

A Robust Oxysalt-Tolerant Bacterium Marinobacter sp. for Simultaneous Nitrification and Denitrification of Hypersaline Wastewater


{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
1. Introduction
2. Materials and Methods
2.1. Isolation and Identification
2.2. Response Surface Methodology Optimization
2.3. Assessment of Salt Tolerance
2.4. Effects of Different Oxysalts on Bacterial Growth and Nitrogen Removal
2.5. SND Capability at 11% Na2SO4
2.6. Enzyme Activity and Functional Genes
2.7. Analytical Methods
3. Results and Discussion
3.1. Identifying the Isolated Strain Y2
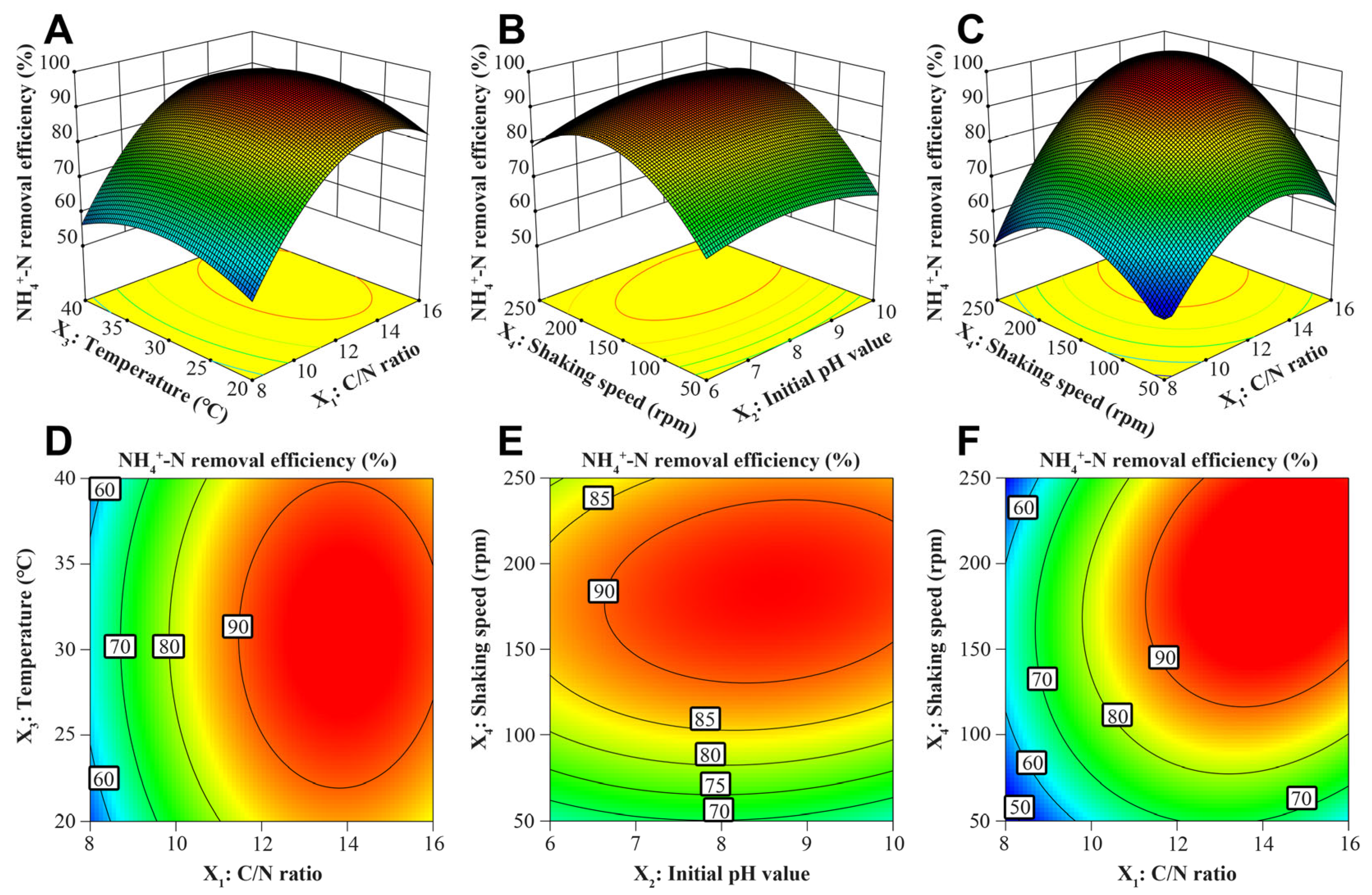
3.2. Optimization of the SND at High Salinity
3.3. Oxysalt-Tolerant Capabilities
3.4. SND Performance Using Single Nitrogen Sources
3.5. SND Performance Using Mixed Nitrogen Sources
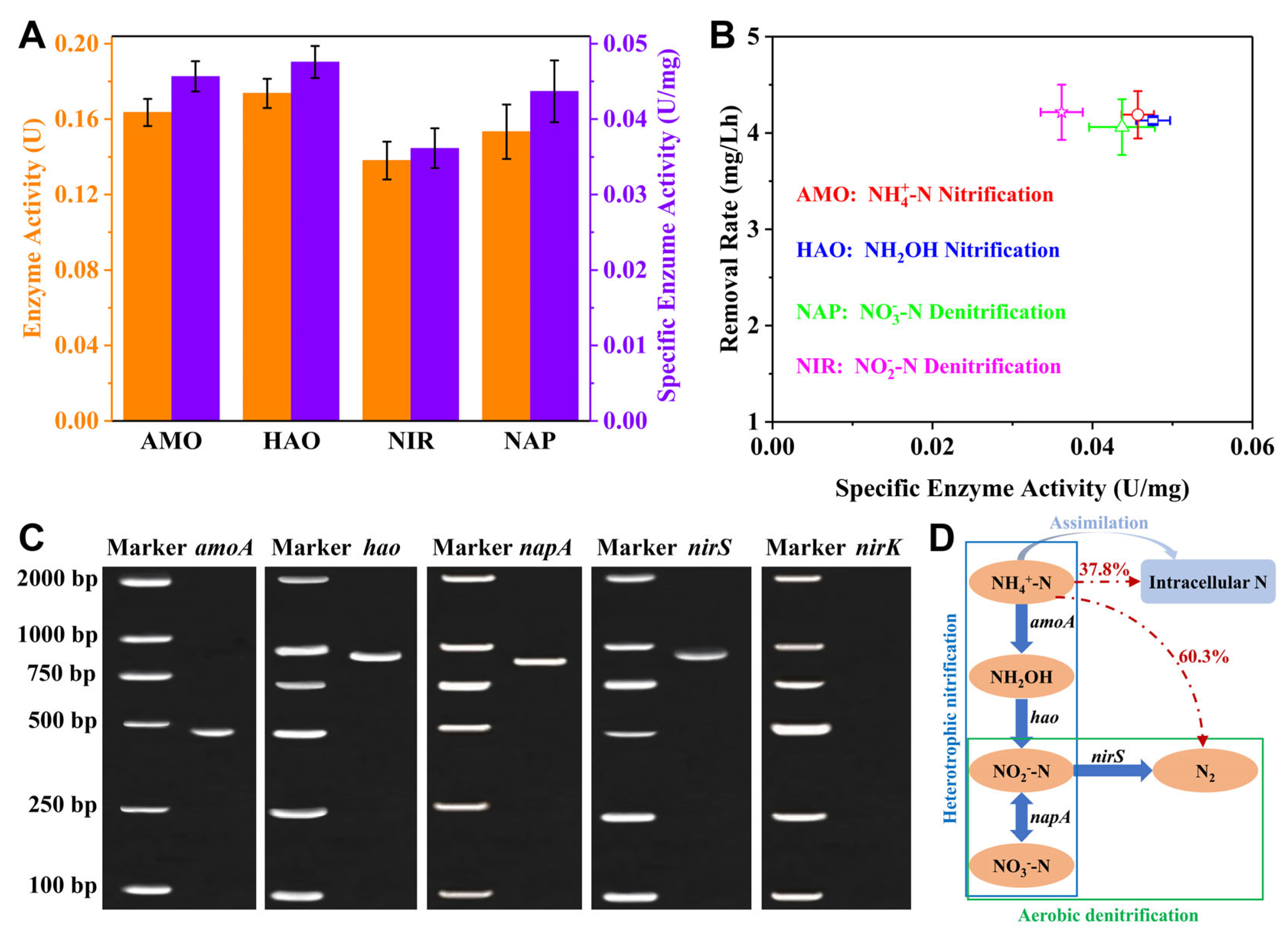
3.6. Nitrogen Removal Enzymes and Functional Genes for the SND
4. Conclusions
Supplementary Materials
Author Contributions
Funding
Data Availability Statement
Conflicts of Interest
References
- Chen, B.; Qaisar, M.; Xiao, J.; Li, W.; Li, J.; Cai, J. Combined acute effect of salinity and substrate concentration on simultaneous sulfide and nitrite removal process. Sep. Purif. Technol. 2023, 305, 122544. [Google Scholar] [CrossRef]
- Shen, N.; Guo, H.; Yao, T.; Xu, L.; Gao, Y.; Yang, P. Treatment of PickleWastewater under Varying Salinity Conditions within the Sequencing Batch Biofilm Reactor System. Water 2024, 16, 1312. [Google Scholar] [CrossRef]
- Liu, Y.; Yin, P.; Zhou, J.; Ma, Y.; Lai, X.; Lin, J.; Peng, H.; Shu, H.; Huang, W. Removal of Nitrogen and Phosphorus by a Novel Salt-Tolerant Strain Pseudomonas sediminis D4. Water 2025, 17, 502. [Google Scholar] [CrossRef]
- Zhang, Q.; Qin, S.; Yuan, C.; Fang, D.; Chen, W. A novel biofilm reactor startup with HN-AD bacteria inoculation for high ammonia-salt wastewater treatment: Performance, microbial characteristics and salt-tolerant mechanism. J. Water Process Eng. 2024, 59, 105032. [Google Scholar] [CrossRef]
- Li, P.; Wang, Y.; Liu, Y.; Wang, S.; Peng, Y. The Effect of Salinity on N2O Emissions during Domestic Wastewater Partial Nitrification Treatment in a Sequencing Batch Reactor. Water 2023, 15, 3502. [Google Scholar] [CrossRef]
- Lin, L.; Pratt, S.; Crick, O.; Xia, J.; Duan, H.; Ye, L. Salinity effect on freshwater Anammox bacteria: Ionic stress and ion composition. Water Res. 2021, 188, 116432. [Google Scholar] [CrossRef]
- Ng, K.K.; Shi, X.; Ng, H.Y. Evaluation of system performance and microbial communities of a bioaugmented anaerobic membrane bioreactor treating pharmaceutical wastewater. Water Res. 2015, 81, 311–324. [Google Scholar] [CrossRef] [PubMed]
- Ng, K.K.; Shi, X.; Ong, S.L.; Lin, C.-F.; Ng, H.Y. An innovative of aerobic bio-entrapped salt marsh sediment membrane reactor for the treatment of high-saline pharmaceutical wastewater. Chem. Eng. J. 2016, 295, 317–325. [Google Scholar] [CrossRef]
- Srivastava, A.; Parida, V.K.; Majumder, A.; Gupta, B.; Gupta, A.K. Treatment of saline wastewater using physicochemical, biological, and hybrid processes: Insights into inhibition mechanisms, treatment efficiencies and performance enhancement. J. Environ. Chem. Eng. 2021, 9, 105775. [Google Scholar] [CrossRef]
- Wang, J.; Zhou, J.; Wang, Y.; Wen, Y.; He, L.; He, Q. Efficient nitrogen removal in a modified sequencing batch biofilm reactor treating hypersaline mustard tuber wastewater: The potential multiple pathways and key microorganisms. Water Res. 2020, 177, 115734. [Google Scholar] [CrossRef]
- Han, F.; Zhao, C.; Zhang, W.; Jiao, T.; Zhang, Z.; Zhou, W. Responses of halophilic microbial communities to changes in salt composition: Comparison between autotrophic nitrification and heterotrophic ammonia assimilation biosystems. Bioresour. Technol. 2023, 386, 129500. [Google Scholar] [CrossRef] [PubMed]
- Li, Q.; He, Y.; Wang, B.; Weng, N.; Zhang, L.; Wang, K.; Tian, F.; Lyu, M.; Wang, S. Heterotrophic Nitrification–Aerobic Denitrification by Bacillus sp. L2: Mechanism of Denitrification and Strain Immobilization. Water 2024, 16, 416. [Google Scholar] [CrossRef]
- Wang, M.; He, J.; Zhang, J. Evaluating the performance and microbial community of aerobic granular sludge under different salinities. J. Water Process Eng. 2024, 58, 104891. [Google Scholar] [CrossRef]
- Cui, Y.; Cui, Y.-W.; Huang, J.-L. A novel halophilic Exiguobacterium mexicanum strain removes nitrogen from saline wastewater via heterotrophic nitrification and aerobic denitrification. Bioresour. Technol. 2021, 333, 125189. [Google Scholar] [CrossRef]
- Hu, J.; Yan, J.; Wu, L.; Bao, Y.; Yu, D.; Li, J. Simultaneous nitrification and denitrification of hypersaline wastewater by a robust bacterium Halomonas salifodinae from a repeated-batch acclimation. Bioresour. Technol. 2021, 341, 125818. [Google Scholar] [CrossRef]
- Hu, J.; Yan, J.; Wu, L.; Bao, Y.; Yu, D.; Li, J. Insight into halotolerance of a robust heterotrophic nitrifying and aerobic denitrifying bacterium Halomonas salifodinae. Bioresour. Technol. 2022, 351, 126925. [Google Scholar] [CrossRef]
- Blázquez, E.; Baeza, J.A.; Gabriel, D.; Guisasola, A. Treatment of real flue gas desulfurization wastewater in an autotrophic biocathode in view of elemental sulfur recovery: Microbial communities involved. Sci. Total Environ. 2019, 657, 945–952. [Google Scholar] [CrossRef]
- Han, Y.-L.; Zhang, X.-Z.; Liu, H.-B.; Rittmann, B.E.; Zhao, H.-P. Novel Sulfate Reduction Coupled to Simultaneous Nitrification and Autotrophic Denitrification Process for Removing Nitrogen and Organics from Saline Wastewater. Environ. Sci. Technol. 2023, 57, 10733–10744. [Google Scholar] [CrossRef]
- Mukhopadhyay, R.; Adhikari, T.; Sarkar, B.; Barman, A.; Paul, R.; Patra, A.K.; Sharma, P.C.; Kumar, P. Fe-exchanged nano-bentonite outperforms Fe3O4 nanoparticles in removing nitrate and bicarbonate from wastewater. J. Hazard. Mater. 2019, 376, 141–152. [Google Scholar] [CrossRef]
- Xu, Z.; Wang, J.; Qiu, J.; Cao, H.; Xie, Y. Unexpectedly Enhanced Organics Removal in Persulfate Oxidation with High Concentration of Sulfate: The Origin and the Selectivity. Environ. Sci. Technol. 2023, 57, 14442–14451. [Google Scholar] [CrossRef]
- Cheng, Q.; Xu, L.; Cheng, F.; Pan, G.; Zhou, Q. Bicarbonate-rich wastewater as a carbon fertilizer for culture of Dictyosphaerium sp. of a giant pyrenoid. J. Clean. Prod. 2018, 202, 439–443. [Google Scholar]
- Glass, C.; Silverstein, J. Denitrification of high-nitrate, high-salinity wastewater. Water Res. 1999, 33, 223–229. [Google Scholar] [CrossRef]
- Wang, R.; Zheng, P.; Ding, A.-Q.; Zhang, M.; Ghulam, A.; Yang, C.; Zhao, H.-P. Effects of inorganic salts on denitrifying granular sludge: The acute toxicity and working mechanisms. Bioresour. Technol. 2016, 204, 65–70. [Google Scholar] [CrossRef]
- Ji, B.; Zhang, H.; Zhou, L.; Yang, J.; Zhang, K.; Yuan, X.; Ma, J.; Qian, Y. Effect of the rapid increase of salinity on anoxic-oxic biofilm reactor for treatment of high-salt and high-ammonia-nitrogen wastewater. Bioresour. Technol. 2021, 337, 125363. [Google Scholar] [CrossRef]
- Shi, X.; Li, J.; Wang, X.; Zhang, X.; Tang, L. Effect of the gradual increase of Na2SO4 on performance and microbial diversity of aerobic granular sludge. J. Environ. Manag. 2021, 292, 112696. [Google Scholar] [CrossRef]
- Li, B.; Yun, Y.; Liu, G.; Li, C.; Li, X.; Hilal, M.; Yang, W.; Wang, M. Direct contact membrane distillation with softening Pre-treatment for effective reclaiming flue gas desulfurization wastewater. Sep. Purif. Technol. 2021, 277, 119637. [Google Scholar] [CrossRef]
- Chen, A.-L.; Su, X.; Xing, Z.-L.; Xu, F.-Q.; Chen, S.-J.; Xiang, J.-X.; Li, J.; Liu, H.; Zhao, T.-T. Effect mechanism of individual and combined salinity on the nitrogen removal yield of heterotrophic nitrification-aerobic denitrification bacteria. Environ. Res. 2022, 214, 113834. [Google Scholar] [CrossRef]
- Chen, H.; Zhao, X.; Cheng, Y.; Jiang, M.; Li, X.; Xue, G. Iron Robustly Stimulates Simultaneous Nitrification and Denitrification Under Aerobic Conditions. Environ. Sci. Technol. 2018, 52, 1404–1412. [Google Scholar] [CrossRef]
- Zhao, B.; He, Y.L.; Hughes, J.; Zhang, X.F. Heterotrophic nitrogen removal by a newly isolated Acinetobacter calcoaceticus HNR. Bioresour. Technol. 2010, 101, 5194–5200. [Google Scholar] [CrossRef]
- APHA. Standard Methods for the Examination of Water and Wastewater; APHA: Washington, DC, USA, 1998. [Google Scholar]
- Frear, D.S.; Burrell, R.C. Spectrophotometric Method for Determining Hydroxylamine Reductase Activity in Higher Plants. Anal. Chem. 1955, 27, 1664–1665. [Google Scholar] [CrossRef]
- Anh, H.T.H.; Shahsavari, E.; Bott, N.J.; Ball, A.S. The application of Marinobacter hydrocarbonoclasticu as a bioaugmentation agent for the enhanced treatment of non-sterile fish wastewater. J. Environ. Manag. 2021, 291, 112658. [Google Scholar] [CrossRef] [PubMed]
- Dou, L.; Zhang, M.; Pan, L.; Liu, L.; Su, Z. Sulfide removal characteristics, pathways and potential application of a novel chemolithotrophic sulfide-oxidizing strain, Marinobacter sp. SDSWS8. Environ. Res. 2022, 212, 113176. [Google Scholar] [CrossRef] [PubMed]
- Zhang, K.; Jiang, S.; Zhang, J.; Zheng, J.; Li, P.; Wang, S.; Bi, R.; Gao, L. Phycoremediation and valorization of hypersaline pickled mustard wastewater via Chaetoceros muelleri and indigenous bacteria. Bioresour. Technol. 2024, 393, 130172. [Google Scholar] [CrossRef]
- Hu, X.; Su, J.; Ali, A.; Wang, Z.; Wu, Z. Heterotrophic nitrification and biomineralization potential of Pseudomonas sp. HXF1 for the simultaneous removal of ammonia nitrogen and fluoride from groundwater. Bioresour. Technol. 2021, 323, 124608. [Google Scholar] [CrossRef]
- Zhang, Y.; Xu, J.; Dong, X.; Wang, J.; Liu, C.; Liu, J. Optimization of nitrogen removal conditions based on response surface methodology and nitrogen removal pathway of Paracoccus sp. QD-19. Sci. Total Environ. 2024, 908, 168348. [Google Scholar] [CrossRef]
- D’Aquino, A.; Kalinainen, N.; Auvinen, H.; Andreottola, G.; Puhakka, J.A.; Palmroth, M.R.T. Effects of inorganic ions on autotrophic denitrification by Thiobacillus denitrificans and on heterotrophic denitrification by an enrichment culture. Sci. Total Environ. 2023, 901, 165940. [Google Scholar] [CrossRef] [PubMed]
- Ji, J.; Peng, Y.; Wang, B.; Mai, W.; Li, X.; Zhang, Q.; Wang, S. Effects of salinity build-up on the performance and microbial community of partial-denitrification granular sludge with high nitrite accumulation. Chemosphere 2018, 209, 53–60. [Google Scholar] [CrossRef]
- Li, W.; Li, H.; Liu, Y.-D.; Zheng, P.; Shapleigh, J.P. Salinity-Aided Selection of Progressive Onset Denitrifiers as a Means of Providing Nitrite for Anammox. Environ. Sci. Technol. 2018, 52, 10665–10672. [Google Scholar] [CrossRef]
- Asamoto, C.K.; Rempfert, K.R.; Luu, V.H.; Younkin, A.D.; Kopf, S.H. Enzyme-Specific Coupling of Oxygen and Nitrogen Isotope Fractionation of the Nap and Nar Nitrate Reductases. Environ. Sci. Technol. 2021, 55, 5537–5546. [Google Scholar] [CrossRef]
- Liang, X.; Gan, L.; He, T.; Chen, M.; Zhang, M.; Wu, Q. The coexisted nitrate and nitrite as a driving force for the aerobic denitrification of Peribacillus sp. EM-C3. Environ. Technol. Innov. 2023, 32, 103299. [Google Scholar] [CrossRef]
- Sparacino-Watkins, C.; Stolz, J.F.; Basu, P. Nitrate and periplasmic nitrate reductases. Chem. Soc. Rev. 2014, 43, 676–706. [Google Scholar] [CrossRef] [PubMed]
- Jiang, X.; Wang, H.; Wu, P.; Lei, Y.; Deng, L.; Wang, W. Enhanced narH gene expression contributing to nitrite accumulation in simultaneous nitrification and denitrification under Na+ stress instead of K+ stress. Chem. Eng. J. 2024, 479, 147637. [Google Scholar] [CrossRef]
- Shi, S.; He, X.; He, L.; Fan, X.; Shu, B.; Zhou, J.; He, Q. Overlooked pathways of endogenous simultaneous nitrification and denitrification in anaerobic/aerobic/anoxic sequencing batch reactors with organic supplementation. Water Res. 2023, 230, 119493. [Google Scholar] [CrossRef] [PubMed]
- Wen, X.; Cui, L.; Lin, H.; Zhu, W.; Shao, Z.; Wang, Y. Comparison of nitrification performance in SBR and SBBR with response to NaCl salinity shock: Microbial structure and functional genes. Environ. Res. 2024, 252, 118917. [Google Scholar] [CrossRef]
- Ke, X.; Liu, C.; Tang, S.-Q.; Guo, T.-T.; Pan, L.; Xue, Y.-P.; Zheng, Y.-G. Characterization of Acinetobacter indicus ZJB20129 for heterotrophic nitrification and aerobic denitrification isolated from an urban sewage treatment plant. Bioresour. Technol. 2022, 347, 126423. [Google Scholar] [CrossRef]
- Yang, L.; Ren, Y.-X.; Liang, X.; Zhao, S.-Q.; Wang, J.-P.; Xia, Z.-H. Nitrogen removal characteristics of a heterotrophic nitrifier Acinetobacter junii YB and its potential application for the treatment of high-strength nitrogenous wastewater. Bioresour. Technol. 2015, 193, 227–233. [Google Scholar] [CrossRef]




Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2025 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Hu, J.; Xu, B.; Gao, J.; Yan, J.; Fan, G. A Robust Oxysalt-Tolerant Bacterium Marinobacter sp. for Simultaneous Nitrification and Denitrification of Hypersaline Wastewater. Water 2025, 17, 1435. https://doi.org/10.3390/w17101435
Hu J, Xu B, Gao J, Yan J, Fan G. A Robust Oxysalt-Tolerant Bacterium Marinobacter sp. for Simultaneous Nitrification and Denitrification of Hypersaline Wastewater. Water. 2025; 17(10):1435. https://doi.org/10.3390/w17101435
Chicago/Turabian StyleHu, Jie, Bing Xu, Jie Gao, Jiabao Yan, and Guozhi Fan. 2025. "A Robust Oxysalt-Tolerant Bacterium Marinobacter sp. for Simultaneous Nitrification and Denitrification of Hypersaline Wastewater" Water 17, no. 10: 1435. https://doi.org/10.3390/w17101435
APA StyleHu, J., Xu, B., Gao, J., Yan, J., & Fan, G. (2025). A Robust Oxysalt-Tolerant Bacterium Marinobacter sp. for Simultaneous Nitrification and Denitrification of Hypersaline Wastewater. Water, 17(10), 1435. https://doi.org/10.3390/w17101435