
Comparing Four Evapotranspiration Partitioning Methods from Eddy Covariance Considering Turbulent Mixing in a Poplar Plantation

 ,
,
Abstract
1. Introduction
2. Materials and Methods
2.1. The Site Introduction
2.2. Evapotranspiration Partitioning and Data Processing
2.2.1. The Double-Layer Eddy Covariance (DLEC) Method
2.2.2. The Determination of the Coupling State across the Canopy Vertical Layer
3. Results and Discussion
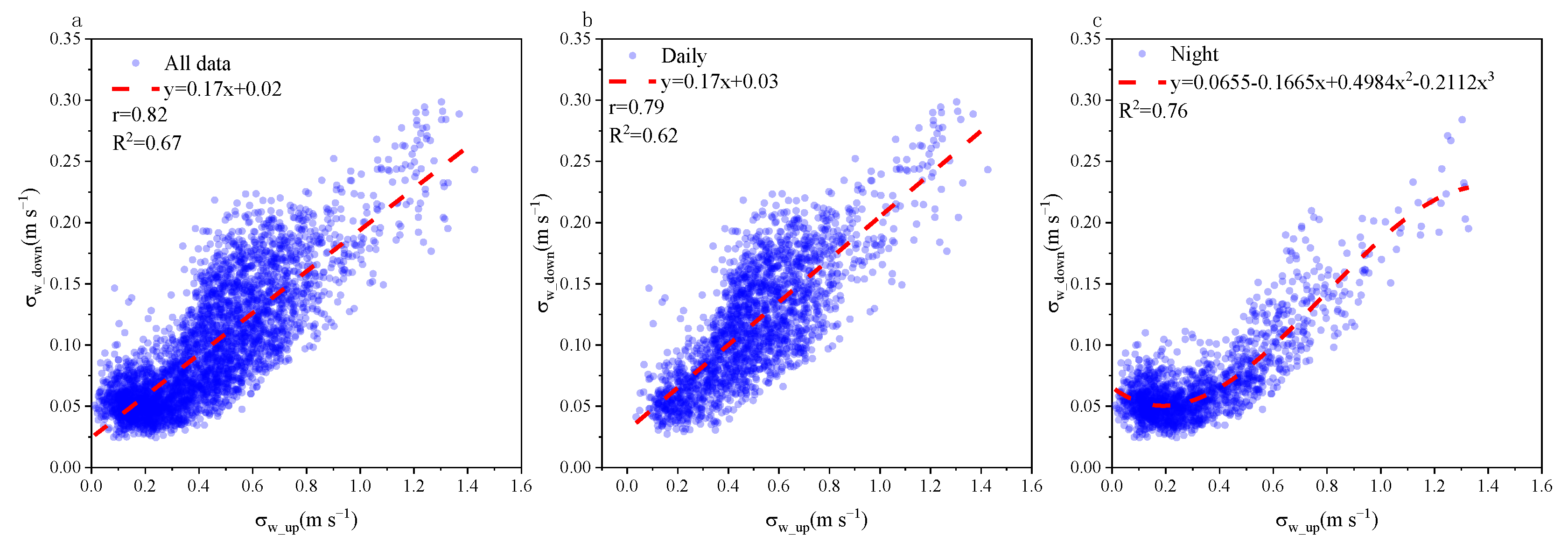
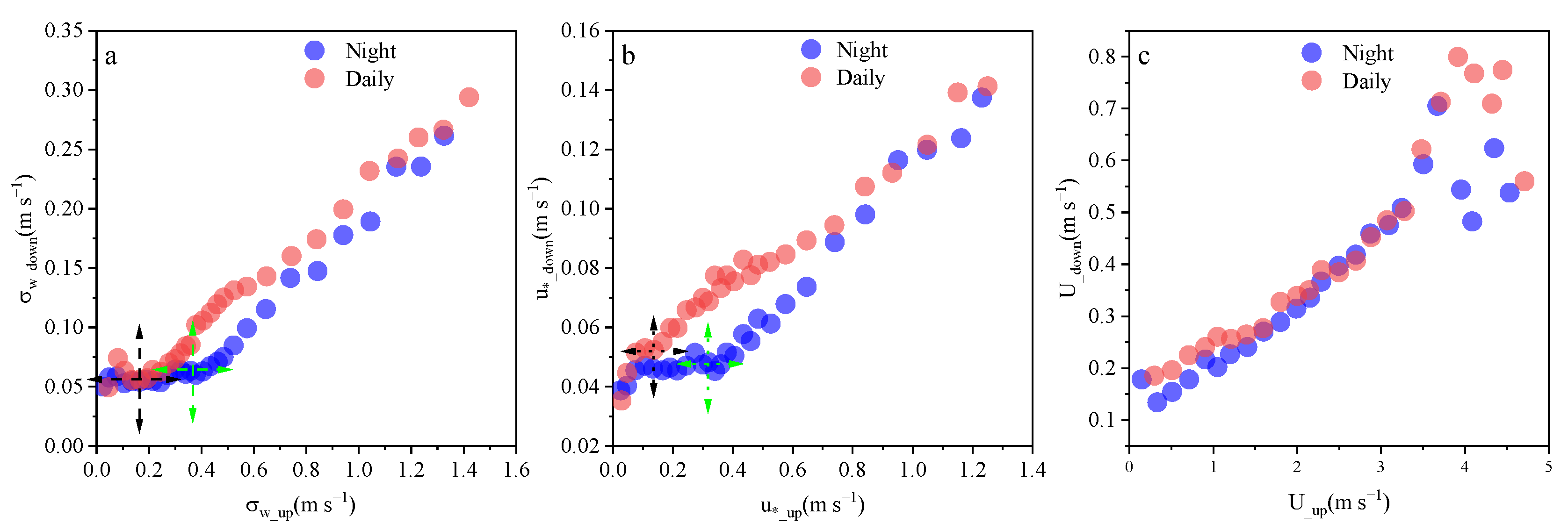
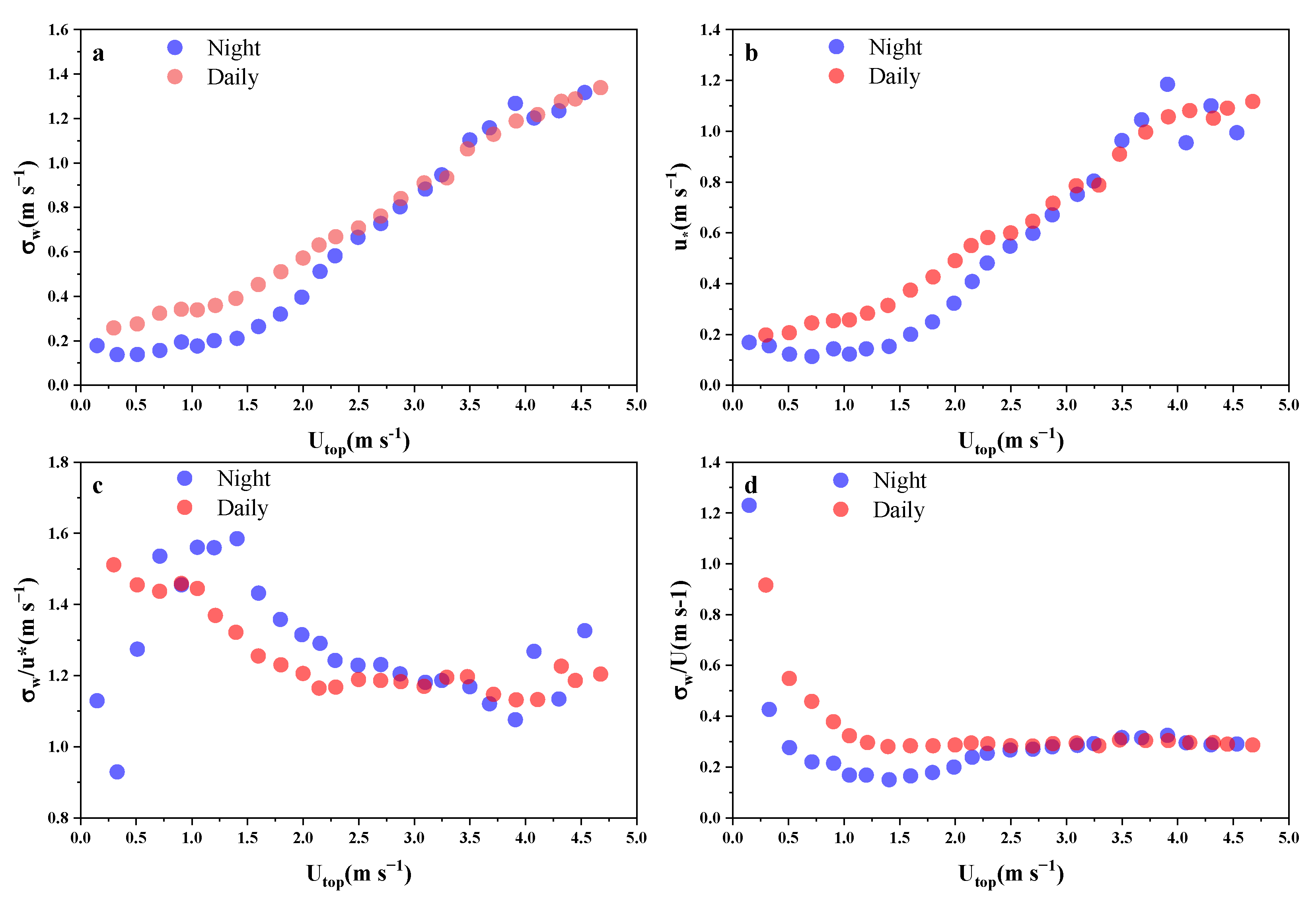
3.1. Coupling State of Airflow Crossing the Vertical Layer of the Plantation Canopy
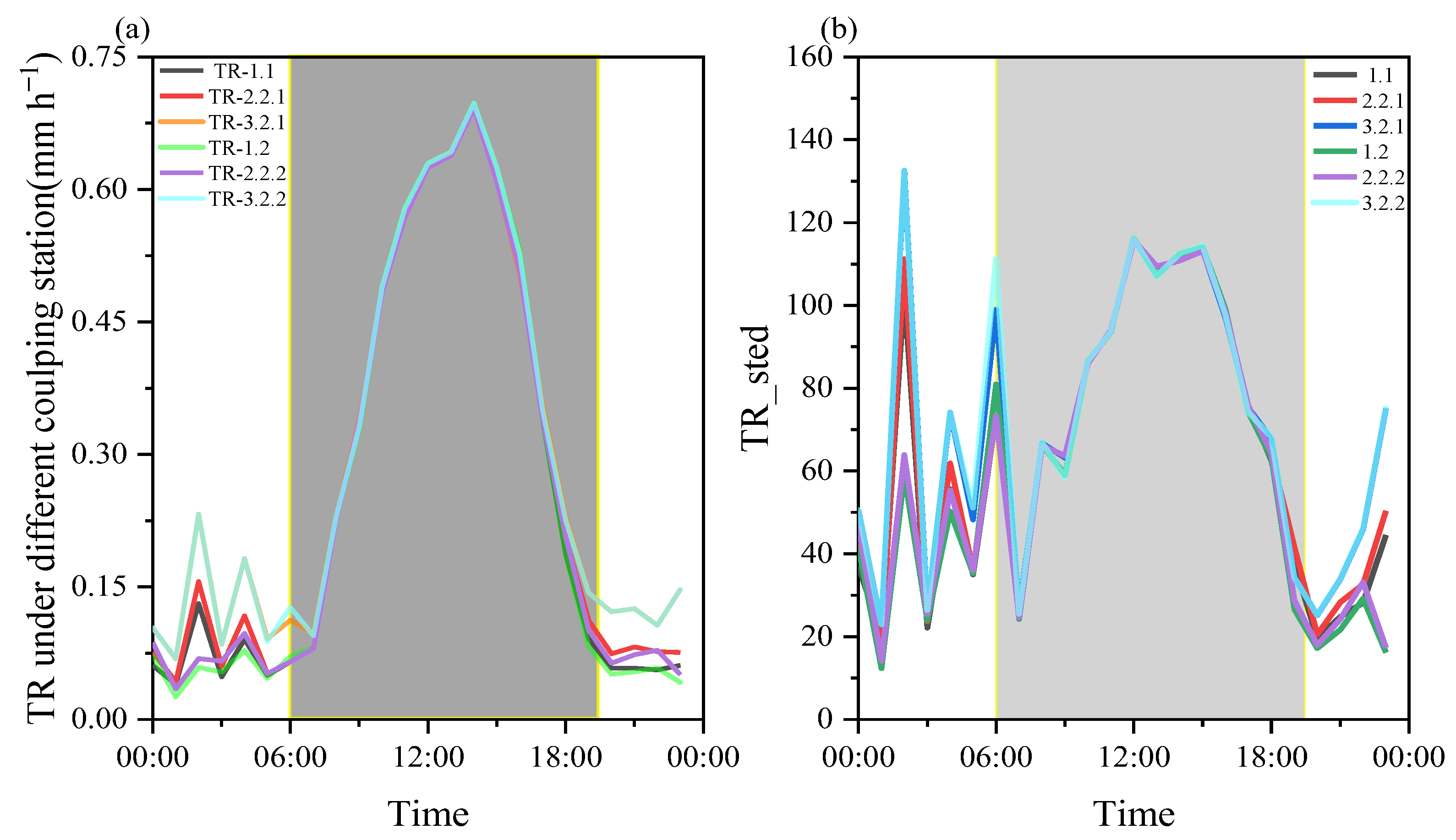
3.2. Do Different Coupling States Obviously Affect the Daily Transpiration in a Sparse Canopy?
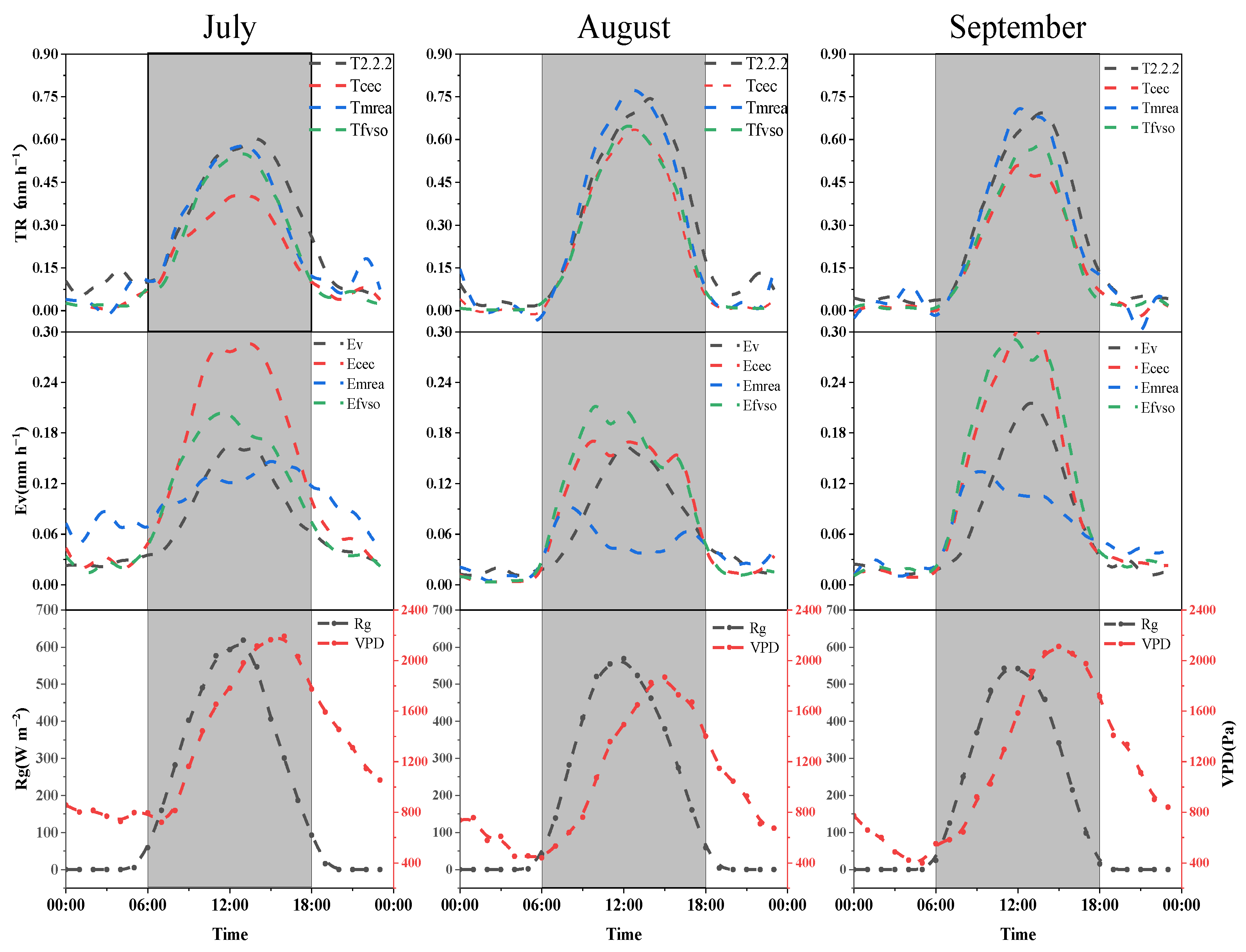
3.3. Comparison of the Results of Different Partitioning Methods
3.4. Uncertainties and Limitations of Separation Methods
4. Conclusions
Author Contributions
Funding
Data Availability Statement
Conflicts of Interest
Appendix A. Methods of ET Partitioning
Appendix A.1. The Modified Relaxed Eddy Accumulation (MREA) Method
Appendix A.2. The Conditional Eddy Covariance (CEC) Method
Appendix A.3. The Flux Variance Similarity (FVS) Method
References
- Wang, K.; Dickinson, R.E. A review of global terrestrial evapotranspiration: Observation, modeling, climatology, and climatic variability. Rev. Geophys. 2012, 50, RG2005. [Google Scholar] [CrossRef]
- Lobell, D.B.; Field, C.B. Estimation of the CO2 fertilization effect using growth rate anomalies of CO2 and crop yields since 1961. Glob. Chang. Biol. 2008, 14, 451. [Google Scholar]
- Cao, L.; Bala, G.; Caldeira, K.; Nemani, R.; Ban-Weiss, G. Importance of carbon dioxide physiological forcing to future climate change. Proc. Natl. Acad. Sci. USA 2010, 107, 9513–9518. [Google Scholar] [CrossRef] [PubMed]
- Field, C.B.; Jackson, R.B.; Mooney, H.A. Stomatal responses to increased CO2: Implications from the plant to the global scale. Plant Cell Environ. 1995, 18, 1214–1225. [Google Scholar] [CrossRef]
- Dai, A. Increasing drought under global warming in observations and models. Nat. Clim. Chang. 2013, 3, 52–58. [Google Scholar] [CrossRef]
- Zhou, S.; Yu, B.; Zhang, Y.; Huang, Y.; Wang, G. Partitioning evapotranspiration based on the concept of underlying water use efficiency. Water Resour. Res. 2016, 52, 1160–1175. [Google Scholar] [CrossRef]
- Scott, R.L.; Biederman, J.A. Partitioning evapotranspiration using long-term carbon dioxide and water vapor fluxes. Geophys. Res. Lett. 2017, 44, 6833–6840. [Google Scholar] [CrossRef]
- Nelson, J.A.; Carvalhais, N.; Cuntz, M.; Delpierre, N.; Knauer, J.; Ogée, J.; Migliavacca, M.; Reichstein, M.; Jung, M. Coupling Water and Carbon Fluxes to Constrain Estimates of Transpiration: The TEA Algorithm. J. Geophys. Res. Biogeosci. 2018, 123, 3617–3632. [Google Scholar] [CrossRef]
- Paul-Limoges, E.; Wolf, S.; Schneider, F.D.; Longo, M.; Moorcroft, P.; Gharun, M.D.A. Partitioning evapotranspiration with concurrent eddy covariance measurements in a mixed forest. Agric. For. Meteorol. 2020, 280, 107786. [Google Scholar] [CrossRef]
- Kool, D.; Agam, N.; Lazarovitch, N.; Heitman, J.; Sauer, T.; Ben-Gal, A. A review of approaches for evapotranspiration partitioning. Agric. For. Meteorol. 2014, 184, 56–70. [Google Scholar] [CrossRef]
- Rana, G.; De Lorenzi, F.; Palatella, L.; Martinelli, N.; Ferrara, R.M. Field scale recalibration of the sap flow thermal dissipation method in a Mediterranean vineyard. Agric. For. Meteorol. 2019, 269–270, 169–179. [Google Scholar] [CrossRef]
- Wilson, K.B.; Hanson, P.J.; Mulholland, P.J.; Baldocchi, D.D.; Wullschleger, S.D. A comparison of methods for determining forest evapotranspiration and its components: Sap-flow, soil water budget, eddy covariance and catchment water balance. Agric. For. Meteorol. 2001, 106, 153–168. [Google Scholar] [CrossRef]
- Daamen, C.C.; Simmonds, L.P.; Wallace, J.S.; Laryea, K.B.; Sivakumar, M.V.K. Use of microlysimeters to measure evaporation from sandy soils. Agric. For. Meteorol. 1993, 65, 159–173. [Google Scholar] [CrossRef]
- Lobit, P.; Mpandeli, N.S.; Annandale, J.G.; Jovanovic, N.Z.; du Sautoy, N. Validation of the soil evaporation subroutine of the SWB-2D model in a hedgerow peach orchard. S. Afr. J. Plant Soil 2004, 21, 220–229. [Google Scholar] [CrossRef]
- Detto, M.; Katul, G.; Mancini, M.; Montaldo, N.; Albertson, J.D. Surface heterogeneity and its signature in higher-order scalar similarity relationships. Agric. For. Meteorol. 2008, 148, 902–916. [Google Scholar] [CrossRef]
- Scanlon, T.M.; Albertson, J.D. Turbulent transport of carbon dioxide and water vapor within a vegetation canopy during unstable conditions: Identification of episodes using wavelet analysis. J. Geophys. Res. Atmos. 2001, 106, 7251–7262. [Google Scholar] [CrossRef]
- Zahn, E.; Bou-Zeid, E.; Good, S.P.; Katul, G.G.; Thomas, C.K.; Ghannam, K.; Smith, J.A.; Chamecki, M.; Dias, N.L.; Fuentes, J.D.; et al. Direct partitioning of eddy-covariance water and carbon dioxide fluxes into ground and plant components. Agric. For. Meteorol. 2022, 315, 108790. [Google Scholar] [CrossRef]
- Scanlon, T.M.; Sahu, P. On the correlation structure of water vapor and carbon dioxide in the atmospheric surface layer: A basis for flux partitioning. Water Resour. Res. 2008, 44. [Google Scholar] [CrossRef]
- Scanlon, T.M.; Kustas, W.P. Partitioning carbon dioxide and water vapor fluxes using correlation analysis. Agric. For. Meteorol. 2010, 150, 89–99. [Google Scholar] [CrossRef]
- Jocher, G.; Ottosson Löfvenius, M.; De Simon, G.; Hörnlund, T.; Linder, S.; Lundmark, T.; Marshall, J.; Nilsson, M.B.; Näsholm, T.; Tarvainen, L.; et al. Apparent winter CO2 uptake by a boreal forest due to decoupling. Agric. For. Meteorol. 2017, 232, 23–34. [Google Scholar] [CrossRef]
- Jocher, G.; Marshall, J.; Nilsson, M.B.; Linder, S.; De Simon, G.; Hörnlund, T.; Lundmark, T.; Näsholm, T.; Löfvenius, M.O.; Tarvainen, L.; et al. Impact of Canopy Decoupling and Subcanopy Advection on the Annual Carbon Balance of a Boreal Scots Pine Forest as Derived From Eddy Covariance. J. Geophys. Res. Biogeosci. 2018, 123, 303–325. [Google Scholar] [CrossRef]
- Roupsard, O.; Bonnefond, J.M.; Irvine, M.; Berbigier, P.; Nouvellon, Y.; Dauzat, J.; Taga, S.; Hamel, O.; Jourdan, C.; Saint-André, L.; et al. Partitioning energy and evapo-transpiration above and below a tropical palm canopy. Agric. For. Meteorol. 2006, 139, 252–268. [Google Scholar] [CrossRef]
- Thomas, C.K.; Martin, J.G.; Law, B.E.; Davis, K. Toward biologically meaningful net carbon exchange estimates for tall, dense canopies: Multi-level eddy covariance observations and canopy coupling regimes in a mature Douglas-fir forest in Oregon. Agric. For. Meteorol. 2013, 173, 14–27. [Google Scholar] [CrossRef]
- Denmead, O.T.; Bradley, E.F. Flux-Gradient Relationships in a Forest Canopy. In The Forest-Atmosphere Interaction: Proceedings of the Forest Environmental Measurements Conference Held at Oak Ridge, Tennessee, 23–28 October 1983; Hutchison, B.A., Hicks, B.B., Eds.; Springer: Dordrecht, The Netherlands, 1985; pp. 421–442. [Google Scholar]
- Vickers, D.; Irvine, J.; Martin, J.G.; Law, B.E. Nocturnal subcanopy flow regimes and missing carbon dioxide. Agric. For. Meteorol. 2012, 152, 101–108. [Google Scholar] [CrossRef]
- Vickers, D.; Mahrt, L. Quality control and flux sampling problems for tower and aircraft data. J. Atmos. Ocean. Technol. 1997, 14, 512–526. [Google Scholar] [CrossRef]
- Goulden, M.L.; Munger, J.W.; Fan, S.M.; Daube, B.C.; Wofsy, S.C. Measurements of carbon sequestration by long-term eddy covariance: Methods and a critical evaluation of accuracy. Glob. Chang. Biol. 1996, 2, 169–182. [Google Scholar] [CrossRef]
- Suyker, A.E.; Verma, S.B.; Burba, G.G. Interannual variability in net CO2 exchange of a native tallgrass prairie. Glob. Chang. Biol. 2003, 9, 255–265. [Google Scholar] [CrossRef]
- Acevedo, O.C.; Moraes, O.L.L.; Degrazia, G.A.; Fitzjarrald, D.R.; Manzi, A.O.; Campos, J.G. Is friction velocity the most appropriate scale for correcting nocturnal carbon dioxide fluxes? Agric. For. Meteorol. 2009, 149, 1–10. [Google Scholar] [CrossRef]
- Foken, T.; Wichura, B. Tools for quality assessment of surface-based flux measurements. Agric. For. Meteorol. 1996, 78, 83–105. [Google Scholar] [CrossRef]
- Thomas, C.; Foken, T. Re-evaluation of integral turbulence characteristics and their parameterisations. In Proceedings of the 15th Symposium on Boundary Layers and Turbulence, Wageningen, The Netherlands, 15–19 July 2002. [Google Scholar]
- Paul-Limoges, E.; Wolf, S.; Eugster, W.; Hörtnagl, L.; Buchmann, N. Below-canopy contributions to ecosystem CO2 fluxes in a temperate mixed forest in Switzerland. Agric. For. Meteorol. 2017, 247, 582–596. [Google Scholar] [CrossRef]
- Kowalska, N.; Jocher, G.; Šigut, L.; Pavelka, M. Does Below-Above Canopy Air Mass Decoupling Impact Temperate Floodplain Forest CO2 Exchange? Atmosphere 2022, 13, 437. [Google Scholar] [CrossRef]
- Foken, T.; Aubinet, M.; Leuning, R. The Eddy Covariance Method, in Eddy Covariance: A Practical Guide to Measurement and Data Analysis; Aubinet, M., Vesala, T., Papale, D., Eds.; Springer: Dordrecht, The Netherlands, 2012; pp. 1–19. [Google Scholar]
- Aubinet, M.; Vesala, T.; Papale, D. Eddy Covariance: A Practical Guide to Measurement and Data Analysis; Springer Science & Business Media: Berlin/Heidelberg, Germany, 2012. [Google Scholar]
- Wagle, P.; Skaggs, T.H.; Gowda, P.H.; Northup, B.K.; Neel, J.P.S.; Anderson, R.G. Evaluation of Water Use Efficiency Algorithms for Flux Variance Similarity-Based Evapotranspiration Partitioning in C3 and C4 Grain Crops. Water Resour. Res. 2021, 57, e2020WR028866. [Google Scholar] [CrossRef]
- Thomas, C.; Martin, J.G.; Goeckede, M.; Siqueira, M.B.; Foken, T.; Law, B.E.; Loescher, H.; Katul, G. Estimating daytime subcanopy respiration from conditional sampling methods applied to multi-scalar high frequency turbulence time series. Agric. For. Meteorol. 2008, 148, 1210–1229. [Google Scholar] [CrossRef]
- Sulman, B.N.; Roman, D.T.; Scanlon, T.M.; Wang, L.; Novick, K.A. Comparing methods for partitioning a decade of carbon dioxide and water vapor fluxes in a temperate forest. Agric. For. Meteorol. 2016, 226–227, 229–245. [Google Scholar] [CrossRef]
- Roth, B.E.; Slatton, K.C.; Cohen, M.J. On the potential for high-resolution lidar to improve rainfall interception estimates in forest ecosystems. Front. Ecol. Environ. 2007, 5, 421–428. [Google Scholar] [CrossRef]
- Gash, J.H.C.; Lloyd, C.R.; Lachaud, G. Estimating sparse forest rainfall interception with an analytical model. J. Hydrol. 1995, 170, 79–86. [Google Scholar] [CrossRef]
- Gash, J.H.C.; Valente, F.; David, J.S. Estimates and measurements of evaporation from wet, sparse pine forest in Portugal. Agric. For. Meteorol. 1999, 94, 149–158. [Google Scholar] [CrossRef]
- Palatella, L.; Rana, G.; Vitale, D. Towards a flux-partitioning procedure based on the direct use of high-frequency eddy-covariance data. Bound. Layer Meteorol. 2014, 153, 327–337. [Google Scholar] [CrossRef]
- Scanlon, T.M.; Schmidt, D.F.; Skaggs, T.H. Correlation-based flux partitioning of water vapor and carbon dioxide fluxes: Method simplification and estimation of canopy water use efficiency. Agric. For. Meteorol. 2019, 279, 107732. [Google Scholar] [CrossRef]
- Baker, J.M. Conditional sampling revisited. Agric. For. Meteorol. 2000, 104, 59–65. [Google Scholar] [CrossRef]
- Pattey, E.; Desjardins, R.L.; Rochette, P. Accuracy of the relaxed eddy-accumulation technique, evaluated using CO2 flux measurements. Bound. Layer Meteorol. 1993, 66, 341–355. [Google Scholar] [CrossRef]
- Skaggs, T.H.; Anderson, R.G.; Alfieri, J.G.; Scanlon, T.M.; Kustas, W.P. Fluxpart: Open source software for partitioning carbon dioxide and water vapor fluxes. Agric. For. Meteorol. 2018, 253–254, 218–224. [Google Scholar] [CrossRef]
- Wang, W.; Smith, J.A.; Ramamurthy, P.; Baeck, M.L.; Bou-Zeid, E.; Scanlon, T.M. On the correlation of water vapor and CO2: Application to flux partitioning of evapotranspiration. Water Resour. Res. 2016, 52, 9452–9469. [Google Scholar] [CrossRef]
- Klosterhalfen, A.; Moene, A.; Schmidt, M.; Scanlon, T.; Vereecken, H.; Graf, A. Sensitivity analysis of a source partitioning method for H2O and CO2 fluxes based on high frequency eddy covariance data: Findings from field data and large eddy simulations. Agric. For. Meteorol. 2019, 265, 152–170. [Google Scholar] [CrossRef]

{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Decoupling State | Mixed Indicators | Proportion of Valid Daytime Data (%) | Proportion of Valid Data at Night (%) | Proportion of Total Valid Data (%) | Methods |
|---|---|---|---|---|---|
| - | No index | 80.21 | 52.51 | 67.9 | 1.1 |
| *** | 70.64 | 42.3 | 58.04 | 1.2 | |
| Night | Single-level u* | 80.21 | 24.06 | 55.22 | 2.1.1 |
| *** | 70.64 | 21.01 | 53.42 | 2.1.2 | |
| Single-level σw | 80.21 | 32.21 | 58.86 | 2.2.1 | |
| *** | 70.64 | 26.74 | 56.41 | 2.2.2 | |
| Double-level u* | 80.21 | 8.95 | 48.49 | 2.3.1 | |
| *** | 70.64 | 7.58 | 47.88 | 2.3.2 | |
| Double-level σw | 80.21 | 13.68 | 50.6 | 2.4.1 | |
| *** | 70.64 | 10.95 | 49.38 | 2.4.2 | |
| Daily | Double-level u* | 55.57 | 8.95 | 34.86 | 3.1.1 |
| *** | 50.89 | 7.58 | 32.22 | 3.1.2 | |
| Double-level σw | 71.64 | 13.68 | 45.84 | 3.2.1 | |
| *** | 64.59 | 10.95 | 41.93 | 3.2.2 |
| Month | TR2.2.2 | CEC | MREA | FVS | ||
|---|---|---|---|---|---|---|
| Solution Ratio | TR | 7 | 62.18 | 96.16 | 67.63 | 62.99 |
| 8 | 50.40 | 91.06 | 57.59 | 50.54 | ||
| 9 | 49.45 | 78.62 | 48.90 | 48.36 | ||
| Daily Average | TR | 7 | 6.06 | 3.16 | 5.35 | 4.56 |
| 8 | 6.13 | 4.43 | 5.64 | 4.64 | ||
| 9 | 5.36 | 3.55 | 4.90 | 3.86 | ||
| Ev | 7 | 1.65 | 2.88 | 2.30 | 2.14 | |
| 8 | 1.42 | 1.73 | 0.91 | 1.90 | ||
| 9 | 1.58 | 2.33 | 1.45 | 2.49 | ||
| TR/ET | 7 | 78.62 | 56.20 | 69.90 | 68.07 | |
| 8 | 81.18 | 71.86 | 86.13 | 70.98 | ||
| 9 | 77.26 | 60.33 | 77.11 | 60.83 |
| Parameters | Month | CEC | MREA | FVS | |
|---|---|---|---|---|---|
| TR | 1:1 slope | 7 | 0.64 | 0.92 | 0.83 |
| 8 | 0.81 | 1.01 | 0.82 | ||
| 9 | 0.65 | 0.94 | 0.69 | ||
| R | 7 | 0.90 | 0.95 | 0.92 | |
| 8 | 0.90 | 0.97 | 0.90 | ||
| 9 | 0.86 | 0.96 | 0.93 | ||
| RMSE | 7 | 13.58 | 16.33 | 16.62 | |
| 8 | 24.46 | 17.04 | 29.07 | ||
| 9 | 17.28 | 15.52 | 17.09 | ||
| Ev | 1:1 slope | 7 | 1.99 | 0.95 | 1.23 |
| 8 | 1.80 | 0.65 | 1.26 | ||
| 9 | 1.81 | 0.87 | 1.66 | ||
| R | 7 | 0.80 | 0.57 | 0.78 | |
| 8 | 0.62 | 0.45 | 0.73 | ||
| 9 | 0.78 | 0.41 | 0.73 | ||
| RMSE | 7 | 20.65 | 13.97 | 13.33 | |
| 8 | 23.03 | 7.50 | 10.00 | ||
| 9 | 21.78 | 18.57 | 21.26 |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2024 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Wang, X.; Zhou, Y.; Huang, H.; Gao, X.; Sun, S.; Meng, P.; Zhang, J. Comparing Four Evapotranspiration Partitioning Methods from Eddy Covariance Considering Turbulent Mixing in a Poplar Plantation. Water 2024, 16, 1548. https://doi.org/10.3390/w16111548
Wang X, Zhou Y, Huang H, Gao X, Sun S, Meng P, Zhang J. Comparing Four Evapotranspiration Partitioning Methods from Eddy Covariance Considering Turbulent Mixing in a Poplar Plantation. Water. 2024; 16(11):1548. https://doi.org/10.3390/w16111548
Chicago/Turabian StyleWang, Xin, Yu Zhou, Hui Huang, Xiang Gao, Shoujia Sun, Ping Meng, and Jinsong Zhang. 2024. "Comparing Four Evapotranspiration Partitioning Methods from Eddy Covariance Considering Turbulent Mixing in a Poplar Plantation" Water 16, no. 11: 1548. https://doi.org/10.3390/w16111548
APA StyleWang, X., Zhou, Y., Huang, H., Gao, X., Sun, S., Meng, P., & Zhang, J. (2024). Comparing Four Evapotranspiration Partitioning Methods from Eddy Covariance Considering Turbulent Mixing in a Poplar Plantation. Water, 16(11), 1548. https://doi.org/10.3390/w16111548