
Green Infrastructure Microbial Community Response to Simulated Pulse Precipitation Events in the Semi-Arid Western United States

 ,
, 
Abstract
1. Introduction
- Do pulses of water, equal in magnitude to moderate- to high-intensity storm events, stimulate SGI microbial community growth and resource acquisition?
- Do pulsed water additions within SGI induce changes to soil N pools and fluxes?
- To what extent are microbial physiology and soil N dynamics controlled by plant diversity vs. ecological stoichiometry?
2. Materials and Methods
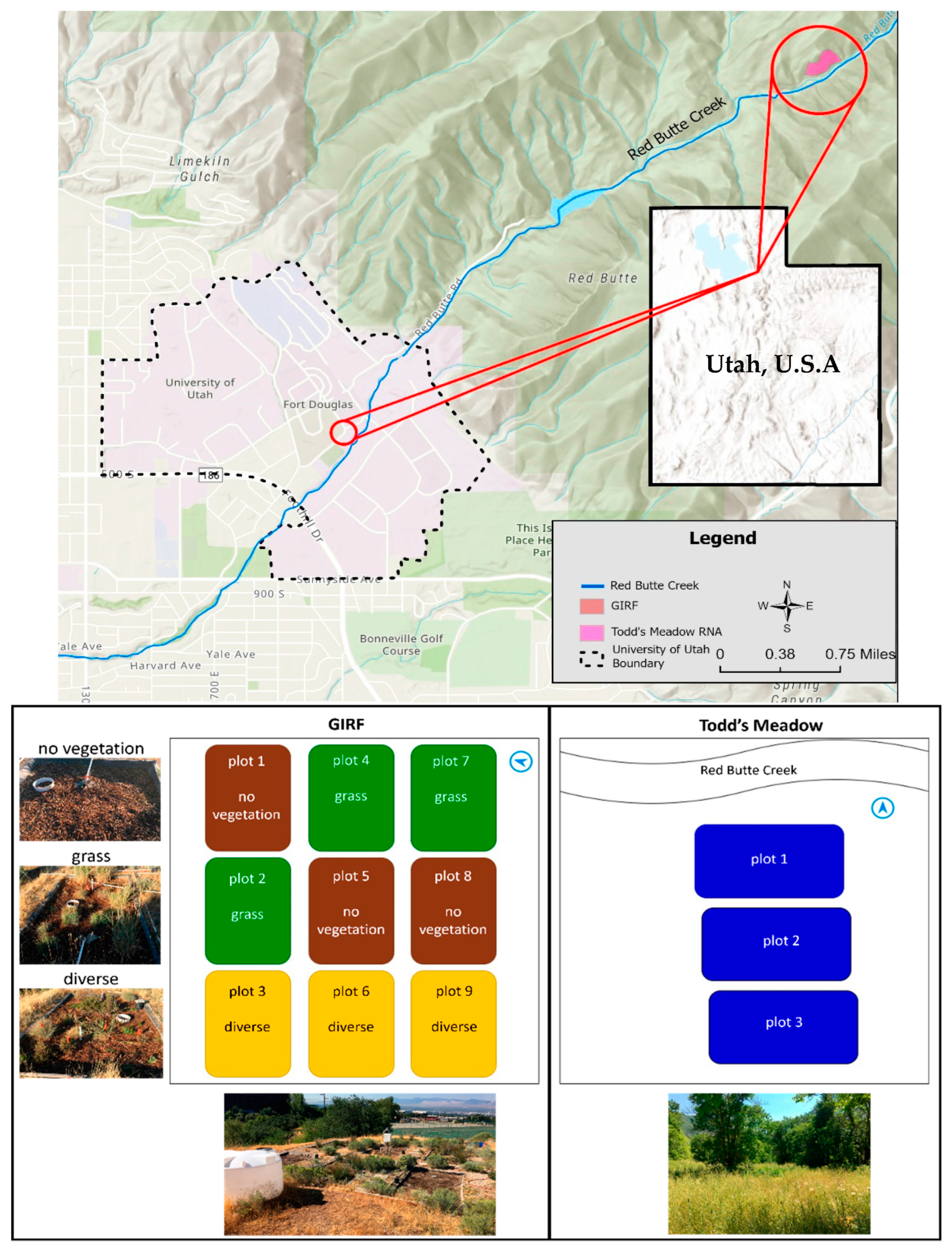
2.1. Site Descriptions
2.1.1. Green Infrastructure Research Facility (GIRF)
2.1.2. Research Natural Area (RNA)—Todd’s Meadow
2.2. Local Climate during Study Period
2.3. Conceptual Model
2.4. Field Sampling and Processing
2.4.1. Stormwater Experimental and Natural Pulses
2.4.2. Soil Sampling Technique and Frequency
2.4.3. Lab Processing
2.5. Analytical Assays
2.5.1. Gravimetric Soil Moisture, pH, Organic Matter Content, and N Pools and Fluxes
2.5.2. Biomass Quantification
2.5.3. Ecoenzyme Activity
2.5.4. Proteolytic Rate
2.6. Mathematical and Statistical Analyses
2.6.1. Two-Way Repeated Measures Analysis of Variance
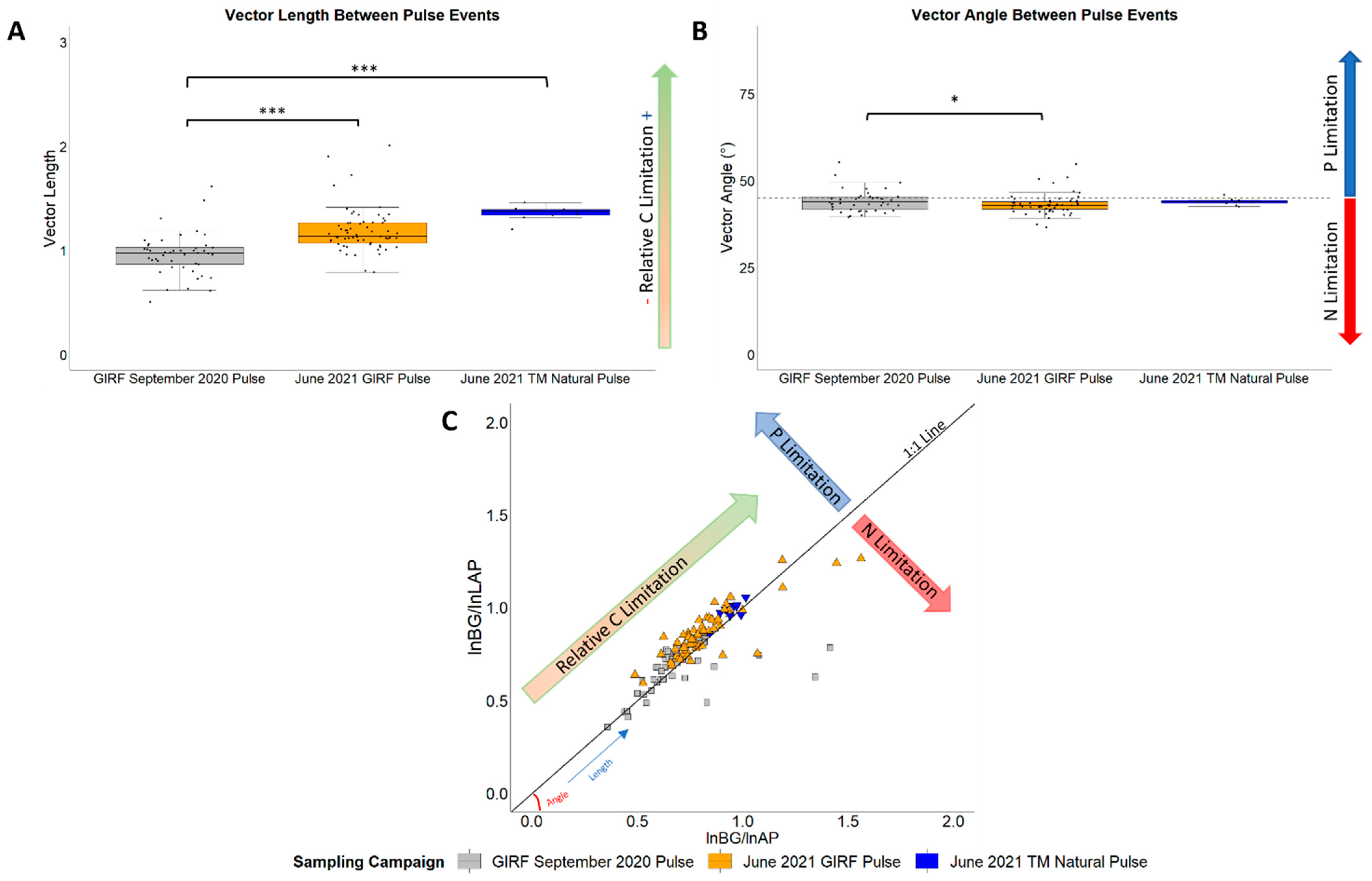
2.6.2. Vector Analysis
2.6.3. Pearson Correlation
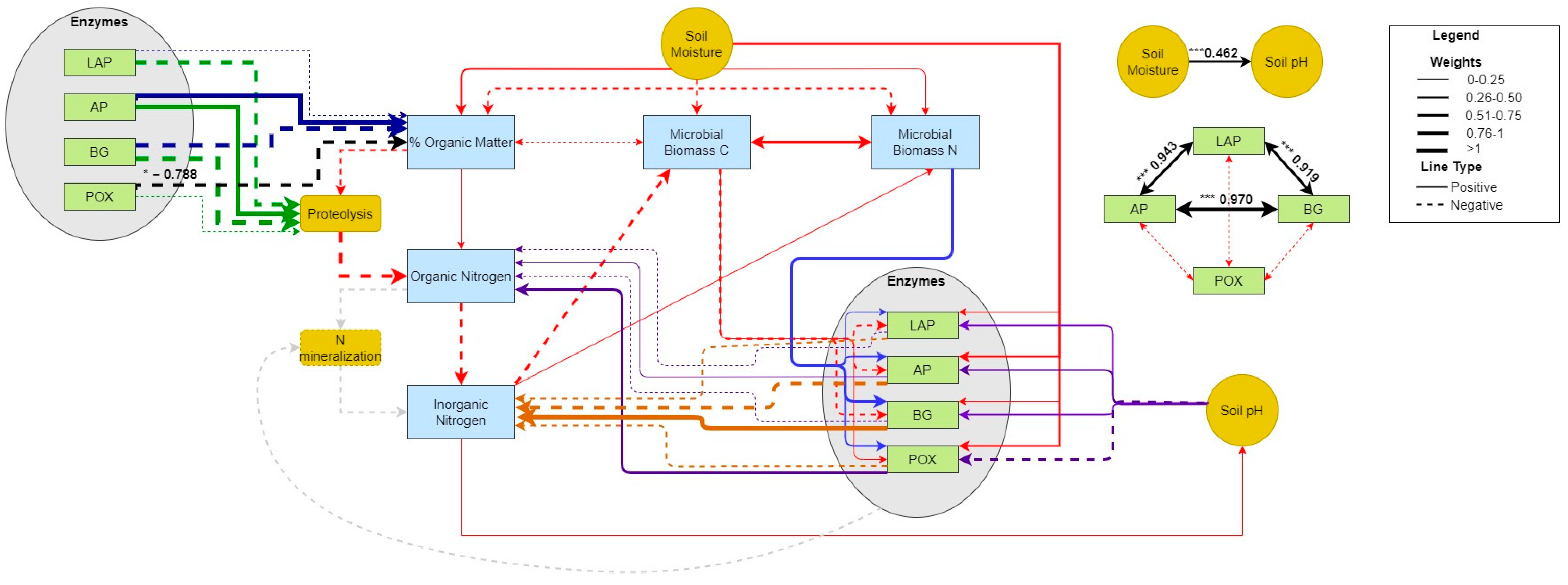
2.6.4. Piecewise Structural Equation Modeling
3. Results
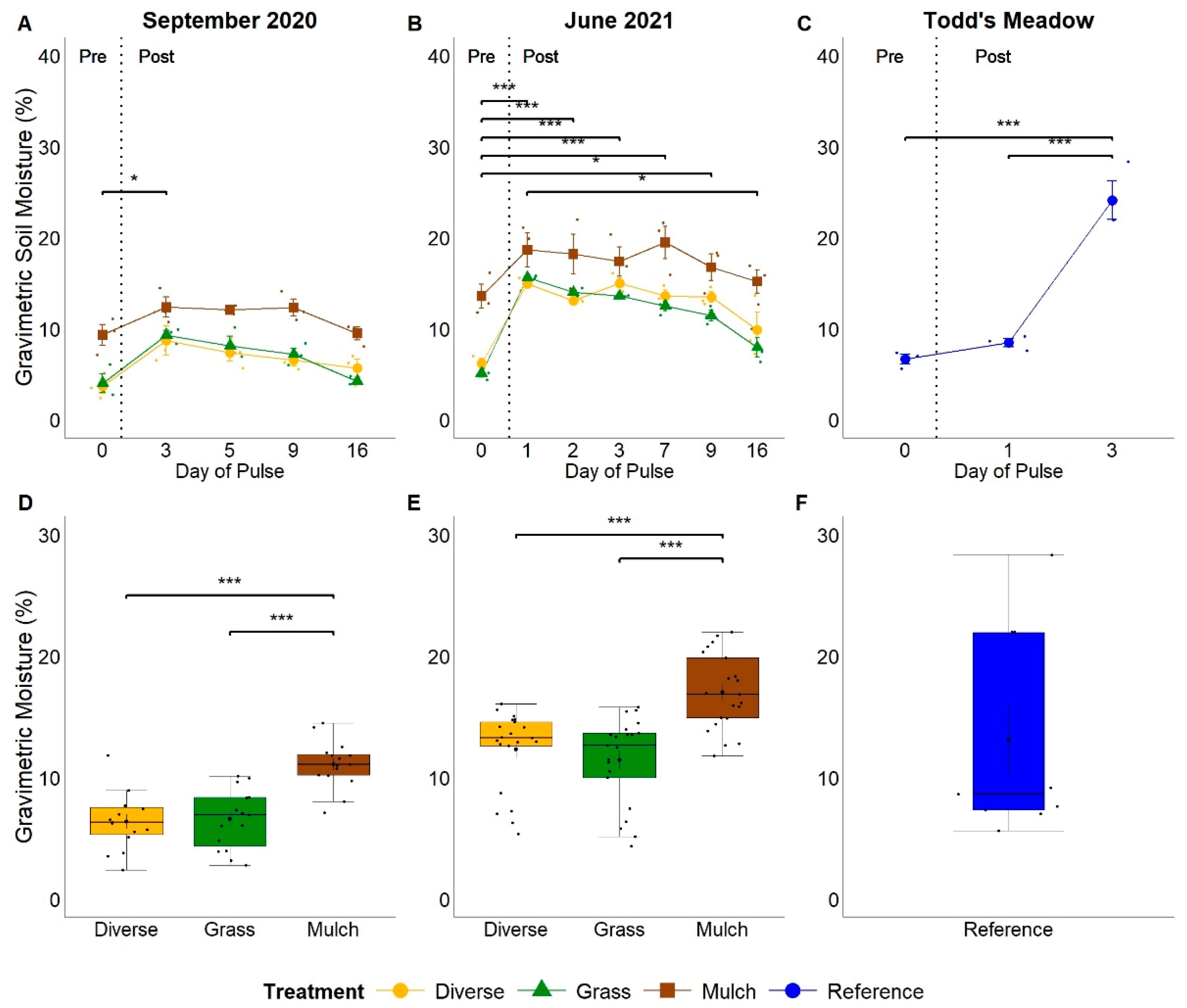
3.1. Gravimetric Soil Moisture and pH
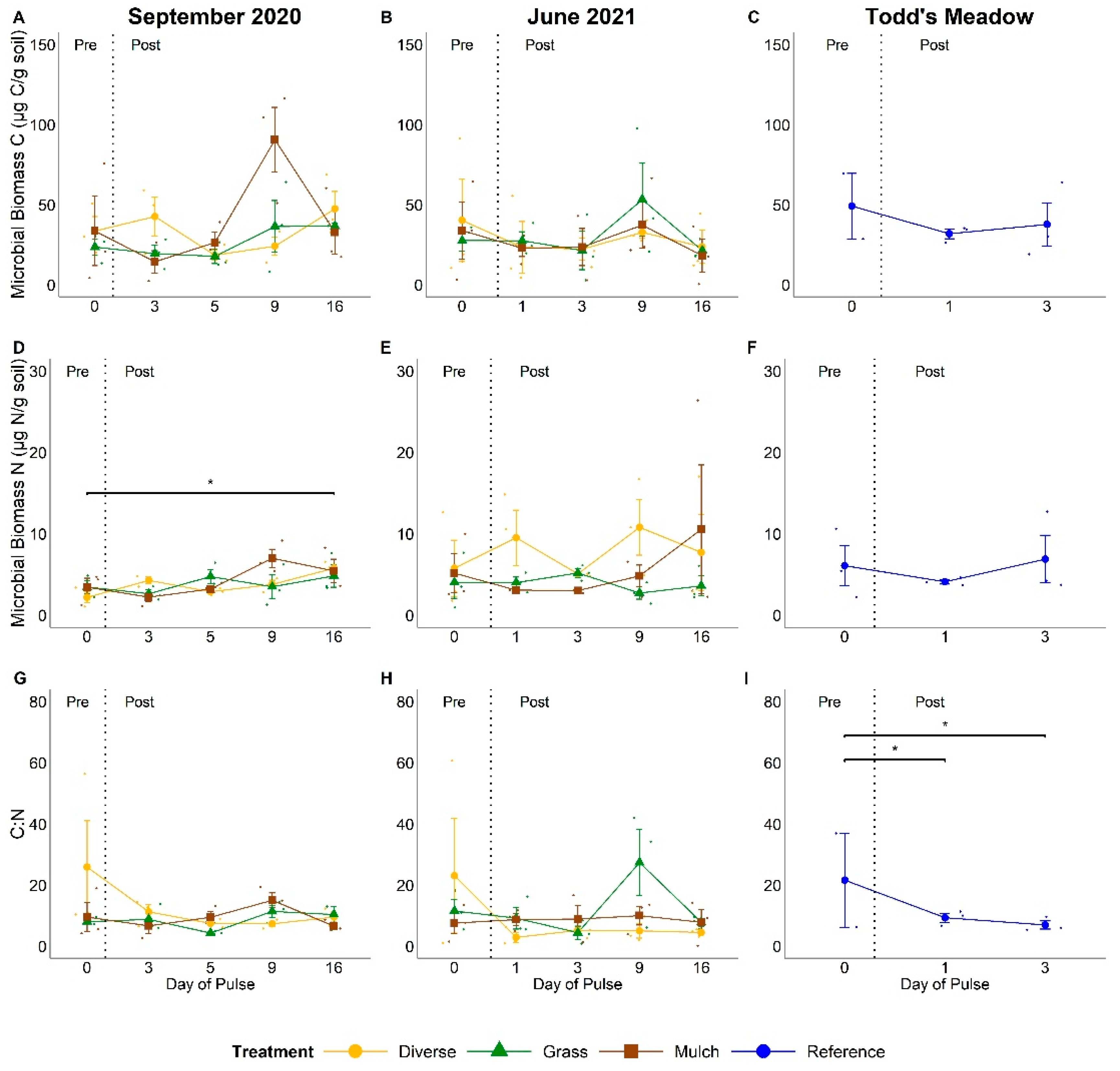
3.2. Microbial Biomass C and N Content
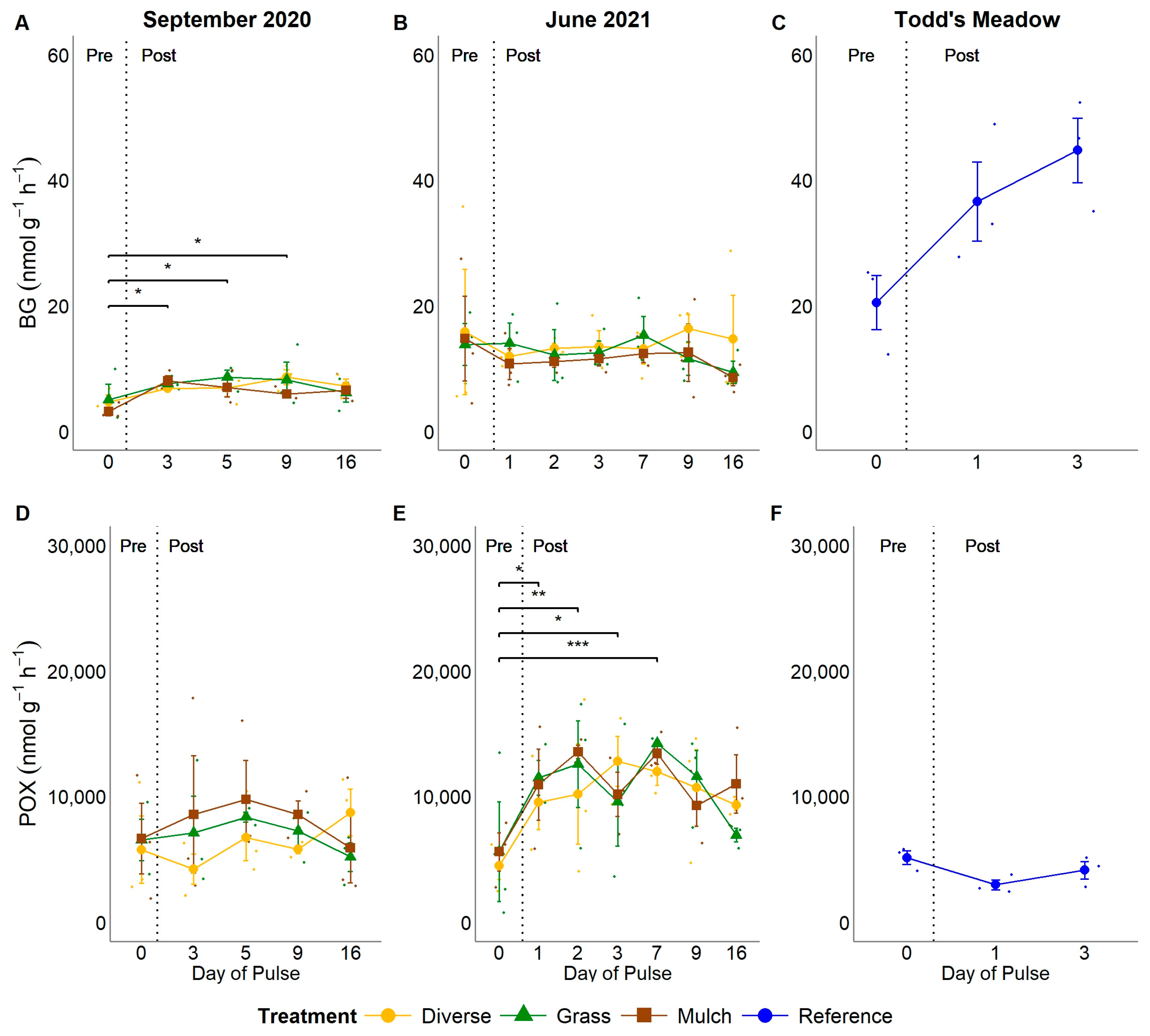
3.3. Ecoenzyme Activity Rates, Ratios, and Vector Analyses
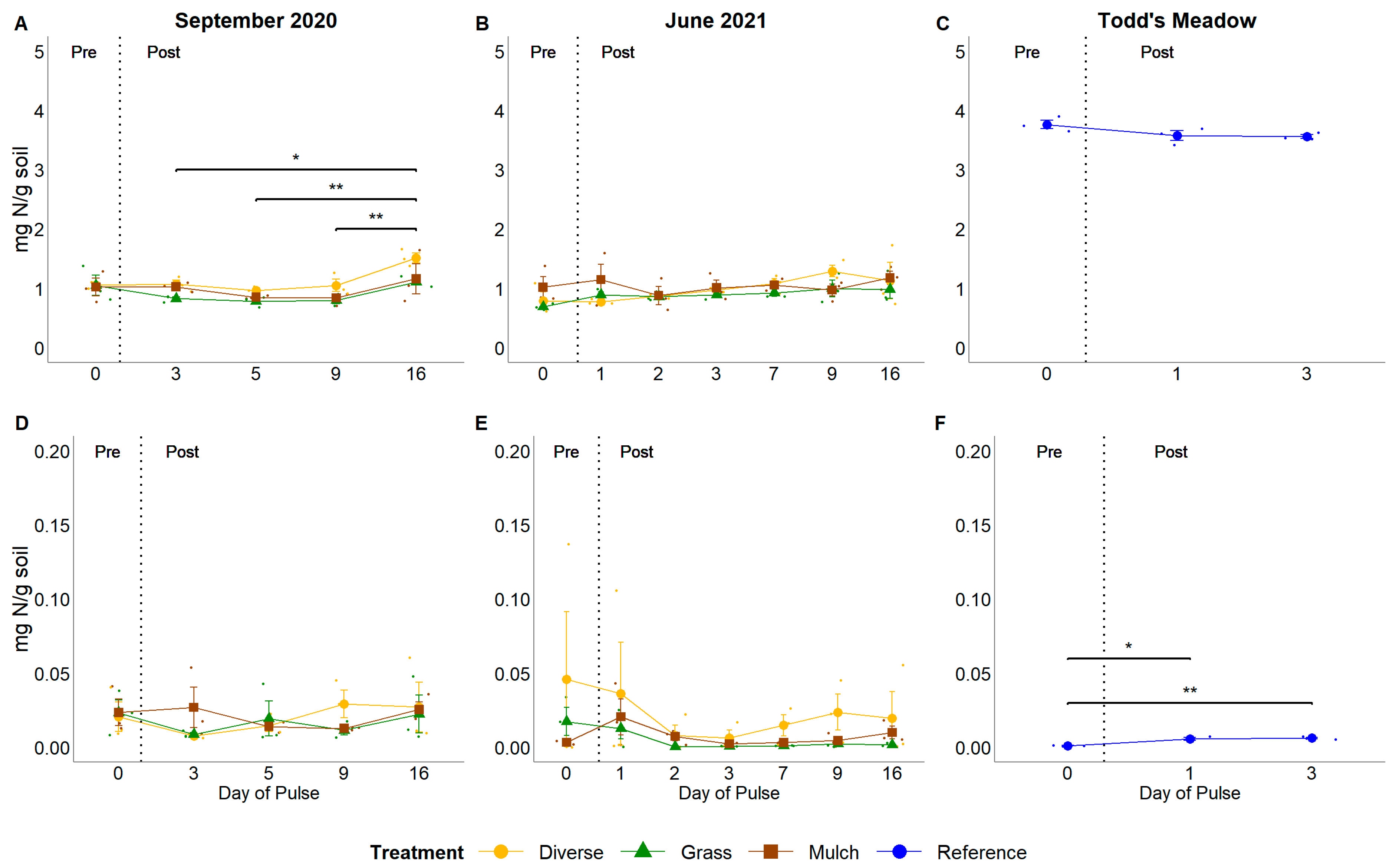
3.4. Soil Organic Matter Content and Pools and Fluxes of Soil N
3.5. Pearson Correlation and Piecewise Structural Equation Modeling
4. Discussion
4.1. Do Pulses of Water Equal in Magnitude to Moderate- to High-Intensity Storm Events Stimulate SGI Microbial Community Growth and Resource Acquisition?
4.2. Do Pulsed Water Additions Induce Changes to Soil N Pools and Fluxes?
4.3. To What Extent Are Microbial Physiology and Soil N Dynamics Controlled by Plant Diversity vs. Ecological Stoichiometry?
4.4. Study Limitations
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Gold, A.C.; Thompson, S.P.; Piehler, M.F. Nitrogen Cycling Processes within Stormwater Control Measures: A Review and Call for Research. Water Res. 2019, 149, 578–587. [Google Scholar] [CrossRef] [PubMed]
- Meerow, S.; Natarajan, M.; Krantz, D. Green Infrastructure Performance in Arid and Semi-Arid Urban Environments. Urban Water J. 2021, 18, 275–285. [Google Scholar] [CrossRef]
- Golden, H.E.; Hoghooghi, N. Green Infrastructure and Its Catchment-scale Effects: An Emerging Science. WIREs Water 2018, 5, e1254. [Google Scholar] [CrossRef]
- Walsh, C.J.; Roy, A.H.; Feminella, J.W.; Cottingham, P.D.; Groffman, P.M.; Ii, R.P.M. The Urban Stream Syndrome: Current Knowledge and the Search for a Cure. J. N. Am. Benthol. Soc. 2005, 24, 706–723. [Google Scholar] [CrossRef]
- Booth, D.B.; Roy, A.H.; Smith, B.; Capps, K.A. Global Perspectives on the Urban Stream Syndrome. Freshw. Sci. 2016, 35, 412–420. [Google Scholar] [CrossRef]
- Osman, M.; Yusof, K.W.; Takaijudin, H.; Goh, H.W.; Malek, M.A.; Azizan, N.A.; Ghani, A.A.; Abdurrasheed, A.S. A Review of Nitrogen Removal for Urban Stormwater Runoff in Bioretention System. Sustainability 2019, 11, 5415. [Google Scholar] [CrossRef]
- Galloway, J.N.; Townsend, A.R.; Erisman, J.W.; Bekunda, M.; Cai, Z.; Freney, J.R.; Martinelli, L.A.; Seitzinger, S.P.; Sutton, M.A. Transformation of the Nitrogen Cycle: Recent Trends, Questions, and Potential Solutions. Science 2008, 320, 889–892. [Google Scholar] [CrossRef] [PubMed]
- Decina, S.M.; Hutyra, L.R.; Templer, P.H. Hotspots of Nitrogen Deposition in the World’s Urban Areas: A Global Data Synthesis. Front. Ecol. Environ. 2020, 18, 92–100. [Google Scholar] [CrossRef]
- Collins, S.L.; Sinsabaugh, R.L.; Crenshaw, C.; Green, L.; Porras-Alfaro, A.; Stursova, M.; Zeglin, L.H. Pulse Dynamics and Microbial Processes in Aridland Ecosystems. J. Ecol. 2008, 96, 413–420. [Google Scholar] [CrossRef]
- Sinsabaugh, R.L.; Shah, J.J.F. Ecoenzymatic Stoichiometry and Ecological Theory. Annu. Rev. Ecol. Evol. Syst. 2012, 43, 313–343. [Google Scholar] [CrossRef]
- Deeb, M.; Groffman, P.M.; Joyner, J.L.; Lozefski, G.; Paltseva, A.; Lin, B.; Mania, K.; Cao, D.L.; McLaughlin, J.; Muth, T.; et al. Soil and Microbial Properties of Green Infrastructure Stormwater Management Systems. Ecol. Eng. 2018, 125, 68–75. [Google Scholar] [CrossRef]
- Ali, W.; Takaijudin, H.; Yusof, K.W.; Osman, M.; Abdurrasheed, A.S. The Common Approaches of Nitrogen Removal in Bioretention System. Sustainability 2021, 13, 2575. [Google Scholar] [CrossRef]
- Skorobogatov, A.; He, J.; Chu, A.; Valeo, C.; van Duin, B. The Impact of Media, Plants and Their Interactions on Bioretention Performance: A Review. Sci. Total Environ. 2020, 715, 136918. [Google Scholar] [CrossRef] [PubMed]
- Fowdar, H.; Payne, E.; Schang, C.; Zhang, K.F.; Deletic, A.; McCarthy, D. How Well Do Stormwater Green Infrastructure Respond to Changing Climatic Conditions? J. Hydrol. 2021, 603, 126887. [Google Scholar] [CrossRef]
- Zhu, W.-X.; Dillard, N.D.; Grimm, N.B. Urban Nitrogen Biogeochemistry: Status and Processes in Green Retention Basins. Biogeochemistry 2004, 71, 177–196. [Google Scholar] [CrossRef]
- Houdeshel, C.D.; Hultine, K.R.; Johnson, N.C.; Pomeroy, C.A. Evaluation of Three Vegetation Treatments in Bioretention Gardens in a Semi-Arid Climate. Landsc. Urban Plan. 2015, 135, 62–72. [Google Scholar] [CrossRef]
- Collins, S.L.; Belnap, J.; Grimm, N.B.; Rudgers, J.A.; Dahm, C.N.; D’Odorico, P.; Litvak, M.; Natvig, D.O.; Peters, D.C.; Pockman, W.T.; et al. A Multiscale, Hierarchical Model of Pulse Dynamics in Arid-Land Ecosystems. Annu. Rev. Ecol. Evol. Syst. 2014, 45, 397–419. [Google Scholar] [CrossRef]
- USGCRP. Impacts, Risks, and Adaptation in the United States. In Fourth National Climate Assessment; Reidmiller, D.R., Avery, C.W., Easterling, D.R., Kunkel, K.E., Lewis, K.L.M., Maycock, T.K., Stewart, B.C., Eds.; U.S. Global Change Research Program: Washington, DC, USA, 2018; Volume II, p. 1515. [Google Scholar] [CrossRef]
- Georgescu, M.; Broadbent, A.M.; Wang, M.; Krayenhoff, E.S.; Moustaoui, M. Precipitation Response to Climate Change and Urban Development over the Continental United States. Environ. Res. Lett. 2021, 16, 044001. [Google Scholar] [CrossRef]
- Wijesiri, B.; Liu, A.; Goonetilleke, A. Impact of Global Warming on Urban Stormwater Quality: From the Perspective of an Alternative Water Resource. J. Clean. Prod. 2020, 262, 121330. [Google Scholar] [CrossRef]
- Borken, W.; Matzner, E. Reappraisal of Drying and Wetting Effects on C and N Mineralization and Fluxes in Soils. Glob. Chang. Biol. 2009, 15, 808–824. [Google Scholar] [CrossRef]
- Loreau, M.; Naeem, S.; Inchausti, P.; Bengtsson, J.; Grime, J.P.; Hector, A.; Hooper, D.U.; Huston, M.A.; Raffaelli, D.; Schmid, B.; et al. Ecology: Biodiversity and Ecosystem Functioning: Current Knowledge and Future Challenges. Science 2001, 294, 804–808. [Google Scholar] [CrossRef]
- Hooper, D.U.; Chapin, F.S.; Ewel, J.J.; Hector, A.; Inchausti, P.; Lavorel, S.; Lawton, J.H.; Lodge, D.M.; Loreau, M.; Naeem, S.; et al. Effects of Biodiversity on Ecosystem Functioning: A Consensus of Current Knowledge. Ecol. Monogr. 2005, 75, 3–35. [Google Scholar] [CrossRef]
- de Bello, F.; Lavorel, S.; Díaz, S.; Harrington, R.; Cornelissen, J.H.C.; Bardgett, R.D.; Berg, M.P.; Cipriotti, P.; Feld, C.K.; Hering, D.; et al. Towards an Assessment of Multiple Ecosystem Processes and Services via Functional Traits. Biodivers. Conserv. 2010, 19, 2873–2893. [Google Scholar] [CrossRef]
- Cardinale, B.J.; Duffy, J.E.; Gonzalez, A.; Hooper, D.U.; Perrings, C.; Venail, P.; Narwani, A.; MacE, G.M.; Tilman, D.; Wardle, D.A.; et al. Biodiversity Loss and Its Impact on Humanity. Nature 2012, 486, 59–67. [Google Scholar] [CrossRef]
- Oliver, T.H.; Heard, M.S.; Isaac, N.J.B.; Roy, D.B.; Procter, D.; Eigenbrod, F.; Freckleton, R.; Hector, A.; Orme, C.D.L.; Petchey, O.L.; et al. Biodiversity and Resilience of Ecosystem Functions. Trends Ecol. Evol. 2015, 30, 673–684. [Google Scholar] [CrossRef] [PubMed]
- Houdeshel, C.D.; Pomeroy, C.A.; Hultine, K.R. Bioretention Design for Xeric Climates Based on Ecological Principles. J. Am. Water Resour. Assoc. 2012, 48, 1178–1190. [Google Scholar] [CrossRef]
- Sterner, R.W.; Elser, J.J. Ecological Stoichiometry: The Biology of Elements from Molecules to the Biosphere; Princeton University Press: Princeton, NJ, USA, 2002; p. 439. ISBN 9780691074917. [Google Scholar]
- Sinsabaugh, R.L.; Hill, B.H.; Follstad Shah, J.J. Ecoenzymatic Stoichiometry of Microbial Organic Nutrient Acquisition in Soil and Sediment. Nature 2009, 462, 795–798. [Google Scholar] [CrossRef] [PubMed]
- Zeglin, L.H.; Bottomley, P.J.; Jumpponen, A.; Rice, C.W.; Arango, M.; Lindsley, A.; McGowan, A.; Mfombep, P.; Myrold, D.D. Altered Precipitation Regime Affects the Function and Composition of Soil Microbial Communities on Multiple Time Scales. Ecology 2013, 94, 2334–2345. [Google Scholar] [CrossRef]
- Graham, E.B.; Knelman, J.E.; Schindlbacher, A.; Siciliano, S.; Breulmann, M.; Yannarell, A.; Beman, J.M.; Abell, G.; Philippot, L.; Prosser, J.; et al. Microbes as Engines of Ecosystem Function: When Does Community Structure Enhance Predictions of Ecosystem Processes? Front. Microbiol. 2016, 7, 214. [Google Scholar] [CrossRef] [PubMed]
- Moorhead, D.L.; Sinsabaugh, R.L.; Hill, B.H.; Weintraub, M.N. Vector Analysis of Ecoenzyme Activities Reveal Constraints on Coupled C, N and P Dynamics. Soil Biol. Biochem. 2016, 93, 1–7. [Google Scholar] [CrossRef]
- Noy-Meir, I. Desert Ecosystems: Environment and Producers. Annu. Rev. Ecol. Evol. Syst. 1973, 4, 25–51. [Google Scholar] [CrossRef]
- Xu, X.; Thornton, P.E.; Post, W.M. A Global Analysis of Soil Microbial Biomass Carbon, Nitrogen and Phosphorus in Terrestrial Ecosystems. Glob. Ecol. Biogeogr. 2013, 22, 737–749. [Google Scholar] [CrossRef]
- United States Department of Agriculture Natural Resources Conservation Service. USDA Plants Database. Available online: https://plants.sc.egov.usda.gov/home (accessed on 14 May 2020).
- Evenden, A.G.; Moeur, M.; Shelly, J.S.; Kimball, S.F.; Wellner, C.A. Research Natural Areas on National Forest System Lands in Idaho, Montana, Nevada, Utah, and Western Wyoming: A Guidebook for Scientists, Managers, and Educators; General Technical Report RMRS-STR-69; US Department of Agriculture, Forest Service, Rocky Mountain Research Station: Ogden, UT, USA, 2001; p. 21.
- Woodward, L.; Harvey, J.L.; Donaldson, K.M.; Shiozaki, J.J.; Leishman, G.W.; Broderick, J.H. Soil Survey of Salt Lake Area, Utah; US Department of Agriculture, Soil Conservation Service: Salt Lake County, UT, USA, 1974; pp. 55–56.
- Red Butte Canyon Research Natural Area. Available online: https://redbuttecanyon.net/ (accessed on 14 March 2022).
- Follstad Shah, J.J.; Bares, R.; Bowen, B.B.; Bowen, G.J.; Bowling, D.R.; Eiriksson, D.P.; Fasoli, B.; Fiorella, R.P.; Hallar, A.G.; Hinners, S.J.; et al. The Wasatch Environmental Observatory: A Mountain to Urban Research Network in the Semi-Arid Western US. Hydrol. Process. 2021, 35, e14352. [Google Scholar] [CrossRef]
- University of Utah—Wasatch Environmental Observatory Wasatch Environmental Observatory Red Butte Network: Todds Meadow Climate (RB_TM_C) Quality Controlled Data. Available online: http://www.hydroshare.org/resource/79ae0f0efe2447fe9a5ab5c15427b2d8 (accessed on 14 March 2022).
- University of Utah—Wasatch Environmental Observatory Wasatch Environmental Observatory Red Butte Network: Green Infrastructure Climate (RM_GIRF_C) Quality Controlled Data. Available online: http://www.hydroshare.org/resource/e5935762e9054fc49570f02d1a28ed8a (accessed on 14 March 2022).
- University of Utah—Wasatch Environmental Observatory Green Infrastructure Research Facility (GIRF) Sensor Data. Available online: http://www.hydroshare.org/resource/331588a2bfe44206a5290d660d9097b6 (accessed on 14 March 2022).
- National Drought Mitigation Center. U.S. Drought Monitor. Available online: https://droughtmonitor.unl.edu/ (accessed on 8 March 2023).
- Sinsabaugh, R.L.; Lauber, C.L.; Weintraub, M.N.; Ahmed, B.; Allison, S.D.; Crenshaw, C.; Contosta, A.R.; Cusack, D.; Frey, S.; Gallo, M.E.; et al. Stoichiometry of Soil Enzyme Activity at Global Scale. Ecol. Lett. 2008, 11, 1252–1264. [Google Scholar] [CrossRef] [PubMed]
- Weintraub, S. NEON User Guide to Soil Inorganic Nitrogen Pools and Transformations (NEON.DP1.10086); US National Science Foundation, National Ecological Observatory Network, Terrestrial Observation System: Alexandria, VI, USA, 2020; p. 18.
- Brookes, P.; Landman, A.; Pruden, G.; Jenkinson, D. Chloroform Fumigation and the Release of Soil Nitrogen: A Rapid Direct Extraction Method to Measure Microbial Biomass Nitrogen in Soil. Soil Biol. Biochem. 1985, 17, 837–842. [Google Scholar] [CrossRef]
- Weintraub, S.R.; Brooks, P.D.; Bowen, G.J. Interactive Effects of Vegetation Type and Topographic Position on Nitrogen Availability and Loss in a Temperate Montane Ecosystem. Ecosystems 2017, 20, 1073–1088. [Google Scholar] [CrossRef]
- Hill, B.H.; Elonen, C.M.; Seifert, L.R.; May, A.A.; Tarquinio, E. Microbial Enzyme Stoichiometry and Nutrient Limitation in US Streams and Rivers. Ecol. Indic. 2012, 18, 540–551. [Google Scholar] [CrossRef]
- Bach, C.E.; Warnock, D.D.; Van Horn, D.J.; Weintraub, M.N.; Sinsabaugh, R.L.; Allison, S.D.; German, D.P. Measuring Phenol Oxidase and Peroxidase Activities with Pyrogallol, l-DOPA, and ABTS: Effect of Assay Conditions and Soil Type. Soil Biol. Biochem. 2013, 67, 183–191. [Google Scholar] [CrossRef]
- Sinsabaugh, R.L. Phenol Oxidase, Peroxidase and Organic Matter Dynamics of Soil. Soil Biol. Biochem. 2010, 42, 391–404. [Google Scholar] [CrossRef]
- Weintraub, M.N.; Schimel, J.P. Seasonal Protein Dynamics in Alaskan Arctic Tundra Soils. Soil Biol. Biochem. 2005, 37, 1469–1475. [Google Scholar] [CrossRef]
- R Core Team. R: A Language and Environment for Statistical Computing; R Foundation for Statistical Computing: Vienna, Austria, 2020; Available online: https://www.R-project.org/ (accessed on 1 May 2020).
- Lefcheck, J.S. PiecewiseSEM: Piecewise Structural Equation Modelling in r for Ecology, Evolution, and Systematics. Methods Ecol. Evol. 2016, 7, 573–579. [Google Scholar] [CrossRef]
- Esch, E.H.; Lipson, D.; Cleland, E.E. Direct and Indirect Effects of Shifting Rainfall on Soil Microbial Respiration and Enzyme Activity in a Semi-Arid System. Plant Soil 2017, 411, 333–346. [Google Scholar] [CrossRef]
- Bi, J.; Zhang, N.; Liang, Y.; Yang, H.; Ma, K. Interactive Effects of Water and Nitrogen Addition on Soil Microbial Communities in a Semiarid Steppe. J. Plant Ecol. 2012, 5, 320–329. [Google Scholar] [CrossRef]
- Pavao-Zuckerman, M.A.; Sookhdeo, C. Nematode Community Response to Green Infrastructure Design in a Semiarid City. J. Environ. Qual. 2017, 46, 687–694. [Google Scholar] [CrossRef] [PubMed]
- Li, Z.; Zeng, Z.; Tian, D.; Wang, J.; Fu, Z.; Zhang, F.; Zhang, R.; Chen, W.; Luo, Y.; Niu, S. Global Patterns and Controlling Factors of Soil Nitrification Rate. Glob. Change Biol. 2020, 26, 4147–4157. [Google Scholar] [CrossRef] [PubMed]
- Bettez, N.D.; Groffman, P.M. Denitrification Potential in Stormwater Control Structures and Natural Riparian Zones in an Urban Landscape. Environ. Sci. Technol. 2012, 46, 10909–10917. [Google Scholar] [CrossRef] [PubMed]
- Stursova, M.; Crenshaw, C.L.; Sinsabaugh, R.L. Microbial Responses to Long-Term N Deposition in a Semiarid Grassland. Microb. Ecol. 2006, 51, 90–98. [Google Scholar] [CrossRef] [PubMed]
- Stursova, M.; Sinsabaugh, R.L. Stabilization of Oxidative Enzymes in Desert Soil May Limit Organic Matter Accumulation. Soil Biol. Biochem. 2008, 40, 550–553. [Google Scholar] [CrossRef]
- Tiemann, L.K.; Billings, S.A. Changes in Variability of Soil Moisture Alter Microbial Community C and N Resource Use. Soil Biol. Biochem. 2011, 43, 1837–1847. [Google Scholar] [CrossRef]
- McPhillips, L.; Walter, M.T. Hydrologic Conditions Drive Denitrification and Greenhouse Gas Emissions in Stormwater Detention Basins. Ecol. Eng. 2015, 85, 67–75. [Google Scholar] [CrossRef]
- Payne, E.G.I.; Fletcher, T.D.; Russell, D.G.; Grace, M.R.; Cavagnaro, T.R.; Evrard, V.; Deletic, A.; Hatt, B.E.; Cook, P.L.M. Temporary Storage or Permanent Removal? The Division of Nitrogen between Biotic Assimilation and Denitrification in Stormwater Biofiltration Systems. PLoS ONE 2014, 9, e0090890. [Google Scholar] [CrossRef]
- Zinger, Y.; Prodanovic, V.; Zhang, K.F.; Fletcher, T.D.; Deletic, A. The Effect of Intermittent Drying and Wetting Stormwater Cycles on the Nutrient Removal Performances of Two Vegetated Biofiltration Designs. Chemosphere 2021, 267, 129294. [Google Scholar] [CrossRef] [PubMed]
- Rivers, E.N.; Morse, J.L. Variability of Potential Soil Nitrogen Cycling Rates in Stormwater Bioretention Facilities. Sustainability 2022, 14, 2175. [Google Scholar] [CrossRef]
- Shrestha, P.; Hurley, S.E.; Adair, E.C. Soil Media CO2 and N2O Fluxes Dynamics from Sand-Based Roadside Bioretention Systems. Water 2018, 10, 185. [Google Scholar] [CrossRef]
- Ladwig, L.M.; Sinsabaugh, R.L.; Collins, S.L.; Thomey, M.L. Soil Enzyme Responses to Varying Rainfall Regimes in Chihuahuan Desert Soils. Ecosphere 2015, 6, 1–10. [Google Scholar] [CrossRef]
- Wang, R.; Filley, T.R.; Xu, Z.; Wang, X.; Li, M.H.; Zhang, Y.; Luo, W.; Jiang, Y. Coupled Response of Soil Carbon and Nitrogen Pools and Enzyme Activities to Nitrogen and Water Addition in a Semi-Arid Grassland of Inner Mongolia. Plant Soil 2014, 381, 323–336. [Google Scholar] [CrossRef]
- Zhang, B.; Chen, L.; Guo, Q.; Lian, J.; Yao, Y. Evaluation of Ammonia and Nitrate Distribution and Reduction within Stormwater Green Infrastructure with Different Woody Plants under Multiple Influencing Factors. J. Environ. Manag. 2022, 302, 114086. [Google Scholar] [CrossRef] [PubMed]
- Barron, N.J.; Hatt, B.; Jung, J.; Chen, Y.; Deletic, A. Seasonal Operation of Dual-Mode Biofilters: The Influence of Plant Species on Stormwater and Greywater Treatment. Sci. Total Environ. 2020, 715, 136680. [Google Scholar] [CrossRef]
- Ivanov, V.Y.; Fatichi, S.; Jenerette, G.D.; Espeleta, J.F.; Troch, P.A.; Huxman, T.E. Hysteresis of Soil Moisture Spatial Heterogeneity and the “Homogenizing” Effect of Vegetation. Water Resour. Res. 2010, 46, W09521. [Google Scholar] [CrossRef]
- Månsson, K.; Bengtson, P.; Falkengren-Grerup, U.; Bengtsson, G. Plant-Microbial Competition for Nitrogen Uncoupled from Soil C:N Ratios. Oikos 2009, 118, 1908–1916. [Google Scholar] [CrossRef]
- Cui, Y.; Moorhead, D.L.; Peng, S.; Sinsabaugh, R.L. New Insights into the Patterns of Ecoenzymatic Stoichiometry in Soil and Sediment. Soil Biol. Biochem. 2023, 177, 108910. [Google Scholar] [CrossRef]
- Cleveland, C.C.; Liptzin, D. C:N:P Stoichiometry in Soil: Is There a “Redfield Ratio” for the Microbial Biomass? Biogeochemistry 2007, 85, 235–252. [Google Scholar] [CrossRef]
- Sinsabaugh, R.L.; Shah, J.J.F.; Findlay, S.G.; Kuehn, K.A.; Moorhead, D.L. Scaling Microbial Biomass, Metabolism and Resource Supply. Biogeochemistry 2015, 122, 175–190. [Google Scholar] [CrossRef]
- Díaz, S.; Kattge, J.; Cornelissen, J.H.C.; Wright, I.J.; Lavorel, S.; Dray, S.; Reu, B.; Kleyer, M.; Wirth, C.; Colin Prentice, I.; et al. The Global Spectrum of Plant Form and Function. Nature 2016, 529, 167–171. [Google Scholar] [CrossRef]
- Moreau, D.; Pivato, B.; Bru, D.; Busset, H.; Deau, F.; Faivre, C.; Matejicek, A.; Strbik, F.; Philippot, L.; Mougel, C. Plant Traits Related to Nitrogen Uptake Influence Plantmicrobe Competition. Ecology 2015, 96, 2300–2310. [Google Scholar] [CrossRef] [PubMed]
- Austin, A.T.; Yahdjian, L.; Stark, J.M.; Belnap, J.; Porporato, A.; Norton, U.; Ravetta, D.A.; Schaeffer, S.M. Water Pulses and Biogeochemical Cycles in Arid and Semiarid Ecosystems. Oecologia 2004, 141, 221–235. [Google Scholar] [CrossRef] [PubMed]
- Dijkstra, F.A.; Augustine, D.J.; Brewer, P.; von Fischer, J.C. Nitrogen Cycling and Water Pulses in Semiarid Grasslands: Are Microbial and Plant Processes Temporally Asynchronous? Oecologia 2012, 170, 799–808. [Google Scholar] [CrossRef] [PubMed]
- Reisinger, A.J.; Groffman, P.M.; Rosi-Marshall, E.J. Nitrogen-Cycling Process Rates across Urban Ecosystems. FEMS Microbiol. Ecol. 2016, 92, fiw198. [Google Scholar] [CrossRef]
- Hillebrand, H.; Cowles, J.M.; Lewandowska, A.; Van de Waal, D.B.; Plum, C. Think Ratio! A Stoichiometric View on Biodiversity-Ecosystem Functioning Research. Basic Appl. Ecol. 2014, 15, 465–474. [Google Scholar] [CrossRef]




{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Enzyme | EC | Abbreviation | Substrate | Example Source |
|---|---|---|---|---|
| β-1,4-glucosidase | EC 3.2.1.21 | BG | 4-MUB-β-D-glucoside | C from simple sugars |
| Leucyl aminopeptidase | EC 3.4.11.1 | LAP | L-Leucine-7-amido- 4-methylcoumarin | N from amino acid chains |
| Alkaline phosphatase | EC 3.1.3.1 | AP | 4-MUB-phosphate | P from phospholipids |
| Phenol oxidase | EC 1.10.3 | POX | L-3,4-dihydroxyphenylalanine | C from lignin |
| Enzyme | Sept 2020 Experimental Pulse Average Activity Rate (nmol g−1 h−1) | June 2021 Experimental Pulse Average Activity Rate (nmol g−1 h−1) | 2021 Todd’s Meadow Natural Pulse Average Activity Rate (nmol g−1 h−1) |
|---|---|---|---|
| Acid Phosphatase (AP) | 17.63 ± 1.17 | 22.87 ± 1.13 | 39.39 ± 3.62 |
| Leucyl Aminopeptidase (LAP) | 15.49 ± 0.51 | 17.49 ± 0.52 | 35.26 ± 4.12 |
| β-1,4-Glucosidase (BG) | 6.72 ± 0.37 | 12.82 ± 0.73 | 33.96 ± 4.44 |
| Phenol Oxidase (POX) | 7010 ± 536 | 10237 ± 524 | 4071 ± 520 |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2024 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Hastings, Y.D.; Smith, R.M.; Mann, K.A.; Brewer, S.; Goel, R.; Hinners, S.J.; Shah, J.F. Green Infrastructure Microbial Community Response to Simulated Pulse Precipitation Events in the Semi-Arid Western United States. Water 2024, 16, 1931. https://doi.org/10.3390/w16131931
Hastings YD, Smith RM, Mann KA, Brewer S, Goel R, Hinners SJ, Shah JF. Green Infrastructure Microbial Community Response to Simulated Pulse Precipitation Events in the Semi-Arid Western United States. Water. 2024; 16(13):1931. https://doi.org/10.3390/w16131931
Chicago/Turabian StyleHastings, Yvette D., Rose M. Smith, Kyra A. Mann, Simon Brewer, Ramesh Goel, Sarah Jack Hinners, and Jennifer Follstad Shah. 2024. "Green Infrastructure Microbial Community Response to Simulated Pulse Precipitation Events in the Semi-Arid Western United States" Water 16, no. 13: 1931. https://doi.org/10.3390/w16131931
APA StyleHastings, Y. D., Smith, R. M., Mann, K. A., Brewer, S., Goel, R., Hinners, S. J., & Shah, J. F. (2024). Green Infrastructure Microbial Community Response to Simulated Pulse Precipitation Events in the Semi-Arid Western United States. Water, 16(13), 1931. https://doi.org/10.3390/w16131931