, 415, 862–862. [Google Scholar]
- Freeman, C.; Gresswell, R.; Guasch, H.; Hudson, J.; Lock, M.A.; Reynolds, B.; Sabater, F.; Sabater, S. The role of drought in the impact of climatic-change on the microbiota of peatland streams. Freshw. Biol. 1994, 32, 223–230. [Google Scholar]
- Tang, R.; Clark, J.M.; Bond, T.; Graham, N.; Hughes, D.; Freeman, C. Assessment of potential climate change impacts on peatland dissolved organic carbon release and drinking water treatment from laboratory experiments. Environ. Pollut. 2013, 173, 270–277. [Google Scholar] [PubMed]
- Whitworth, K.L.; Baldwin, D.S.; Kerr, J.L. Drought, floods and water quality: Drivers of a severe hypoxic blackwater event in a major river system (the Southern Murray-Darling Basin, Australia). J. Hydrol. 2012, 450, 190–198. [Google Scholar]
- Hamilton, S.K.; Sippel, S.J.; Calheiros, D.F.; Melack, J.M. An anoxic event and other biogeochemical effects of the pantanal wetland on the Paraguay River. Limnol. Oceanogr. 1997, 42, 257–272. [Google Scholar]
- Fontenot, Q.C.; Rutherford, D.A.; Kelso, W.E. Effects of environmental hypoxia associated with the annual flood pulse on the distribution of larval sunfish and shad in the Atchafalaya River basin, Louisiana. Trans. Am. Fish. Soc. 2001, 130, 107–116. [Google Scholar]
- Marin-Spiotta, E.; Gruley, K.E.; Crawford, J.; Atkinson, E.E.; Miesel, J.R.; Greene, S.; Cardona-Correa, C.; Spencer, R.G.M. Paradigm shifts in soil organic matter research affect interpretations of aquatic carbon cycling: Transcending disciplinary and ecosystem boundaries. Biogeochemistry 2014, 117, 279–297. [Google Scholar]
- Vidon, P.; Wagner, L.E.; Soyeux, E. Changes in the character of DOC in streams during storms in two midwestern watersheds with contrasting land uses. Biogeochemistry 2008, 88, 257–270. [Google Scholar]
- Inamdar, S.; Finger, N.; Singh, S.; Mitchell, M.; Levia, D.; Bais, H.; Scott, D.; McHale, P. Dissolved organic matter (DOM) concentration and quality in a forested mid-Atlantic watershed, USA. Biogeochemistry 2012, 108, 55–76. [Google Scholar]
- Fellman, J.B.; Hood, E.; Edwards, R.T.; D’Amore, D.V. Changes in the concentration, biodegradability, and fluorescent properties of dissolved organic matter during stormflows in coastal temperate watersheds. J. Geophys. Res. Biogeosci. 2009, 114. [Google Scholar] [CrossRef]
- Winterdahl, M.; Erlandsson, M.; Futter, M.N.; Weyhenmeyer, G.A.; Bishop, K. Intra-Annual variability of organic carbon concentrations in running waters: Drivers along a climatic gradient. Glob. Biogeochem. Cycle 2014, 28, 451–464. [Google Scholar]
- Sanderman, J.; Lohse, K.A.; Baldock, J.A.; Amundson, R. Linking soils and streams: Sources and chemistry of dissolved organic matter in a small coastal watershed. Water Resour. Res. 2009, 45. [Google Scholar] [CrossRef]
- Jencso, K.G.; McGlynn, B.L.; Gooseff, M.N.; Wondzell, S.M.; Bencala, K.E.; Marshall, L.A. Hydrologic connectivity between landscapes and streams: Transferring reach-and plot-scale understanding to the catchment scale. Water Resour. Res. 2009, 45. [Google Scholar] [CrossRef]
- Amon, R.M.W.; Rinehart, A.J.; Duan, S.; Louchouarn, P.; Prokushkin, A.; Guggenberger, G.; Bauch, D.; Stedmon, C.; Raymond, P.A.; Holmes, R.M.; et al. Dissolved organic matter sources in large arctic rivers. Geochim. Cosmochim. Acta 2012, 94, 217–237. [Google Scholar]
- Walker, S.A.; Amon, R.M.W.; Stedmon, C.A. Variations in high-latitude riverine fluorescent dissolved organic matter: A comparison of large arctic rivers. J. Geophys. Res. Biogeosci. 2013, 118, 1689–1702. [Google Scholar]
- Laudon, H.; Tetzlaff, D.; Soulsby, C.; Carey, S.; Seibert, J.; Buttle, J.; Shanley, J.; McDonnell, J.J.; McGuire, K. Change in winter climate will affect dissolved organic carbon and water fluxes in mid-to-high latitude catchments. Hydrol. Process. 2013, 27, 700–709. [Google Scholar]
- Foley, J.A.; DeFries, R.; Asner, G.P.; Barford, C.; Bonan, G.; Carpenter, S.R.; Chapin, F.S.; Coe, M.T.; Daily, G.C.; Gibbs, H.K.; et al. Global consequences of land use. Science 2005, 309, 570–574. [Google Scholar] [PubMed]
- Findlay, S.; Quinn, J.M.; Hickey, C.W.; Burrell, G.; Downes, M. Effects of land use and riparian flowpath on delivery of dissolved organic carbon to streams. Limnol. Oceanogr. 2001, 46, 345–355. [Google Scholar]
- Houghton, R.A. Land-Use change and the carbon-cycle. Glob. Chang. Biol. 1995, 1, 275–287. [Google Scholar]
- Gough, R.; Holliman, P.J.; Willis, N.; Jones, T.G.; Freeman, C. Influence of habitat on the quantity and composition of leachable carbon in the O2 horizon: Potential implications for potable water treatment. Lake Reserv. Manag. 2012, 28, 282–292. [Google Scholar]
- Yallop, A.R.; Clutterbuck, B.; Thacker, J. Increases in humic dissolved organic carbon export from upland peat catchments: The role of temperature, declining sulphur deposition and changes in land management. Clim. Res. 2010, 45, 43–56. [Google Scholar]
- Bricker, S.B.; Longstaf, B.; Dennison, W.; Jones, A.; Boicourt, K.; Wicks, C.; Woerner, J. Effects of nutrient enrichment in the nation’s estuaries: A decade of change. Harmful Algae 2008, 8, 21–32. [Google Scholar]
- Carpenter, S.R.; Caraco, N.F.; Correll, D.L.; Howarth, R.W.; Sharpley, A.N.; Smith, V.H. Nonpoint pollution of surface waters with phosphorus and nitrogen. Ecol. Appl. 1998, 8, 559–568. [Google Scholar]
- Fellman, J.B.; Hood, E.; Spencer, R.G.M. Fluorescence spectroscopy opens new windows into dissolved organic matter dynamics in freshwater ecosystems: A review. Limnol. Oceanogr. 2010, 55, 2452–2462. [Google Scholar]
- Wilson, H.F.; Xenopoulos, M.A. Effects of agricultural land use on the composition of fluvial dissolved organic matter. Nat. Geosci. 2009, 2, 37–41. [Google Scholar]
- Schlesinger, W. Biogeochemistry: An Analysis of Global Change; Academic Press: San Diego, CA, USA, 1991. [Google Scholar]
- Mierle, G.; Ingram, R. The role of humic substances in the mobilization of mercury from watersheds. Water Air Soil Pollut. 1991, 56, 349–357. [Google Scholar]
- Fabris, R.; Chowa, C.W.K.; Drikas, M.; Eikebrokk, B. Comparison of NOM character in selected Australian and Norwegian drinking waters. Water Res. 2008, 42, 4188–4196. [Google Scholar] [PubMed]
- Chow, A.T.; Guo, F.M.; Gao, S.D.; Breuer, R.S. Trihalomethane reactivity of water- and sodium hydroxide-extractable organic carbon fractions from peat soils. J. Environ. Qual. 2006, 35, 114–121. [Google Scholar] [PubMed]
- Fleck, J.A.; Bossio, D.A.; Fujii, R. Dissolved organic carbon and disinfection by-product precursor release from managed peat soils. J. Environ. Qual. 2004, 33, 465–475. [Google Scholar] [PubMed]
- Diaz, F.J.; Chow, A.T.; O’Geen, A.T.; Dahlgren, R.A.; Wong, P.K. Restored wetlands as a source of disinfection byproduct precursors. Environ. Sci. Technol. 2008, 42, 5992–5997. [Google Scholar] [PubMed]
- Saleh, D.K. Organic Carbon Trends, Loads, and Yields to the Sacramento-San Joaquin Delta, California, Water Years 1980–2000, 2nd ed.; Water Resources Investigation Report 03-4070; U.S. Geological Survey: Sacramento, CA, USA, 2007. [Google Scholar]
- Ledesma, J.L.; Köhler, S.J.; Futter, M.N. Long-Term dynamics of dissolved organic carbon: Implications for drinking water supply. Sci. Total Environ. 2012, 432, 1–11. [Google Scholar] [PubMed]
- Gough, R.; Holliman, P.J.; Willis, N.; Freeman, C. Dissolved organic carbon and trihalomethane precursor removal at a UK upland water treatment works. Sci. Total Environ. 2014, 468, 228–239. [Google Scholar] [PubMed]
- United States Environmental Protection Agency (US EPA). National primary drinking water regulations: Stage 2 disinfectants and disinfection byproducts rule; National primary and secondary drinking water regulations: Approval of analytical methods for chemical contaminants. Federal Register 2003, 68, 49548–49681. [Google Scholar]
- United States Environmental Protection Agency (US EPA). National primary drinking water regulations: Stage 2 disinfectants and disinfection byproducts rule. Federal Register 2006, 71, 388–493. [Google Scholar]
- Haaland, S.; Hongve, D.; Laudon, H.; Riise, G.; Vogt, R.D. Quantifying the drivers of the increasing colored organic matter in boreal surface waters. Environ. Sci. Technol. 2010, 44, 2975–2980. [Google Scholar] [PubMed]
- Eckard, R.S.; Hernes, P.J.; Bergamaschi, B.A.; Stepanauskas, R.; Kendall, C. Landscape scale controls on the vascular plant component of dissolved organic carbon across a freshwater delta. Geochim. Cosmochim. Acta 2007, 71, 5968–5984. [Google Scholar]
- Norwood, D.L.; Johnson, J.D.; Christman, R.F.; Hass, J.R.; Bobenrieth, M.J. Reactions of chlorine with selected aromatic models of aquatic humic material. Environ. Sci. Technol. 1980, 14, 187–190. [Google Scholar]
- Hanna, J.V.; Johnson, W.D.; Quezada, R.A.; Wilson, M.A.; Lu, X.Q. Characterization of aqueous humic substances before and after chlorination. Environ. Sci. Technol. 1991, 25, 1160–1164. [Google Scholar]
- Arnold, W.A.; Bolotin, J.; von Gunten, U.; Hofstetter, T.B. Evaluation of functional groups responsible for chloroform formation during water chlorination using compound specific isotope analysis. Environ. Sci. Technol. 2008, 42, 7778–7785. [Google Scholar] [PubMed]
- Hendricks, D.W. Water Treatment Unit Processes: Physical and Chemical. Civil and Environmental Engineering; CRC/Taylor & Francis: Boca Raton, FL, USA, 2006. [Google Scholar]
- Archer, A.D.; Singer, P.C. An evaluation of the relationship between SUVA and NOM coagulation using the ICR database. J. Am. Water Works Assoc. 2006, 98, 110–123. [Google Scholar]
- Archer, A.D.; Singer, P.C. Effect of SUVA and enhanced coagulation on removal of TOX precursors. J. Am. Water Works Assoc. 2006, 98, 97–107. [Google Scholar]
- Gang, D.C.; Clevenger, T.E.; Banerji, S.K. Effects of alum coagulation on speciation and distribution of trihalomethanes (THMs) and haloacetic acids (HAAs). J. Environ. Sci. Health Part A Toxic Hazard. Subst. Environ. Eng. 2005, 40, 521–534. [Google Scholar]
- Johnstone, D.W.; Sanchez, N.P.; Miller, C.M. Parallel factor analysis of excitation-emission matrices to assess drinking water disinfection byproduct formation during a peak formation period. Environ. Eng. Sci. 2009, 26, 1551–1559. [Google Scholar]
- Hussain, S.; van Leeuwen, J.; Chow, C.; Beecham, S.; Kamruzzaman, M.; Wang, D.S.; Drikas, M.; Aryal, R. Removal of organic contaminants from river and reservoir waters by three different aluminum-based metal salts: Coagulation adsorption and kinetics studies. Chem. Eng. J. 2013, 225, 394–405. [Google Scholar]
- Reckhow, D.A.; Rees, P.L.S.; Bryan, D. Watershed sources of disinfectant byproduct precursors. Water Supply 2004, 4, 61–69. [Google Scholar]
- Clark, J.M.; Chapman, P.J.; Heathwaite, A.L.; Adamson, J.K. Suppression of dissolved organic carbon by sulfate induced acidification during simulated droughts. Environ. Sci. Technol. 2006, 40, 1776–1783. [Google Scholar] [PubMed]
© 2014 by the authors; licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Pagano, T.; Bida, M.; Kenny, J.E. Trends in Levels of Allochthonous Dissolved Organic Carbon in Natural Water: A Review of Potential Mechanisms under a Changing Climate. Water 2014, 6, 2862-2897. https://doi.org/10.3390/w6102862
Pagano T, Bida M, Kenny JE. Trends in Levels of Allochthonous Dissolved Organic Carbon in Natural Water: A Review of Potential Mechanisms under a Changing Climate. Water. 2014; 6(10):2862-2897. https://doi.org/10.3390/w6102862
Chicago/Turabian StylePagano, Todd, Morgan Bida, and Jonathan E. Kenny. 2014. "Trends in Levels of Allochthonous Dissolved Organic Carbon in Natural Water: A Review of Potential Mechanisms under a Changing Climate" Water 6, no. 10: 2862-2897. https://doi.org/10.3390/w6102862
APA StylePagano, T., Bida, M., & Kenny, J. E. (2014). Trends in Levels of Allochthonous Dissolved Organic Carbon in Natural Water: A Review of Potential Mechanisms under a Changing Climate. Water, 6(10), 2862-2897. https://doi.org/10.3390/w6102862