
Successional Pathways of Riparian Vegetation Following Weir Gate Operations: Insights from the Geumgang River, South Korea


Abstract
1. Introduction
2. Materials and Methods
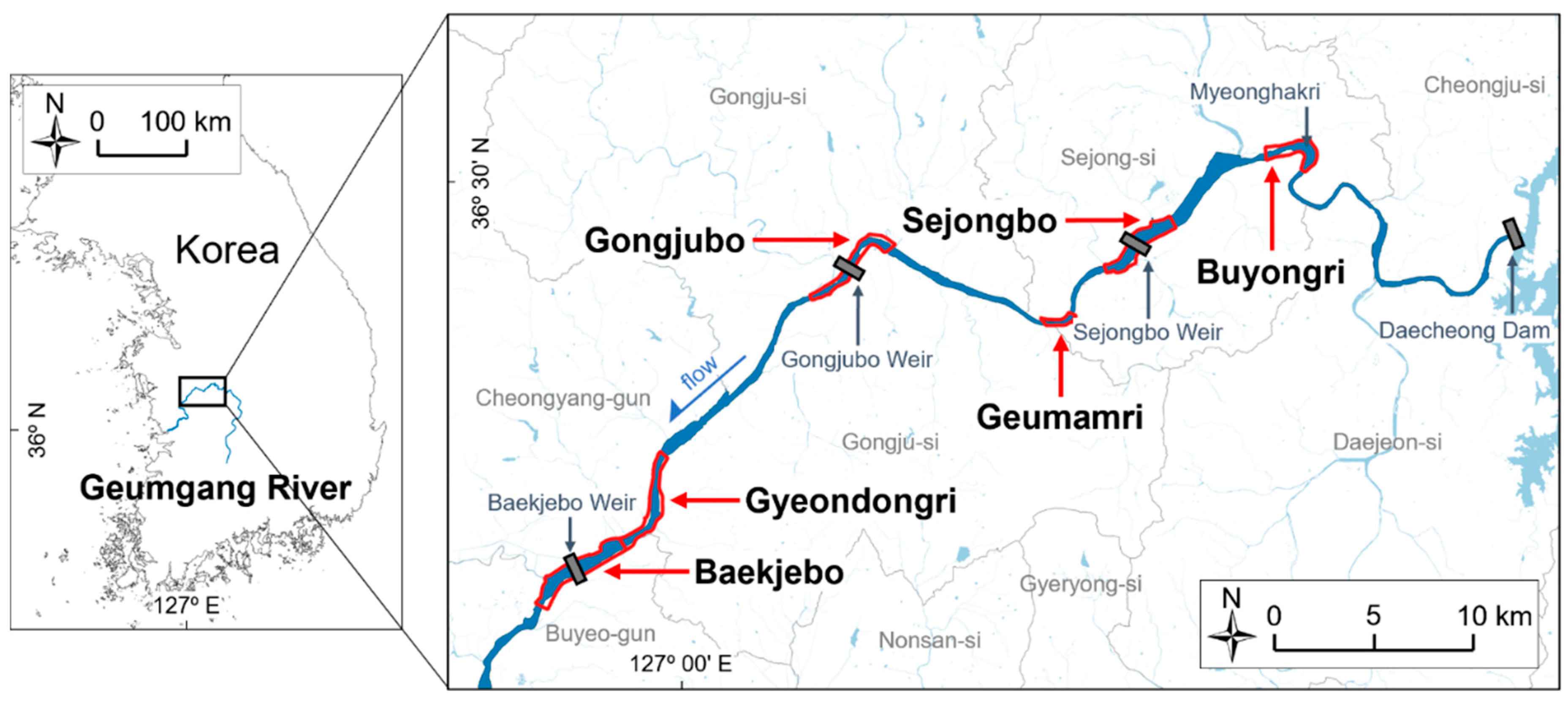
2.1. Study Site
2.2. Hydrological and Geomorphological Changes by the Weir Operation
2.3. Field Survey
2.4. Remote Sensing
2.5. Statistical Analysis
3. Results
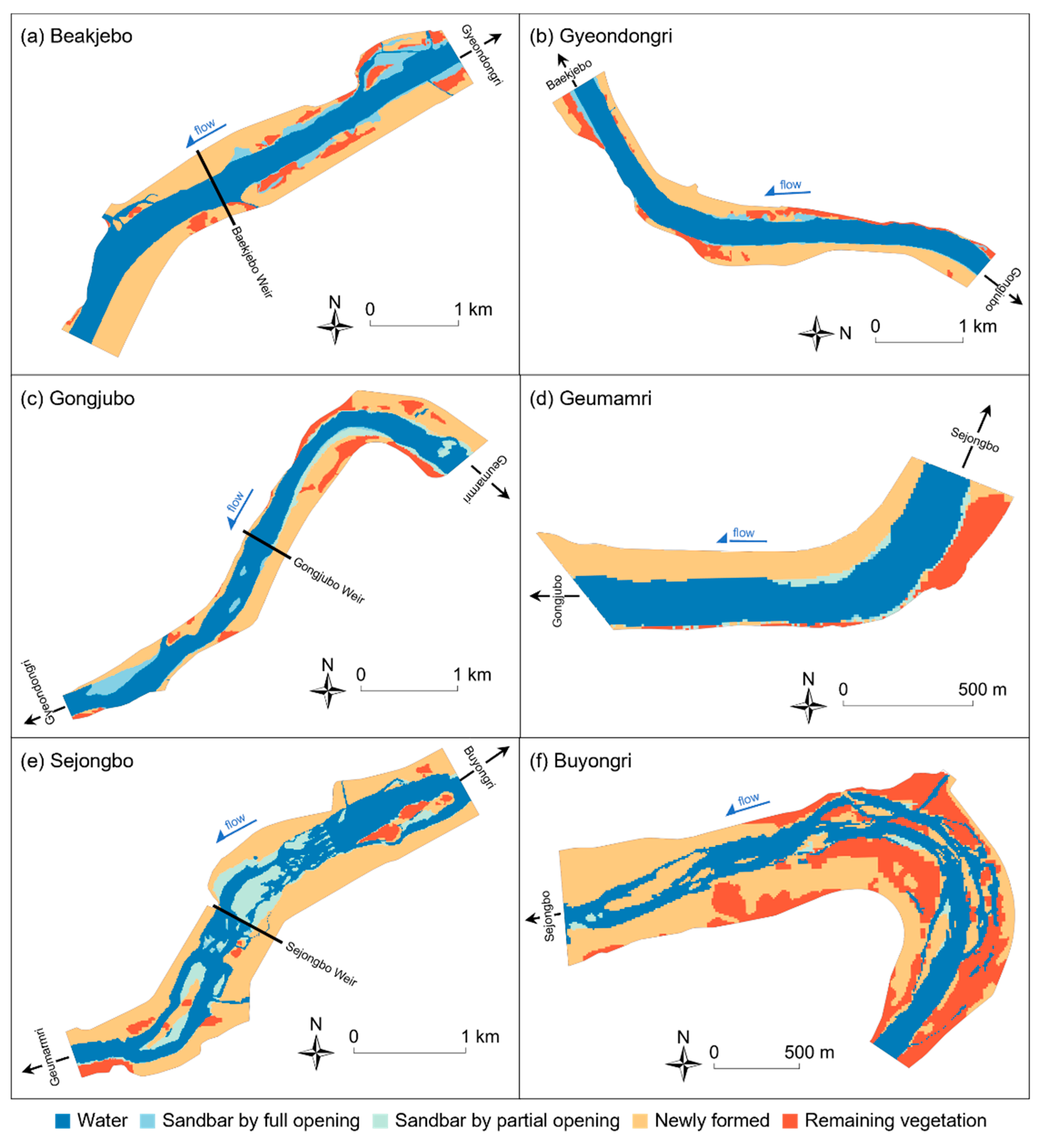
3.1. Geomorphological Changes
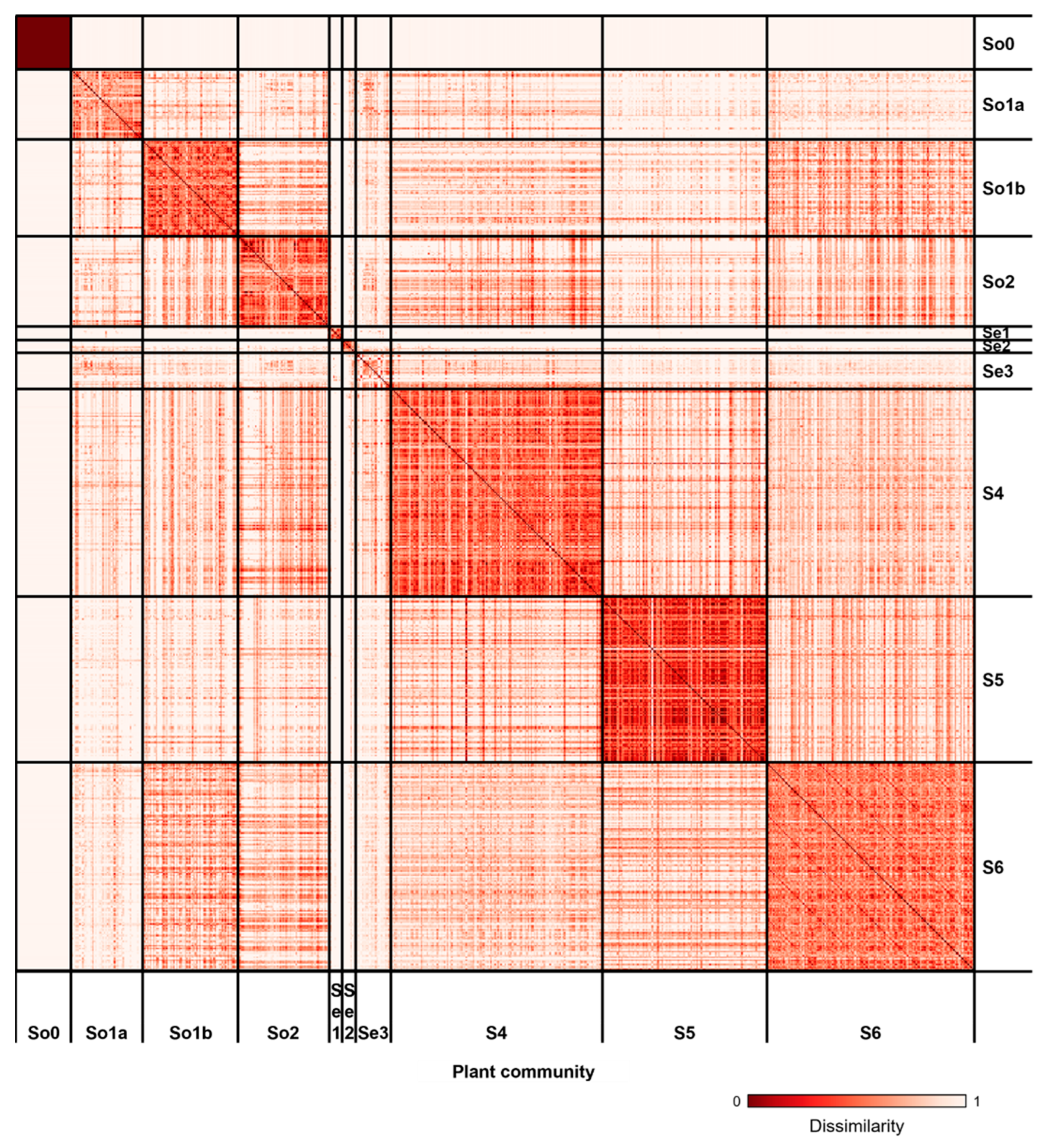
3.2. Vegetation Classification
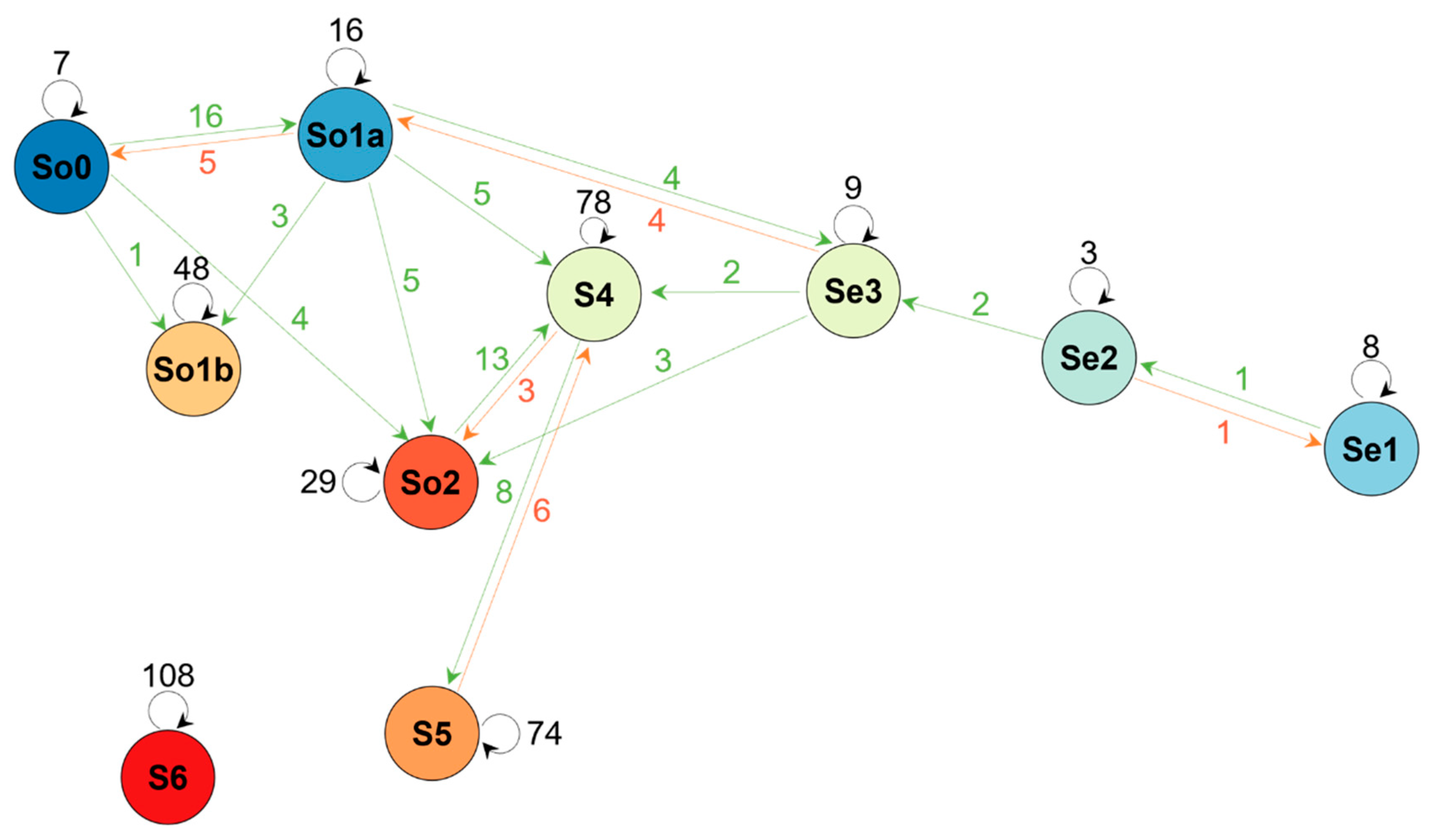
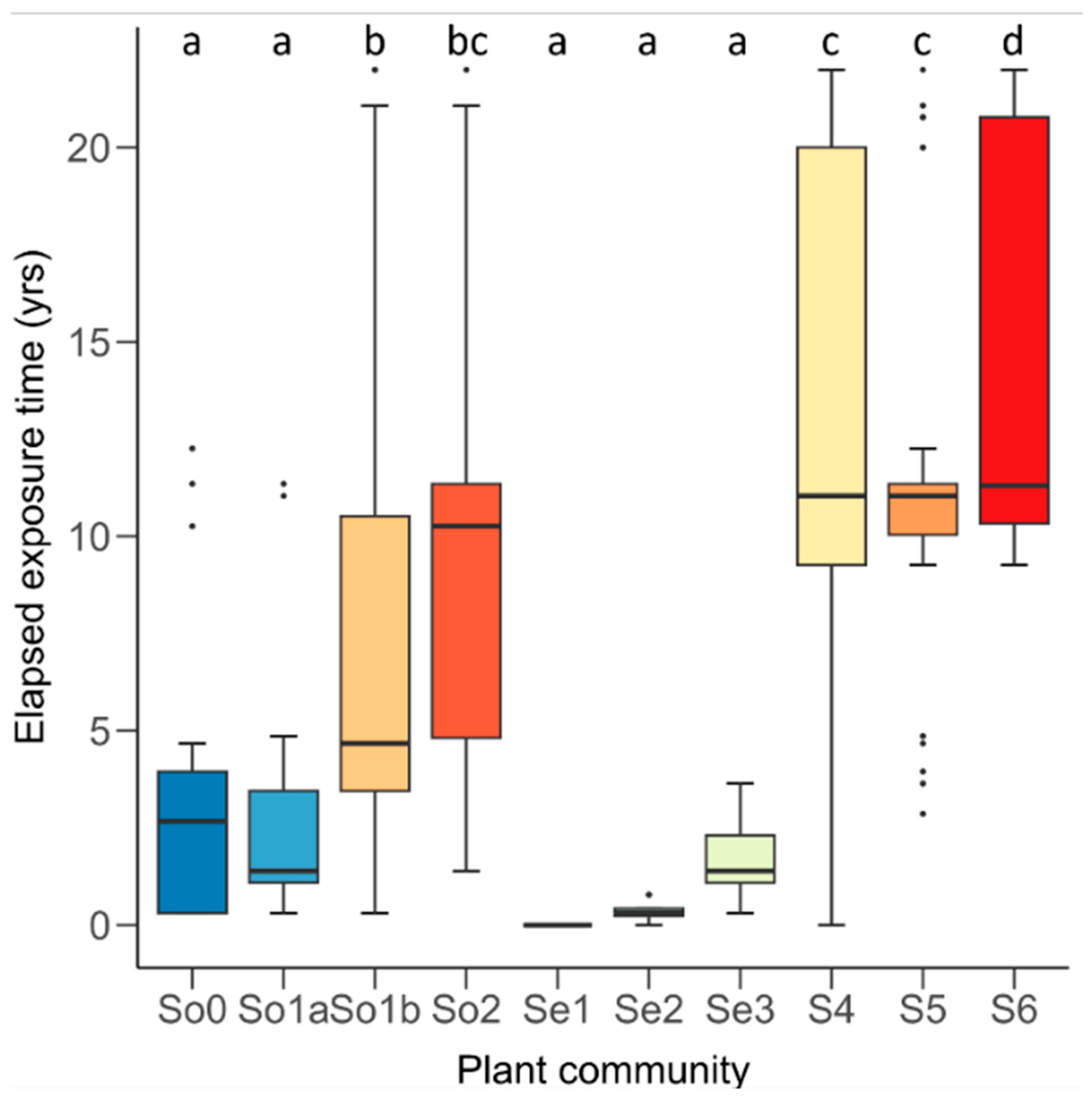
3.3. Vegetation Changes
4. Discussion
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Data Availability Statement
Conflicts of Interest
References
- Ghosh, S.; Mistri, B. Geographic concerns on flood climate and flood hydrology in monsoon-dominated Damodar River basin, Eastern India. Geogr. J. 2015, 2015, 486740. [Google Scholar] [CrossRef]
- Wright, S.A.; Kaplinski, M. Flow structures and sandbar dynamics in a canyon river during a controlled flood, Colorado River, Arizona. J. Geophys. Res. 2011, 116, F01019. [Google Scholar] [CrossRef]
- Blom, C.W.P.M.; Bögemann, G.M.; Laan, P.; van der Sman, A.J.M.; van de Steeg, H.M.; Voesenek, L.A.C.J. Adaptations to flooding in plants from river areas. Aquat. Bot. 1990, 38, 29–47. [Google Scholar] [CrossRef]
- Lee, Y.K.; Kim, J.W. Riparian Vegetation of South Korea; Keimyung University Press: Daegu, Republic of Korea, 2005. [Google Scholar]
- Merritt, D.M.; Cooper, D.J. Riparian vegetation and channel change in response to river regulation: A comparative study of regulated and unregulated streams in the Green River Basin, USA. River Res. Appl. 2000, 16, 543–564. [Google Scholar] [CrossRef]
- Lee, C.; Lee, K.; Kim, H.; Baek, D.; Kim, W.; Woo, H.; Kim, D. Impacts of an extreme flood event on the riparian vegetation of a monsoonal cobble-bed stream in southern Korea: A multiscale fluvial biogeomorphic framework. River Res. Appl. 2022, 38, 1101–1114. [Google Scholar] [CrossRef]
- Corenblit, D.; Steiger, J.; Gurnell, A.M.; Tabacchi, E.; Roques, L. Control of sediment dynamics by vegetation as a key function driving biogeomorphic succession within fluvial corridors. J. Br. Stud. 2009, 34, 1790–1810. [Google Scholar] [CrossRef]
- van Andel, J.; Bakker, J.P.; Grootjans, A.P. Mechanisms of vegetation succession: A review of concepts and perspectives. Acta Bot. Neerl. 1993, 42, 413–433. [Google Scholar]
- Bornette, G.; Amoros, C.; Castella, C.; Beffy, C.J. Succession and fluctuation in the aquatic vegetation of two former Rhône River channels. Vegetatio 1994, 110, 171–184. [Google Scholar] [CrossRef]
- Gergely, A.; Hahn, I.; Mészáros-Draskovits, R.; Simon, T.; Szabó, M.; Barabás, S. Vegetation succession in a newly exposed Danube riverbed. Appl. Veg. Sci. 2001, 4, 35–40. [Google Scholar] [CrossRef]
- Zhao, B.; Yan, Y.; Guo, H.; He, M.; Gu, Y.; Li, B. Monitoring rapid vegetation succession in estuarine wetland using time series MODIS-based indicators: An application in the Yangtze River Delta area. Ecol. Indic. 2009, 9, 346–356. [Google Scholar] [CrossRef]
- Prach, K.; Petřík, P.; Brož, Z.; Song, J.S. Vegetation succession on river sediments along the Nakdong River, South Korea. Folia Geobot. 2014, 49, 507–519. [Google Scholar] [CrossRef]
- Bourgeois, B.; Boutin, C.; Vanasse, A.; Poulin, M. Divergence between riparian seed banks and standing vegetation increases along successional trajectories. J. Veg. Sci. 2017, 28, 787–797. [Google Scholar] [CrossRef]
- Williams, B.S.; D’Amico, E.; Kastens, J.H.; Thorp, J.H.; Flotemersch, J.E.; Thoms, M.C. Automated riverine landscape characterization: GIS-based tools for watershed-scale research, assessment, and management. Environ. Monit. Assess. 2013, 185, 7485–7499. [Google Scholar] [CrossRef]
- Sukhodolov, A.N. Field-based research in fluvial hydraulics: Potential, paradigms and challenges. J. Hydraul. Res. 2015, 53, 1–19. [Google Scholar] [CrossRef]
- Marcus, W.A.; Fonstad, M.A. Remote sensing of rivers: The emergence of a subdiscipline in the river sciences. Earth Surf. Process. Landf. 2010, 35, 1867–1872. [Google Scholar] [CrossRef]
- Mouat, D.A.; Lancaster, J. Use of remote sensing and GIS to identify vegetation change in the upper San Pedro River watershed, Arizona. Geocarto Int. 1996, 11, 55–67. [Google Scholar] [CrossRef]
- Temesgen, H.; Nyssen, J.; Zenebe, A.; Haregeweyn, N.; Kindu, M.; Lemenih, M.; Haile, M. Ecological succession and land use changes in a lake retreat area (Main Ethiopian Rift Valley). J. Arid Environ. 2013, 91, 53–60. [Google Scholar] [CrossRef]
- Han, M.; Brierley, G.; Li, B.; Li, Z.; Li, X. Impacts of flow regulation on geomorphic adjustment and riparian vegetation succession along an anabranching reach of the Upper Yellow River. Catena 2020, 190, 104561. [Google Scholar] [CrossRef]
- Huylenbroeck, L.; Laslier, M.; Dufour, S.; Georges, B.; Lejeune, P.; Michez, A. Using remote sensing to characterize riparian vegetation: A review of available tools and perspectives for managers. J. Environ. Manag. 2020, 267, 110652. [Google Scholar] [CrossRef]
- Rusnák, M.; Goga, T.; Michaleje, L.; Šulc Michalková, M.; Máčka, Z.; Bertalan, L.; Kidová, A. Remote sensing of riparian ecosystems. Remote Sens. 2022, 14, 2645. [Google Scholar] [CrossRef]
- Lara-Alvarez, C.; Flores, J.J.; Rodriguez-Rangel, H.; Lopez-Farias, R. A literature review on satellite image time series forecasting: Methods and applications for remote sensing. Wiley Interdiscip. Rev. Data Min. Knowl. Discov. 2024, 14, e1528. [Google Scholar] [CrossRef]
- Corenblit, D.; Tabacchi, E.; Steiger, J.; Gurnell, A.M. Reciprocal interactions and adjustments between fluvial landforms and vegetation dynamics in river corridors: A review of complementary approaches. Earth-Sci. Rev. 2007, 84, 56–86. [Google Scholar] [CrossRef]
- Camporeale, C.; Perucca, E.; Ridolfi, L.; Gurnell, A.M. Modeling the interactions between river morphodynamics and riparian vegetation. Rev. Geophys. 2013, 51, 379–414. [Google Scholar] [CrossRef]
- Del Tánago, M.G.; Martínez-Fernández, V.; Aguiar, F.C.; Bertoldi, W.; Dufour, S.; de Jalón, D.G.; Garófano-Gómez, V.; Mandzukovski, D.; Rodríguez-González, P.M. Improving river hydromorphological assessment through better integration of riparian vegetation: Scientific evidence and guidelines. J. Environ. Manag. 2021, 292, 112730. [Google Scholar] [CrossRef] [PubMed]
- Lee, K.; Lee, C.; Baek, D.; Park, G.; Shim, T.; Kim, W.; Cho, K.-H.; Kim, D. Vegetation destruction during an extreme flood: Multilevel modelling of an entire river in southern Korea. Hydrol. Process. 2023, 37, e15051. [Google Scholar] [CrossRef]
- Dias-Silva, K.; Vieira, T.B.; de Matos, T.P.; Juen, L.; Simião-Ferreira, J.; Hughes, R.M.; Júnior, P.D.M. Measuring stream habitat conditions: Can remote sensing substitute for field data? Sci. Total Environ. 2021, 788, 147617. [Google Scholar] [CrossRef] [PubMed]
- HRFCO (Hangang River Flood Control Office). Korean River Catalog; Ministry of Land Transport and Maritime Affairs: Seoul, Republic of Korea, 2014. [Google Scholar]
- Choi, I.C.; Shin, H.J.; Nguyen, T.T.; Tenhunen, J. Water policy reforms in South Korea: A historical review and ongoing challenges for sustainable water governance and management. Water 2017, 9, 717. [Google Scholar] [CrossRef]
- Kim, D.; Lee, C.; Kim, H.; Ock, G.; Cho, K.-H. Changes in landscape characteristics of stream habitats with the construction and operation of river-crossing structures in the Geum-gang River, South Korea. Ecol. Resil. Infrastruct. 2021, 8, 64–78. [Google Scholar] [CrossRef]
- Ock, G.; Choi, M.; Kim, J.-C.; Park, H.-G.; Han, J.H. Evaluation of habitat diversity changes by weir operation of the Sejongbo Weir in Geum River using high-resolution aerial photographs. Ecol. Resil. Infrastruct. 2020, 7, 366–373. [Google Scholar] [CrossRef]
- MOLTM (Ministry of Land, Transport and Maritime). Master Plan of Four Major Rivers Restoration; Ministry of Land, Transport and Maritime: Gwacheon, Republic of Korea, 2009. [Google Scholar]
- MOLIT (Ministry of Land, Infrastructure and Transport). Study on River Change Monitoring and Evaluation; Ministry of Land, Infrastructure and Transport: Sejong, Republic of Korea, 2016. [Google Scholar]
- Lee, C.; Kim, H.; Cho, K.H. Spatial distribution and successional changes of riparian vegetation on sandbars exposed after watergate-opening of weirs in the Geumgang River, South Korea. Ecol. Resil. Infrastruct. 2022, 9, 194–205. [Google Scholar] [CrossRef]
- MOE (Ministry of Environment). Aquatic Ecosystem Monitoring in the Weirs of Geum River; Ministry of Environment: Sejong, Republic of Korea, 2021. [Google Scholar]
- MOE (Ministry of Environment). Research on the Effect of Weir Construction on Freshwater Ecosystem; Ministry of Environment: Sejong, Republic of Korea, 2010. [Google Scholar]
- Lee, C.; Cho, K.H. Development of a method for tracking sandbar formation by weir-gate opening using multispectral satellite imagery in the Geumgang River, South Korea. Ecol. Resil. Infrastruct. 2023, 10, 135–142. [Google Scholar] [CrossRef]
- MOLIT (Ministry of Land, Infrastructure and Transport); MOE (Ministry of Environment); MAFRA (Ministry of Agriculture, Food and Rural Affairs). The Report on the Optimal Linkage Operation Plan Between Dam-Weir-Reservoir; Ministry of Land, Infrastructure and Transport, Ministry of Environment, Ministry of Agriculture, Food and Rural Affairs: Sejong, Republic of Korea, 2017. [Google Scholar]
- MOE (Ministry of Environment). Dam Operation and Monitoring Plan; Ministry of Environment: Sejong, Republic of Korea, 2022. [Google Scholar]
- Ellenberg, D.; Mueller-Dombois, D. Aims and Methods of Vegetation Ecology; Wiley: New York, NY, USA, 1974. [Google Scholar]
- Van der Maarel, E.; Franklin, J. Vegetation Ecology, 2nd ed.; Wiley-Blackwell: Hoboken, NJ, USA, 2012. [Google Scholar]
- Gao, B.C. NDWI—A normalized difference water index for remote sensing of vegetation liquid water from space. Remote Sens. Environ. 1996, 58, 257–266. [Google Scholar] [CrossRef]
- Carlson, T.N.; Ripley, D.A. On the relation between NDVI, fractional vegetation cover, and leaf area index. Remote Sens. Environ. 1997, 62, 241–252. [Google Scholar] [CrossRef]
- Otsu, N. A threshold selection method from gray-level histograms. IEEE Trans. Syst. Man Cybern. Syst. 1979, 9, 62–66. [Google Scholar]
- Piedelobo, L.; Taramelli, A.; Schiavon, E.; Valentini, E.; Molina, J.L.; Nguyen Xuan, A.; González-Aguilera, D. Assessment of green infrastructure in Riparian zones using Copernicus programme. Remote Sens. 2019, 11, 2967. [Google Scholar] [CrossRef]
- Valerio, F.; Godinho, S.; Ferraz, G.; Pita, R.; Gameiro, J.; Silva, B.; Marques, A.T.; Silva, J.P. Multi-temporal remote sensing of inland surface waters: A fusion of Sentinel-1 & 2 data applied to small seasonal ponds in semiarid environments. Int. J. Appl. Earth Obs. Geoinf. 2024, 135, 104283. [Google Scholar] [CrossRef]
- Salvoldi, M.; Siaki, G.; Sprintsin, M.; Karnieli, A. Burned area mapping using multi-temporal sentinel-2 data by applying the relative differenced aerosol-free vegetation index (RdAFRI). Remote Sens. 2020, 12, 2753. [Google Scholar] [CrossRef]
- Pau, G.; Fuchs, F.; Sklyar, O.; Boutros, M.; Huber, W. EBImage—An R package for image processing with applications to cellular phenotypes. J. Bioinform. 2010, 26, 979–981. [Google Scholar] [CrossRef]
- R Core Team. R: A Language and Environment for Statistical Computing. Available online: https://www.r-project.org (accessed on 30 June 2023).
- QGIS.org. QGIS Geographic Information System. Available online: http://www.qgis.org (accessed on 30 June 2023).
- Roberts, D.W. Vegetation classification by two new iterative reallocation optimization algorithms. Plant Ecol. 2015, 216, 741–758. [Google Scholar] [CrossRef]
- Aho, K.; Roberts, D.W.; Weaver, T. Using geometric and non-geometric internal evaluators to compare eight vegetation classification methods. J. Veg. Sci. 2008, 19, 549–562. [Google Scholar] [CrossRef]
- Pakgohar, N.; Eshaghi Rad, J.; Gholami, G.; Alijanpour, A.; Roberts, D.W. A comparative study of hard clustering algorithms for vegetation data. J. Veg. Sci. 2021, 32, e13042. [Google Scholar] [CrossRef]
- Oksanen, J.; Simpson, G.; Blanchet, F.; Kindt, R.; Legendre, P.; Minchin, P.R.; O’Hara, R.B.; Solymos, P.; Stevens, M.H.H.; Szoecs, E.; et al. Vegan: Community Ecology Package. Available online: https://CRAN.R-project.org/package=vegan (accessed on 15 December 2022).
- Hakkenberg, C.R.; Peet, R.K.; Urban, D.L.; Song, C. Modeling plant composition as community continua in a forest landscape with LiDAR and hyperspectral remote sensing. Ecol. Appl. 2018, 28, 177–190. [Google Scholar] [CrossRef] [PubMed]
- Lengyel, A.; Roberts, D.W.; Botta-Dukát, Z. Comparison of silhouette-based reallocation methods for vegetation classification. J. Veg. Sci. 2021, 32, e12984. [Google Scholar] [CrossRef]
- Roberts, D.W. Optpart: Optimal Partitioning of Similarity Relations. R Package Version 3.0-3. Available online: https://CRAN.R-project.org/package=optpart (accessed on 1 June 2023).
- De Cáceres, M.; Legendre, P. Associations between species and groups of sites: Indices and statistical inference. Ecology 2009, 90, 3566–3574. [Google Scholar] [CrossRef] [PubMed]
- Pielou, E.C. A quick method of determining the diversity of foraminiferal assemblages. J. Paleontol. 1979, 53, 1237–1242. [Google Scholar]
- Choung, Y.; Min, B.M.; Lee, K.S.; Cho, K.-H.; Joo, K.Y.; Hyun, J.-O.; Na, R.H.; Oh, H.K.; Nam, G.-H.; Lee, J. Categorized wetland preference and life forms of the vascular plants in the Korean Peninsula. J. Ecol. Environ. 2021, 45, 1–6. [Google Scholar] [CrossRef]
- Laliberté, E.; Legendre, P.A. Distance-based framework for measuring functional diversity from multiple traits. Ecology 2010, 91, 299–305. [Google Scholar] [CrossRef]
- Csardi, G.; Nepusz, T. igraph: Network Analysis and Visualization in R. Available online: https://zenodo.org/records/14736815 (accessed on 8 July 2023).
- Poff, N.L.; Allan, J.D.; Bain, M.B.; Karr, J.R.; Prestegaard, K.L.; Richter, B.D.; Sparks, R.E.; Stromberg, J.C. The natural flow regime. BioScience 1997, 47, 769–784. [Google Scholar] [CrossRef]
- Ligon, F.K.; Dietrich, W.E.; Trush, W.J. Downstream ecological effects of dams. BioScience 1995, 45, 183–192. [Google Scholar] [CrossRef]
- Nilsson, C.; Berggren, K. Alterations of riparian ecosystems caused by river regulation. BioScience 2000, 50, 783–792. [Google Scholar] [CrossRef]
- Yang, J.; Li, E.H.; Yang, C.; Xia, Y.; Zhou, R. Effects of south-to-north water diversion project cascade dams on riparian vegetation along the middle and lower reaches of the Hanjiang River, China. Front. Plant Sci. 2022, 13, 849010. [Google Scholar] [CrossRef] [PubMed]
- Bellmore, J.R.; Duda, J.J.; Craig, L.S.; Greene, S.L.; Torgersen, C.E.; Collins, M.J.; Vittum, K.M. Status and trends of dam removal research in the United States. WIREs Water 2017, 4, e1164. [Google Scholar] [CrossRef]
- Doyle, M.W.; Stanley, E.H.; Harbor, J.M. Stream ecosystem response to small dam removal: Lessons from the heartland. Geomorphology 2005, 71, 227–244. [Google Scholar] [CrossRef]
- Magilligan, F.J.; Graber, B.E.; Nislow, K.H.; Chipman, J.W.; Sneddon, C.S.; Fox, C.A. River restoration by dam removal: Enhancing connectivity at watershed scales. Elem. Sci. Anth. 2016, 4, 000108. [Google Scholar] [CrossRef]
- Bednarek, A.T. Undamming rivers: A review of the ecological impacts of dam removal. Environ. Manag. 2001, 27, 803–814. [Google Scholar] [CrossRef]
- Kim, J.W.; Lee, S.E.; Lee, J. Hwasan wetland vegetation in Gunwi, South Korea: With a phytosociological focus on alder Alnus japonica forests. Korean J. Ecol. Environ. 2017, 50, 70–78. [Google Scholar] [CrossRef]
- Corenblit, D.; Piégay, H.; Arrignon, F.; González-Sargas, E.; Bonis, A.; Davies, N.S.; Ebengo, D.M.; Garófano-Gómez, V.; Gurnell, A.M.; Henry, A.L.; et al. Interactions between vegetation and river morphodynamics. Part I: Research clarifications and challenges. Earth-Sci. Rev. 2024, 253, 104769. [Google Scholar] [CrossRef]
- Cho, H.-J. Mechanisms and Model of Early Succession of Riparian Vegetation in a Sandy Stream. Ph.D. Thesis, Inha University, Incheon, Republic of Korea, 2012. [Google Scholar]
- Jin, S.-N. Modeling the Distribution of Floodplain Vegetation in the Medium Sized Stream of the Central Korean Peninsula. Ph.D. Thesis, Inha University, Incheon, Republic of Korea, 2017. [Google Scholar]
- Lim, B.S.; Seol, J.; Kim, A.R.; An, J.H.; Lim, C.H.; Lee, C.S. Succession of the abandoned rice fields restores the riparian forest. Int. J. Environ. Res. Public Health 2022, 19, 10416. [Google Scholar] [CrossRef]
- Walker, L.R.; Del Moral, R. Primary Succession and Ecosystem Rehabilitation; Cambridge University Press: Cambridge, UK, 2003. [Google Scholar]
- Magdaleno, F.; Fernández, J.A. Hydromorphological alteration of a large Mediterranean river: Relative role of high and low flows on the evolution of riparian forests and channel morphology. River Res. Appl. 2011, 27, 374–387. [Google Scholar] [CrossRef]
- Sanchis-Ibor, C.; Segura-Beltrán, F.; Navarro-Gómez, A. Channel forms and vegetation adjustment to damming in a Mediterranean gravel-bed river (Serpis River, Spain). River Res. Appl. 2019, 35, 37–47. [Google Scholar] [CrossRef]
- Song, F.; Zhang, W.; Yuan, T.; Ji, Z.; Cao, Z.; Xu, B.; Lu, L.; Zou, S. UAV quantitative remote sensing of riparian zone vegetation for river and lake health assessment: A review. Remote Sens. 2024, 16, 3560. [Google Scholar] [CrossRef]
- Seok, J.E.; Lim, B.S.; Moon, J.S.; Kim, G.S.; Lee, C.S. Spatial distribution of vegetation on stream bars and the riparian zone reflects successional pattern due to fluid dynamics of river. Water 2023, 15, 1493. [Google Scholar] [CrossRef]
- Casado, A.; Peiry, J.L.; Campo, A.M. Geomorphic and vegetation changes in a meandering dryland river regulated by a large dam, Sauce Grande River, Argentina. Geomorphology 2016, 268, 21–34. [Google Scholar] [CrossRef]
- Woo, H.; Park, M.; Cho, K.-H.; Cho, H.; Chung, S. Recruitment and succession of riparian vegetation in alluvial river regulated by upstream dams—Focused on the Nakdong River downstream Andong and Imha Dams. J. Korea Water Resour. 2010, 43, 455–469. [Google Scholar] [CrossRef]
- Müllerová, A.; Řehounková, K.; Prach, K. Succession of aquatic and littoral vegetation in disused sandpits. Land Degrad. Dev. 2022, 33, 257–268. [Google Scholar] [CrossRef]
- Cho, H.J.; Jin, S.N.; Lee, H.; Marrs, R.H.; Cho, K.-H. The relationship between the soil seed bank and above-ground vegetation in a sandy floodplain, South Korea. Ecol. Resil. Infrastruct. 2018, 5, 145–155. [Google Scholar] [CrossRef]
- Lee, P.H.; Son, S.G.; Kim, C.S.; Oh, K.H. Population dynamics of Salix nipponica and S. koreensis during the riverbed sedimentation in the wetland of the Nam-River. J. Wet. Res. 2000, 2, 95–107. [Google Scholar]
- Azami, K.; Fukuyama, A.; Asaeda, T.; Takechi, Y.; Nakazawa, S.; Tanida, K. Conditions of establishment for the Salix community at lower-than-normal water levels along a dam reservoir shoreline. Landsc. Ecol. Eng. 2013, 9, 227–238. [Google Scholar] [CrossRef]
- Ward, J.V. An expansive perspective of riverine landscapes: Pattern and process across scales. GAIA 1997, 6, 52–60. [Google Scholar] [CrossRef]
- MOE (Ministry of Environment). Aquatic Ecosystem Monitoring in the Weirs of Geum River; Ministry of Environment: Sejong, Republic of Korea, 2016. [Google Scholar]
- Gudžinskas, Z.; Taura, L. Scirpus radicans (Cyperaceae) a newly-discovered native species in Lithuania: Population, habitats and threats. Biodivers. Data J. 2021, 9, e65674. [Google Scholar] [CrossRef]
- Corenblit, D.; Piégay, H.; Arrignon, F.; González-Sargas, E.; Bonis, A.; Ebengo, D.M.; Garófano-Gómez, V.; Gurnell, A.M.; Henry, A.L.; Hortobágyi, B.; et al. Interactions between vegetation and river morphodynamics. Part II: Why is a functional trait framework important? Earth-Sci. Rev. 2024, 253, 104709. [Google Scholar]
- Ghimire, P.; Lei, D.; Juan, N. Effect of image fusion on vegetation index quality—A comparative study from Gaofen-1, Gaofen-2, Gaofen-4, Landsat-8 OLI and MODIS Imagery. Remote Sens. 2020, 12, 1550. [Google Scholar] [CrossRef]
- Wang, Q.; Blackburn, G.A.; Onojeghuo, A.O.; Dash, J.; Zhou, L.; Zhang, Y.; Atkinson, P.M. Fusion of Landsat 8 OLI and Sentinel-2 MSI data. IEEE Trans. Geosci. Remote Sens. 2017, 55, 3885–3899. [Google Scholar] [CrossRef]
- Guan, H.; Su, Y.; Hu, T.; Chen, J.; Guo, Q. An object-based strategy for improving the accuracy of spatiotemporal satellite imagery fusion for vegetation-mapping applications. Remote Sens. 2019, 11, 2927. [Google Scholar] [CrossRef]
- Jovanovic, N.; Garcia, C.L.; Bugan, R.D.; Teich, I.; Rodriguez, C.M.G. Validation of remotely-sensed evapotranspiration and NDWI using ground measurements at Riverlands, South Africa. Water SA 2014, 40, 211–220. [Google Scholar] [CrossRef]
- Wen, J.; Wu, X.; You, D.; Ma, X.; Ma, D.; Wang, J.; Xiao, Q. The main inherent uncertainty sources in trend estimation based on satellite remote sensing data. Theor. Appl. Climatol. 2023, 151, 915–934. [Google Scholar] [CrossRef]
- Ashok, A.; Rani, H.P.; Jayakumar, K.V. Monitoring of dynamic wetland changes using NDVI and NDWI based Landsat imagery. Remote Sens. Appl. Soc. Environ. 2021, 23, 100547. [Google Scholar] [CrossRef]
- Lee, J.; Lim, J.; Lee, J.; Park, J.; Won, M. Ground-based NDVI network: Early validation practice with Sentinel-2 in South Korea. Sensors 2024, 24, 1892. [Google Scholar] [CrossRef]




{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Event | Year | ||||||||||||||||||||||||||||||||||||||||||||||||||||||
|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|
| 2010 | 2011 | 2012 | ... | 2017 | 2018 | 2019 | 2020 | 2021 | 2022 | ||||||||||||||||||||||||||||||||||||||||||||||
| Weir construction | ◀ | ||||||||||||||||||||||||||||||||||||||||||||||||||||||
| Water gate operation | |||||||||||||||||||||||||||||||||||||||||||||||||||||||
| Baekjebo Weir | ○ | ||||||||||||||||||||||||||||||||||||||||||||||||||||||
| Gongjubo Weir | ◐ | ○ | |||||||||||||||||||||||||||||||||||||||||||||||||||||
| Sejongbo Weir | ◐ | ○ | |||||||||||||||||||||||||||||||||||||||||||||||||||||
| Satellite image | |||||||||||||||||||||||||||||||||||||||||||||||||||||||
| Baekjebo Weir | ★ | ★ | ★ | ||||||||||||||||||||||||||||||||||||||||||||||||||||
| Gongjubo Weir | ★ | ★ | ★ | ★ | |||||||||||||||||||||||||||||||||||||||||||||||||||
| Sejongbo Weir | ★ | ★ | ★ | ★ | |||||||||||||||||||||||||||||||||||||||||||||||||||
| Field survey | ★ | ★ | ★ | ★ | |||||||||||||||||||||||||||||||||||||||||||||||||||
| Section | Area (ha) | ||||
|---|---|---|---|---|---|
| Newly Exposed Sandbar | Pre-Existing Vegetation | Total | |||
| By Full Opening | By Partial Opening | Newly Formed | Remaining | ||
| Baekjebo | 34.0 (14%) | - | 171.4 (71%) | 35.1 (15%) | 240.5 (100%) |
| Upstream | 34.0 (22%) | - | 91.7 (59%) | 29.7 (19%) | 155.4 (100%) |
| Downstream | 0 (0%) | - | 79.7 (94%) | 5.4 (6%) | 85.1 (100%) |
| Gyeondongri | 13.3 (10%) | - | 88.4 (67%) | 29.4 (23%) | 131.1 (100%) |
| Gongjubo | 13.8 (10%) | 15.5 (11%) | 86.9 (61%) | 25.0 (18%) | 141.2 (100%) |
| Upstream | 1.8 (2%) | 15.5 (17%) | 51.5 (57%) | 21.5 (24%) | 90.3 (100%) |
| Downstream | 12.0 (23%) | 0 (0%) | 35.4 (70%) | 3.5 (7%) | 50.9 (100%) |
| Geumarmri | 0.1 (0%) | 2.2 (8%) | 18.6 (72%) | 5.2 (20%) | 26.1 (100%) |
| Sejongbo | 15.8 (8%) | 36.3 (19%) | 141.4 (72%) | 14.6 (8%) | 193.7 (100%) |
| Upstream | 14.4 (13%) | 11.8 (10%) | 81.7 (73%) | 6.6 (6%) | 114.5 (100%) |
| Downstream | 1.4 (2%) | 10.1 (13%) | 59.7 (75%) | 8.0 (10%) | 79.2 (100%) |
| Buyongri | 1.8 (2%) | 0.9 (1%) | 85.8 (60%) | 54.0 (37%) | 142.5 (100%) |
| Community | Indicator Species | ||||
|---|---|---|---|---|---|
| Name | Abb. | Scientific Name | Abb. | Indicator Value | p |
| Bare bar | So0 | - | - | - | - |
| Persicaria lapathifolia | So1a | Persicaria lapathifolia | Pl | 0.860 | <0.001 |
| Chenopodium album | Cb | 0.352 | 0.014 | ||
| Salix triandra subsp. nipponica | So1b | Salix triandra subsp. nipponica | St | 0.933 | <0.001 |
| Phalaris arundinacea | So2 | Phalaris arundinacea | Ph | 0.719 | <0.001 |
| Potamogeton crispus | Se1 | Potamogeton crispus | Pc | 1.000 | <0.001 |
| Spirodela polyrhiza | So | 0.691 | <0.001 | ||
| Myriophyllum spicatum | My | 0.663 | <0.001 | ||
| Trapa japonica | Tj | 0.662 | <0.001 | ||
| Hydrilla verticillata | Hv | 0.302 | 0.030 | ||
| Ceratophyllum demersum | Cr | 0.298 | 0.030 | ||
| Hemistepta lyrata | Hl | 0.291 | 0.037 | ||
| Nymphoides peltata | Np | 0.279 | 0.031 | ||
| Typha angustifolia | Se2 | Typha angustifolia | Ta | 0.999 | <0.001 |
| Cyperus amuricus | Ca | 0.334 | 0.021 | ||
| Lindernia crustacea | Li | 0.333 | 0.015 | ||
| Lindernia micrantha | Lm | 0.333 | 0.014 | ||
| Zizania latifolia | Zl | 0.331 | 0.009 | ||
| Actinostemma lobatum | Al | 0.306 | 0.017 | ||
| Carex leiorhyncha | Cl | 0.303 | 0.023 | ||
| Juncus decipiens | Jd | 0.293 | 0.026 | ||
| Lindernia procumbens | Lp | 0.265 | 0.041 | ||
| Scirpus radicans | Se3 | Scirpus radicans | Sr | 0.714 | <0.001 |
| Leersia japonica | Lr | 0.435 | <0.001 | ||
| Paspalum distichum | Pi | 0.349 | 0.012 | ||
| Panicum dichotomiflorum | Pd | 0.281 | 0.026 | ||
| Phragmites australis | S4 | Phragmites australis | Pa | 0.797 | <0.001 |
| Humulus japonicus | Hj | 0.408 | 0.026 | ||
| Miscanthus sacchariflorus | S5 | Miscanthus sacchariflorus | Ms | 0.901 | <0.001 |
| Salix pierotii | S6 | Salix pierotii | Sp | 0.945 | <0.001 |
| Salix chaenomeloides | Sc | 0.485 | <0.001 | ||
| Achyranthes bidentata var. japonica | Ab | 0.349 | 0.016 | ||
| Glycine soja | Gs | 0.325 | 0.022 | ||
| Sicyos angulatus | Sg | 0.319 | 0.041 | ||
| Community | Properties of Dominant Species | No. of Species | Diversity | Height (m) | Coverage (%) | Tree Age (Years) | n | |
|---|---|---|---|---|---|---|---|---|
| Growth Form | Hydrological Type | |||||||
| So0 | - | - | - | - | - | - | - | 37 |
| So1a | Annual herb | Hygrophyte | 4.0 ± 2.2 | 1.17 ± 0.70 | 0.4 ± 0.2 | 51 ± 33 | 0.1 ± 0.3 | 50 |
| So1b | Perennial subtree | Hygrophyte | 3.9 ± 2.8 | 1.04 ± 0.67 | 3.8 ± 2.2 | 93 ± 13 | 5.1 ± 4.0 | 68 |
| So2 | Perennial herb | Hygrophyte | 2.7 ± 2.3 | 0.66 ± 0.71 | 0.8 ± 0.6 | 65 ± 30 | 0.3 ± 0.8 | 60 |
| Se1 | Perennial herb | Submergent | 3.0 ± 1.4 | 0.99 ± 0.45 | 0.4 ± 0.1 | 59 ± 29 | - | 11 |
| Se2 | Perennial herb | Emergent | 3.1 ± 1.4 | 0.96 ± 0.49 | 1.4 ± 0.5 | 54 ± 18 | - | 8 |
| Se3 | Perennial herb | Emergent | 3.5 ± 1.8 | 0.97 ± 0.59 | 0.4 ± 0.2 | 80 ± 21 | - | 26 |
| S4 | Perennial herb | Emergent | 3.3 ± 1.8 | 0.96 ± 0.52 | 1.4 ± 0.6 | 83 ± 25 | 0.1 ± 0.6 | 145 |
| S5 | Perennial herb | Hygrophyte | 3.2 ± 1.6 | 0.90 ± 0.50 | 1.6 ± 0.4 | 96 ± 10 | 0.5 ± 1.6 | 114 |
| S6 | Perennial tree | Hygrophyte | 7.2 ± 3.7 | 1.75 ± 0.52 | 7.2 ± 3.3 | 98 ± 07 | 12.6 ± 5.3 | 144 |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2025 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Lee, C.; Cho, K.-H. Successional Pathways of Riparian Vegetation Following Weir Gate Operations: Insights from the Geumgang River, South Korea. Water 2025, 17, 1006. https://doi.org/10.3390/w17071006
Lee C, Cho K-H. Successional Pathways of Riparian Vegetation Following Weir Gate Operations: Insights from the Geumgang River, South Korea. Water. 2025; 17(7):1006. https://doi.org/10.3390/w17071006
Chicago/Turabian StyleLee, Cheolho, and Kang-Hyun Cho. 2025. "Successional Pathways of Riparian Vegetation Following Weir Gate Operations: Insights from the Geumgang River, South Korea" Water 17, no. 7: 1006. https://doi.org/10.3390/w17071006
APA StyleLee, C., & Cho, K.-H. (2025). Successional Pathways of Riparian Vegetation Following Weir Gate Operations: Insights from the Geumgang River, South Korea. Water, 17(7), 1006. https://doi.org/10.3390/w17071006