

Application of General Unified Threshold Models to Predict Time-Varying Survival of Mayfly Nymphs Exposed to Three Neonicotinoids



{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
1. Introduction
2. Materials and Methods
2.1. Experimental Data Source
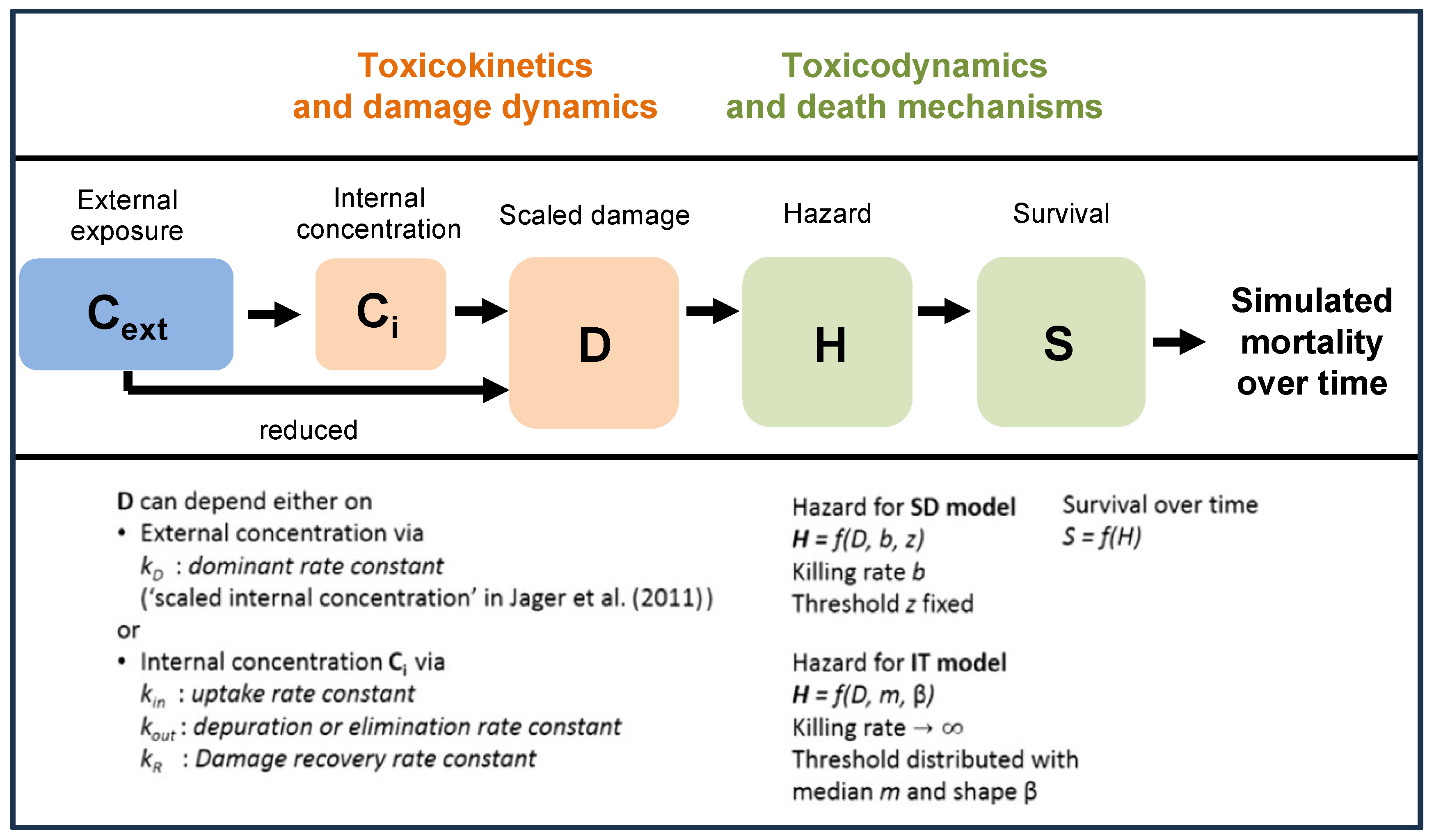
2.2. GUTS Models Evaluated
2.3. Model Calibrations and Quality Assessment of Model Predictions
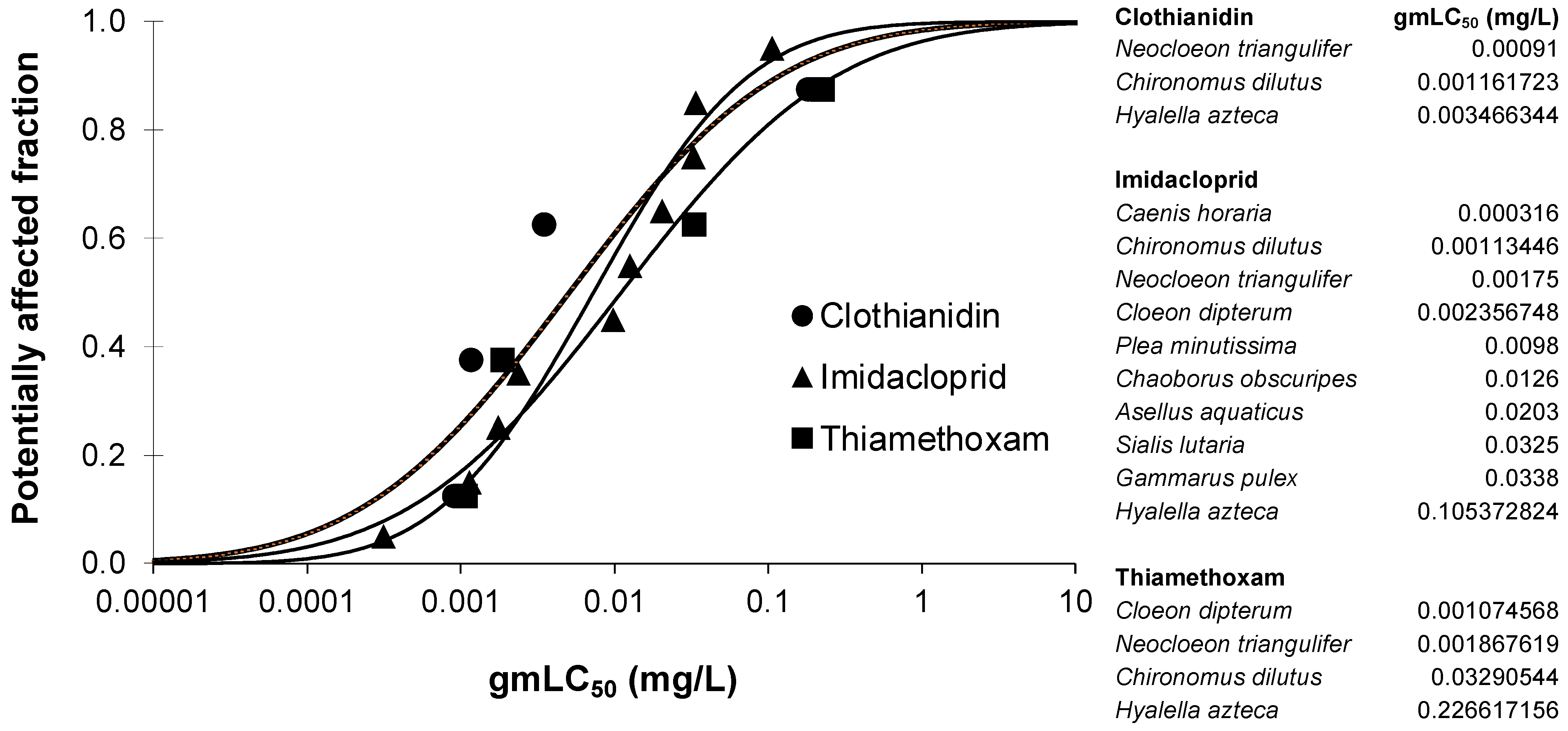
2.4. Species Sensitivity Distributions (SSDs)
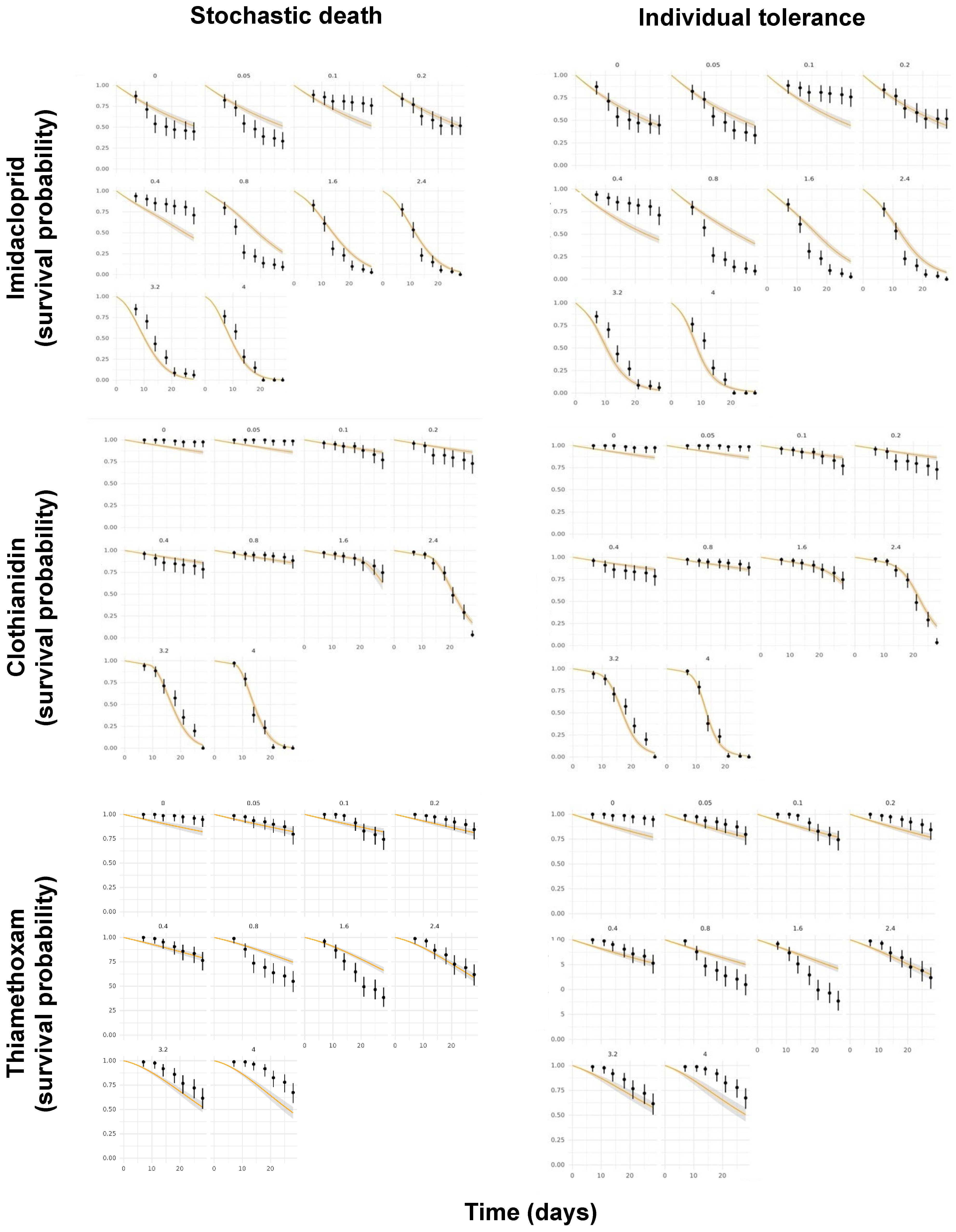
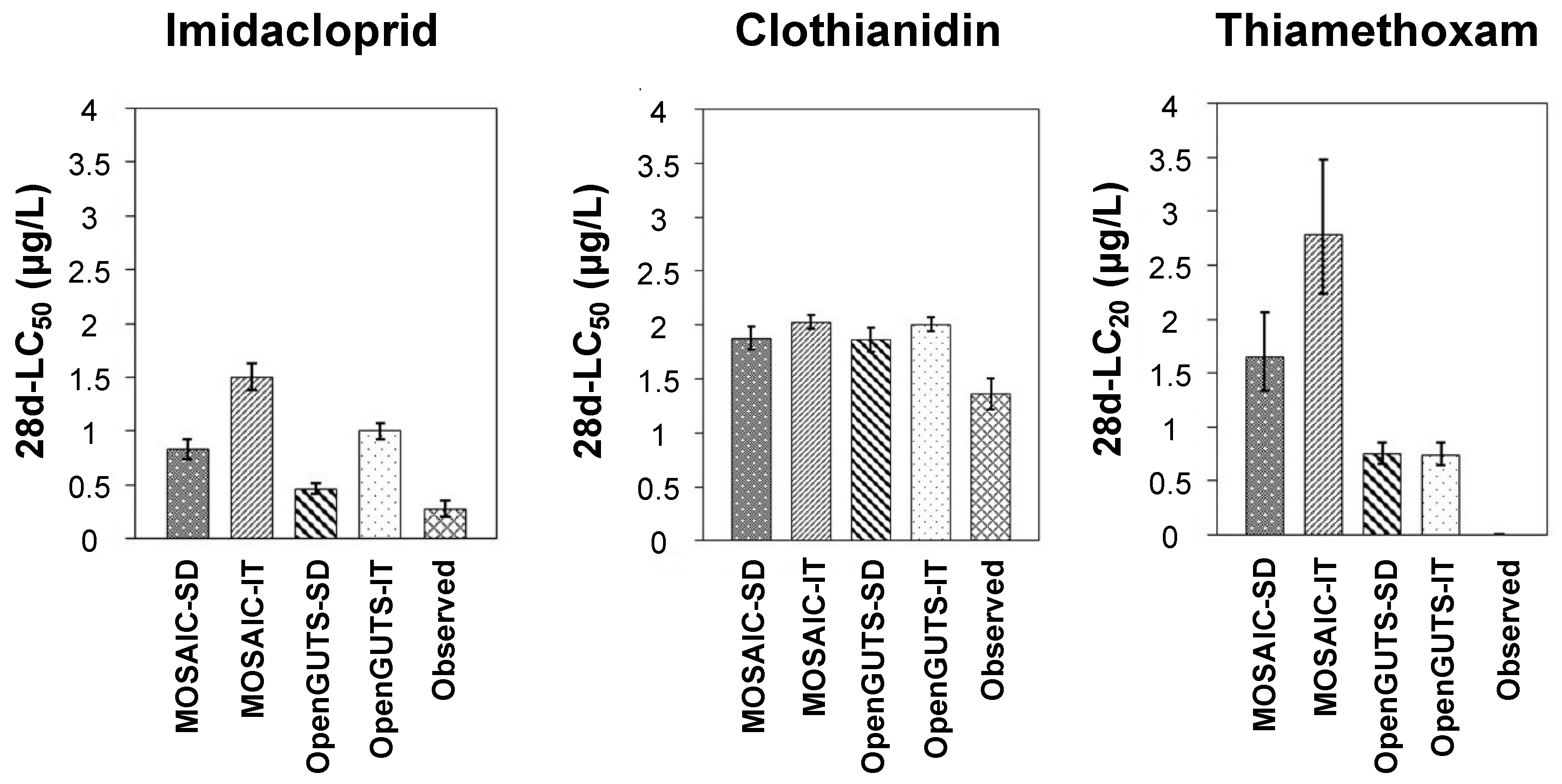
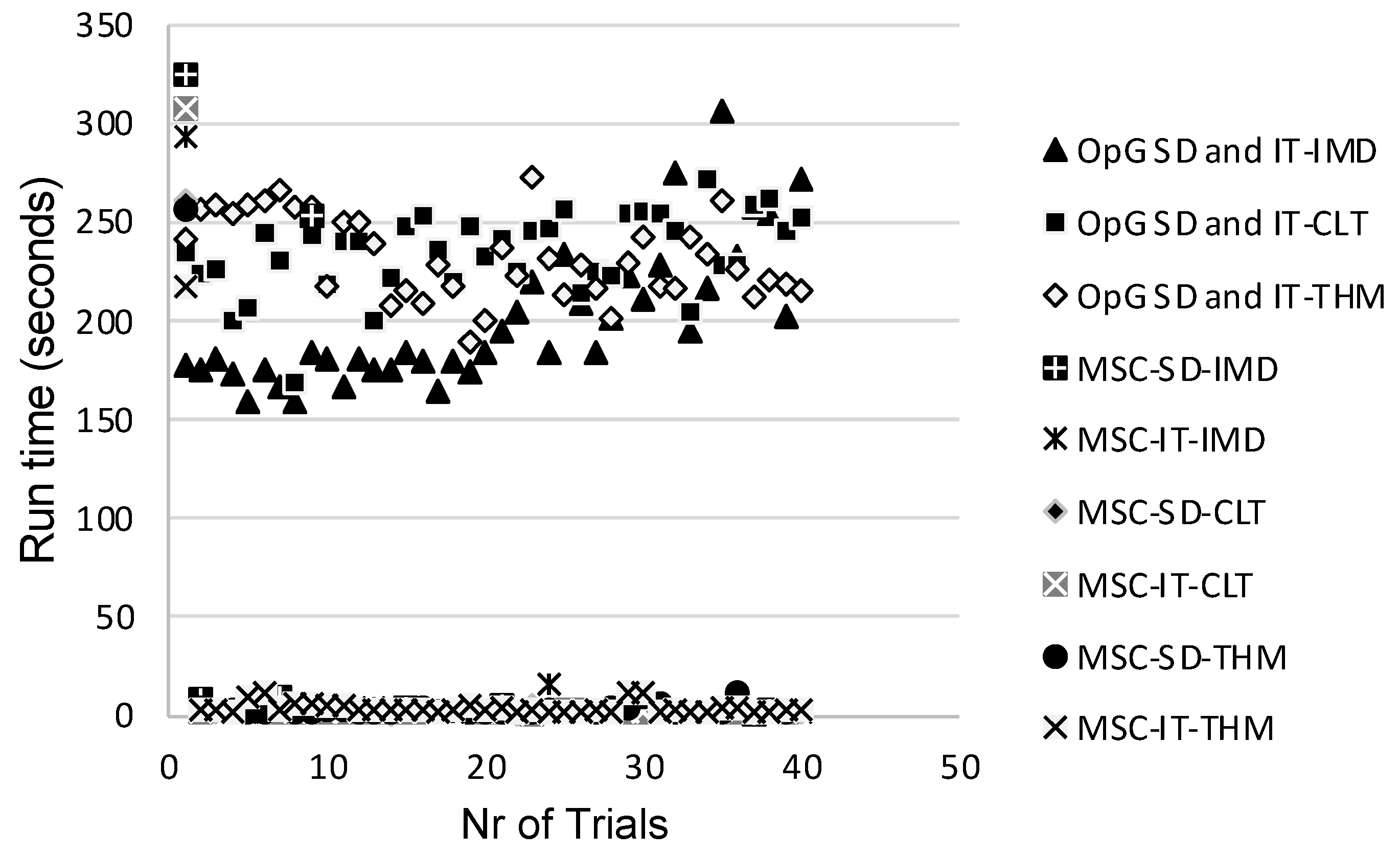
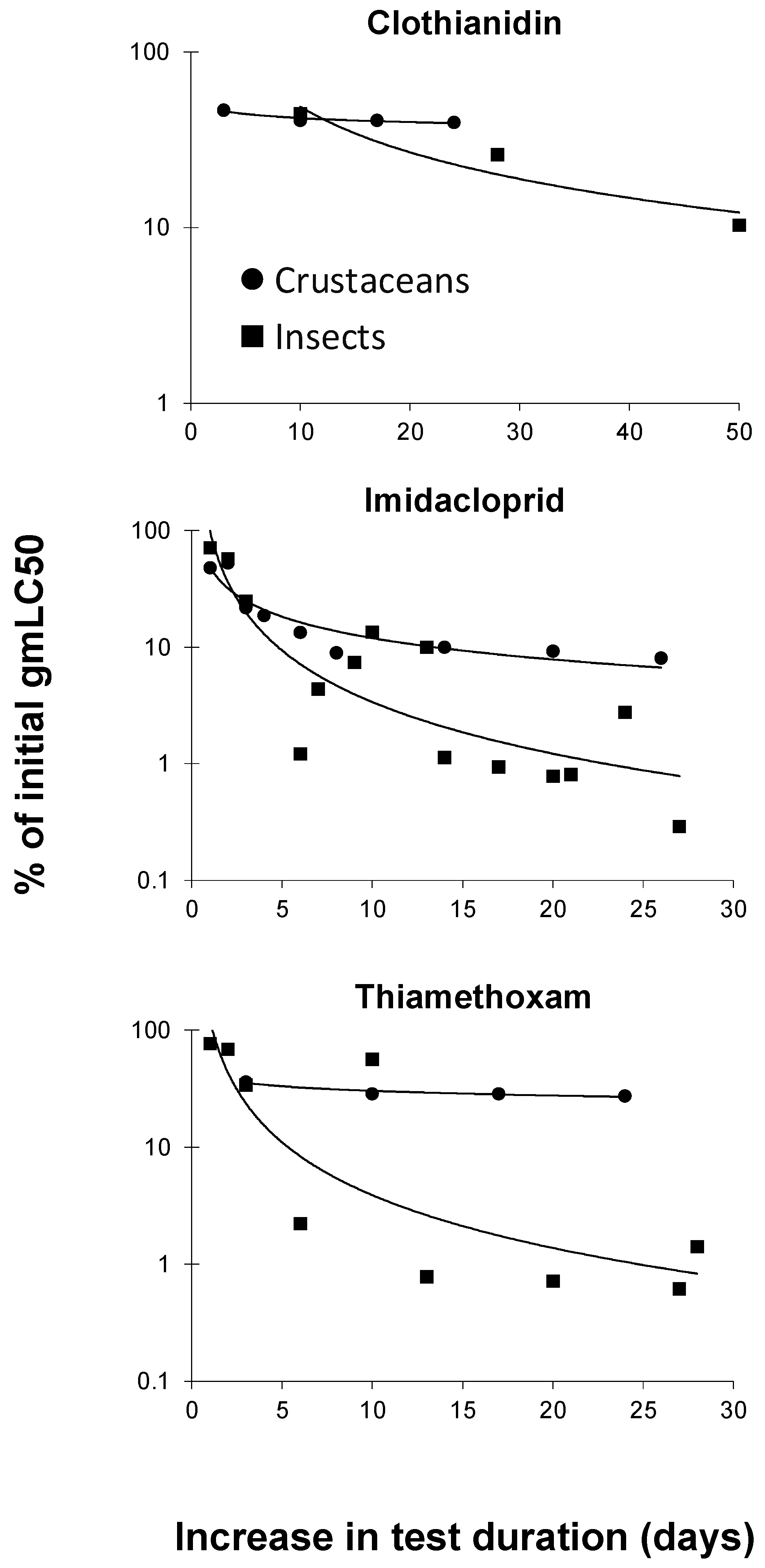
3. Results and Discussion
3.1. Relative Toxicities of Test Compounds
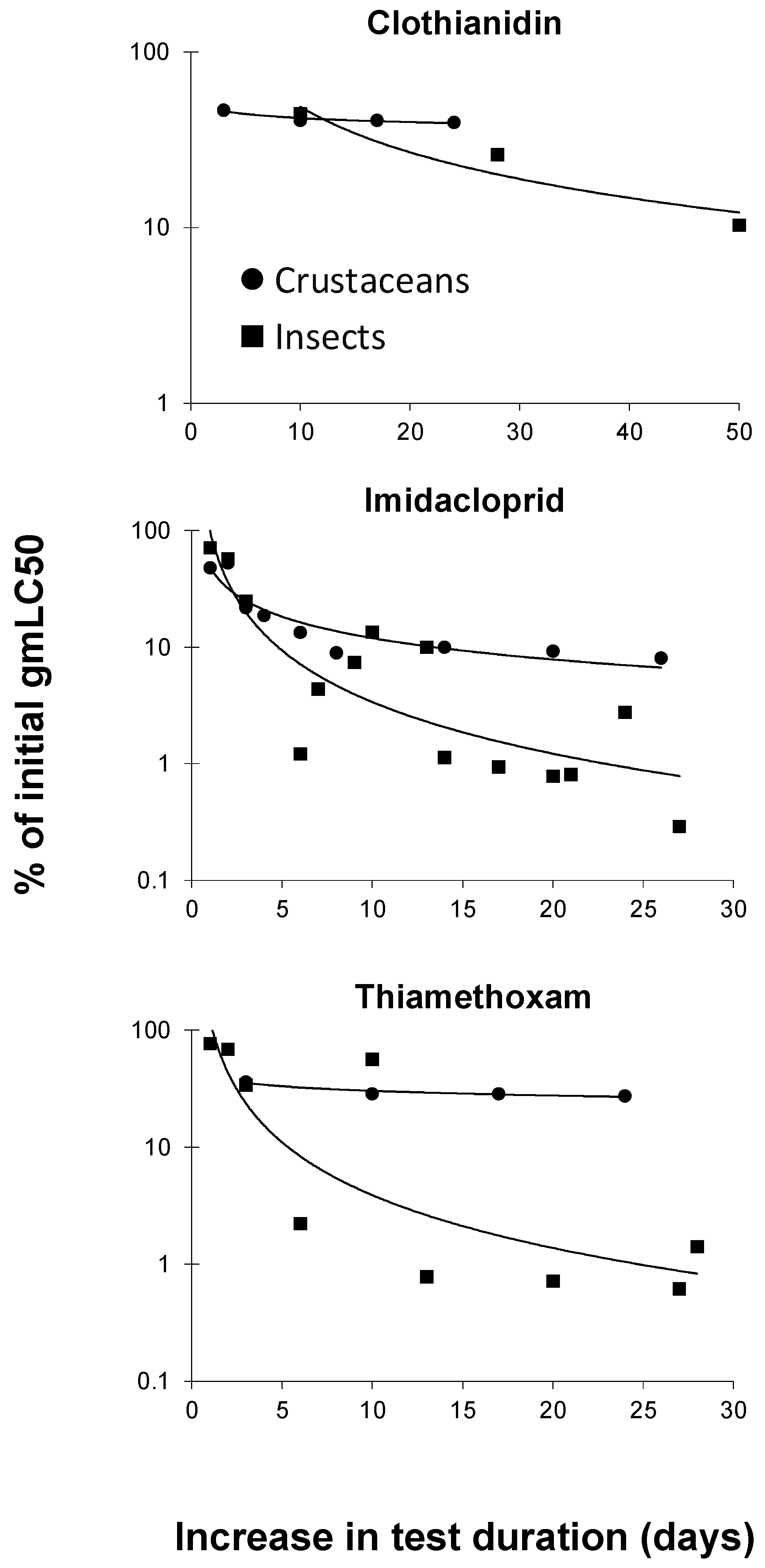
3.2. Toxicity of Neonicotinoids Increased with Increasing Test Duration
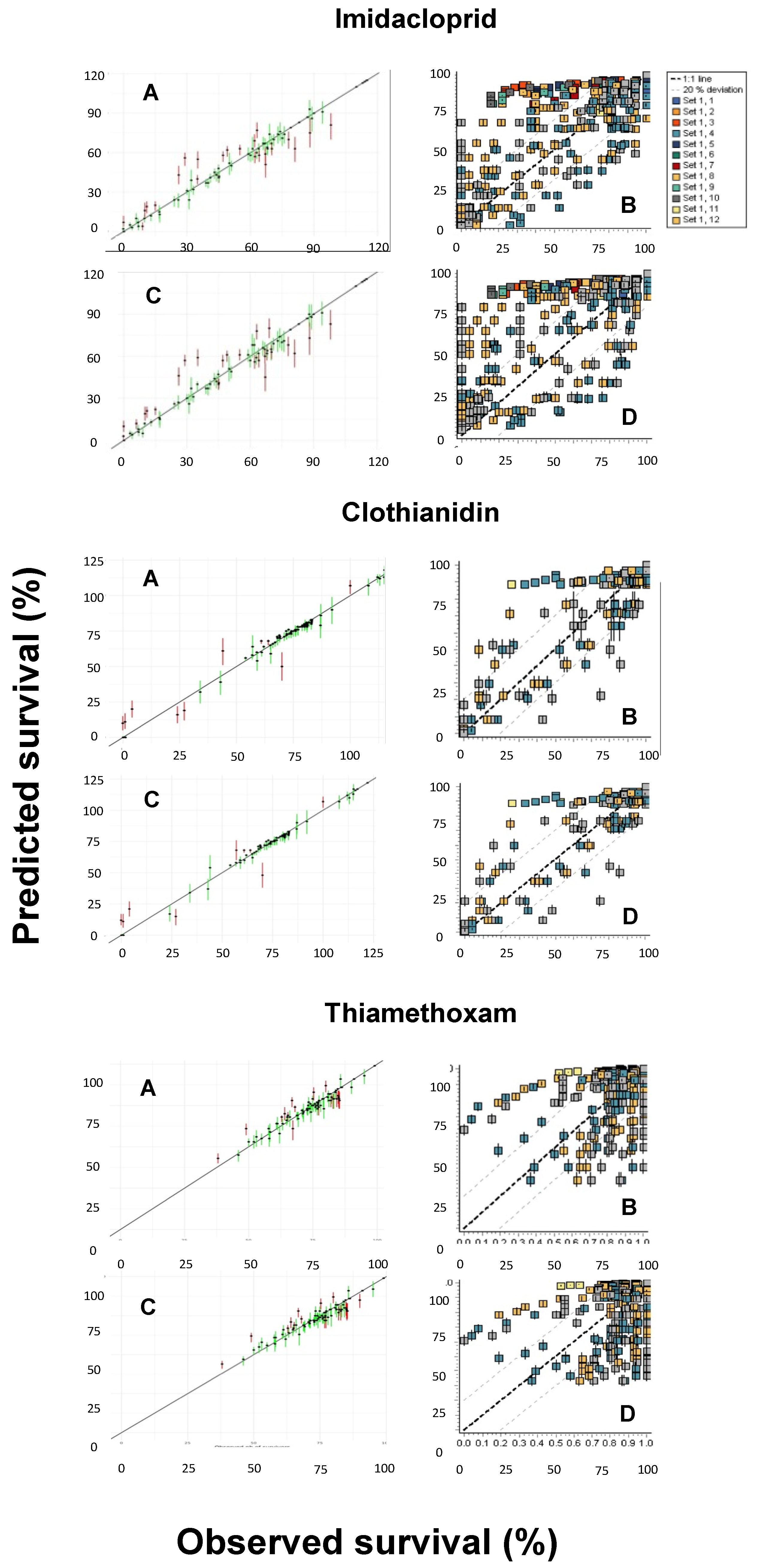
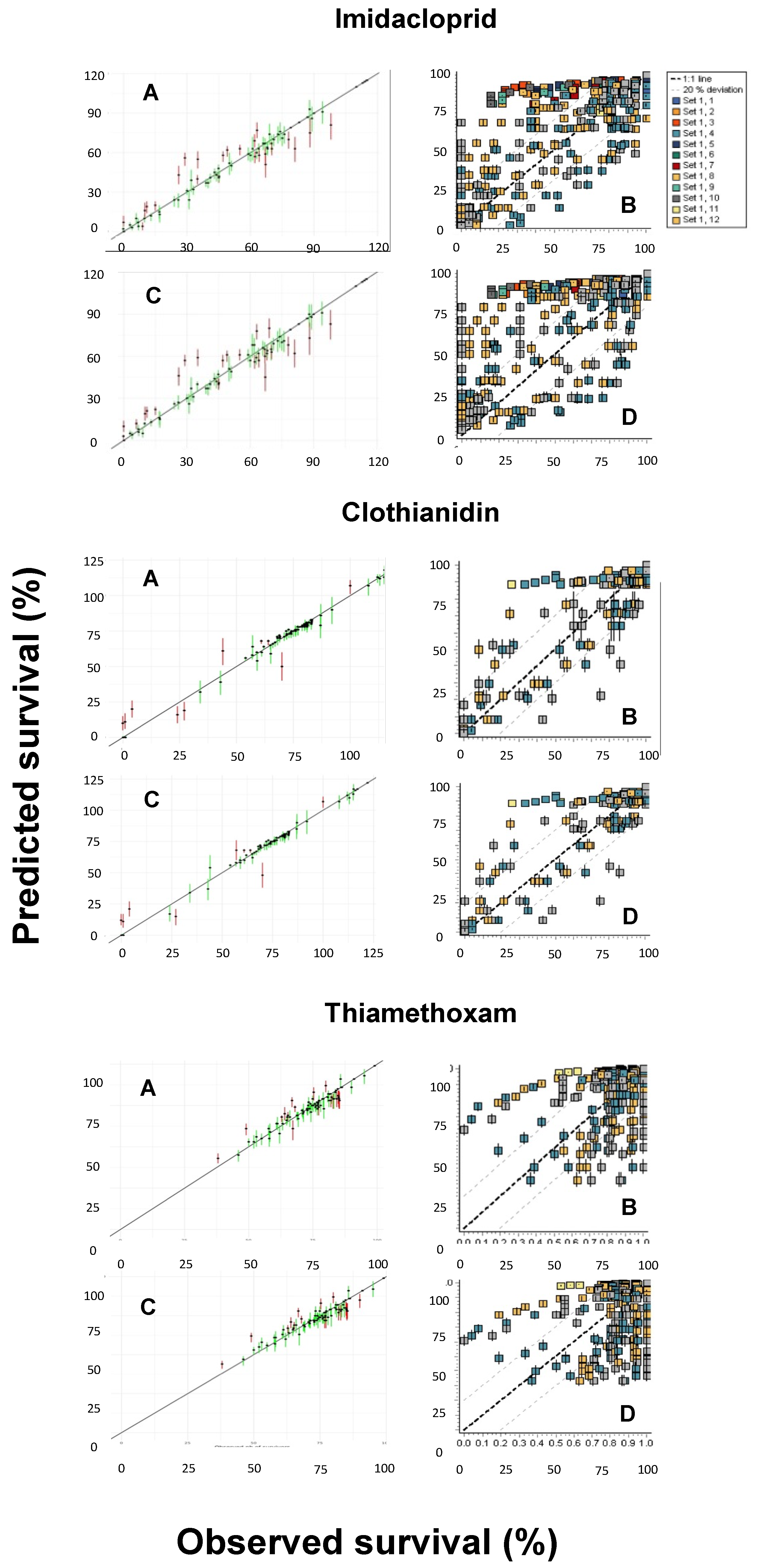
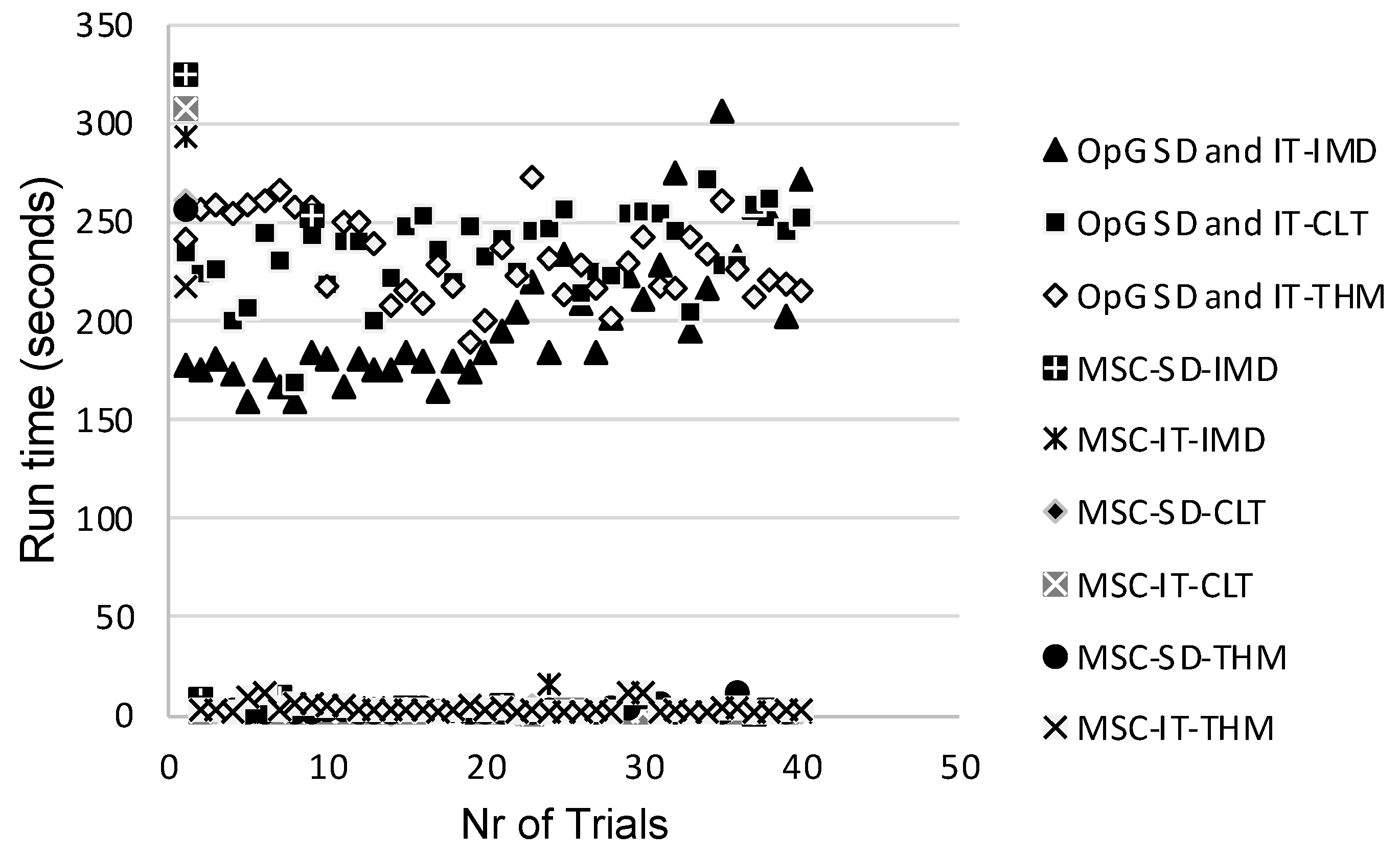
3.3. Comparison of Model Performances
3.4. Predicted versus Observed Toxicity Values
3.5. Concluding Remarks and Indications for Future Research
Author Contributions
Funding
Data Availability Statement
Conflicts of Interest
References
- UNEP. Environmental and Health Impacts of Pesticides and Fertilizers and Ways of Minimizing Them: Synthesis Report; United Nations Environment Program: Nairobi, Kenya, 2022; ISBN 9789280739299. [Google Scholar]
- Grandview Research Agrochemicals Market Size To Reach $289.56 Billion by 2030. Available online: https://www.grandviewresearch.com/press-release/global-agrochemicals-market (accessed on 17 January 2024).
- Capri, E.; Trevisan, M. Prediction of Environmental Concentrations (PECs) by Mathematical Model Application in Europe. Pestic. Outlook 1998, 9, 26–30. [Google Scholar]
- Daam, M.A.; Van Den Brink, P.J. Risk Assessment of Agrochemicals on Irrigation Water Quality. Acta Hortic. 2011, 922, 41–47. [Google Scholar] [CrossRef]
- Van den Brink, P.J.; Daam, M.A. Multispecies Environmental Testing Designs. In Encyclopedia of Toxicology, 3rd ed.; Elsevier: Amsterdam, The Netherlands, 2014; pp. 399–402. ISBN 9780123864543. [Google Scholar]
- Beketov, M.A.; Liess, M. Acute and Delayed Effects of the Neonicotinoid Insecticide Thiacloprid on Seven Freshwater Arthropods. Environ. Toxicol. Chem. 2008, 27, 461–470. [Google Scholar] [CrossRef] [PubMed]
- EFSA. Guidance on Tiered Risk Assessment for Plant Protection Products for Aquatic Organisms in Edge-of-Field Surface Waters. EFSA J. 2013, 11, 3290. [Google Scholar] [CrossRef]
- FOCUS Surface Water Scenarios in the EU Evaluation Process under 91/414/EEC. Report of the FOCUS Working Group on Surface Water Scenarios. EC Document Reference SANCO/4802/2001-Rev.2; EUROPEAN COMMISSION: Brussels, Belgium, 2001.
- Focks, A.; Belgers, D.; Boerwinkel, M.C.; Buijse, L.; Roessink, I.; Van den Brink, P.J. Calibration and Validation of Toxicokinetic-Toxicodynamic Models for Three Neonicotinoids and Some Aquatic Macroinvertebrates. Ecotoxicology 2018, 27, 992–1007. [Google Scholar] [CrossRef] [PubMed]
- Casallanovo, F.; Santos, G.S.; Cione, A.P.; Simone, D.M.; de Oliveira Kaminski, T.S.; Chen, W. Qualitative Analysis of the Most Locally Relevant Runoff and Erosion Parameters for Constructing Brazilian Scenarios. Integr. Environ. Assess. Manag. 2023, 19, 1374–1384. [Google Scholar] [CrossRef] [PubMed]
- Casallanovo, F.; Simone, D.M.; Santos, G.S.; de Oliveira Kaminski, T.S.; Cione, A.P.; Peranginangin, N. Estimating Pesticide Environmental Concentrations in Latin America: The Importance of Developing Local Scenarios. Integr. Environ. Assess. Manag. 2021, 17, 901–904. [Google Scholar] [CrossRef]
- Daam, M.A.; Cerejeira, M.J.; van den Brink, P.J.; Brock, T.C.M. Is It Possible to Extrapolate Results of Aquatic Microcosm and Mesocosm Experiments with Pesticides between Climate Zones in Europe? Environ. Sci. Pollut. Res. 2011, 18, 123–126. [Google Scholar] [CrossRef] [PubMed]
- Baudrot, V.; Charles, S. Recommendations to Address Uncertainties in Environmental Risk Assessment Using Toxicokinetic-Toxicodynamic Models. Sci. Rep. 2019, 9, 11432. [Google Scholar] [CrossRef]
- Ashauer, R.; Escher, B.I. Advantages of Toxicokinetic and Toxicodynamic Modelling in Aquatic Ecotoxicology and Risk Assessment. J. Environ. Monit. 2010, 12, 2056–2061. [Google Scholar] [CrossRef]
- Jager, T.; Albert, C.; Preuss, T.G.; Ashauer, R. General Unified Threshold Model of Survival—A Toxicokinetic-Toxicodynamic Framework for Ecotoxicology. Environ. Sci. Technol. 2011, 45, 2529–2540. [Google Scholar] [CrossRef]
- Jager, T. Robust Likelihood-Based Approach for Automated Optimization and Uncertainty Analysis of Toxicokinetic-Toxicodynamic Models. Integr. Environ. Assess. Manag. 2021, 17, 388–397. [Google Scholar] [CrossRef]
- Brock, T.; Arena, M.; Cedergreen, N.; Charles, S.; Duquesne, S.; Ippolito, A.; Klein, M.; Reed, M.; Teodorovic, I.; van den Brink, P.J.; et al. Application of General Unified Threshold Models of Survival Models for Regulatory Aquatic Pesticide Risk Assessment Illustrated with an Example for the Insecticide Chlorpyrifos. Integr. Environ. Assess. Manag. 2021, 17, 243–258. [Google Scholar] [CrossRef] [PubMed]
- EFSA. Scientific Opinion on the State of the Art of Toxicokinetic/Toxicodynamic (TKTD) Effect Models for Regulatory Risk Assessment of Pesticides for Aquatic Organisms. EFSA J. 2018, 16, e05377. [Google Scholar] [CrossRef]
- Larras, F.; Charles, S.; Chaumot, A.; Pelosi, C.; Le Gall, M.; Mamy, L.; Beaudouin, R. A Critical Review of Effect Modeling for Ecological Risk Assessment of Plant Protection Products. Environ. Sci. Pollut. Res. 2022, 29, 43448–43500. [Google Scholar] [CrossRef] [PubMed]
- Van den Brink, P.J.; Van Smeden, J.M.; Bekele, R.S.; Dierick, W.; De Gelder, D.M.; Noteboom, M.; Roessink, I. Acute and Chronic Toxicity of Neonicotinoids to Nymphs of a Mayfly Species and Some Notes on Seasonal Differences. Environ. Toxicol. Chem. 2016, 35, 128–133. [Google Scholar] [CrossRef] [PubMed]
- Morrissey, C.A.; Mineau, P.; Devries, J.H.; Sanchez-Bayo, F.; Liess, M.; Cavallaro, M.C.; Liber, K. Neonicotinoid Contamination of Global Surface Waters and Associated Risk to Aquatic Invertebrates: A Review. Environ. Int. 2015, 74, 291–303. [Google Scholar] [CrossRef] [PubMed]
- Roessink, I.; Merga, L.B.; Zweers, H.J.; Van den Brink, P.J. The Neonicotinoid Imidacloprid Shows High Chronic Toxicity to Mayfly Nymphs. Environ. Toxicol. Chem. 2013, 32, 1096–1100. [Google Scholar] [CrossRef]
- Sibley, P.; Lagadic, L.; McCoole, M.; Norberg-King, T.; Roessink, I.; Soucek, D.; Watson-Leung, T.; Wirtz, J. Mayflies in Ecotoxicity Testing: Methodological Needs and Knowledge Gaps. Integr. Environ. Assess. Manag. 2020, 16, 292–293. [Google Scholar] [CrossRef]
- Macaulay, S.J.; Hageman, K.J.; Alumbaugh, R.E.; Lyons, S.M.; Piggott, J.J.; Matthaei, C.D. Chronic Toxicities of Neonicotinoids to Nymphs of the Common New Zealand Mayfly Deleatidium spp. Environ. Toxicol. Chem. 2019, 38, 2459–2471. [Google Scholar] [CrossRef]
- Schneeweiss, A.; Juvigny-Khenafou, N.P.D.; Osakpolor, S.; Scharmüller, A.; Scheu, S.; Schreiner, V.C.; Ashauer, R.; Escher, B.I.; Leese, F.; Schäfer, R.B. Three Perspectives on the Prediction of Chemical Effects in Ecosystems. Glob. Chang. Biol. 2023, 29, 21–40. [Google Scholar] [CrossRef] [PubMed]
- Van Vlaardingen, P.L.A.; Traas, T.P.; Wintersen, A.M.; Aldenberg, T. ETX 2.0. A Program. to Calculate Hazardous Concentrations and Fraction Affected, Based on Normally Distributed Toxicity Data. 2004. Available online: https://www.researchgate.net/publication/27452022_ETX_20_A_Program_to_Calculate_Hazardous_Concentrations_and_Fraction_Affected_Based_on_Normally_Distributed_Toxicity_Data (accessed on 15 March 2021).
- Aldenberg, T.; Jaworska, J.S. Uncertainty of the Hazardous Concentration and Fraction Affected for Normal Species Sensitivity Distributions. Ecotoxicol. Environ. Saf. 2000, 46, 1–18. [Google Scholar] [CrossRef] [PubMed]
- Zhang, A.; Kayser, H.; Maienfisch, P.; Casida, J.E. Insect Nicotinic Acetylcholine Receptor: Conserved Neonicotinoid Specificity of [3H]Imidacloprid Binding Site. J. Neurochem. 2000, 75, 1294–1303. [Google Scholar] [CrossRef] [PubMed]
- Iwasa, T.; Motoyama, N.; Ambrose, J.T.; Roe, R.M. Mechanism for the Differential Toxicity of Neonicotinoid Insecticides in the Honey Bee, Apis mellifera. Crop Prot. 2004, 23, 371–378. [Google Scholar] [CrossRef]
- Manjon, C.; Troczka, B.J.; Zaworra, M.; Beadle, K.; Randall, E.; Hertlein, G.; Singh, K.S.; Zimmer, C.T.; Homem, R.A.; Lueke, B.; et al. Unravelling the Molecular Determinants of Bee Sensitivity to Neonicotinoid Insecticides. Curr. Biol. 2018, 28, 1137–1143.e5. [Google Scholar] [CrossRef] [PubMed]
- Yamamoto, I.; Tomizawa, M.; Saito, T.; Miyamoto, T.; Walcott, E.C.; Sumikawa, K. Structural Factors Contributing to Insecticidal and Selective Actions of Neonicotinoids. Arch. Insect Biochem. Physiol. 1998, 37, 24–32. [Google Scholar] [CrossRef]
- Vehovszky, Á.; Farkas, A.; Ács, A.; Stoliar, O.; Székács, A.; Mörtl, M.; Gyori, J. Neonicotinoid Insecticides Inhibit Cholinergic Neurotransmission in a Molluscan (Lymnaea stagnalis) Nervous System. Aquat. Toxicol. 2015, 167, 172–179. [Google Scholar] [CrossRef]
- Raby, M.; Zhao, X.; Hao, C.; Poirier, D.G.; Sibley, P.K. Chronic Toxicity of 6 Neonicotinoid Insecticides to Chironomus dilutus and Neocloeon triangulifer. Environ. Toxicol. Chem. 2018, 37, 2727–2739. [Google Scholar] [CrossRef] [PubMed]
- Song, M.Y.; Brown, J.J. Influence of Fluctuating Salinity on Insecticide Tolerance of Two Euryhaline Arthropods. J. Econ. Entomol. 2006, 99, 745–751. [Google Scholar] [CrossRef]
- Nyman, A.M.; Schirmer, K.; Ashauer, R. Toxicokinetic-Toxicodynamic Modelling of Survival of Gammarus pulex in Multiple Pulse Exposures to Propiconazole: Model Assumptions, Calibration Data Requirements and Predictive Power. Ecotoxicology 2012, 21, 1828–1840. [Google Scholar] [CrossRef]
- He, A.; Liu, X.; Qu, L.; Gao, Y.; Feng, J.; Zhu, L. Comparison of the General Threshold Model of Survival and Dose–Response Models in Simulating the Acute Toxicity of Metals to Danio rerio. Environ. Toxicol. Chem. 2019, 38, 2169–2177. [Google Scholar] [CrossRef] [PubMed]
- Baudrot, V.; Veber, P.; Gence, G.; Charles, S. Fit Reduced GUTS Models Online: From Theory to Practice. Integr. Environ. Assess. Manag. 2018, 14, 625–630. [Google Scholar] [CrossRef]
- Baudrot, V.; Preux, S.; Ducrot, V.; Pave, A.; Charles, S. New Insights to Compare and Choose TKTD Models for Survival Based on an Interlaboratory Study for Lymnaea stagnalis Exposed to Cd. Environ. Sci. Technol. 2018, 52, 1582–1590. [Google Scholar] [CrossRef]
- Andersen, J.S.; Holst, H.; Spliid, H.; Andersen, H.; Baun, A.; Nyholm, N. Continuous Ecotoxicological Data Evaluated Relative to a Control Response. J. Agric. Biol. Environ. Stat. 1998, 3, 405–420. [Google Scholar] [CrossRef]
- Huang, A.; Roessink, I.; van den Brink, N.W.; van den Brink, P.J. Size- and Sex-Related Sensitivity Differences of Aquatic Crustaceans to Imidacloprid. Ecotoxicol. Environ. Saf. 2022, 242, 113917. [Google Scholar] [CrossRef] [PubMed]
- Bart, S.; Jager, T.; Robinson, A.; Lahive, E.; Spurgeon, D.J.; Ashauer, R. Predicting Mixture Effects over Time with Toxicokinetic-Toxicodynamic Models (Guts): Assumptions, Experimental Testing, and Predictive Power. Environ. Sci. Technol. 2021, 55, 2430–2439. [Google Scholar] [CrossRef]
- Tagliati, A.; Hazlerigg, C.; Brignall, S.; Beinum, W. Van Exploring GUTS in the Environmental Risk Assessment of Pesticides. In Proceedings of the Poster Presentation a the SETAC EUROPE 32 ND Annual Meeting, Copenhagen, Denmark, 15–19 May 2022. [Google Scholar]
- Charles, S.; Ratier, A.; Baudrot, V.; Multari, G.; Siberchicot, A.; Wu, D.; Lopes, C. Taking Full Advantage of Modelling to Better Assess Environmental Risk Due to Xenobiotics—The All-in-One Facility MOSAIC. Environ. Sci. Pollut. Res. 2022, 29, 29244–29257. [Google Scholar] [CrossRef]




Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2024 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Lourenço, V.S.C.; Figueiredo, N.L.; Daam, M.A. Application of General Unified Threshold Models to Predict Time-Varying Survival of Mayfly Nymphs Exposed to Three Neonicotinoids. Water 2024, 16, 1082. https://doi.org/10.3390/w16081082
Lourenço VSC, Figueiredo NL, Daam MA. Application of General Unified Threshold Models to Predict Time-Varying Survival of Mayfly Nymphs Exposed to Three Neonicotinoids. Water. 2024; 16(8):1082. https://doi.org/10.3390/w16081082
Chicago/Turabian StyleLourenço, Vanessa S. C., Neusa L. Figueiredo, and Michiel A. Daam. 2024. "Application of General Unified Threshold Models to Predict Time-Varying Survival of Mayfly Nymphs Exposed to Three Neonicotinoids" Water 16, no. 8: 1082. https://doi.org/10.3390/w16081082
APA StyleLourenço, V. S. C., Figueiredo, N. L., & Daam, M. A. (2024). Application of General Unified Threshold Models to Predict Time-Varying Survival of Mayfly Nymphs Exposed to Three Neonicotinoids. Water, 16(8), 1082. https://doi.org/10.3390/w16081082