

Shallow Hydrostratigraphy Beneath Marsh Platforms: Insights from Electrical Resistivity Tomography


Abstract
1. Introduction
2. Materials and Methods
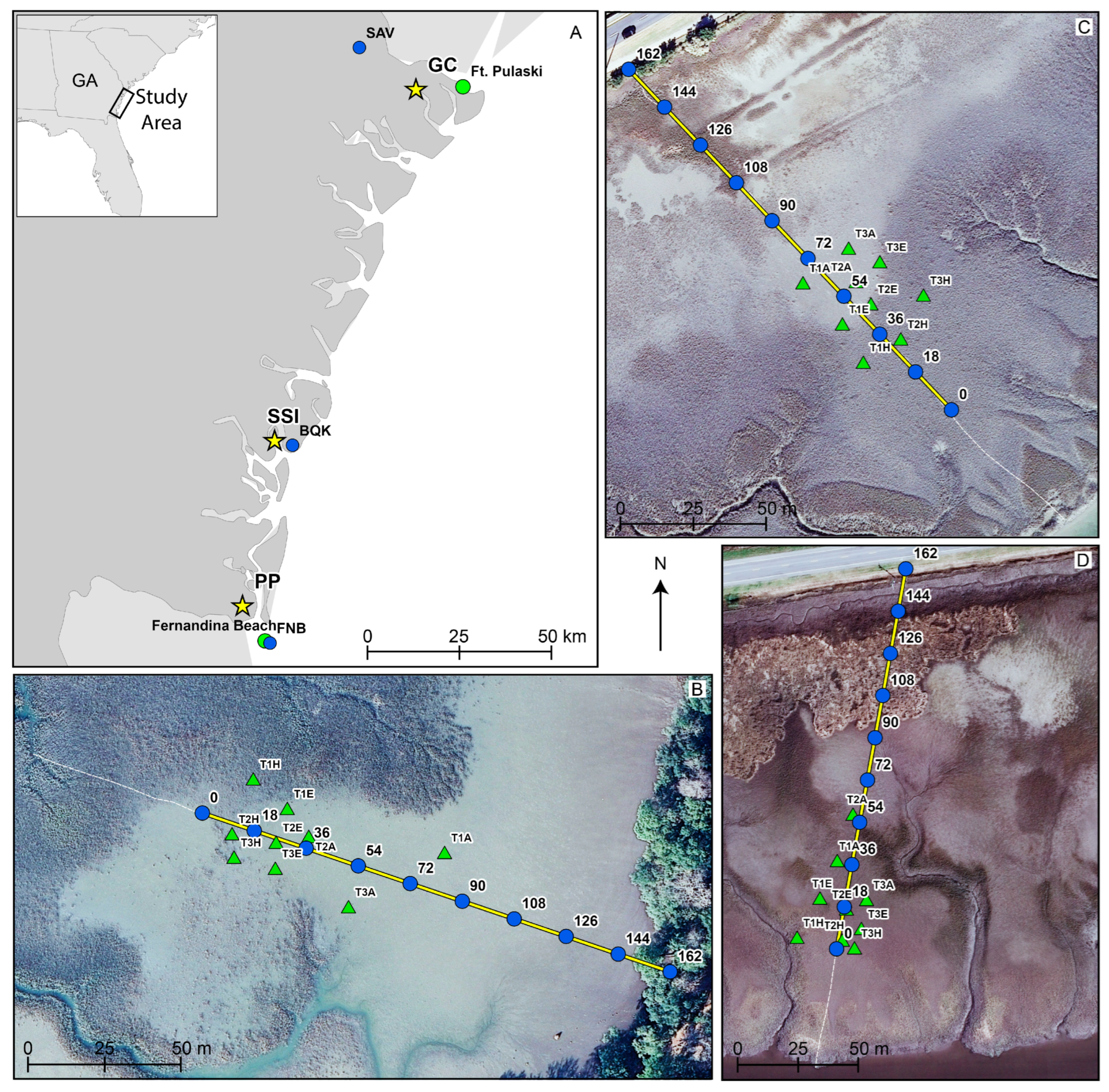
2.1. Study Sites
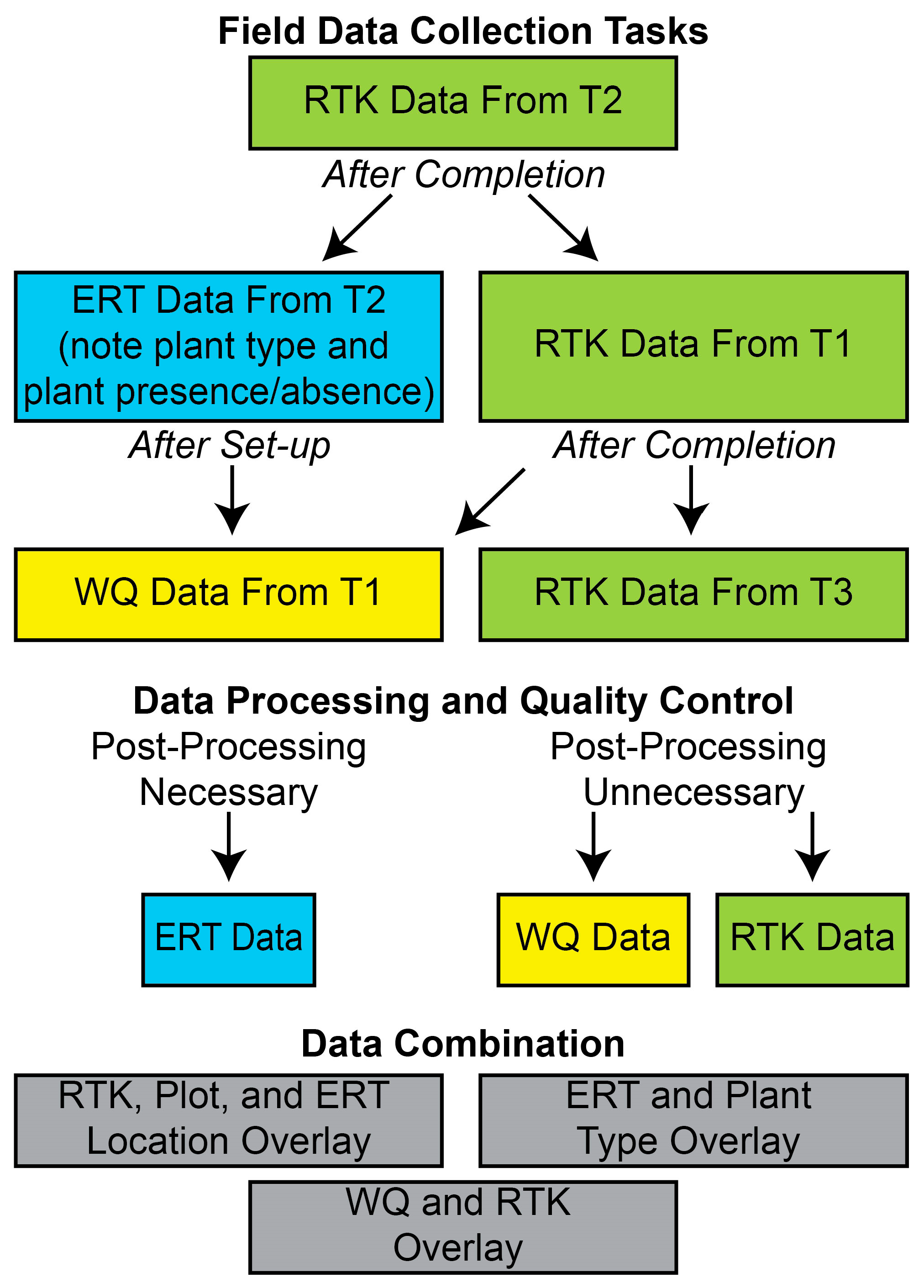
2.2. Data Collection and Processing
3. Results
4. Discussion
5. Conclusions
Author Contributions
Funding
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Costanza, R.; d’Arge, R.; de Groot, R.; Farber, S.; Grasso, M.; Hannon, B.; Limburg, K.; Naeem, S.; O’Neill, R.V.; Paruelo, J.; et al. The value of the world’s ecosystem services and natural capital. Nature 1997, 387, 253–260. [Google Scholar] [CrossRef]
- Mitsch, W.J.; Gosselink, J.G. Wetlands, 3rd ed.; John Wiley and Sons: Hoboken, NJ, USA, 2000. [Google Scholar]
- Elsey-Quirk, T.; Lynn, A.; Jacobs, M.D.; Diaz, R.; Cronin, J.; Wang, L.; Huang, H.; Justic, D. Vegetation dieback in the Mississippi River Delta triggered by acute drought and chronic relative sea-level rise. Nat. Commun. 2024, 15, 3518. [Google Scholar] [CrossRef] [PubMed]
- Hughes, A.L.; Wilson, A.M.; Morris, J.T. Hydrologic variability in a salt marsh: Assessing the links between drought and acute marsh dieback. Estuar. Coast. Shelf Sci. 2012, 111, 95–106. [Google Scholar] [CrossRef]
- Marsh, A.; Blum, L.K.; Christian, R.R.; Ramsey III, E.; Rangoonwala, A. Response and resilience of Spartina alterniflora to sudden dieback. J. Coast. Conserv. 2016, 20, 335–350. [Google Scholar] [CrossRef]
- Morton, R.A.; Tiling, G.; Ferina, N.F. Causes of hot-spot wetland loss in the Mississippi delta plain. Environ. Geosci. 2003, 10, 71–80. [Google Scholar] [CrossRef]
- Schepers, L.; Kirwan, M.; Guntenspergen, G.; Temmerman, S. Spatio-temporal development of vegetation die-off in a submerging coastal marsh. Limnol. Oceanogr. 2017, 62, 137–150. [Google Scholar] [CrossRef]
- Alber, M.; Swenson, E.M.; Adamowicz, S.C.; Mendelssohn, I.A. Salt marsh dieback: An overview of recent events in the US. Estuar. Coast. Shelf Sci. 2008, 80, 1–11. [Google Scholar] [CrossRef]
- Ogburn, M.B.; Alber, M. An investigation of salt marsh dieback in Georgia using field transplants. Estuaries Coasts 2006, 29, 54–62. [Google Scholar] [CrossRef]
- Day, J.W.; Christian, R.R.; Boesch, D.M.; Yáñez-Arancibia, A.; Morris, J.; Twilley, R.R.; Naylor, L.; Schaffner, L.; Stevenson, C. Consequences of Climate Change on the Ecogeomorphology of Coastal Wetlands. Estuaries Coasts 2008, 31, 477–491. [Google Scholar] [CrossRef]
- Baustian, J.J.; Mendelssohn, I.A.; Hester, M.W. Vegetation’s importance in regulating surface elevation in a coastal salt marsh facing elevated rates of sea level rise. Glob. Change Biol. 2012, 18, 3377–3382. [Google Scholar] [CrossRef]
- Coleman, D.J.; Kirwan, M.L. The effect of a small vegetation dieback event on salt marsh sediment transport. Earth Surf. Process. Landf. 2019, 44, 944–952. [Google Scholar] [CrossRef]
- Krest, J.M.; Moore, W.S.; Gardner, L.R.; Morris, J.T. Marsh nutrient export supplied by groundwater discharge: Evidence from radium measurements. Glob. Biogeochem. Cycles 2000, 14, 167–176. [Google Scholar] [CrossRef]
- Morris, J.T.; Sundareshwar, P.V.; Nietch, C.T.; Kjerfve, B.; Cahoon, D.R. Responses of coastal wetlands to rising sea level. Ecology 2002, 83, 2869–2877. [Google Scholar] [CrossRef]
- Materne, M.; Bush, T.; Houck, M. Plant Guide for Smooth Cordgrass (Spartina alterniflora). USDA-Natural Resources Conservation Service. 2022. Available online: https://www.nrcs.usda.gov/plantmaterials/njpmcpg13933.pdf (accessed on 3 January 2025).
- Eleuterius, L.N. Vegetative Morphology and Anatomy of the Salt Marsh Rush, Juncus roemerianus. Gulf Res. Rep. 1976, 5, 1–10. [Google Scholar] [CrossRef]
- Moore, W.S.; Blanton, J.O.; Joye, J. Estimates of flushing times, submarine groundwater discharge, and nutrient fluxes to Okatee Estuary, South Carolina. J. Geophys. Res. 2006, 111, C09006. [Google Scholar] [CrossRef]
- Swarzenski, P.W.; Izbicki, J.A. Coastal groundwater dynamics off Santa Barbara, California: Combining geochemical tracers, electromagnetic seepmeters, and electrical resistivity. Estuar. Coast. Shelf Sci. 2009, 83, 77–89. [Google Scholar] [CrossRef]
- Dimova, N.T.; Swarzenski, P.W.; Dulaiova, H.; Glenn, C.R. Utilizing multichannel electrical resistivity methods to examine the dynamics of the fresh water-seawater interface in two Hawaiian groundwater systems. J. Geophys. Res. 2012, 117, C02012. [Google Scholar] [CrossRef]
- Swarzenski, P.W.; Burnett, W.C.; Greenwood, W.J.; Herut, B.; Peterson, R.; Dimova, N.; Shalem, Y.; Yechieli, Y.; Weinstein, Y. Combined time-series resistivity and geochemical tracer techniques to examine submarine groundwater discharge at Dor Beach, Israel. Geophys. Res. Lett. 2006, 33, L24405. [Google Scholar] [CrossRef]
- U.S. Climate Data. Available online: https://www.usclimatedata.com/ (accessed on 13 November 2024).
- Wiegert, R.G.; Pomeroy, L.R.; Wiebe, W.J. Ecology of salt marshes: An introduction. In The Ecology of a Salt Marsh; Pomeroy, L.R., Wiegert, R.G., Eds.; Springer: New York, NY, USA, 1981; pp. 3–19. [Google Scholar]
- Tides and Currents (Datums for 8670870, Fort Pulaski GA). Available online: https://tidesandcurrents.noaa.gov/datums.html?datum=MLLW&units=1&epoch=0&id=8670870&name=Fort+Pulaski&state=GA (accessed on 13 November 2024).
- Tides and Currents (Datums for 8720030, Fernandina Beach FL). Available online: https://tidesandcurrents.noaa.gov/datums.html?datum=MLLW&units=1&epoch=0&id=8720030&name=Fernandina+Beach&state=FL (accessed on 13 November 2024).
- Tides and Currents (Relative Sea Level Trend Fort Pulaski). Available online: https://tidesandcurrents.noaa.gov/sltrends/sltrends_station.shtml?id=8670870 (accessed on 13 November 2024).
- Tides and Currents (Relative Sea Level Trend Fernandina Beach). Available online: https://tidesandcurrents.noaa.gov/sltrends/sltrends_station.shtml?id=8720030 (accessed on 13 November 2024).
- Dibble, M. Examination of Salt Marsh Dieback Development and Recovery Using Historical Aerial Photography and Climatic Conditions. Bachelor’s Thesis, Georgia Southern University, Statesboro, GA, USA, 2017. [Google Scholar]
- Trimble R8 GNSS Receiver User Guide. Available online: https://trl.trimble.com/docushare/dsweb/Get/Document-666215/R8-R6-5800_v400A_UserGuide.pdf (accessed on 23 July 2024).
- Rinaldi, V.; Guichon, M.; Ferrero, V.; Serrano, C.; Ponti, N. Resistivity Survey of the Subsurface Conditions in the Estuary of the Rio de la Plata. J. Geotech. Geoenviron. Eng. 2006, 132, 72–79. [Google Scholar] [CrossRef]
- Loke, M.H.; Chambers, J.E.; Rucker, D.F.; Kuras, O.; Wilkinson, P.B. Recent developments in the direct-current geoelectrical imaging method. J. Appl. Geophys. 2013, 95, 135–156. [Google Scholar] [CrossRef]
- Advanced Geosciences Incorporated Instruction Manual for EarthImager 2D Resistivity and IP Inversion Software. Available online: http://www.agiusa.com/ (accessed on 13 November 2024).
- Nguyen, F.; Garambois, S.; Jongmans, D.; Pirard, E.; Loke, M.H. Image processing of 2D resistivity data for imaging faults. J. Appl. Geophys. 2005, 57, 260–277. [Google Scholar] [CrossRef]
- Caputo, R.; Piscitelli, S.; Oliveto, A.; Rizzo, E.; Lapenna, V. The use of electrical resistivity tomographies in active tectonics: Examples from the Tyrnavos Basin, Greece. J. Geodyn. 2003, 36, 19–35. [Google Scholar] [CrossRef]
- Hoffmann, R.; Dietrich, P. An approach to determine equivalent solutions to the geoelectrical 2D inversion problem. J. Appl. Geophys. 2004, 56, 79–91. [Google Scholar] [CrossRef]
- Oldenburg, D.W.; Li, Y. Estimating depth of investigation in dc resistivity and IP surveys. Geophysics 1999, 64, 403–416. [Google Scholar] [CrossRef]
- Michaels, R.E.; Zieman, J.C. Fiddler crab (Uca spp.) burrows have little effect on surrounding sediment oxygen concentrations. J. Exp. Mar. Biol. Ecol. 2013, 448, 104–113. [Google Scholar] [CrossRef]
- Hemond, H.F.; Fifield, J.L. Subsurface flow in salt marsh peat: A model and field study. Limnol. Oceanogr. 1982, 27, 126–136. [Google Scholar] [CrossRef]
- Granse, D.; Titschack, J.; Ainouche, M.; Jensen, K.; Koop-Jackobsen, K. Subsurface aeration of tidal wetland soils: Root-system structure and aerenchyma connectivity in Spartina (Poaceae). Sci. Total Environ. 2022, 802, 149771. [Google Scholar] [CrossRef]
- Li, H.; Li, L.; Lockington, D. Aeration for plant root respiration in a tidal marsh. Water Resour. Res. 2005, 41, W06023. [Google Scholar] [CrossRef]
- Xin, P.; Jin, G.; Li, L.; Barry, D.A. Effects of crab burrows on pore water flows in salt marshes. Adv. Water Resour. 2009, 32, 439–449. [Google Scholar] [CrossRef]
- Silva Filho, A.M.; Silva, J.R.S.; Fernandes, G.M.; Morais, L.D.; Coimbra, A.P.; Calixto, W.P. Root system analysis and influence of moisture on soil electrical properties. Energies 2021, 14, 6951. [Google Scholar] [CrossRef]
- Archie, G.E. The Electrical Resistivity Log as an Aid in Determining Some Reservoir Characteristics. Trans. AIME 1942, 146, 54–62. [Google Scholar] [CrossRef]
- Papen, M.; Hanssens, D.; De Smedt, P.; Walraevens, K.; Hermans, T. Combining resistivity and frequency domain electromagnetic methods to investigate submarine groundwater discharge in the littoral zone. Hydrol. Earth Syst. Sci. 2020, 24, 3539–3555. [Google Scholar] [CrossRef]
- Koontz, E.L.; Parker, S.M.; Stearns, A.E.; Roberts, B.J.; Young, C.M.; Windham-Myers, L.; Oikawa, P.Y.; Megonigal, J.P.; Noyce, G.L.; Buskey, E.J.; et al. Controls on spatial variation in porewater methane concentrations across United States tidal wetlands. Sci. Total Environ. 2024, 957, 177290. [Google Scholar] [CrossRef] [PubMed]
- Harvey, J.W.; Germann, P.F.; Odum, W.E. Geomorphological control of subsurface hydrology in the creekbank zone of tidal marshes. Estuar. Coast. Shelf Sci. 1987, 25, 677–691. [Google Scholar] [CrossRef]
- Gardner, L.R.; Porter, D.E. Stratigraphy and geologic history of a southeastern salt marsh basin, North Inlet, South Carolina, USA. Wetl. Ecol. Manag. 2001, 9, 371–385. [Google Scholar] [CrossRef]
- Gardner, L.R. Role of stratigraphy in governing pore water seepage from salt marsh sediments. Water Resour. Res. 2007, 43, W07502. [Google Scholar] [CrossRef]
- Wilson, A.M.; Moore, W.S.; Joye, S.B.; Anderson, J.L.; Schutte, C.A. Storm-driven groundwater flow in a salt marsh. Water Resour. Res. 2011, 47, W02535. [Google Scholar] [CrossRef]
- Wilson, A.M.; Evans, T.; Moore, W.; Schutte, C.A.; Joye, S.B.; Hughes, A.H.; Anderson, J.L. Groundwater controls ecological zonation of salt marsh macrophytes. Ecology 2015, 96, 840–849. [Google Scholar] [CrossRef]
- Alexander, C.R.; Hodgson, J.Y.S.; Brandes, J.A. Sedimentary processes and products in a mesotidal salt marsh environment: Insights from Groves Creek, Georgia. Geo-Mar. Lett. 2017, 37, 345–359. [Google Scholar] [CrossRef]
- Lonard, R.I.; Judd, F.W.; Stalter, R. Biological flora of coastal dunes and wetlands: Borrichia frutescens (L.) DC. J. Coast. Res. 2015, 31, 749–757. [Google Scholar] [CrossRef]
- Pennings, S.C.; Grant, M.-B.; Bertness, M.D. Plant zonation in low-latitude salt marshes: Disentangling the roles of flooding, salinity and competition. J. Ecol. 2005, 93, 159–167. [Google Scholar] [CrossRef]
- Thibodeau, P.M.; Gardner, L.R.; Reeves, H.W. The role of groundwater flow in controlling the spatial distribution of soil salinity and rooted macrophytes in a southeastern salt marsh, USA. Mangroves Salt Marshes 1998, 2, 1–13. [Google Scholar] [CrossRef]
- Gardner, L.R.; Reeves, H.W. Seasonal patterns in the soil water balance of a Spartina marsh site at North Inlet, South Carolina, USA. Wetlands 2002, 22, 467–477. [Google Scholar] [CrossRef]
- Montalvo, M.S.; Grande, E.; Graswell, A.E.; Visser, A.; Arora, B.; Seybold, E.C.; Tatariw, C.; Haskiins, J.C.; Endris, C.A.; Gerbl, F.; et al. A Fresh Take: Seasonal Changes in Terrestrial Freshwater Inputs Impact Salt Marsh Hydrology and Vegetation Dynamics. Estuaries Coasts 2024, 47, 2389–2405. [Google Scholar] [CrossRef]
- Wilson, A.M.; Morris, J.T. The influence of tidal forcing on groundwater flow and nutrient exchange in a salt marsh-dominated estuary. Biogeochemistry 2012, 108, 27–38. [Google Scholar] [CrossRef]
- Wilson, A.M.; Gardner, L.R. Tidally driven groundwater flow and solute exchange in a marsh: Numerical simulations. Water Resour. Res. 2006, 42, W01405. [Google Scholar] [CrossRef]
- Xin, P.; Gibbes, B.; Ling, L.; Song, Z.; Lockington, D. Soil saturation index of salt marshes subjected to spring-neap tides: A new variable for describing marsh soil aeration condition. Hydrol. Process. 2010, 24, 2564–2577. [Google Scholar] [CrossRef]
- Wiegert, R.G.; Freeman, B.J. Tidal Salt Marshes of the Southeast Atlantic Coast: A Community Profile; U.S. Department of the Interior, Fish and Wildlife Service: Washington, DC, USA, 1990. [CrossRef]
- McKee, K.L.; Patrick, W.H., Jr. The relationship of smooth cordgrass (Spartina alterniflora) to tidal datums: A review. Estuaries 1988, 11, 143–151. [Google Scholar] [CrossRef]
- Hladik, C.M.; Alber, M.; Schalles, J.F. Salt marsh elevation and habitat mapping using hyperspectral and LIDAR data. Remote Sens. Environ. 2013, 139, 318–330. [Google Scholar] [CrossRef]
- Hladik, C.; Alber, M. Accuracy assessment and correction of a LIDAR-derived salt marsh digital elevation model. Remote Sens. Environ. 2012, 121, 224–235. [Google Scholar] [CrossRef]
- Gardner, L.R. Role of geomorphic and hydraulic parameters in governing pore water seepage from salt marsh sediments. Water Resour. Res. 2005, 41, W07010. [Google Scholar] [CrossRef]
- Schultz, G.; Ruppel, C. Constraints on hydraulic parameters and implications for groundwater flux across the upland-estuary interface. J. Hydrol. 2002, 260, 255–269. [Google Scholar] [CrossRef]
- Moffett, K.B.; Gorelick, S.M. Relating salt marsh pore water geochemistry patterns to vegetation zones and hydrologic influences. Water Resour. Res. 2016, 52, 1729–1745. [Google Scholar] [CrossRef]



{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Parameter | Grays Creek (GC) | St. Simons Island (SSI) | Point Peter (PP) |
|---|---|---|---|
| Minimum Elevation (m) | 0.03 | −0.50 | −0.48 |
| Maximum Elevation (m) | 0.92 | 1.79 | 0.98 |
| Elevation Change (m) | 0.88 | 2.28 | 1.45 |
| Median Elevation (m) | 0.72 | 0.70 | 0.58 |
| Mean Elevation (m) | 0.70 | 0.74 | 0.55 |
| Standard Deviation (m) | 0.18 | 0.36 | 0.28 |
| Transect Length (m) | 221 | 216 | 212 |
| Electrode 0 Distance to Creek (m) | 56 | 51 | 47 |
| Sample Date | Precipitation (cm) | Tidal Amplitude (m) |
|---|---|---|
| Gray’s Creek (GC) | ||
| 10/09/2015 | 1.2 | 2.3 |
| 03/04/2016 | 1.7 | 1.8 |
| St. Simons Island (SSI) | ||
| 10/11/2015 | 0.4 | 1.9 |
| 03/06/2016 | 1.0 | 1.7 |
| Point Peter (PP) | ||
| 10/10/2015 | 3.0 | 1.7 |
| 03/05/2016 | 0.5 | 1.5 |
| Sample Date | Depth | Salinity | SPC | Depth | Salinity | SPC | Depth | Salinity | SPC |
|---|---|---|---|---|---|---|---|---|---|
| Grays Creek | GC T1A | GC T1E | GC T1H | ||||||
| 11/14/2014 | 40 | 37.5 | 56,329 | 30 | 36.3 | 54,050 | 30 | 35.8 | 54,024 |
| 03/20/2015 | 90 | 29.0 | 44,801 | 90 | 26.8 | 41,650 | 90 | 30.0 | 46,206 |
| 07/31/2015 | 90 | 24.3 | 38,580 | 90 | 23.8 | 37,590 | 90 | 22.2 | 35,390 |
| 10/09/2015 | - | - | - | 90 | 26.9 | 33,854 | 90 | 23.7 | 32,063 |
| 03/04/2016 | 90 | 25.2 | 39,379 | 90 | 23.1 | 36,438 | 90 | 22.8 | 36,118 |
| 07/22/2016 | 90 | 19.8 | 31,766 | 90 | 19.8 | 31,820 | 90 | 18.5 | 29,880 |
| St. Simons | SSI T1A | SSI T1E | SSI T1H | ||||||
| 11/01/2014 | 70 | 35.6 | 53,867 | 50 | 33.7 | 51,160 | 30 | 32.6 | 49,761 |
| 03/21/2015 | 90 | 38.3 | 55,286 | 60 | 34.4 | 51,829 | 90 | 36.6 | 52,648 |
| 07/25/2015 | 90 | 35.2 | 53,796 | 90 | 27.5 | 42,830 | 90 | 31.3 | 48,200 |
| 10/11/2015 | 90 | 36.0 | 48,002 | - | - | - | - | - | - |
| 03/06/2016 | 90 | 35.9 | 54,289 | - | - | - | - | - | - |
| 07/24/2016 | 90 | 34.1 | 51,882 | 60 | 30.9 | 47,596 | 90 | 28.6 | 44,410 |
| Point Peter | PP T1A | PP T1E | PP T1H | ||||||
| 11/02/2014 | 40 | 20.3 | 32,409 | 50 | 15.5 | 25,282 | 50 | 17.0 | 27,629 |
| 03/28/2015 | 90 | 26.9 | 42,665 | 90 | 32.2 | 54,160 | 90 | 22.5 | 35,562 |
| 07/26/2015 | 90 | 25.0 | 39,412 | 90 | 22.8 | 36,084 | 90 | 18.4 | 24,773 |
| 10/10/2015 | - | - | - | 90 | 22.8 | 38,115 | 90 | 23.0 | - |
| 03/05/2016 | - | - | - | 90 | 24.6 | 38,710 | 90 | 22.6 | 35,721 |
| 07/23/2016 | - | - | - | 90 | 21.6 | 34,300 | 90 | 17.3 | 28,308 |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2025 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Kelly, J.L.; Hladik, C.M. Shallow Hydrostratigraphy Beneath Marsh Platforms: Insights from Electrical Resistivity Tomography. Water 2025, 17, 144. https://doi.org/10.3390/w17020144
Kelly JL, Hladik CM. Shallow Hydrostratigraphy Beneath Marsh Platforms: Insights from Electrical Resistivity Tomography. Water. 2025; 17(2):144. https://doi.org/10.3390/w17020144
Chicago/Turabian StyleKelly, Jacque L., and Christine M. Hladik. 2025. "Shallow Hydrostratigraphy Beneath Marsh Platforms: Insights from Electrical Resistivity Tomography" Water 17, no. 2: 144. https://doi.org/10.3390/w17020144
APA StyleKelly, J. L., & Hladik, C. M. (2025). Shallow Hydrostratigraphy Beneath Marsh Platforms: Insights from Electrical Resistivity Tomography. Water, 17(2), 144. https://doi.org/10.3390/w17020144