
Determining Water Use Patterns for the Betula platyphylla in the Southern Greater Khingan Mountains and Its Adaptation to Drought


 ,
,  ,
,
Abstract
1. Introduction
2. Materials and Methods
2.1. Study Area
2.2. Environmental and Meteorological Measurements
2.3. Experiment Design and Sampling
2.4. Isotopic Analysis
2.5. Data Analyses
3. Results
3.1. Precipitation and Soil Moisture Characteristics
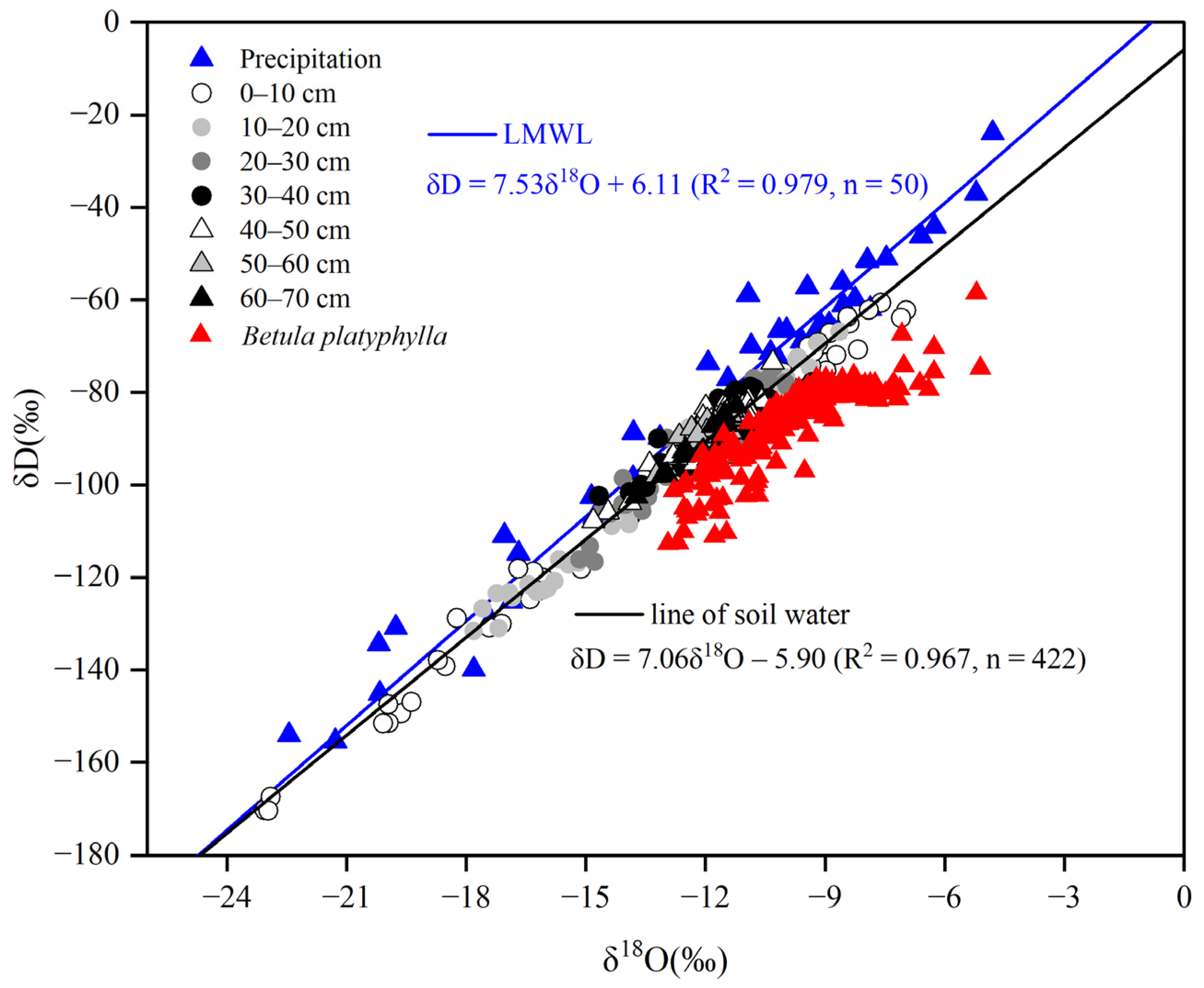
3.2. Precipitation, Soil Water, Xylem Water and Isotopic Composition
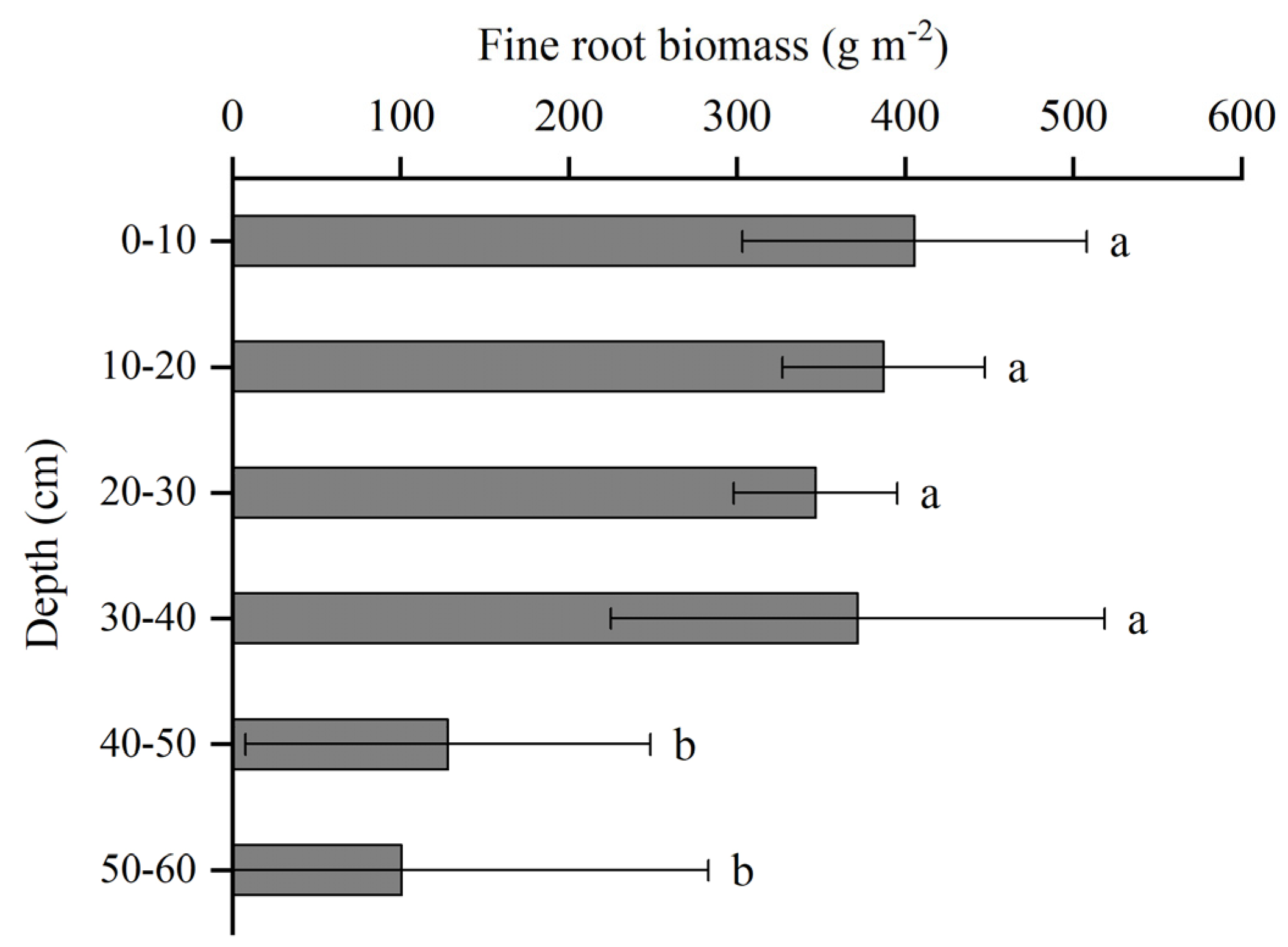
3.3. Root Distribution
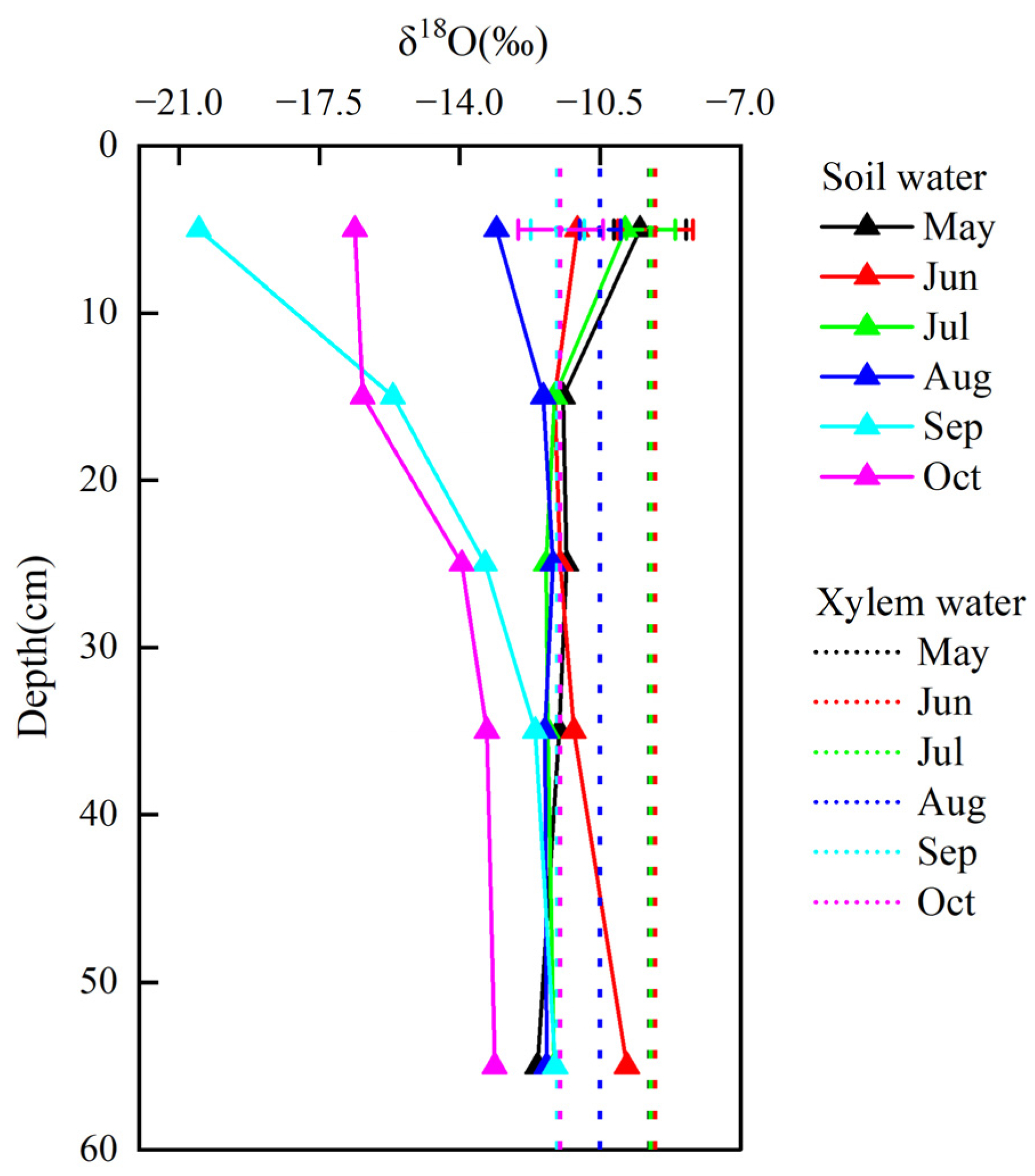
3.4. Soil Moisture and Isotopic Composition
3.5. Depth of Water Utilization
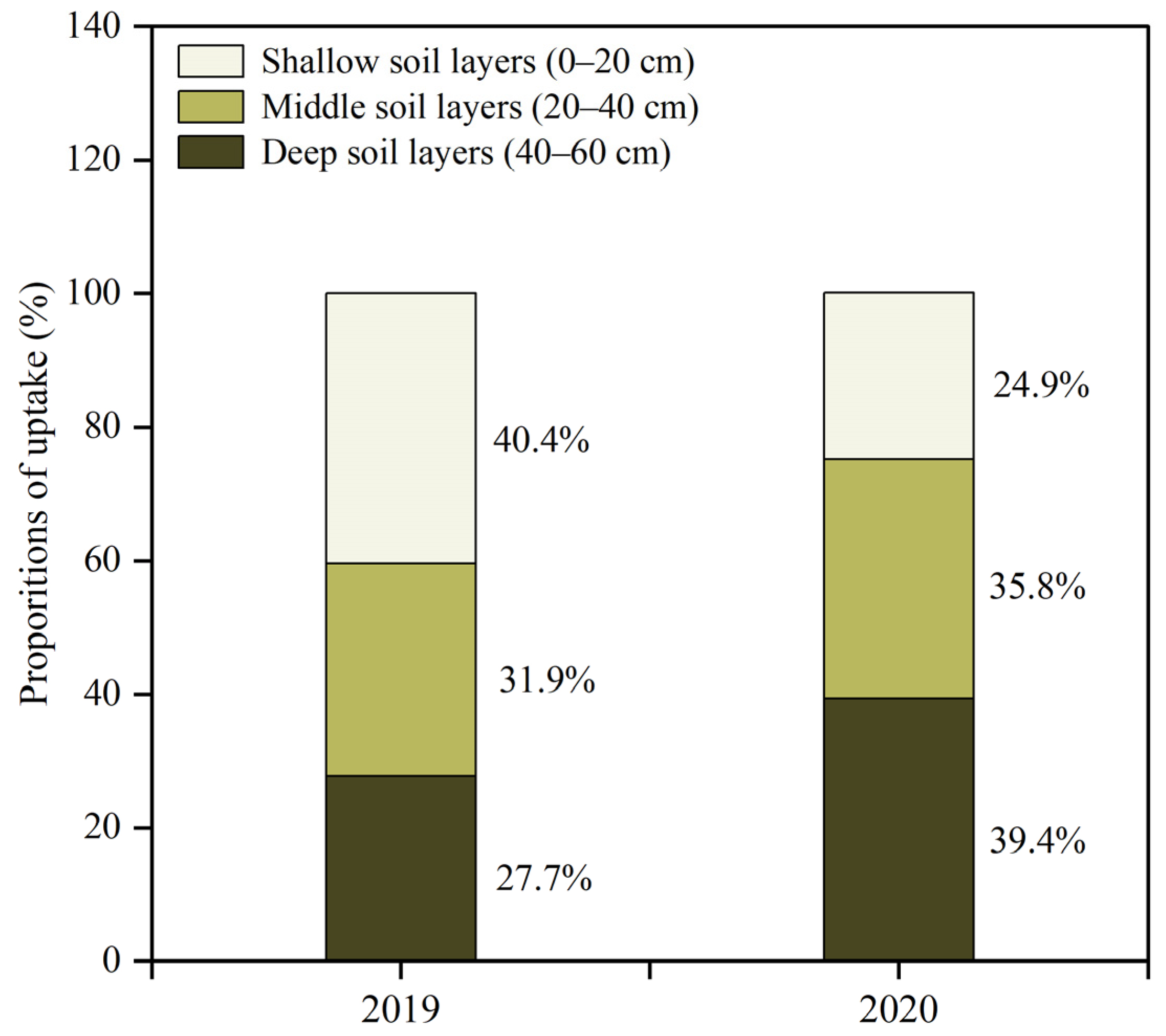
3.6. Seasonal and Interannual Variations in the Proportion of Betula platyphylla Water Uptake
4. Discussion
5. Conclusions
Author Contributions
Funding
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Zeng, N.; Yao, H.; Zhou, M.; Zhao, P.; Dech, J.P.; Zhang, B.; Lu, X. Species-specific determinants of mortality and recruitment in the forest-steppe ecotone of northeast China. For. Chron. 2016, 92, 336–344. [Google Scholar] [CrossRef]
- Xu, C.; Liu, H.; Hampe, A. Hydraulic adaptability promotes tree life spans under climate dryness. Glob. Ecol. Biogeogr. 2021, 31, 51–61. [Google Scholar] [CrossRef]
- Brodribb, T.J.; Cochard, H.; Dominguez, C.R.; Hultine, K. Measuring the pulse of trees; using the vascular system to predict tree mortality in the 21st century. Conserv. Physiol. 2019, 7, coz046. [Google Scholar] [CrossRef] [PubMed]
- Choat, B.; Jansen, S.; Brodribb, T.J.; Cochard, H.; Delzon, S.; Bhaskar, R.; Bucci, S.J.; Feild, T.S.; Gleason, S.M.; Hacke, U.G.; et al. Global convergence in the vulnerability of forests to drought. Nature 2012, 491, 752–755. [Google Scholar] [CrossRef]
- Boisvenue, C.; Running, S.W. Impacts of climate change on natural forest productivity—Evidence since the middle of the 20th century. Glob. Change Biol. 2006, 12, 862–882. [Google Scholar] [CrossRef]
- Clark, J.S.; Iverson, L.; Woodall, C.W.; Allen, C.D.; Bell, D.M.; Bragg, D.C.; D’Amato, A.W.; Davis, F.W.; Hersh, M.H.; Ibanez, I.; et al. The impacts of increasing drought on forest dynamics, structure, and biodiversity in the United States. Glob. Change Biol. 2016, 22, 2329–2352. [Google Scholar] [CrossRef]
- Miguez-Macho, G.; Fan, Y. Spatiotemporal origin of soil water taken up by vegetation. Nature 2021, 598, 624–628. [Google Scholar] [CrossRef] [PubMed]
- Cui, Y.; Ma, J.; Sun, W.; Sun, J.; Duan, Z. A preliminary study of water use strategy of desert plants in Dunhuang, China. J. Arid. Land 2014, 7, 73–81. [Google Scholar] [CrossRef]
- Liu, H.; Park Williams, A.; Allen, C.D.; Guo, D.; Wu, X.; Anenkhonov, O.A.; Liang, E.; Sandanov, D.V.; Yin, Y.; Qi, Z.; et al. Rapid warming accelerates tree growth decline in semi-arid forests of Inner Asia. Glob. Change Biol. 2013, 19, 2500–2510. [Google Scholar] [CrossRef]
- Köstner, B.; Granier, A.; Cermák, J. Sapflow measurements in forest stands: Methods and uncertainties. Ann. Des. Sci. For. 1998, 55, 13–27. [Google Scholar] [CrossRef]
- Zhang, Y.; Zhang, M.; Qu, D.; Duan, W.; Wang, J.; Su, P.; Guo, R. Water Use Strategies of Dominant Species (Caragana korshinskii and Reaumuria soongorica) in Natural Shrubs Based on Stable Isotopes in the Loess Hill, China. Water 2020, 12, 1923. [Google Scholar] [CrossRef]
- Zhao, G.; Li, X.; Wu, H.; Zhang, S.; Li, G. Study on plant water use in Myricaria squamosa with stable hydrogen isotope tracer in Qing-hai Lake basin. Chin. J. Plant Ecol. 2013, 37, 1091–1100. [Google Scholar] [CrossRef]
- Campbell, G.S.; Norman, J. An Introduction to Environmental Biophysics; Springer: New York, NY, USA, 1998. [Google Scholar]
- Chen, J.; Xu, Q.; Gao, D.; Song, A.; Hao, Y.; Ma, Y. Differential water use strategies among selected rare and endangered species in West Ordos Desert of China. J. Plant Ecol. 2016, 10, 660–669. [Google Scholar] [CrossRef]
- Nie, Y.-P.; Chen, H.-S.; Wang, K.-L.; Tan, W.; Deng, P.-Y.; Yang, J. Seasonal water use patterns of woody species growing on the continuous dolostone outcrops and nearby thin soils in subtropical China. Plant Soil 2010, 341, 399–412. [Google Scholar] [CrossRef]
- Eggemeyer, K.D.; Awada, T.; Harvey, F.E.; Wedin, D.A.; Zhou, X.; Zanner, C.W. Seasonal changes in depth of water uptake for encroaching trees Juniperus virginiana and Pinus ponderosa and two dominant C4 grasses in a semiarid grassland. Tree Physiol. 2008, 29, 157–169. [Google Scholar] [CrossRef]
- Sun, Z.; Long, X.; Ma, R. Water uptake by saltcedar (Tamarix ramosissima) in a desert riparian forest: Responses to intra-annual water table fluctuation. Hydrol. Process. 2015, 30, 1388–1402. [Google Scholar] [CrossRef]
- Wu, H.; Li, J.; Zhang, C.; He, B.; Zhang, H.; Wu, X.; Li, X.-Y. Determining root water uptake of two alpine crops in a rainfed cropland in the Qinghai Lake watershed: First assessment using stable isotopes analysis. Field Crops Res. 2018, 215, 113–121. [Google Scholar] [CrossRef]
- Zhang, H.; Cao, J.; Wang, H.; Song, B.; Jia, G.; Liu, Z.; Yu, X.; Zeng, J. Water utilization characteristics of the degraded poplar shelterbelts in Zhangbei, Hebei, China. Chin. J. Appl. Ecol. 2018, 29, 1381–1388. [Google Scholar] [CrossRef]
- Wenping, D.; Guodong, J.; Yuanqiu, L.; Qi, C.; Jiahui, H.; Linsheng, W.; Zhangling; Xiaojun, L.; Jianbo, J.; Songli, P. Long-term study on the seasonal water uptake of Platycladus orientalis in the Beijing mountain area, northern China. Agric. For. Meteorol. 2021, 307, 108531. [Google Scholar] [CrossRef]
- Su, W.; Jia, D.; Gao, R.; Lu, J.; Lu, F.; Zhao, H.; Wang, F. Water use characteristics of artificial sand-fixing vegetation on the southern edge of Hun-shandake Sandy Land, Inner Mongolia, China. Chin. J. Appl. Ecol. 2021, 32, 1980–1988. [Google Scholar] [CrossRef]
- Yang, L.; Jia, D.; Gao, R.; Su, W.; Lu, F.; Wendu, R. Characteristics of Hydrogen and Oxygen Isotopes in Different Water Bodies of Salix Gordejevii Forest in the South Edgeof Hunshandake Sand Land. Earth Environ. 2022, 50, 630–638. [Google Scholar] [CrossRef]
- Xie, C.; Zhao, L.; Meng, F.; Dong, X.; Liu, Q.; Ma, L. Water sources of plants in the forest ecosystem in the upper reaches ofthe Heihe River Basin. J. Lanzhou Univ. Nat. Sci. 2020, 56, 503–508. [Google Scholar] [CrossRef]
- Elkin, C.; Gutierrez, A.G.; Leuzinger, S.; Manusch, C.; Temperli, C.; Rasche, L.; Bugmann, H. A 2 °C warmer world is not safe for ecosystem services in the European Alps. Glob. Change Biol. 2013, 19, 1827–1840. [Google Scholar] [CrossRef]
- Zeng, N. Tree Mortality Dynamic and the Ecosystem Response in Broad-Leaved Forest in Southern Greater Khingan Mountains. Ph.D. Thesis, Inner Mongolia Agricultural University, Hohhot, China, 2017. [Google Scholar]
- Mao, Q.; Watanabe, M.; Koike, T. Growth characteristics of two promising tree species for afforestation, birch and larch in the northeastern part of Asia. Eurasian J. For. Res. 2010, 13, 76. [Google Scholar]
- Erdene-Ochir, T.; Ishiguri, F.; Nezu, I.; Tumenjargal, B.; Baasan, B.; Chultem, G.; Ohshima, J.; Yokota, S. Utilization potential of naturally regenerated Mongolian Betula platyphylla wood based on growth characteristics and wood properties. Silva Fenn. 2020, 54, 10284. [Google Scholar] [CrossRef]
- Talskih, A.I.; Kopanina, A.V.; Vlasova, I.I. Features of the structural response of the bark and wood of birch (Betula platyphylla, Betulaceae) in the landscapes of sea coasts, magmatic and mud volcanoes of Sakhalin and the Kuril Islands. Geosyst. Transit. Zones 2022, 6, 360–379. [Google Scholar] [CrossRef]
- Zhao, X.; Mao, Z.; Xu, J. Gas Exchange, Chlorophyll and Growth Responses of Betula Platyphylla Seedlings to Elevated CO2 and Nitrogen. Int. J. Biol. 2010, 2, 149. [Google Scholar] [CrossRef]
- Sun, S. Effects of Seasonal Drought on Physiology and Growth of Betula Platyphylla; Northeast Forestry University: Harbin, China, 2020. [Google Scholar]
- Xue, J. Effects of Metabolism and Wood Structure of Betulla Platphilla by Drought Stress; Northeast Forestry University: Harbin, China, 2007. [Google Scholar]
- He, M.; Wei, J.; Shi, L.; Zhou, M.; Zhao, P. The response of radial growth and death of Populus davidiana to regional climate change in southern Greater Khingan Mountains. Chin. J. Ecol. 2018, 37, 3237–3244. [Google Scholar] [CrossRef]
- Wu, Y.; Zhao, P.; Zhou, M.; Liu, Z.; Yao, H.; Wei, J.; Shu, Y.; Li, J.; Xiang, C.; Zhou, L. Environmental Factors Driving the Transpiration of a Betula platyphylla Sukaczev Forest in a Semi-arid Region in North China during Different Hydrological Years. Forests 2022, 13, 1729. [Google Scholar] [CrossRef]
- Wu, H.; Zhang, X.; Xiaoyan, L.; Li, G.; Huang, Y. Seasonal variations of deuterium and oxygen-18 isotopes and their response to moisture source for precipitation events in the subtropical monsoon region. Hydrol. Process. 2015, 29, 90–102. [Google Scholar] [CrossRef]
- Wang, X.; Jia, G.; Deng, W.; Liu, Z.; Liu, Z.; Qiu, G.; Li, W. Long-term water use characteristics and patterns of typical tree species in seasonal drought regions. Chin. J. Appl. Ecol. 2021, 32, 1943–1950. [Google Scholar] [CrossRef]
- Moore, J.W.; Semmens, B.X. Incorporating uncertainty and prior information into stable isotope mixing models. Ecol. Lett. 2008, 11, 470–480. [Google Scholar] [CrossRef]
- Parnell, A.C.; Inger, R.; Bearhop, S.; Jackson, A.L. Source partitioning using stable isotopes: Coping with too much variation. PLoS ONE 2010, 5, e9672. [Google Scholar] [CrossRef] [PubMed]
- Craig, H. Isotopic variations in meteoric waters. Science 1961, 133, 1703. [Google Scholar] [CrossRef]
- Ellsworth, P.Z.; Williams, D.G. Hydrogen isotope fractionation during water uptake by woody xerophytes. Plant Soil 2007, 291, 93–107. [Google Scholar] [CrossRef]
- Hu, J.; Moore, D.J.P.; Burns, S.P.; Monson, R.K. Longer growing seasons lead to less carbon sequestration by a subalpine forest. Glob. Change Biol. 2010, 16, 771–783. [Google Scholar] [CrossRef]
- Yang, A.; Fu, Z.; Wang, L.; Xiao, F.; Wang, L.; Fan, P.; Zhang, J. Strategies on Water Utilization of Poplar in Horqin Sandy Land of Northern China. J. Beijing For. Univ. 2018, 40, 63–72. [Google Scholar] [CrossRef]
- Ma, Y. Study on Hydrological Process of Freshwater Wetland Forests in Tangpu Reservoir Area Based on Stable Hydrogen and Oxygen Isotope Techniques; Chinese Academy of Forestry: Beijing, China, 2019. [Google Scholar]
- Gao, D. Characteristics of Stable Hydrogen and Oxygen Isotopes in the Typical Forest Hydrological Processes of Mt. Dinghu; Chinese Academy of Forestry: Beijing, China, 2017. [Google Scholar]
- Wang, J.; Fu, B.; Lu, N.; Zhang, L. Seasonal variation in water uptake patterns of three plant species based on stable isotopes in the semi-arid Loess Plateau. Sci. Total Environ. 2017, 609, 27–37. [Google Scholar] [CrossRef]
- McDowell, G.N.; Allen, D.C. Darcy’s law predicts widespread forest mortality under climate warming. Nat. Clim. Change 2015, 5, 669–672. [Google Scholar] [CrossRef]




{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Sample Trees | Diameter at Breast Height (cm) | Tree Height (m) |
|---|---|---|
| B1 | 29.5 | 13.9 |
| B2 | 25 | 13 |
| B3 | 29.5 | 14 |
| B4 | 29 | 14.3 |
| B5 | 25.4 | 13.3 |
| Year and Month | P (mm) | 0–5 cm | 10–20 cm | 20–40 cm | 40–80 cm |
|---|---|---|---|---|---|
| 2019 | 390.4 | 0.17 | 0.15 | 0.17 | 0.16 |
| May | 97.9 | 0.27 a | 0.21 a | 0.19 a | 0.15 a |
| June | 78 | 0.23 bd | 0.24 b | 0.27 bcf | 0.17 bc |
| July | 97.9 | 0.29 c | 0.27 c | 0.28 cf | 0.18 bcdf |
| August | 94.8 | 0.26 ad | 0.22 a | 0.24 d | 0.18 df |
| September | 4.6 | 0.17 e | 0.17 d | 0.19 ae | 0.18 bcd |
| 2020 | 501.5 | 0.20 | 0.21 | 0.23 | 0.20 |
| May | 108.7 | 0.28 ac | 0.25 bc | 0.21 e | 0.16 e |
| June | 84.6 | 0.27 ac | 0.27 c | 0.29 f | 0.17 b |
| July | 57.4 | 0.24 bd | 0.25 b | 0.28 cf | 0.18 f |
| August | 120 | 0.22 b | 0.22 ae | 0.26 b | 0.18 df |
| September | 70.4 | 0.28 ac | 0.24 be | 0.27 bc | 0.18 cdf |
| Soil Layers | Soil Depth (cm) | 2019 | 2020 | ||||||
|---|---|---|---|---|---|---|---|---|---|
| July | August | September | May | June | July | August | September | ||
| Shallow | 0–10 cm | 39.4 | 18.4 | 22.8 | 27.9 | 18.3 | 22.1 | 13.2 | 3.7 |
| 10–20 cm | 13.2 | 19 | 19.3 | 14.8 | 16.3 | 16.3 | 17.1 | 6.3 | |
| Middle | 20–30 cm | 13.3 | 17.3 | 16.9 | 14.2 | 16.3 | 14.5 | 18.4 | 10.7 |
| 30–40 cm | 12.2 | 16.1 | 15.2 | 14.3 | 16.5 | 16.5 | 17.4 | 19.4 | |
| Deep | 40–50 cm | 11.7 | 15.3 | 13.1 | 14.5 | 16.7 | 15.6 | 17.8 | 29.3 |
| 50–60 cm | 10.2 | 13.9 | 12.6 | 14.3 | 15.9 | 15 | 16.1 | 30.6 | |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2024 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Zhao, P.; Wu, Y.; Yao, H.; Li, Y.; Liu, B.; Zhou, M.; Shu, Y.; Zhou, L.; Xiang, C. Determining Water Use Patterns for the Betula platyphylla in the Southern Greater Khingan Mountains and Its Adaptation to Drought. Water 2024, 16, 927. https://doi.org/10.3390/w16070927
Zhao P, Wu Y, Yao H, Li Y, Liu B, Zhou M, Shu Y, Zhou L, Xiang C. Determining Water Use Patterns for the Betula platyphylla in the Southern Greater Khingan Mountains and Its Adaptation to Drought. Water. 2024; 16(7):927. https://doi.org/10.3390/w16070927
Chicago/Turabian StyleZhao, Pengwu, Yiheng Wu, Huaxia Yao, Yuanqiao Li, Bingbing Liu, Mei Zhou, Yang Shu, Liwen Zhou, and Changlin Xiang. 2024. "Determining Water Use Patterns for the Betula platyphylla in the Southern Greater Khingan Mountains and Its Adaptation to Drought" Water 16, no. 7: 927. https://doi.org/10.3390/w16070927
APA StyleZhao, P., Wu, Y., Yao, H., Li, Y., Liu, B., Zhou, M., Shu, Y., Zhou, L., & Xiang, C. (2024). Determining Water Use Patterns for the Betula platyphylla in the Southern Greater Khingan Mountains and Its Adaptation to Drought. Water, 16(7), 927. https://doi.org/10.3390/w16070927