
Driving Factors and Variability of Cyanobacterial Blooms in Qionghai Lake, Yunnan–Guizhou Plateau, China


{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
1. Introduction
2. Materials and Methods
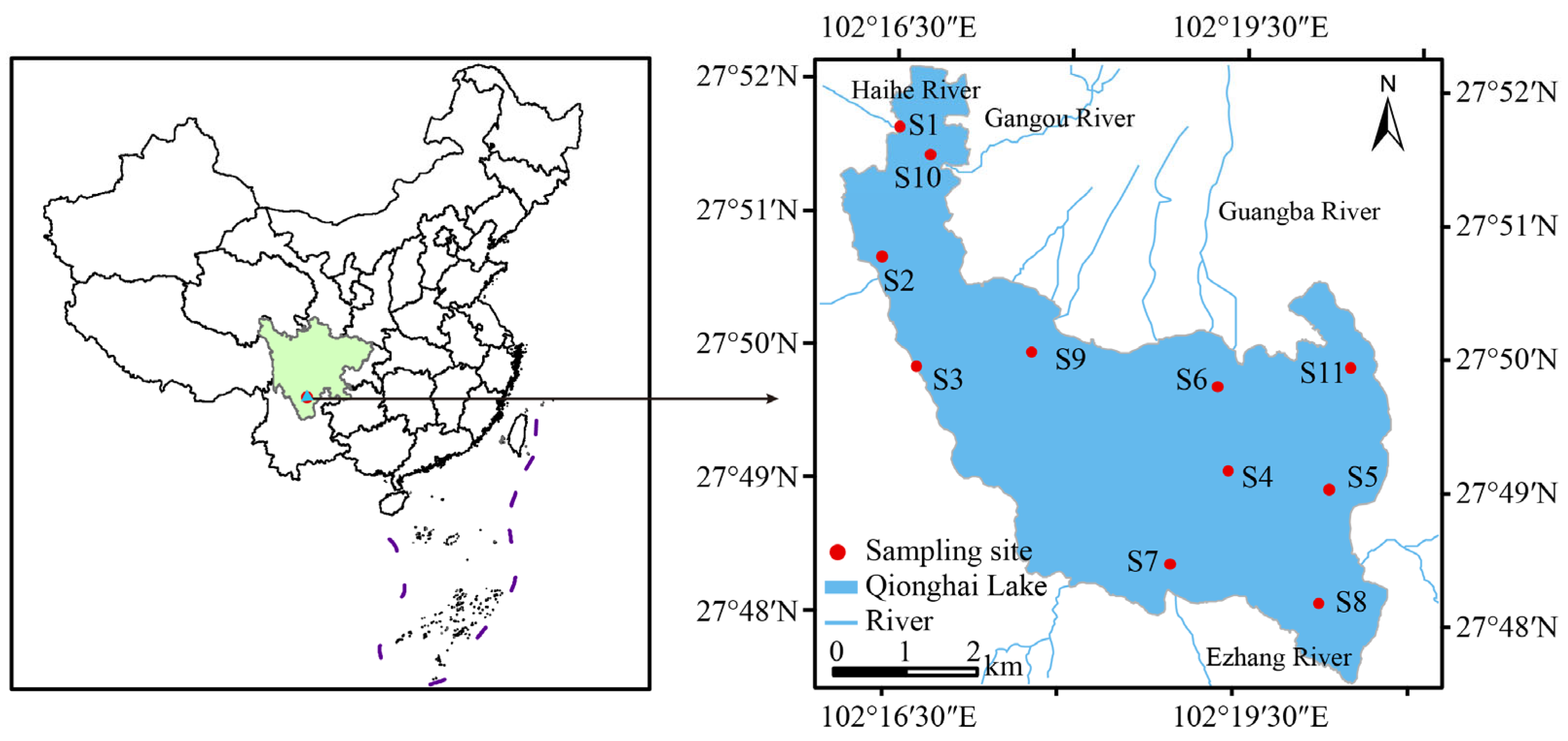
2.1. Study Area
2.2. Sampling Methods
2.3. Analytical Methods and Data Source
2.4. Statistical Analysis
3. Results and Discussion
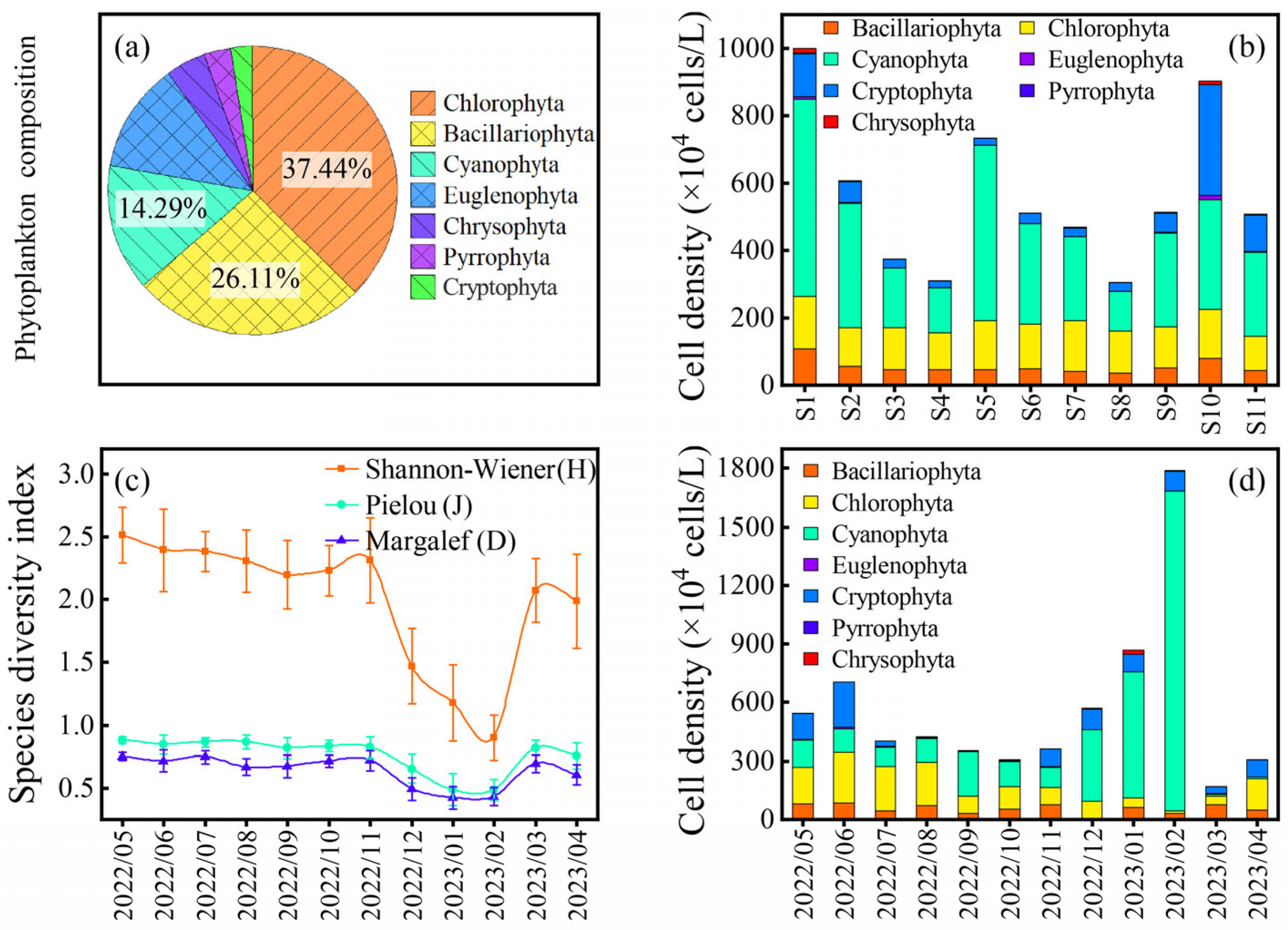
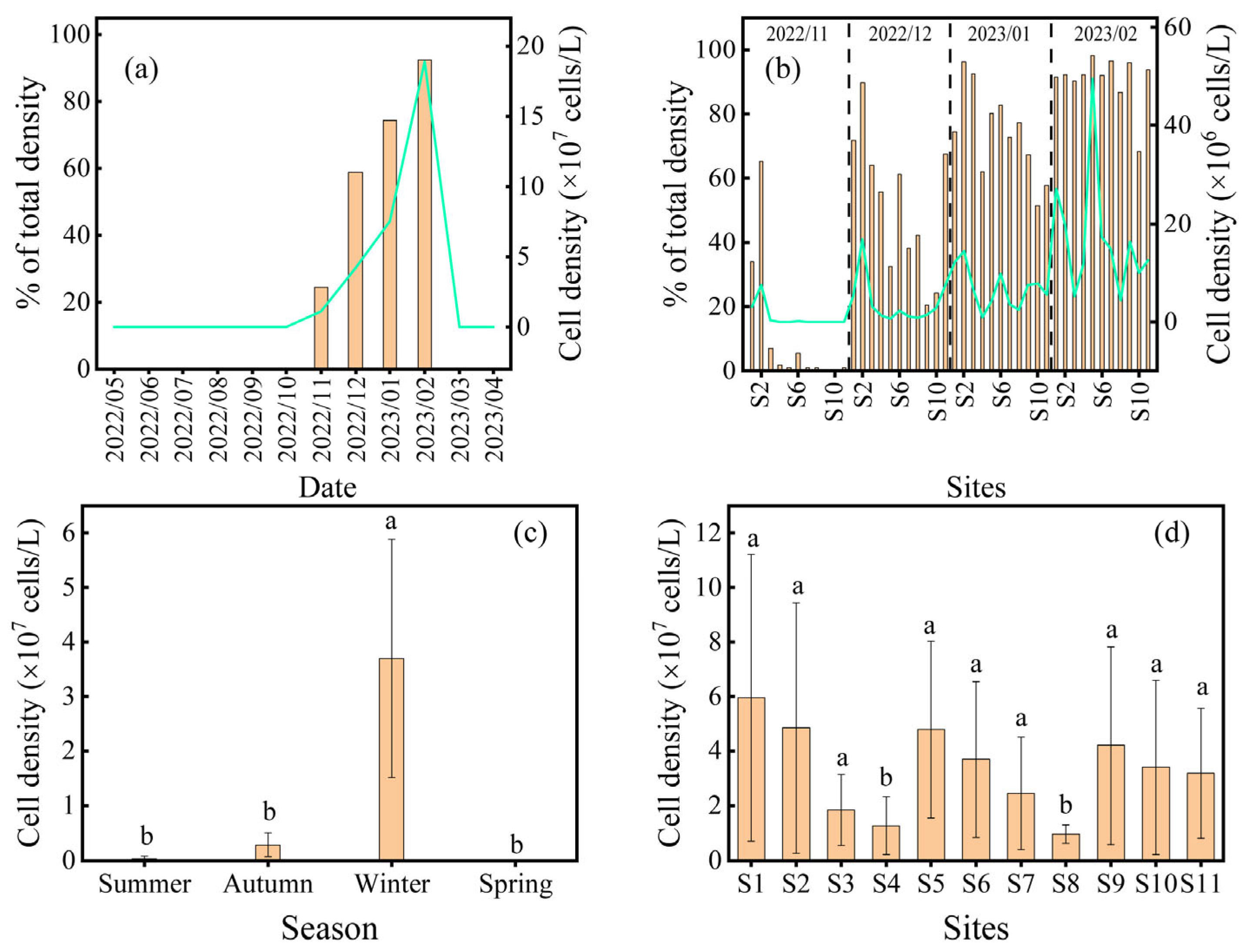
3.1. Changes in Phytoplankton
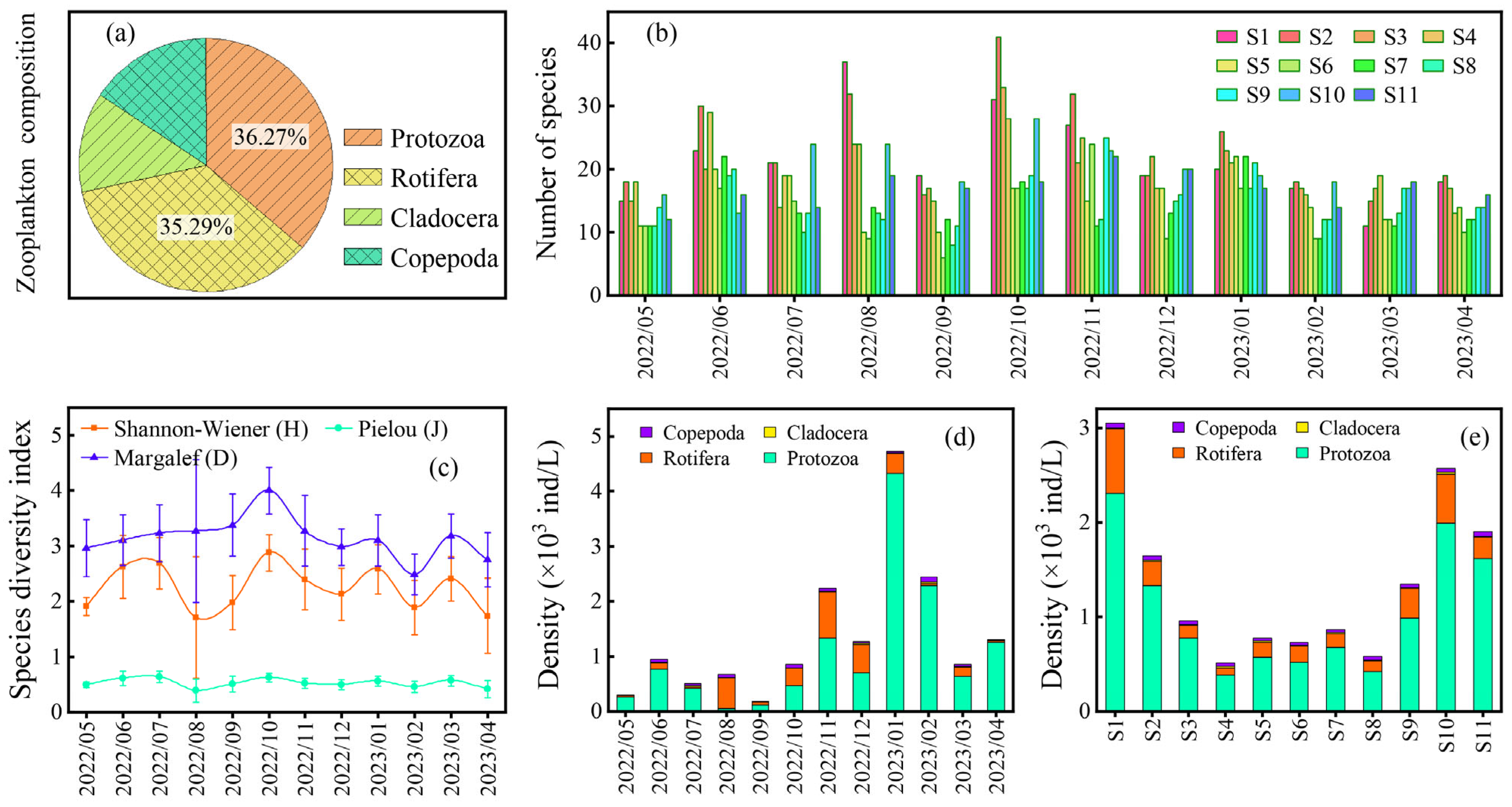
3.2. Effects of Zooplankton on Cyanobacterial Bloom
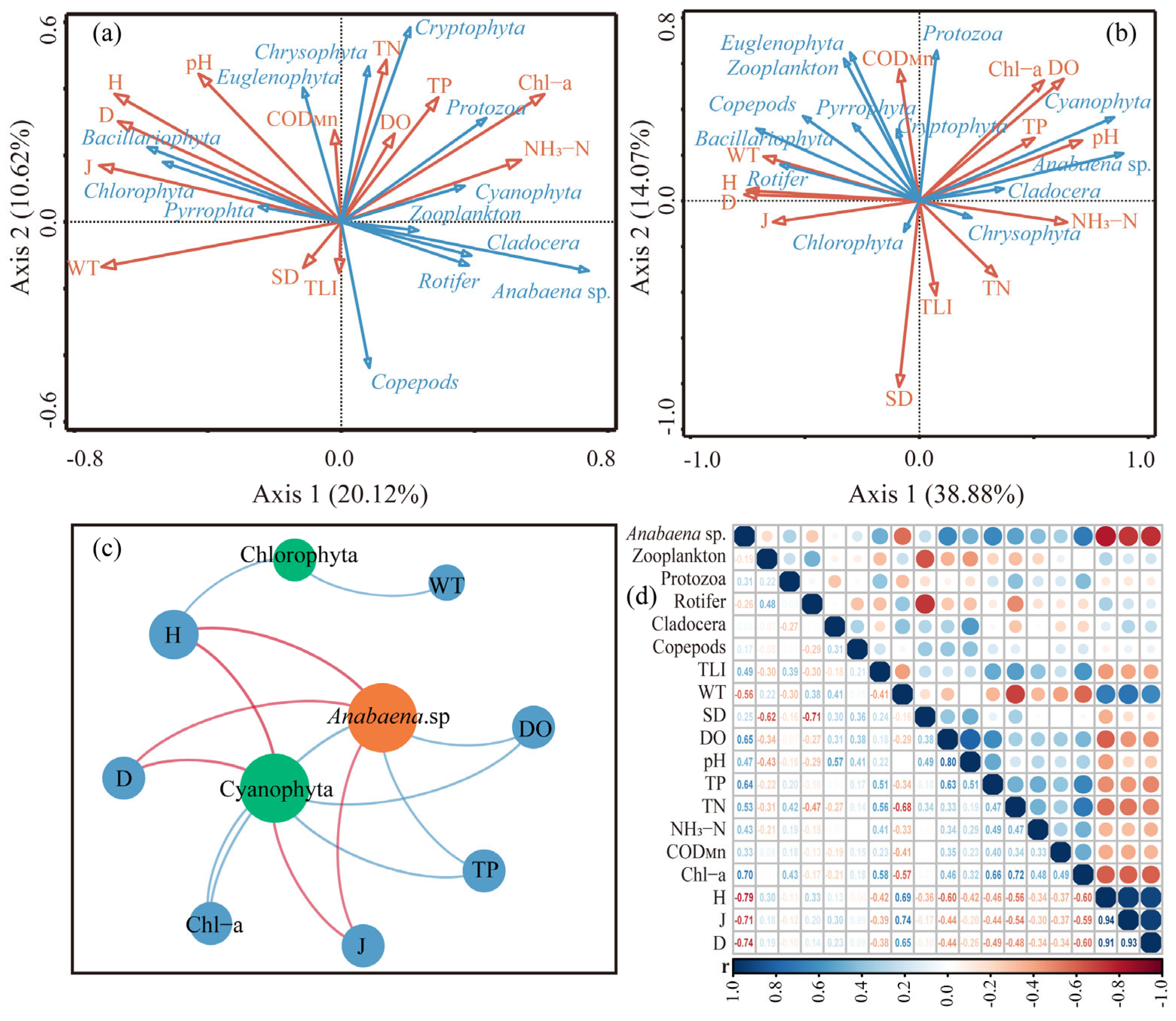
3.3. Effects of Biochemical Indicators on Cyanobacterial Bloom
3.3.1. Effects Analysis on Temporal Scale
3.3.2. Effects Analysis on Spatial Scale
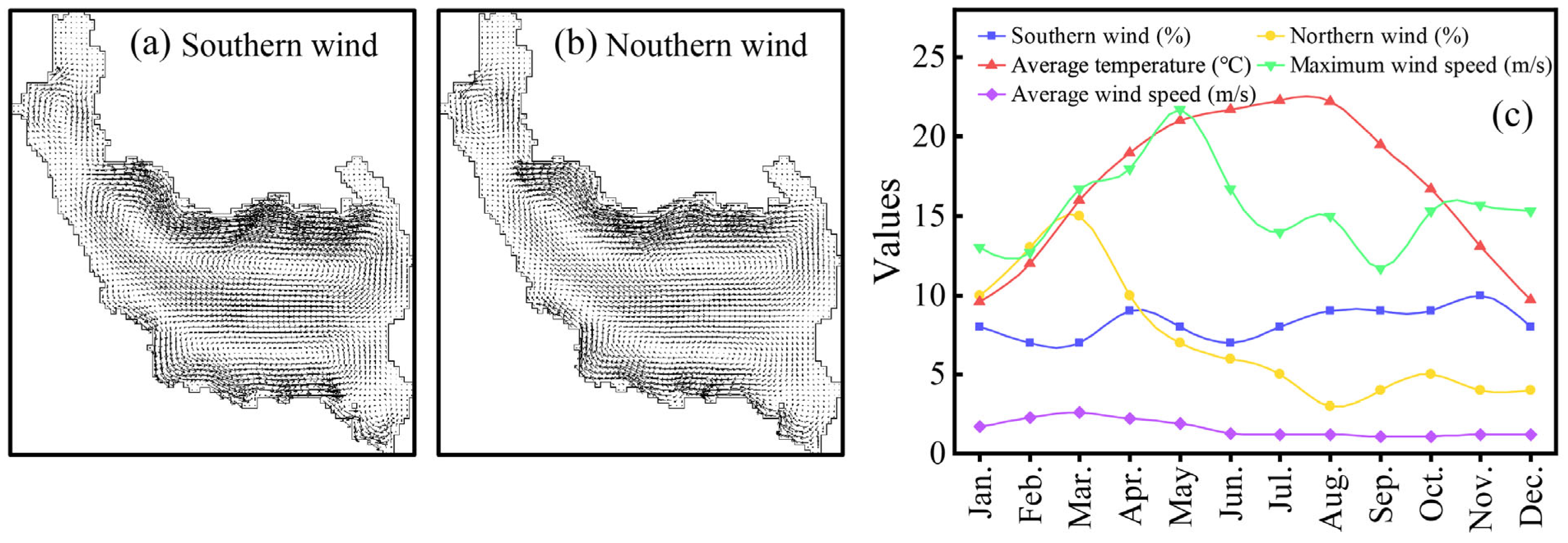
3.4. Ecological Mechanism of Cyanobacteria Bloom Occurrence in Qionghai Lake
4. Conclusions
Supplementary Materials
Author Contributions
Funding
Data Availability Statement
Conflicts of Interest
References
- Amorim, C.A.; Moura, A.d.N. Ecological impacts of freshwater algal blooms on water quality, plankton biodiversity, structure, and ecosystem functioning. Sci. Total Environ. 2021, 758, 143605. [Google Scholar] [CrossRef] [PubMed]
- Hou, X.; Feng, L.; Dai, Y.; Hu, C.; Gibson, L.; Tang, J.; Lee, Z.; Wang, Y.; Cai, X.; Liu, J.; et al. Global mapping reveals increase in lacustrine algal blooms over the past decade. Nat. Geosci. 2022, 15, 130–134. [Google Scholar] [CrossRef]
- Peng, J.; Chen, J.; Liu, S.; Liu, T.; Cao, M.; Nanding, N.; Zhuang, L.; Bao, A.; De Maeyer, P. Dynamics of algal blooms in typical low-latitude plateau lakes: Spatiotemporal patterns and driving factors. Environ. Pollut. 2024, 345, 123453. [Google Scholar] [CrossRef] [PubMed]
- Qin, B.; Zhang, Y.; Zhu, G.; Gao, G. Eutrophication control of large shallow lakes in China. Sci. Total Environ. 2023, 881, 163494. [Google Scholar] [CrossRef]
- Antunes, J.T.; Leão, P.N.; Vasconcelos, V.M. Cylindrospermopsis raciborskii: Review of the distribution, phylogeography, and ecophysiology of a global invasive species. Front. Microbiol. 2015, 6, 473. [Google Scholar] [CrossRef]
- Huisman, J.; Codd, G.A.; Paerl, H.W.; Ibelings, B.W.; Verspagen, J.M.H.; Visser, P.M. Cyanobacterial blooms. Nat. Rev. Microbiol. 2018, 16, 471–483. [Google Scholar] [CrossRef]
- Ran, J.; Xiang, R.; He, J.; Zheng, B. Spatiotemporal variation and driving factors of water quality in Yunnan-Guizhou plateau lakes, China. J. Contam. Hydrol. 2023, 254, 104141. [Google Scholar] [CrossRef]
- Wang, J.-H.; Yang, C.; He, L.-Q.-S.; Dao, G.-H.; Du, J.-S.; Han, Y.-P.; Wu, G.-X.; Wu, Q.-Y.; Hu, H.-Y. Meteorological factors and water quality changes of Plateau Lake Dianchi in China (1990–2015) and their joint influences on cyanobacterial blooms. Sci. Total Environ. 2019, 665, 406–418. [Google Scholar] [CrossRef]
- Cao, J.; Hou, Z.; Li, Z.; Chu, Z.; Yang, P.; Zheng, B. Succession of phytoplankton functional groups and their driving factors in a subtropical plateau lake. Sci. Total Environ. 2018, 631–632, 1127–1137. [Google Scholar] [CrossRef]
- Yong, C.; Wang, Y.Q. Qionghai Lake, Sichuan, China: Environmental degradation and the need for multidimensional management. Mt. Res. Dev. 2003, 23, 65–72. [Google Scholar] [CrossRef]
- Liu, Y.; Guo, H.C.; Zou, R.; Wang, L.J. Neural network modeling for regional hazard assessment of debris flow in Lake Qionghai Watershed, China. Environ. Geol. 2006, 49, 968–976. [Google Scholar] [CrossRef]
- Wei, X.; Chen, N. Development of debris flows in Guanba River and its effect on sediment deposition in Qionghai Lake of Sichuan. Acta Geogr. Sin. 2018, 3, 81–91. [Google Scholar]
- Wu, S.; Tian, C.; Li, B.; Wang, J.; Wang, Z. Ecological environment health assessment of lake water ecosystem system based on simulated annealing-projection pursuit: A case study of plateau lake. Sustain. Cities Soc. 2022, 86, 104131. [Google Scholar] [CrossRef]
- Ran, J.; Xiang, R.; Li, J.; Xiao, K.; Zheng, B. Spatiotemporal variations in the water quality of Qionghai Lake, Yunnan-Guizhou Plateau, China. Water 2022, 14, 2451. [Google Scholar] [CrossRef]
- Wang, G.; Wang, Y.; Wei, Z.; He, W.; Zhang, T.; Ma, X. Geochemical records of Qionghai Lake sediments in southwestern China linked to late Quaternary climate changes. Palaeogeogr. Palaeoclimatol. Palaeoecol. 2020, 560, 109902. [Google Scholar] [CrossRef]
- Hu, H.J.; Wei, Y.X. The Freshwater Algae of China-Systematics, Taxonomy and Ecology; Science Press: Beijing, China, 2006. [Google Scholar]
- Han, M.S. Freshwater Plankton Atlas; Agricultural Publishing House: Beijing, China, 1980. [Google Scholar]
- Institute of Zoology Chinese Academy of Sciences. Fauna Sinica: Freshwater Copepoda; Science Press: Beijing, China, 1979. [Google Scholar]
- Jiang, X.Z.; Du, N.S. Fauna Sinica: Freshwater Cladocera; Science Press: Beijing, China, 1979. [Google Scholar]
- Xiang, R.; Liu, T.; Chu, Z.; Wang, X.; Zheng, B.; Jia, H. Effects of dissolved organic matter derived from two herbs on the growth, physiology, and physico-chemical characteristics of four bloom-forming algae species. J. Environ. Manag. 2023, 336, 117559. [Google Scholar] [CrossRef]
- GB 3838-2002; Environmental Quality Standards for Surface Water. MEP (Ministry of Environmental Protection P.R. China): Beijing, China, 2002.
- Secchi, P.A. Relazione delle esperienze fatte a bordo della pontificia pirocorvetta Imacolata Concezione per determinare la trasparenza del mare. Mem. PA Secchi. Il Nuovo C. G. Fis. Chim. E Stor. Nat. 1864, 20, 205–237. [Google Scholar]
- Zhang, S. Eutrophication of the Three Gorges Reservoir after its first filling. Trop. Sub-Trop. Reserv. Limnol. China Theory Pract. 2012, 91, 293–309. [Google Scholar]
- Peng, Y.; Fan, M.; Song, J.; Cui, T.; Li, R. Assessment of plant species diversity based on hyperspectral indices at a fine scale. Sci. Rep. 2018, 8, 4776. [Google Scholar] [CrossRef]
- Wei, Y.; Ding, D.; Gu, T.; Jiang, T.; Qu, K.; Sun, J.; Cui, Z. Different responses of phytoplankton and zooplankton communities to current changing coastal environments. Environ. Res. 2022, 215, 114426. [Google Scholar] [CrossRef]
- He, J.; Wu, X.; Zhang, Y.; Zheng, B.; Meng, D.; Zhou, H.; Lu, L.; Deng, W.; Shao, Z.; Qin, Y. Management of water quality targets based on river-lake water quality response relationships for lake basins—A case study of Dianchi Lake. Environ. Res. 2020, 186, 109479. [Google Scholar] [CrossRef]
- Gabarron, M.; Faz, A.; Acosta, J.A. Use of multivariable and redundancy analysis to assess the behavior of metals and arsenic in urban soil and road dust affected by metallic mining as a base for risk assessment. J. Environ. Manag. 2018, 206, 192–201. [Google Scholar] [CrossRef]
- Li, L.; Li, G.; Li, G. Winter and spring succession of the phytoplankton community in Dianchi Lake under the Influences of environment change. J. Hydroecol. 2020, 41, 57–68. [Google Scholar]
- Triest, L.; Stiers, I.; Van Onsem, S. Biomanipulation as a nature-based solution to reduce cyanobacterial blooms. Aquat. Ecol. 2016, 50, 461–483. [Google Scholar] [CrossRef]
- Xu, M.Q.; Cao, H.; Xie, P.; Deng, D.G.; Feng, W.S.; Xu, H. The temporal and spatial distribution, composition and abundance of Protozoa in Chaohu Lake, China: Relationship with eutrophication. Eur. J. Protistol. 2005, 41, 183–192. [Google Scholar] [CrossRef]
- Xu, H.; Qin, B.; Paerl, H.W.; Peng, K.; Zhang, Q.; Zhu, G.; Zhang, Y. Environmental controls of harmful cyanobacterial blooms in Chinese inland waters. Harmful Algae 2021, 110, 102127. [Google Scholar] [CrossRef]
- Sarkar, S.K. Marine Algal Bloom: Characteristics, Causes and Climate Change Impacts; Springer: Singapore, 2018; pp. 53–109. [Google Scholar]
- Thomann, R.V.; Mueller, J.A. Principles of Surface Water Quality Modeling and Control; Harper & Row Publishers: New York, NY, USA, 1987. [Google Scholar]
- Jia, Z.; Chang, X.; Duan, T.; Wang, X.; Wei, T.; Li, Y. Water quality responses to rainfall and surrounding land uses in urban lakes. J. Environ. Manag. 2021, 298, 113514. [Google Scholar] [CrossRef]
- Dai, X.; Zhou, Y.; Ma, W.; Zhou, L. Influence of spatial variation in land-use patterns and topography on water quality of the rivers inflowing to Fuxian Lake, a large deep lake in the plateau of southwestern China. Ecol. Eng. 2017, 99, 417–428. [Google Scholar] [CrossRef]
- Wei, W.; Gao, Y.; Huang, J.; Gao, J. Exploring the effect of basin land degradation on lake and reservoir water quality in China. J. Clean. Prod. 2020, 268, 122249. [Google Scholar] [CrossRef]
- Walsby, A.E. Gas Vesicles. Microbiol. Rev. 1994, 58, 94–144. [Google Scholar] [CrossRef]
- Yang, Z.; Zhang, M.; Shi, X.; Kong, F.; Ma, R.; Yu, Y. Nutrient reduction magnifies the impact of extreme weather on cyanobacterial bloom formation in large shallow Lake Taihu (China). Water Res. 2016, 103, 302–310. [Google Scholar] [CrossRef]
- Zhang, M.; Zhang, Y.; Yang, Z.; Wei, L.; Yang, W.; Chen, C.; Kong, F. Spatial and seasonal shifts in bloom-forming cyanobacteria in Lake Chaohu: Patterns and driving factors. Phycol. Res. 2016, 64, 44–55. [Google Scholar] [CrossRef]
- Zeng, Y.; Chang, F.; Wen, X.; Duan, L.; Zhang, Y.; Liu, Q.; Zhang, H. Seasonal variation in the water quality and eutrophication of Lake Xingyun in southwestern China. Water 2022, 14, 3677. [Google Scholar] [CrossRef]
- Mazur, M.L.C.; Smith, B.; Bird, B.; McMillan, S.; Pyron, M.; Hauswald, C. Hydrologic connectivity and land cover affect floodplain lake water quality, fish abundance, and fish diversity in floodplain lakes of the Wabash-White River basin. River Res. Appl. 2022, 38, 160–172. [Google Scholar] [CrossRef]
- Glibert, P.M. Harmful algae at the complex nexus of eutrophication and climate change. Harmful Algae 2020, 91, 101583. [Google Scholar] [CrossRef]
- Wu, Y.; Li, L.; Zheng, L.; Dai, G.; Ma, H.; Shan, K.; Wu, H.; Zhou, Q.; Song, L. Patterns of succession between bloom-forming cyanobacteria Aphanizomenon flos-aquae and Microcystis and related environmental factors in large, shallow Dianchi Lake, China. Hydrobiologia 2016, 765, 1–13. [Google Scholar] [CrossRef]
- Zhang, Y.; Zhang, H.; Liu, Q.; Duan, L.; Zhou, Q. Total nitrogen and community turnover determine phosphorus use efficiency of phytoplankton along nutrient gradients in plateau lakes. J. Environ. Sci. 2023, 124, 699–711. [Google Scholar] [CrossRef]
- Li, D.; Chang, F.; Wen, X.; Duan, L.; Zhang, H. Seasonal variations in water quality and algal blooming in hypereutrophic Lake Qilu of southwestern China. Water 2022, 14, 2611. [Google Scholar] [CrossRef]
- Wu, C.; Jiang, C.; Ju, M.; Pan, Z.; Li, Z.; Sun, L.; Geng, H. Simulation and control of the cyanobacterial bloom biomass in a typical plateau lake based on the logistic growth model: A case study of Xingyun Lake. Ecol. Inform. 2024, 82, 102779. [Google Scholar] [CrossRef]
- Jin, Y.; Ren, S.; Wu, Y.; Zhang, X.; Chen, Z.; Xie, B. Microbial community structures and bacteria-Cylindrospermopsis raciborskii interactions in Yilong Lake. FEMS Microbiol. Ecol. 2024, 100, fiae048. [Google Scholar] [CrossRef]


Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2025 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Dong, Y.; Tian, Z.; Li, X.; Xu, D.; Zheng, B. Driving Factors and Variability of Cyanobacterial Blooms in Qionghai Lake, Yunnan–Guizhou Plateau, China. Water 2025, 17, 169. https://doi.org/10.3390/w17020169
Dong Y, Tian Z, Li X, Xu D, Zheng B. Driving Factors and Variability of Cyanobacterial Blooms in Qionghai Lake, Yunnan–Guizhou Plateau, China. Water. 2025; 17(2):169. https://doi.org/10.3390/w17020169
Chicago/Turabian StyleDong, Yanzhen, Zebin Tian, Xiaoyan Li, Dayong Xu, and Binghui Zheng. 2025. "Driving Factors and Variability of Cyanobacterial Blooms in Qionghai Lake, Yunnan–Guizhou Plateau, China" Water 17, no. 2: 169. https://doi.org/10.3390/w17020169
APA StyleDong, Y., Tian, Z., Li, X., Xu, D., & Zheng, B. (2025). Driving Factors and Variability of Cyanobacterial Blooms in Qionghai Lake, Yunnan–Guizhou Plateau, China. Water, 17(2), 169. https://doi.org/10.3390/w17020169