Diversity and Taxonomy of Chrysophytes from the Pasvik State Nature Reserve (Russia)


Abstract
1. Introduction
2. Materials and Methods
- (1)
- The Paz River (N 69°12′46.8″, E 29°18′20.6″); 7 October 2017: environmental variables were not measured;
- (2)
- The Paz River (N 69°00′01.2″, E 28°56′32.0″); 19 June 2019: pH 3.1, temperature 16.5, conductivity 25 µS·cm−1;
- (3)
- A peat bog on the bank of the Paz River (N 69°23.489′, E 29°45.388′); 19 June 2019: environmental variables were not measured.
3. Results

{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Taxon | Site Number | ||
|---|---|---|---|
| 1 | 2 | 3 | |
| Paraphysomonadales | |||
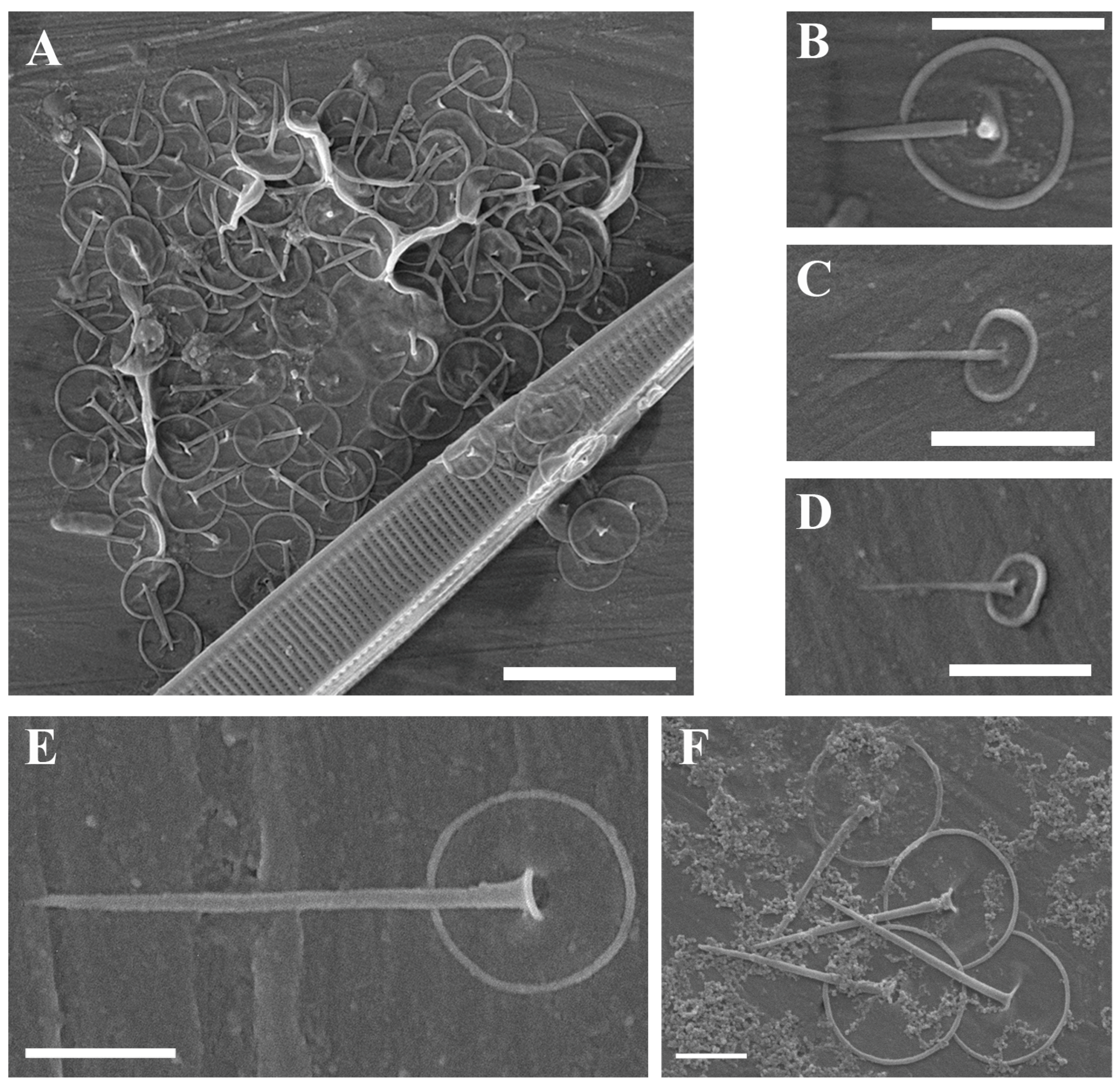
| Paraphysomonas aff. vulgaris brevispina Scoble and Cavalier-Smith (Figure 1A,B) | + | + | |
| Paraphysomonas aff. ovalis Scoble and Cavalier-Smith (Figure 1C,D) | + | ||
| Paraphysomonas cf. acuminata Scoble and Cavalier-Smith (Figure 1E) | + | ||
| Paraphysomonas vulgaris vulgaris Scoble and Cavalier-Smith (Figure 1F) | + | ||
| Synurales | |||
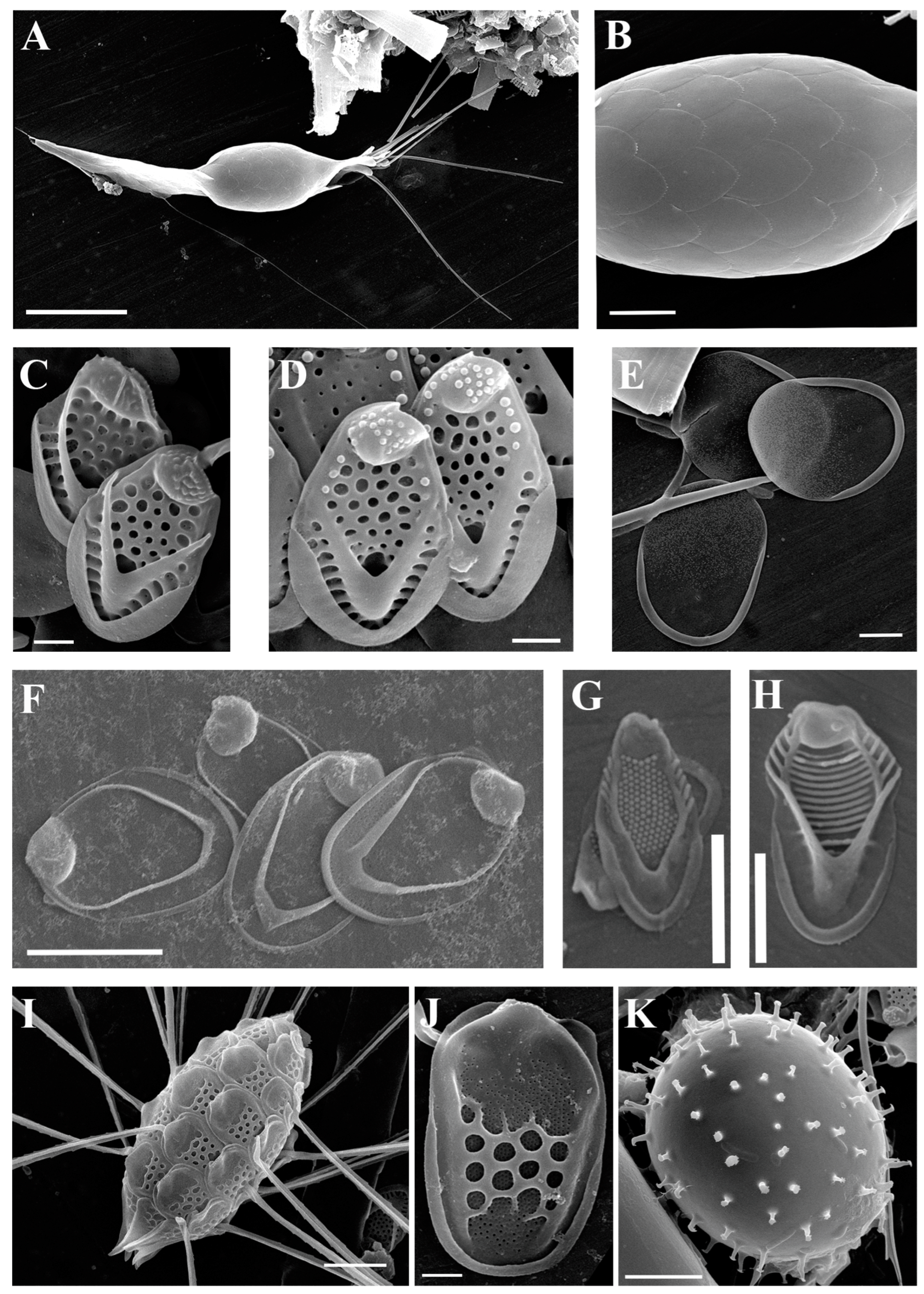
| Mallomonas akrokomos Ruttner (Figure 2A,B) | + | ||
| Mallomonas crassisquama (Asmund) Fott var. crassisquama (Figure 2C) | + | + | |
| Mallomonas crassisquama var. papillosa Siver and Skogstad(Figure 2D) | + | ||
| Mallomonas caudata Iwanoff emend. W. Krieger (Figure 2E) | + | ||
| Mallomonas elongata Reverdin (Figure 2F) | + | + | |
| Mallomonas papillosa K. Harris and D.E. Bradley (Figure 2G) | + | ||
| Mallomonas punctifera Korshikov (Figure 2I,J) | + | ||
| Mallomonas striata Asmund (Figure 2H) | + | ||
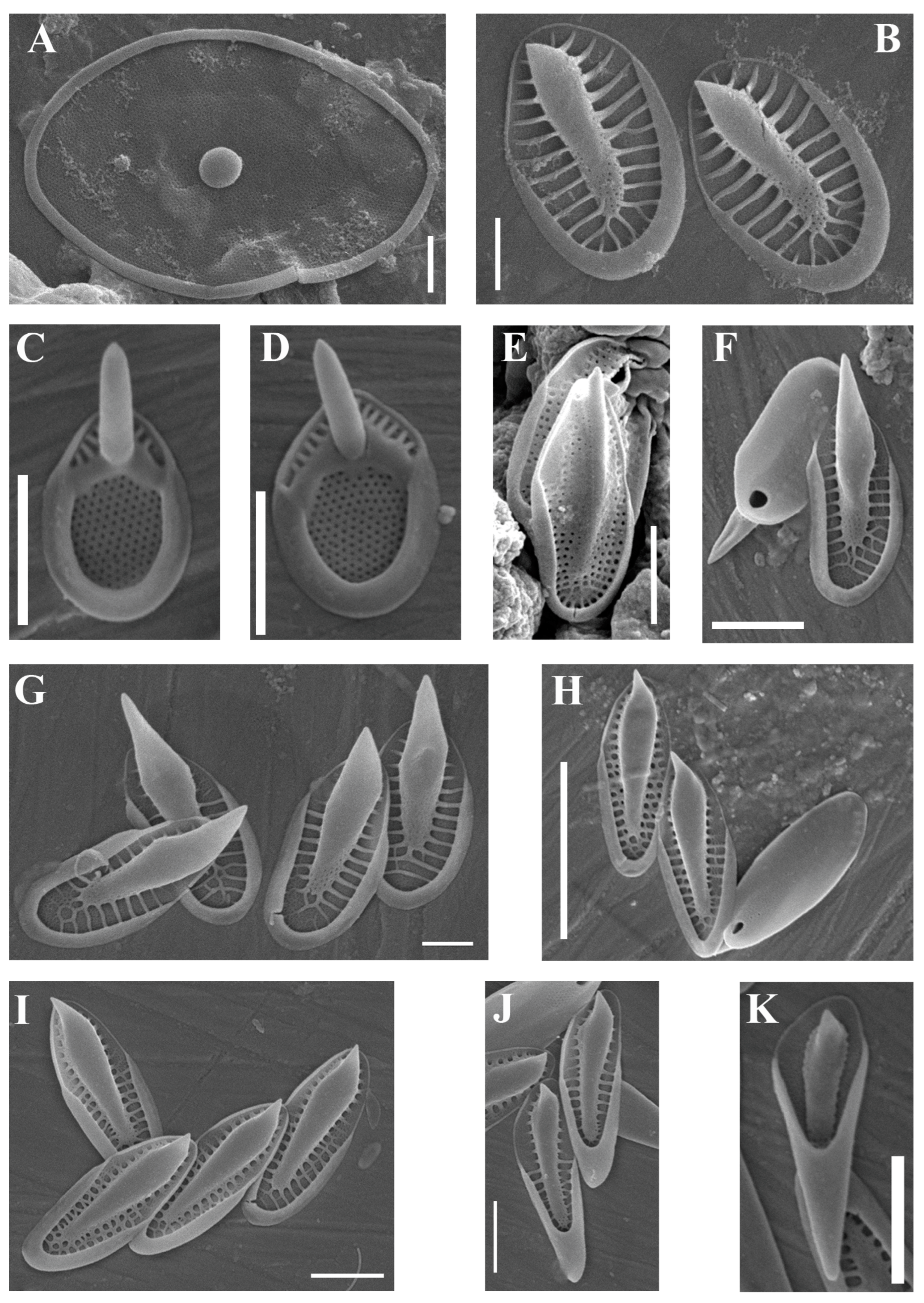
| Neotessella lapponica (Skuja) Jo et al. (Figure 3A) | + | ||
| Synura americana Kynčlová and Škaloud (Figure 3B) | + | ||
| Synura echinulata Korshikov (Figure 3C,D) | + | ||
| Synura macracantha (J.B. Petersen and J.B. Hansen) Asmund (Figure 3E) | + | ||
| Synura skaloudiorum Kapustin sp. nov. (Figure 3F–K) | + | ||
| Chromulinales | |||
| Chrysococcus biporus Skuja | + | ||
| Chrysococcus furcatus K.H. Nicholls | + | ||
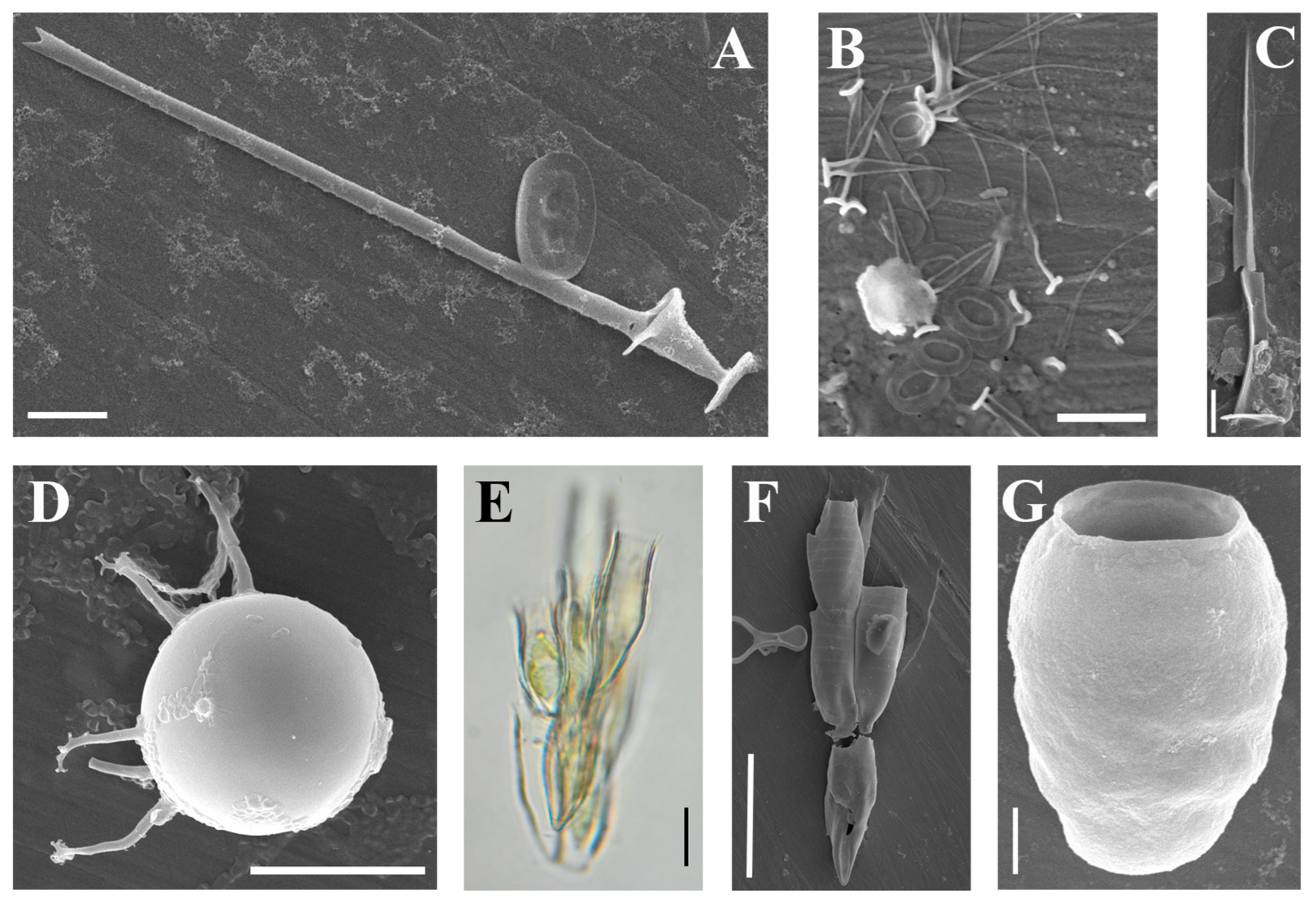
| Chrysosphaerella longispina Lauterborn (Figure 4A) | + | ||
| Chrysosphaerella septentrionalis Kapustin | + | ||
| Lepochromulina calyx Scherffel | + | ||
| Spiniferomonas abei E. Takahashi (Figure 4B) | + | ||
| Spiniferomonas cf. trioralis E. Takahashi (Figure 4C) | + | + | |
| Chrysosaccales | |||
| Chrysastrella paradoxa Chodat (Figure 4D) | + | ||
| Chrysopyxis bipes F. Stein | + | ||
| Chrysothecopsis tubulosus (Pascher) Kapustin | + | ||
| Ochromonadales | |||
| Dinobryon anulatum D.K. Hilliard and Asmund (Figure 4E,F) | + | ||
| Dinobryon bavaricum O.E. Imhof | + | ||
| Dinobryon divergens O.E. Imhof | + | + | |
| Dinobryon sertularia Ehrenberg | + | ||
| Epipyxis utriculus (Ehrenberg) Ehrenberg | + | ||
| Kephyrion starmachii (Czosnowski) Bourrelly (Figure 4G) | + | ||
| Uroglenopsis botrys (Pascher) Pascher | + | ||
3.1. Nomenclatural and Taxonomic Comments on the Selected Taxa
3.1.1. The Correct Authorship of Chrysococcus furcatus
3.1.2. Validation of Kephyrion starmachii
3.1.3. A New Combination in the Genus Chrysothecopsis Conrad
3.1.4. Mallomonas dubia vs. Mallomonas caudata
3.1.5. New Synonyms of Dinobryon annulatum
4. Discussion
4.1. Chrysophyte Flora of the Pasvik State Nature Reserve with Comments on the Selected Taxa
4.2. Morphological Comparison of Synura skaloudiorum with Similar Taxa
5. Conclusions
Funding
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Bock, C.; Olefeld, J.L.; Vogt, J.C.; Albach, D.C.; Boenigk, J. Phylogenetic and functional diversity of Chrysophyceae in inland waters. Org. Divers. Evol. 2022, 22, 327–341. [Google Scholar] [CrossRef]
- Guiry, M.D. How many species of algae are there? A reprise. Four kingdoms, 14 phyla, 63 classes and still growing. J. Phycol. 2024, 60, 214–228. [Google Scholar] [CrossRef]
- Kristiansen, J.; Škaloud, P. Chrysophyta. In Handbook of the Protists, 2nd ed.; Archibald, J.M., Simpson, A.G.B., Slamovits, C.H., Eds.; Springer International Publishing AG: Cham, Switzerland, 2017; pp. 331–366. [Google Scholar] [CrossRef]
- Lengyel, E.; Barreto, S.; Padisák, J.; Stenger-Kovács, C.; Lázár, D.; Buczkó, K. Contribution of silica-scaled chrysophytes to ecosystems services: A review. Hydrobiologia 2023, 850, 2735–2756. [Google Scholar] [CrossRef]
- Charvet, S.; Vincent, W.F.; Lovejoy, C. Chrysophytes and other protists in High Arctic lakes: Molecular gene surveys, pigment signatures and microscopy. Polar Biol. 2012, 35, 733–748. [Google Scholar] [CrossRef]
- Boopathi, T.; Faria, D.G.; Lee, M.-D.; Lee, J.; Chang, M.; Ki, J.-S. A molecular survey of freshwater microeukaryotes in an Arctic reservoir (Svalbard, 79° N) in summer by using next-generation sequencing. Polar Biol. 2015, 38, 179–187. [Google Scholar] [CrossRef]
- Izaguirre, I.; Unrein, F.; Schiaffino, M.R.; Lara, E.; Singer, D.; Balagué, V.; Gasol, J.M.; Massana, R. Phylogenetic diversity and dominant ecological traits of freshwater Antarctic Chrysophyceae. Polar Biol. 2021, 44, 941–957. [Google Scholar] [CrossRef]
- Gusev, E.S.; Guseva, E.E.; Gabyshev, V.A. Taxonomic composition of silica-scaled chrysophytes in rivers and lakes of Yakutia and Magadanskaya oblast (Russia). Nova Hedw. Beih. 2018, 147, 105–117. [Google Scholar] [CrossRef]
- Bessudova, A.; Gabyshev, V.; Firsova, A.; Gabysheva, O.; Bukin, Y.; Likhoshway, Y. Diversity Variation of Silica-Scaled Chrysophytes Related to Differences in Physicochemical Variables in Estuaries of Rivers in an Arctic Watershed. Sustainability 2021, 13, 13768. [Google Scholar] [CrossRef]
- Bessudova, A.; Gabyshev, V.; Bukin, Y.; Gabysheva, O.; Likhoshway, Y.V. Species richness of scaled Chrysophytes in arctic waters in the Tiksi Region (Yakutia, Russia). Acta Biol. Sib. 2022, 8, 431–459. [Google Scholar] [CrossRef]
- Bessudova, A.; Firsova, A.; Bukin, Y.; Kopyrina, L.; Zakharova, Y.; Likhoshway, Y. Under-Ice Development of Silica-Scaled Chrysophytes with Different Trophic Mode in Two Ultraoligotrophic Lakes of Yakutia. Diversity 2023, 15, 326. [Google Scholar] [CrossRef]
- Bessudova, A.; Gabyshev, V.; Likhoshway, Y. Record of two rare taxa from Synura genus (Chrysophyceae) with a description of a new species (Synura tiksiensis sp. nov.) near the arctic settlement of Tiksi, Yakutia, Russia. Phytotaxa 2022, 560, 247–253. [Google Scholar] [CrossRef]
- Sharov, A.N. Phytoplankton from the Lakes of Kola Peninsula; Karelian Research Center RAS: Petrozavodsk, Russia, 2004; pp. 1–98. (In Russian) [Google Scholar]
- Kapustin, D.A.; Kapustina, N.V. New records of Chrysococcus furcatus (Chrysophyceae) in Russia. Inland Wat. Biol. 2018, 11, 384–386. [Google Scholar] [CrossRef]
- Kapustin, D.; Kulikovskiy, M. Chrysosphaerella septentrionalis sp. nov. (Chrysophyceae, Chromulinales), a New Species from the Arctic including the Description of Chrysosphaerellaceae, fam. nov. Plants 2022, 11, 3166. [Google Scholar] [CrossRef]
- Borovichev, E.A.; Boychuk, M.A. Bryophytes of the Pasvik State Nature Reserve; Karelian Research Center RAS: Petrozavodsk, Russia, 2018; pp. 1–123. (In Russian) [Google Scholar]
- Scoble, J.M.; Cavalier-Smith, T. Scale evolution in Paraphysomonadida (Chrysophyceae): Sequence phylogeny and revised taxonomy of Paraphysomonas, new genus Clathromonas, and 25 new species. Eur. J. Protistol. 2014, 50, 551–592. [Google Scholar] [CrossRef]
- Bessudova, A.; Likhoshway, Y. Scaled chrysophytes (Chrysophyceae) of the Boguchany Reservoir. Sovrem. Nauka Aktual. Probl. Teor. Praktiki. Ser. Estestv. Tekhnicheskiye Nauk. 2017, 11, 4–11. (In Russian) [Google Scholar]
- Prokina, K.I.; Philippov, D.A. Heterotrophic Flagellates from Mires of the North Caucasus, Russia. Inland Water Biol. 2021, 14, 500–516. [Google Scholar] [CrossRef]
- Kuzmin, G.V.; Kuzmina, V.A. Armored representatives of the golden algae from the Magadan Region. Novosti Sistem. Nizsh. Rast. 1987, 24, 40–42. (In Russian) [Google Scholar]
- Dolgoff, G.I. Zur Systematik von Trachelomonas Ehrb. Russ. Hydrobiol. Zeit. 1922, 1, 289–291. (In Russian) [Google Scholar]
- Woronichin, N.N. Notiz über Chrysomonadencysten in Plankton der Flüsse Newa und Boljschaja Newka. Acta Inst. Bot. Acad. Sci. USSR 1933, 1, 7. (In Russian) [Google Scholar]
- Deflandre, P. Sur l’abus de l’emploi en paléontologie du nom de genre Trachelomonas. Ann. Protistol. 1934, 4, 151–165. [Google Scholar]
- Nicholls, K.H. Chrysococcus furcatus (Dolg.) comb. nov.: A new name for Chrysastrella furcata (Dolg.) Defl. based on the discovery of the vegetative stage. Phycologia 1981, 20, 16–21. [Google Scholar] [CrossRef]
- Turland, N.J.; Wiersema, J.H.; Barrie, F.R.; Greuter, W.; Hawksworth, D.L.; Herendeen, P.S.; Knapp, S.; Kusber, W.-H.; Li, D.-Z.; Marhold, K.; et al. (Eds.) International Code of Nomenclature for Algae, Fungi, and Plants (Shenzhen Code); Adopted by the Nineteenth International Botanical Congress Shenzhen, China, July 2017; Koeltz Botanical Books: Glashütten, Germany, 2018; 254p, (Regnum Vegetabile 159). [Google Scholar] [CrossRef]
- Czosnowski, J. Materiały do flory wiciowców Polski. Poznań. Towarz. Przyj. Nauk Wydz. Mat.-Przyr. Prace Kom. Biol. 1948, 11, 363–402. [Google Scholar]
- Bourrelly, P. Recherches sur les Chrysophycées. Morphologie, Phylogénie, Systématique. Rev. Algol. Mém. Hors-Sér. 1957, 1, 1–412. [Google Scholar]
- Ellis-Adam, A.C. Some new and interesting benthic Chrysophyceae from a Dutch moorland pool complex. Acta Bot. Neerl. 1983, 32, 1–23. [Google Scholar] [CrossRef]
- Kapustin, D.A.; Kulikovskiy, M.S. (2743) Proposal to conserve the name Mallomonas caudata against M. fastigata (Chrysophyceae: Mallomonadaceae). Taxon 2020, 69, 614–615. [Google Scholar] [CrossRef]
- Andersen, R.A. Report of the Nomenclature Committee for Algae 24. Taxon 2023, 72, 652–653. [Google Scholar] [CrossRef]
- Hilliard, D.K.; Asmund, B. Studies on Chrysophyceae from some ponds and lakes in Alaska. II. Notes on the genera Dinobryon, Hyalobryon and Epipyxis with descriptions of new species. Hydrobiologia 1963, 22, 331–397. [Google Scholar] [CrossRef]
- Skuja, H. Grundzüge der Algenflora und Algenvegetation der Fjeldgegenden um Abisko in Schwedisch-Lappland. Nova Acta Reg. Soc. Sci. Upsal. 1964, 18, 465. [Google Scholar]
- Shi, Z. Some new taxa of Euglenophyta from Xizang (Tibet). Acta Phytotax. Sin. 1984, 22, 337–342. [Google Scholar]
- Pang, W.; Zhuang, J.; Wang, Q. Chrysophytes from the Great Xing’an Mountains, China. Nova Hedw. Beih. 2019, 148, 49–61. [Google Scholar] [CrossRef]
- Kristiansen, J. Flagellates from Finnish Lappland. Bot. Tidsskr. 1964, 59, 315–333. [Google Scholar]
- Němcová, Y.; Pichrtová, M. The rare species Synura lapponica (Synurophyceae) new to the Czech Republic, local vs. global diversity in colonial synurophytes. Biologia 2009, 64, 1070–1075. [Google Scholar] [CrossRef]
- Balonov, I.M.; Kuzmin, G.V. Species of the genus Synura Ehr. (Chrysophyta) in water reservoirs of the Volga cascade. Bot. Zhurn. 1974, 59, 1675–1686. (In Russian) [Google Scholar]
- Balonov, I.M. Genus Synura Ehr. (Chrysophyta). Biology, ecology and systematics. In Biologia, Morphologia i Sistematika Vodnykh Organizmov; Kamshilov, M.M., Ed.; Nauka: Leningrad, Russia, 1976; pp. 61–81. (In Russian) [Google Scholar]
- Kapustin, D.A. An updated checklist of the silica-scaled chrysophytes from Russia. Phytotaxa 2024, in press. [Google Scholar]
- Yermolaev, V.I.; Safonova, T.A. Algae of the genus Dinobryon Ehr. (Chrysophyta) from the water-bodies of Taimyr. Bot. Zhurn. 1974, 59, 556–560. [Google Scholar]
- Siver, P.A.; Skogstad, A. Morphological variation and ecology of Mallomonas crassisquama (Chrysophyceae). Nord. J. Bot. 1988, 7, 99–107. [Google Scholar] [CrossRef]
- Gretz, M.R.; Sommerfeld, M.R.; Wujek, D.E. Scaled Chrysophyceae of Arizona: A Preliminary Survey. J. Arizona-Nevada Acad. Sci. 1979, 14, 75–80. [Google Scholar]
- Voloshko, L.N. Chrysophycean Algae in Water Bodies of the Northern Russia; Renome: St. Petersburg, Russia, 2017; pp. 1–380. (In Russian) [Google Scholar]
- Siver, P.A.; Voloshko, L.N.; Gavrilova, O.V.; Getsen, M.V. The scaled chrysophyte flora of the Bolshezemelskaya tundra (Russia). Nova Hedw. Beih. 2005, 128, 125–150. [Google Scholar]
- Snitko, L.V.; Safronova, T.V.; Snitko, V.P. Chrysophycean algae (Chrysophyceae) in waterbodies of the South Urals and Transural Plateau. Genus Synura (Synuraceae) section Petersenianae. Bot. Zhurn. 2022, 107, 333–349. [Google Scholar]
- Kapustin, D. New and rare species of the loricate golden algae (Chrysophyceae) for the Ukrainian flora from the Polessian Nature Reserve. Novosti Sist. Nizsh. Rast. 2015, 49, 32–46. [Google Scholar] [CrossRef]
- Andersen, R.A. Lepochromulina bursa Scherffel (Chrysophyceae) from Michigan, with ultrastructural observations on the flagellar apparatus and lorica. Nova Hedw. Beih. 2021, 151, 9–26. [Google Scholar] [CrossRef]
- Wislouch, S. Sur les Chrysomonadines des environs de Petrograd. Zhurn. Mikrobiol. 1914, 1, 251–278. [Google Scholar]
- Kulizin, P.V.; Martynenko, N.A.; Gusev, E.S.; Kapustin, D.A.; Vodeneeva, E.L.; Kulikovskiy, M.S. New Species of the Genus Cryptomonas (Cryptophyceae) in the Flora of Russia. Inl. Wat. Biol. 2022, 15, 227–237. [Google Scholar] [CrossRef]
- Andersen, R.A.; Graf, L.; Malakhov, Y.; Yoon, H.S. Rediscovery of the Ochromonas type species Ochromonas triangulata (Chrysophyceae) from its type locality (Lake Veysove, Donetsk region, Ukraine). Phycologia 2017, 56, 591–604. [Google Scholar] [CrossRef]
- Kapustin, D.; Sterlyagova, I.; Patova, E. Morphology of Chrysastrella paradoxa stomatocysts from the Subpolar Urals (Russia) with comments on related morphotypes. Phytotaxa 2019, 402, 295–300. [Google Scholar] [CrossRef]
- Naumenko, Y.V.; Gidora, O.Y. The genus Kephyrion Pascher in the basin of the Sabun River (Western Siberia, Russia). Contemp. Probl. Ecol. 2014, 7, 569–573. [Google Scholar] [CrossRef]
- Němcová, Y.; Nováková, S.; Řezáčová-Škaloudová, M. Synura obesa sp. nov. (Synurophyceae) and other silica-scaled chrysophytes from Abisko (Swedish Lapland). Nova Hedw. 2008, 86, 243–254. [Google Scholar] [CrossRef]
- Siver, P.A.; Lott, A.M. Descriptions of two new species of Synurophyceae from a bog in Newfoundland, Canada: Mallomonas baskettii sp. nov. and Synura kristiansenii sp. nov. Nova Hedw. 2016, 102, 501–511. [Google Scholar] [CrossRef]
- Škaloud, P.; Kynčlová, A.; Benada, O.; Kofroňová, O.; Škaloudová, M. Toward a revision of the genus Synura, section Petersenianae (Synurophyceae, Heterokontophyta): Morphological characterization of six pseudocryptic species. Phycologia 2012, 51, 303–329. [Google Scholar] [CrossRef]
- Škaloud, P.; Škaloudová, M.; Procházková, A.; Němcová, Y. Morphological delineation and distribution patterns of four newly described species within the Synura petersenii species complex (Chrysophyceae, Stramenopiles). Eur. J. Phycol. 2014, 49, 213–229. [Google Scholar] [CrossRef]
- Jo, B.Y.; Kim, J.I.; Škaloud, P.; Siver, P.A.; Shin, W. Multigene phylogeny of Synura (Synurophyceae) and descriptions of four new species based on morphological and DNA evidence. Eur. J. Phycol. 2016, 51, 413–430. [Google Scholar] [CrossRef]
- Škaloud, P.; Škaloudová, M.; Jadrná, I.; Bestová, H.; Pusztai, M.; Kapustin, D.; Siver, P.A. Comparing morphological and molecular estimates of species diversity in the freshwater genus Synura (Stramenopiles): A model for understanding diversity of eukaryotic microorganisms. J. Phycol. 2020, 56, 574–591. [Google Scholar] [CrossRef] [PubMed]
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2024 by the author. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Kapustin, D. Diversity and Taxonomy of Chrysophytes from the Pasvik State Nature Reserve (Russia). Water 2024, 16, 2990. https://doi.org/10.3390/w16202990
Kapustin D. Diversity and Taxonomy of Chrysophytes from the Pasvik State Nature Reserve (Russia). Water. 2024; 16(20):2990. https://doi.org/10.3390/w16202990
Chicago/Turabian StyleKapustin, Dmitry. 2024. "Diversity and Taxonomy of Chrysophytes from the Pasvik State Nature Reserve (Russia)" Water 16, no. 20: 2990. https://doi.org/10.3390/w16202990
APA StyleKapustin, D. (2024). Diversity and Taxonomy of Chrysophytes from the Pasvik State Nature Reserve (Russia). Water, 16(20), 2990. https://doi.org/10.3390/w16202990